
Table of SynGAP1 Isoform α2 (UniProt Q96PV0-1) Missense Variants.
| c.dna | Variant | SGM Consensus | Domain and Structure information: based on WT protein | Annotated databases | Deep learning-based pathogenicity predictions | Folding stability-based pathogenicity predictions | Sequence/structure-based pathogenicity predictions | Phase Separation | Evolutionary/physical properties | Molecular Dynamics-based analysis | DOI | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Domain | IUPred2 | ANCHOR2 | AlphaFold | MobiDB | PhosphoSitePlus | ClinVar | gnomAD | ESM1b | AlphaMissense | FoldX | Rosetta | Foldetta | PremPS | REVEL | PROVEAN | PolyPhen-2 HumDiv | PolyPhen-2 HumVar | FATHMM | SIFT | PSMutPred | PAM | Physical | SASA | Normalized B-factor backbone | Normalized B-factor sidechain | SynGAP Structural Annotation | |||||||||||||||||||||||||||||||||||||||||||||
| Score | Prediction | Score | Prediction | pLDDT | disorder | disorder | LTP | HTP | KL | PTM | Clinical Status | Review | Subm. | ID | Allele count | Allele freq. | LLR score | Prediction | Pathogenicity | Class | Optimized | Average ΔΔG | Prediction | StdDev | ΔΔG | Prediction | ΔΔG | Prediction | ΔΔG | Prediction | Score | Prediction | Score | Prediction | pph2_prob | Prediction | pph2_prob | Prediction | Nervous System Score | Prediction | Prediction | Status | Conservation | Sequences | IP RF | SP RF | Prediction | PAM250 | PAM120 | Hydropathy Δ | MW Δ | Average | Δ | Δ | StdDev | Δ | StdDev | Secondary | Tertiary bonds | Inside out | GAP-Ras interface | At membrane | No effect | MD Alert | Verdict | Description | |||||
| c.3490C>A | L1164M 2D  AIThe SynGAP1 missense variant L1164M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also leans benign. No Foldetta (FoldX‑MD/ Rosetta stability) result is available, so it does not influence the interpretation. Overall, the majority of computational evidence points to a benign impact for the variant, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.517562 | Disordered | 0.853935 | Binding | 0.325 | 0.815 | 0.375 | -5.493 | Likely Benign | 0.757 | Likely Pathogenic | Likely Benign | 0.349 | Likely Benign | -0.29 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 5.34 | Benign | 0.13 | Tolerated | 0.0847 | 0.3378 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3532T>C | Y1178H 2D  AIThe SynGAP1 missense variant Y1178H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence points to a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.568433 | Binding | 0.554 | 0.688 | 0.250 | -2.926 | Likely Benign | 0.775 | Likely Pathogenic | Likely Benign | 0.340 | Likely Benign | -0.78 | Neutral | 0.995 | Probably Damaging | 0.892 | Possibly Damaging | 5.48 | Benign | 0.03 | Affected | 0.2272 | 0.0341 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||||||
| c.3547T>C | Y1183H 2D AIThe SynGAP1 missense variant Y1183H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM, all of which classify the substitution as benign. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. In contrast, AlphaMissense‑Default predicts a pathogenic effect, while AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and no Foldetta stability data are available. Overall, the preponderance of evidence points to a benign impact for Y1183H, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.566480 | Disordered | 0.527818 | Binding | 0.523 | 0.652 | 0.500 | -3.289 | Likely Benign | 0.925 | Likely Pathogenic | Ambiguous | 0.026 | Likely Benign | -0.70 | Neutral | 0.029 | Benign | 0.017 | Benign | 2.75 | Benign | 0.18 | Tolerated | 0.2227 | 0.0243 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||||||
| c.3574C>A | L1192M 2D AIThe SynGAP1 missense variant L1192M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags the variant as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also reports a benign likelihood. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.575842 | Disordered | 0.441757 | Uncertain | 0.762 | 0.609 | 0.625 | -4.591 | Likely Benign | 0.195 | Likely Benign | Likely Benign | 0.092 | Likely Benign | -0.44 | Neutral | 0.606 | Possibly Damaging | 0.287 | Benign | 2.66 | Benign | 0.16 | Tolerated | 0.0669 | 0.2486 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3592T>C | Y1198H 2D AIThe SynGAP1 missense variant Y1198H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and SIFT, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—consistently predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available, so it does not influence the overall assessment. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.626927 | Disordered | 0.439379 | Uncertain | 0.853 | 0.593 | 0.250 | -10.394 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.246 | Likely Benign | -2.86 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.45 | Pathogenic | 0.19 | Tolerated | 0.1975 | 0.0373 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||||||
| c.3613C>A | L1205M 2D AIThe SynGAP1 missense variant L1205M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the variant as damaging. The high‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple in silico predictors points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.552471 | Binding | 0.880 | 0.576 | 0.375 | -9.793 | Likely Pathogenic | 0.945 | Likely Pathogenic | Ambiguous | 0.231 | Likely Benign | -1.73 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.0627 | 0.2297 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3625C>A | L1209M 2D AIThe SynGAP1 missense variant L1209M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL and PROVEAN; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this: AlphaMissense‑Optimized remains uncertain, while Foldetta (a combined FoldX‑MD and Rosetta stability analysis) is not available for this residue. Given the predominance of pathogenic calls and the SGM‑Consensus result, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar entry exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.583711 | Binding | 0.899 | 0.574 | 0.375 | -10.605 | Likely Pathogenic | 0.934 | Likely Pathogenic | Ambiguous | 0.171 | Likely Benign | -1.66 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.0676 | 0.2486 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3646C>A | L1216M 2D AIThe SynGAP1 missense variant L1216M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs 2 benign), and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward a pathogenic interpretation. This conclusion is not contradicted by ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.580690 | Disordered | 0.504713 | Binding | 0.863 | 0.563 | 0.250 | -6.590 | Likely Benign | 0.806 | Likely Pathogenic | Ambiguous | 0.142 | Likely Benign | -1.40 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.16 | Pathogenic | 0.00 | Affected | 0.0612 | 0.2092 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3655T>C | Y1219H 2D AIThe SynGAP1 missense variant Y1219H is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic)—all predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence from both general and high‑accuracy tools indicates that the variant is most likely pathogenic, which does not contradict its current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.613573 | Disordered | 0.474748 | Uncertain | 0.855 | 0.557 | 0.375 | Uncertain | 1 | -9.511 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.363 | Likely Benign | -3.62 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.15 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2234 | 0.0573 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||
| c.3667C>A | L1223M 2D AIThe SynGAP1 missense variant L1223M is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM all predict a pathogenic outcome; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also classifies the variant as benign, and Foldetta results are unavailable. Taken together, the balance of evidence favors a benign effect for L1223M, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.608892 | Disordered | 0.436267 | Uncertain | 0.868 | 0.540 | 0.375 | -5.759 | Likely Benign | 0.358 | Ambiguous | Likely Benign | 0.166 | Likely Benign | -1.29 | Neutral | 0.981 | Probably Damaging | 0.752 | Possibly Damaging | 1.50 | Pathogenic | 0.04 | Affected | 0.0667 | 0.3014 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3670C>A | L1224M 2D AIThe SynGAP1 missense variant L1224M is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign) all indicate a tolerated change. In contrast, polyPhen‑2 (HumDiv and HumVar) and FATHMM predict a pathogenic impact. When focusing on high‑accuracy predictors, AlphaMissense‑Optimized remains benign and the SGM‑Consensus also supports a benign outcome; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this assessment does not conflict with ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.525368 | Disordered | 0.441554 | Uncertain | 0.871 | 0.543 | 0.500 | -5.614 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.085 | Likely Benign | -0.55 | Neutral | 0.994 | Probably Damaging | 0.925 | Probably Damaging | 2.38 | Pathogenic | 0.18 | Tolerated | 0.0660 | 0.2486 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3700C>A | L1234M 2D AIThe SynGAP1 L1234M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence slightly favors a pathogenic interpretation (5 pathogenic vs 4 benign predictions). This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.599170 | Disordered | 0.575096 | Binding | 0.844 | 0.527 | 0.125 | -8.235 | Likely Pathogenic | 0.379 | Ambiguous | Likely Benign | 0.126 | Likely Benign | -0.99 | Neutral | 0.898 | Possibly Damaging | 0.354 | Benign | 1.50 | Pathogenic | 0.03 | Affected | 0.0648 | 0.2944 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3709T>C | Y1237H 2D AIThe SynGAP1 missense variant Y1237H is not reported in ClinVar (ClinVar ID = None) and has no entries in gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic. ESM1b is uncertain, and Foldetta results are unavailable. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. With no conflicting ClinVar evidence, the collective predictions strongly suggest that Y1237H is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.653063 | Disordered | 0.563444 | Binding | 0.842 | 0.535 | 0.125 | -7.985 | In-Between | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.383 | Likely Benign | -4.09 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.45 | Pathogenic | 0.00 | Affected | 0.2221 | 0.0173 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||||||
| c.3721C>A | L1241M 2D AISynGAP1 missense variant L1241M is listed in ClinVar with an Uncertain significance and is not reported in gnomAD. Functional prediction tools show a split verdict: benign calls come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic calls are made by polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is unresolved (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability predictor that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Consequently, the high‑accuracy tools do not converge on a single interpretation. Overall, the predictions are balanced between benign and pathogenic, leaving the variant’s effect uncertain, which aligns with its ClinVar designation of Uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.545602 | Disordered | 0.488880 | Uncertain | 0.828 | 0.541 | 0.375 | Uncertain | 1 | -5.881 | Likely Benign | 0.782 | Likely Pathogenic | Likely Benign | 0.167 | Likely Benign | -1.43 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.65 | Pathogenic | 0.00 | Affected | 0.0781 | 0.2280 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||
| c.3796C>A | L1266M 2D AIThe SynGAP1 missense variant L1266M is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a larger set—polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict pathogenicity. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments are mixed: AlphaMissense‑Optimized yields an uncertain result, SGM‑Consensus remains likely pathogenic, and Foldetta data are unavailable. Overall, the majority of evidence points toward a pathogenic effect for L1266M. This conclusion does not conflict with ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.802655 | Binding | 0.868 | 0.602 | 0.000 | -8.257 | Likely Pathogenic | 0.938 | Likely Pathogenic | Ambiguous | 0.126 | Likely Benign | -1.67 | Neutral | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 2.12 | Pathogenic | 0.00 | Affected | 0.0627 | 0.3113 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3802C>A | L1268M 2D AIThe SynGAP1 missense variant L1268M is reported in gnomAD (ID 6‑33447850‑C‑A) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; a Foldetta stability analysis is not available. Overall, the preponderance of evidence from multiple in silico predictors and consensus methods indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.458154 | Structured | 0.804315 | Binding | 0.859 | 0.629 | 0.000 | 6-33447850-C-A | -4.689 | Likely Benign | 0.140 | Likely Benign | Likely Benign | 0.062 | Likely Benign | -0.07 | Neutral | 0.990 | Probably Damaging | 0.796 | Possibly Damaging | 2.67 | Benign | 0.08 | Tolerated | 3.77 | 5 | 0.0627 | 0.2297 | 2 | 4 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||
| c.3817C>A | L1273M 2D AIThe SynGAP1 missense variant L1273M is catalogued in gnomAD (ID 6‑33447865‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect for the variant. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.773625 | Binding | 0.747 | 0.677 | 0.500 | 6-33447865-C-A | -5.375 | Likely Benign | 0.503 | Ambiguous | Likely Benign | 0.161 | Likely Benign | -1.68 | Neutral | 0.983 | Probably Damaging | 0.874 | Possibly Damaging | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0824 | 0.3932 | 2 | 4 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||
| c.3848C>A | P1283Q 2D  AIThe SynGAP1 missense variant P1283Q is reported in gnomAD (variant ID 6‑33447896‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta’s protein‑folding stability analysis is unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.819686 | Binding | 0.484 | 0.732 | 0.875 | 6-33447896-C-A | -4.028 | Likely Benign | 0.085 | Likely Benign | Likely Benign | 0.102 | Likely Benign | -1.11 | Neutral | 0.991 | Probably Damaging | 0.567 | Possibly Damaging | 2.72 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1195 | 0.2926 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3850C>A | L1284M 2D AIThe SynGAP1 missense variant L1284M is reported in gnomAD (ID 6‑33447898‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from the four benign‑oriented tools). Pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar classification (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.824557 | Binding | 0.441 | 0.748 | 0.875 | 6-33447898-C-A | -5.332 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.49 | Neutral | 0.970 | Probably Damaging | 0.637 | Possibly Damaging | 2.61 | Benign | 0.03 | Affected | 3.77 | 5 | 0.0731 | 0.2092 | 2 | 4 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||
| c.3854C>A | P1285Q 2D AIThe SynGAP1 missense variant P1285Q is reported in gnomAD (variant ID 6‑33447902‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 (HumDiv and HumVar) predict it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign. No Foldetta stability analysis is available, so it does not influence the overall assessment. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.821643 | Binding | 0.557 | 0.759 | 0.750 | 6-33447902-C-A | -4.073 | Likely Benign | 0.100 | Likely Benign | Likely Benign | 0.072 | Likely Benign | -1.67 | Neutral | 0.977 | Probably Damaging | 0.632 | Possibly Damaging | 4.22 | Benign | 0.13 | Tolerated | 4.32 | 2 | 0.1476 | 0.3025 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3871C>A | L1291M 2D AIThe SynGAP1 missense variant L1291M is reported in gnomAD (ID 6‑33447919‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT each predict a pathogenic impact. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (derived from the four benign predictors) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is assigned). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.863458 | Binding | 0.579 | 0.797 | 0.625 | 6-33447919-C-A | -5.625 | Likely Benign | 0.159 | Likely Benign | Likely Benign | 0.045 | Likely Benign | -0.93 | Neutral | 0.970 | Probably Damaging | 0.728 | Possibly Damaging | 2.53 | Benign | 0.03 | Affected | 3.77 | 5 | 0.0767 | 0.2973 | 2 | 4 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||
| c.3904T>A | L1302M 2D AIThe SynGAP1 missense variant L1302M is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of predictions (six benign vs. four pathogenic) support a benign classification. There is no ClinVar entry to contradict this assessment, so the variant is most likely benign based on the available computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | -5.246 | Likely Benign | 0.131 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -1.34 | Neutral | 0.960 | Probably Damaging | 0.799 | Possibly Damaging | 1.54 | Pathogenic | 0.00 | Affected | 0.0929 | 0.3439 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3911C>A | P1304Q 2D AIThe SynGAP1 missense variant P1304Q is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy predictions are consistent: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.885302 | Disordered | 0.886417 | Binding | 0.475 | 0.866 | 0.875 | -4.890 | Likely Benign | 0.090 | Likely Benign | Likely Benign | 0.119 | Likely Benign | -0.84 | Neutral | 0.586 | Possibly Damaging | 0.287 | Benign | 2.84 | Benign | 0.04 | Affected | 0.1512 | 0.4149 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||||
| c.3920C>A | P1307Q 2D AIThe SynGAP1 missense variant P1307Q is listed in ClinVar (ID 982827.0) with an “Uncertain” clinical significance and is present in gnomAD (variant ID 6‑33451794‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.798249 | Disordered | 0.913511 | Binding | 0.491 | 0.901 | 0.875 | Uncertain | 1 | 6-33451794-C-A | -4.227 | Likely Benign | 0.114 | Likely Benign | Likely Benign | 0.192 | Likely Benign | -0.88 | Neutral | 0.988 | Probably Damaging | 0.765 | Possibly Damaging | 2.82 | Benign | 0.03 | Affected | 3.77 | 5 | 0.1629 | 0.4464 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||
| c.3931C>A | L1311M 2D AIThe SynGAP1 missense variant L1311M is reported in gnomAD (ID 6‑33451805‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates “Likely Benign.” In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact. High‑accuracy assessments reinforce the benign view: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence supports a benign classification for L1311M, and this conclusion is not contradicted by any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.968153 | Binding | 0.393 | 0.907 | 0.750 | 6-33451805-C-A | 3 | 1.86e-6 | -4.817 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.060 | Likely Benign | 0.18 | Neutral | 0.936 | Possibly Damaging | 0.632 | Possibly Damaging | 2.73 | Benign | 0.23 | Tolerated | 3.77 | 5 | 0.1068 | 0.4279 | 2 | 4 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||
| c.3938C>A | P1313Q 2D AIThe SynGAP1 missense variant P1313Q is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the consensus of all available predictions strongly supports a benign classification, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.970301 | Binding | 0.452 | 0.902 | 0.750 | -4.533 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.063 | Likely Benign | 0.99 | Neutral | 0.324 | Benign | 0.287 | Benign | 4.31 | Benign | 0.15 | Tolerated | 0.1748 | 0.5651 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||||
| c.3941C>A | P1314Q 2D AIThe SynGAP1 missense variant P1314Q is listed in gnomAD (ID 6‑33451815‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign. No Foldetta stability data are available, so folding‑stability evidence is unavailable. Overall, the preponderance of computational evidence indicates that P1314Q is most likely benign, and this conclusion is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.971592 | Binding | 0.467 | 0.903 | 0.750 | 6-33451815-C-A | -4.222 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.039 | Likely Benign | -0.55 | Neutral | 0.618 | Possibly Damaging | 0.333 | Benign | 4.24 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1612 | 0.4588 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3952C>A | L1318M 2D AIThe SynGAP1 missense variant L1318M is listed in gnomAD (ID 6‑33451826‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar classification (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.968271 | Binding | 0.399 | 0.865 | 0.750 | 6-33451826-C-A | -4.625 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.039 | Likely Benign | -0.62 | Neutral | 0.939 | Possibly Damaging | 0.396 | Benign | 4.01 | Benign | 0.03 | Affected | 3.77 | 5 | 0.1083 | 0.4027 | 2 | 4 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||
| c.3962C>A | P1321Q 2D AIThe SynGAP1 missense variant P1321Q is listed in ClinVar (ID 833687.0) as benign and is present in gnomAD (variant ID 6‑33451836‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification, and AlphaMissense‑Optimized also reports benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence—including the high‑confidence SGM consensus and AlphaMissense‑Optimized—supports a benign impact. This conclusion aligns with the ClinVar benign status, with no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.933505 | Binding | 0.463 | 0.828 | 0.875 | Benign | 1 | 6-33451836-C-A | 1 | 6.58e-7 | -5.594 | Likely Benign | 0.079 | Likely Benign | Likely Benign | 0.055 | Likely Benign | -0.74 | Neutral | 0.659 | Possibly Damaging | 0.034 | Benign | 4.24 | Benign | 0.09 | Tolerated | 3.77 | 5 | 0.1839 | 0.4884 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||
| c.3968C>A | P1323Q 2D AIThe SynGAP1 missense variant P1323Q is reported in gnomAD (ID 6‑33451842‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that P1323Q is most likely benign, and this conclusion is not contradicted by any ClinVar classification (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451842-C-A | -6.134 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.033 | Likely Benign | -0.85 | Neutral | 0.712 | Possibly Damaging | 0.328 | Benign | 3.82 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1791 | 0.4657 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3971C>A | P1324Q 2D AIThe SynGAP1 missense variant P1324Q is reported in gnomAD (ID 6‑33451845‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools, including the high‑accuracy predictors, points to a benign effect. This prediction is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | 6-33451845-C-A | -5.577 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.028 | Likely Benign | -0.69 | Neutral | 0.027 | Benign | 0.010 | Benign | 4.27 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1744 | 0.4736 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3977C>A | P1326Q 2D AIThe SynGAP1 missense variant P1326Q is listed in ClinVar (ID 2806103.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33451851‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar “Uncertain” classification, which remains inconclusive. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | Uncertain | 1 | 6-33451851-C-A | 1 | 6.40e-7 | -5.422 | Likely Benign | 0.128 | Likely Benign | Likely Benign | 0.138 | Likely Benign | -0.86 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.62 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1769 | 0.4457 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||
| c.397C>A | L133M 2D AIThe SynGAP1 missense variant L133M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default; ESM1b and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta results are unavailable. Overall, the majority of standard predictors (four pathogenic vs. three benign) indicate a pathogenic impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.422041 | Structured | 0.718429 | Binding | 0.320 | 0.896 | 0.250 | -7.993 | In-Between | 0.841 | Likely Pathogenic | Ambiguous | 0.095 | Likely Benign | -0.77 | Neutral | 0.877 | Possibly Damaging | 0.580 | Possibly Damaging | 3.57 | Benign | 0.03 | Affected | 0.0856 | 0.2229 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3985C>A | L1329M 2D AIThe SynGAP1 missense change L1329M is recorded in gnomAD (ID 6‑33451859‑C‑A) but has no ClinVar entry. Functional prediction tools split into two consensus groups: benign‑predicted by REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic‑predicted by polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized scores benign, and the SGM‑Consensus likewise indicates benign. Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.924905 | Binding | 0.336 | 0.748 | 0.875 | 6-33451859-C-A | -5.493 | Likely Benign | 0.780 | Likely Pathogenic | Likely Benign | 0.087 | Likely Benign | -1.16 | Neutral | 0.994 | Probably Damaging | 0.990 | Probably Damaging | 3.10 | Benign | 0.00 | Affected | 3.77 | 5 | 0.0914 | 0.4273 | 2 | 4 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||
| c.478C>A | L160M 2D AIThe SynGAP1 missense variant L160M is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; the SGM Consensus is unavailable; and Foldetta, which combines FoldX‑MD and Rosetta stability calculations, has no reported output for this variant. Overall, the balance of evidence leans toward a pathogenic interpretation, but the single high‑accuracy benign prediction and the lack of a ClinVar assertion mean the variant’s clinical significance remains uncertain. This assessment does not contradict any existing ClinVar status, as none is available. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.526760 | Binding | 0.275 | 0.728 | 0.125 | -11.120 | Likely Pathogenic | 0.723 | Likely Pathogenic | Likely Benign | 0.097 | Likely Benign | -0.94 | Neutral | 0.877 | Possibly Damaging | 0.580 | Possibly Damaging | 3.85 | Benign | 0.00 | Affected | 0.0837 | 0.3613 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.517C>A | L173M 2D AIThe SynGAP1 missense variant L173M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Two tools—AlphaMissense‑Default and ESM1b—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields two benign and two uncertain calls, and Foldetta data are unavailable. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.534167 | Disordered | 0.491566 | Uncertain | 0.390 | 0.631 | 0.375 | -7.149 | In-Between | 0.534 | Ambiguous | Likely Benign | 0.129 | Likely Benign | -0.61 | Neutral | 0.940 | Possibly Damaging | 0.564 | Possibly Damaging | 3.96 | Benign | 0.20 | Tolerated | 0.0648 | 0.3123 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.52T>C | Y18H 2D AIThe SynGAP1 missense variant Y18H is listed in gnomAD (ID 6‑33420316‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.446314 | Uncertain | 0.345 | 0.908 | 0.375 | 6-33420316-T-C | -2.871 | Likely Benign | 0.542 | Ambiguous | Likely Benign | 0.045 | Likely Benign | -0.58 | Neutral | 0.872 | Possibly Damaging | 0.114 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2805 | 0.0993 | 2 | 0 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||||
| c.556T>A | L186M 2D AIThe SynGAP1 missense variant L186M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool rates the variant as uncertain, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (5 pathogenic vs. 3 benign) suggest a pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.458154 | Structured | 0.428613 | Uncertain | 0.397 | 0.617 | 0.500 | -11.783 | Likely Pathogenic | 0.933 | Likely Pathogenic | Ambiguous | 0.146 | Likely Benign | -1.58 | Neutral | 0.952 | Possibly Damaging | 0.694 | Possibly Damaging | 3.50 | Benign | 0.00 | Affected | 0.0671 | 0.3396 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.559C>A | L187M 2D AIThe SynGAP1 missense variant L187M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, which would assess protein‑folding stability, has no available output for this variant. Overall, the majority of predictions (five pathogenic vs. three benign) lean toward pathogenicity, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.465241 | Structured | 0.428046 | Uncertain | 0.367 | 0.625 | 0.375 | -8.814 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.136 | Likely Benign | -1.35 | Neutral | 0.971 | Probably Damaging | 0.641 | Possibly Damaging | 3.72 | Benign | 0.02 | Affected | 0.0848 | 0.3613 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.586T>A | L196M 2D AIThe SynGAP1 missense variant L196M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool rates the variant as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result. Overall, more tools predict pathogenicity (5) than benignity (3), and the high‑accuracy predictions do not overturn this trend. Therefore, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.284882 | Structured | 0.429452 | Uncertain | 0.489 | 0.521 | 0.125 | -8.957 | Likely Pathogenic | 0.902 | Likely Pathogenic | Ambiguous | 0.094 | Likely Benign | -1.48 | Neutral | 0.971 | Probably Damaging | 0.691 | Possibly Damaging | 3.64 | Benign | 0.01 | Affected | 0.0689 | 0.2786 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.598T>A | L200M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L200M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign outcome. Only two tools, polyPhen‑2 HumDiv and HumVar, predict pathogenicity. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign; the SGM‑Consensus, derived from the four high‑confidence predictors, is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports a benign effect. Taken together, the overwhelming majority of evidence supports a benign classification for L200M, and this conclusion is consistent with the absence of a ClinVar entry. Thus, the variant is most likely benign, and this assessment does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.366687 | Structured | 0.428168 | Uncertain | 0.687 | 0.453 | 0.125 | -4.107 | Likely Benign | 0.215 | Likely Benign | Likely Benign | 0.08 | Likely Benign | 0.1 | 0.41 | Likely Benign | 0.25 | Likely Benign | -0.25 | Likely Benign | 0.139 | Likely Benign | 0.47 | Neutral | 0.997 | Probably Damaging | 0.960 | Probably Damaging | 4.11 | Benign | 0.31 | Tolerated | 0.0751 | 0.3511 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||
| c.623C>A | P208Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P208Q missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and FATHMM. The majority of tools predict a pathogenic impact: SGM‑Consensus (Likely Pathogenic), FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and Foldetta. Two tools give uncertain results (AlphaMissense‑Optimized and Rosetta) and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as Pathogenic, reinforcing a deleterious prediction. Overall, the evidence strongly favors a pathogenic classification, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.271506 | Structured | 0.399506 | Uncertain | 0.864 | 0.345 | 0.125 | -9.746 | Likely Pathogenic | 0.906 | Likely Pathogenic | Ambiguous | 3.30 | Destabilizing | 0.7 | 0.75 | Ambiguous | 2.03 | Destabilizing | 1.23 | Destabilizing | 0.411 | Likely Benign | -6.78 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.78 | Benign | 0.00 | Affected | 0.1570 | 0.4663 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||
| c.640C>A | L214M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L214M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are PROVEAN and FATHMM, whereas the majority of tools (REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact; the remaining tools (FoldX, Rosetta, Foldetta, premPS) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs two benign) and Foldetta is uncertain. Thus, the available evidence points to a pathogenic effect. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.155435 | Structured | 0.372604 | Uncertain | 0.818 | 0.302 | 0.125 | -9.347 | Likely Pathogenic | 0.958 | Likely Pathogenic | Likely Pathogenic | 0.74 | Ambiguous | 0.3 | 1.43 | Ambiguous | 1.09 | Ambiguous | 0.80 | Ambiguous | 0.646 | Likely Pathogenic | -1.72 | Neutral | 0.997 | Probably Damaging | 0.916 | Probably Damaging | 5.73 | Benign | 0.01 | Affected | 0.0805 | 0.4054 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.736C>A | L246M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L246M has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include PROVEAN and FATHMM, while a majority (REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) predict a pathogenic impact. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized) are uncertain. High‑accuracy methods are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Thus no high‑accuracy tool provides a definitive verdict. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any ClinVar status because no ClinVar record exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.472492 | Structured | 0.302312 | Uncertain | 0.859 | 0.364 | 0.000 | -11.386 | Likely Pathogenic | 0.922 | Likely Pathogenic | Ambiguous | 0.65 | Ambiguous | 0.2 | 0.76 | Ambiguous | 0.71 | Ambiguous | 0.87 | Ambiguous | 0.661 | Likely Pathogenic | -1.79 | Neutral | 0.997 | Probably Damaging | 0.916 | Probably Damaging | 4.72 | Benign | 0.01 | Affected | 0.0710 | 0.3511 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.796C>A | L266M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L266M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Two tools remain uncertain: premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the majority of tools (six benign vs. four pathogenic, with two uncertain) lean toward a benign classification, and this conclusion is not contradicted by ClinVar status. Thus, the variant is most likely benign based on the collective predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.232838 | Structured | 0.297157 | Uncertain | 0.948 | 0.264 | 0.000 | -9.740 | Likely Pathogenic | 0.362 | Ambiguous | Likely Benign | 0.07 | Likely Benign | 0.1 | -0.47 | Likely Benign | -0.20 | Likely Benign | 0.95 | Ambiguous | 0.288 | Likely Benign | -1.66 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.55 | Pathogenic | 0.07 | Tolerated | 0.0581 | 0.2853 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.820C>A | L274M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L274M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, PROVEAN, and AlphaMissense‑Optimized, whereas those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. Three tools (Rosetta, premPS, and ESM1b) returned uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicting benign. Overall, the majority of predictions (seven pathogenic vs. four benign) and the SGM Consensus support a pathogenic classification. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.066181 | Structured | 0.377483 | Uncertain | 0.866 | 0.195 | 0.250 | -7.386 | In-Between | 0.658 | Likely Pathogenic | Likely Benign | 0.24 | Likely Benign | 0.2 | 0.74 | Ambiguous | 0.49 | Likely Benign | 0.81 | Ambiguous | 0.512 | Likely Pathogenic | -1.77 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 0.04 | Pathogenic | 0.01 | Affected | 0.0810 | 0.2202 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.838T>C | Y280H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y280H is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (gnomAD ID: none). Among the available in‑silico predictors, none indicate a benign effect; all 14 tools that produced a classification predict pathogenicity: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No predictions are inconclusive or missing. Based on the unanimous pathogenic predictions and the absence of any benign calls, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status (which is currently unreported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.161087 | Structured | 0.321243 | Uncertain | 0.917 | 0.248 | 0.125 | -9.970 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 3.01 | Destabilizing | 0.3 | 2.58 | Destabilizing | 2.80 | Destabilizing | 1.95 | Destabilizing | 0.588 | Likely Pathogenic | -4.60 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.94 | Pathogenic | 0.01 | Affected | 0.2021 | 0.0212 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.841T>C | Y281H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y281H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: SIFT classifies it as benign, whereas the remaining 13 tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default) predict pathogenicity. High‑accuracy methods provide a clearer picture: AlphaMissense‑Optimized is uncertain, SGM‑Consensus indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic folding instability. With the overwhelming consensus from most predictors and the high‑accuracy tools supporting a damaging effect, the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.098513 | Structured | 0.337647 | Uncertain | 0.927 | 0.254 | 0.000 | -8.522 | Likely Pathogenic | 0.884 | Likely Pathogenic | Ambiguous | 2.65 | Destabilizing | 0.2 | 2.05 | Destabilizing | 2.35 | Destabilizing | 1.63 | Destabilizing | 0.717 | Likely Pathogenic | -3.82 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 0.99 | Pathogenic | 0.16 | Tolerated | 0.2651 | 0.1503 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.856C>A | L286M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L286M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, PROVEAN, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. Two tools (premPS and ESM1b) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of predictions (seven pathogenic vs. five benign) and the SGM Consensus support a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.122885 | Structured | 0.385647 | Uncertain | 0.932 | 0.260 | 0.000 | -7.998 | In-Between | 0.781 | Likely Pathogenic | Likely Benign | -0.06 | Likely Benign | 0.1 | 0.17 | Likely Benign | 0.06 | Likely Benign | 0.98 | Ambiguous | 0.663 | Likely Pathogenic | -1.84 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.54 | Pathogenic | 0.02 | Affected | 0.0789 | 0.3371 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.868C>A | L290M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L290M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split assessment: benign predictions come from REVEL, FoldX, and PROVEAN, whereas pathogenic predictions are reported by polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Tools with uncertain outcomes include Foldetta, premPS, AlphaMissense‑Optimized, and Rosetta. High‑accuracy analyses indicate that AlphaMissense‑Optimized is inconclusive, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains uncertain. Overall, the majority of evidence leans toward a pathogenic effect, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.127496 | Structured | 0.399723 | Uncertain | 0.904 | 0.255 | 0.000 | -8.393 | Likely Pathogenic | 0.901 | Likely Pathogenic | Ambiguous | 0.48 | Likely Benign | 0.2 | 0.98 | Ambiguous | 0.73 | Ambiguous | 0.59 | Ambiguous | 0.438 | Likely Benign | -1.84 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.92 | Pathogenic | 0.02 | Affected | 0.0823 | 0.4167 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||
| c.871T>C | Y291H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y291H is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: the single benign prediction comes from REVEL, while all other evaluated algorithms (FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict pathogenicity, and the SGM‑Consensus score is “Likely Pathogenic.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized indicates pathogenicity; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a likely pathogenic verdict; and Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a destabilizing, pathogenic effect. No prediction or stability result is missing or inconclusive. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.173081 | Structured | 0.383842 | Uncertain | 0.912 | 0.251 | 0.000 | -8.749 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 2.13 | Destabilizing | 0.2 | 2.61 | Destabilizing | 2.37 | Destabilizing | 1.56 | Destabilizing | 0.355 | Likely Benign | -4.04 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.77 | Pathogenic | 0.02 | Affected | 0.2403 | 0.0499 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.949C>A | L317M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L317M missense variant is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools show a split: benign calls come from REVEL, FoldX, Foldetta, and PROVEAN, whereas pathogenic calls arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments give a mixed picture: AlphaMissense‑Optimized is inconclusive, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic, and Foldetta predicts a benign effect on protein stability. No evidence from ClinVar contradicts these findings. Overall, the preponderance of pathogenic predictions and the SGM‑Consensus result suggest that the variant is most likely pathogenic, though the benign signal from Foldetta and other tools indicates uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.106997 | Structured | 0.394031 | Uncertain | 0.874 | 0.240 | 0.125 | -9.558 | Likely Pathogenic | 0.788 | Likely Pathogenic | Ambiguous | 0.14 | Likely Benign | 0.1 | 0.84 | Ambiguous | 0.49 | Likely Benign | 0.94 | Ambiguous | 0.250 | Likely Benign | -1.84 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.71 | Pathogenic | 0.04 | Affected | 0.0937 | 0.3122 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||
| c.953C>A | P318Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P318Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely converge on a deleterious effect: SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate pathogenicity. Only FoldX, Rosetta, and Foldetta provide uncertain results and are therefore not considered evidence for benign impact. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta remains uncertain. Consequently, the variant is most likely pathogenic based on the consensus of predictive algorithms, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.111485 | Structured | 0.400936 | Uncertain | 0.858 | 0.234 | 0.000 | -11.403 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 1.64 | Ambiguous | 0.2 | 1.29 | Ambiguous | 1.47 | Ambiguous | 1.18 | Destabilizing | 0.638 | Likely Pathogenic | -7.05 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.83 | Pathogenic | 0.01 | Affected | 0.1467 | 0.4731 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||
| c.967C>A | L323M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L323M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The remaining tools—Foldetta, premPS, AlphaMissense‑Default, ESM1b, and Rosetta—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is also inconclusive. Taken together, the evidence leans toward a benign impact for the variant, and this assessment does not contradict any ClinVar annotation, as none exists for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.268042 | Structured | 0.428564 | Uncertain | 0.956 | 0.369 | 0.000 | -7.065 | In-Between | 0.427 | Ambiguous | Likely Benign | 0.11 | Likely Benign | 0.1 | 1.82 | Ambiguous | 0.97 | Ambiguous | 0.93 | Ambiguous | 0.270 | Likely Benign | -1.02 | Neutral | 0.997 | Probably Damaging | 0.939 | Probably Damaging | 0.64 | Pathogenic | 0.03 | Affected | 0.0637 | 0.3010 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.973C>A | L325M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L325M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and premPS. Two tools give uncertain results: Foldetta and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. No prediction or folding stability result is missing or inconclusive. Overall, the majority of tools (six pathogenic vs five benign) lean toward pathogenicity, but the high‑accuracy methods and several benign predictions introduce uncertainty. Thus, the variant is most likely pathogenic based on the collective evidence, and this assessment is not contradicted by any ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.352862 | Structured | 0.424577 | Uncertain | 0.955 | 0.436 | 0.000 | -6.725 | Likely Benign | 0.445 | Ambiguous | Likely Benign | 0.22 | Likely Benign | 0.0 | 2.04 | Destabilizing | 1.13 | Ambiguous | 1.05 | Destabilizing | 0.306 | Likely Benign | -0.86 | Neutral | 0.997 | Probably Damaging | 0.939 | Probably Damaging | 1.36 | Pathogenic | 0.01 | Affected | 0.0934 | 0.3942 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.979C>A | L327M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L327M missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, AlphaMissense‑Optimized, and Foldetta, whereas those that predict a pathogenic outcome are SGM‑Consensus, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) labeling it likely pathogenic, and Foldetta predicting a benign effect. Overall, the majority of tools indicate a pathogenic effect, and this assessment does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.335645 | Structured | 0.409189 | Uncertain | 0.939 | 0.490 | 0.000 | -10.320 | Likely Pathogenic | 0.727 | Likely Pathogenic | Likely Benign | 0.20 | Likely Benign | 0.0 | 0.72 | Ambiguous | 0.46 | Likely Benign | 1.07 | Destabilizing | 0.320 | Likely Benign | -1.61 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.53 | Pathogenic | 0.01 | Affected | 0.0806 | 0.3807 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||
| c.982T>C | Y328H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y328H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect. Benign predictions: none. Pathogenic predictions: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. Based on the consensus of all available predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.411940 | Structured | 0.392519 | Uncertain | 0.916 | 0.497 | 0.000 | -10.885 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 2.08 | Destabilizing | 0.0 | 2.43 | Destabilizing | 2.26 | Destabilizing | 1.35 | Destabilizing | 0.516 | Likely Pathogenic | -4.53 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.54 | Pathogenic | 0.02 | Affected | 0.2572 | 0.0703 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.1231A>T | I411F 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I411F is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are limited to FATHMM, while the remaining tools—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact; premPS is uncertain. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No prediction is missing or inconclusive. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.116183 | Structured | 0.339366 | Uncertain | 0.927 | 0.198 | 0.000 | -13.377 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 5.71 | Destabilizing | 0.1 | 4.53 | Destabilizing | 5.12 | Destabilizing | 0.73 | Ambiguous | 0.503 | Likely Pathogenic | -3.69 | Deleterious | 0.998 | Probably Damaging | 0.973 | Probably Damaging | 3.30 | Benign | 0.00 | Affected | 0.0538 | 0.2853 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1360A>T | I454F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I454F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and FATHMM. All other evaluated algorithms—FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. premPS is uncertain and does not influence the overall assessment. Based on the preponderance of pathogenic predictions and the absence of benign evidence, the variant is most likely pathogenic, with no ClinVar status to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.254060 | Structured | 0.312811 | Uncertain | 0.965 | 0.182 | 0.000 | -12.468 | Likely Pathogenic | 0.960 | Likely Pathogenic | Likely Pathogenic | 4.15 | Destabilizing | 0.7 | 2.96 | Destabilizing | 3.56 | Destabilizing | 0.70 | Ambiguous | 0.464 | Likely Benign | -3.95 | Deleterious | 0.998 | Probably Damaging | 0.973 | Probably Damaging | 3.33 | Benign | 0.00 | Affected | 0.0422 | 0.2560 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.148A>T | I50F 2D AIThe SynGAP1 missense variant I50F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for I50F, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.295083 | Structured | 0.449965 | Uncertain | 0.545 | 0.708 | 0.000 | -5.631 | Likely Benign | 0.681 | Likely Pathogenic | Likely Benign | 0.100 | Likely Benign | -1.27 | Neutral | 0.334 | Benign | 0.074 | Benign | 3.75 | Benign | 0.00 | Affected | 0.0483 | 0.2560 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.1501A>T | I501F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I501F missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that indicate a benign effect include REVEL, Rosetta, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise SGM‑Consensus, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely pathogenic (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta as uncertain (a stability method that integrates FoldX‑MD and Rosetta outputs). No predictions are missing or inconclusive. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.079919 | Structured | 0.366596 | Uncertain | 0.886 | 0.153 | 0.000 | -12.113 | Likely Pathogenic | 0.664 | Likely Pathogenic | Likely Benign | 3.06 | Destabilizing | 0.1 | 0.26 | Likely Benign | 1.66 | Ambiguous | 0.71 | Ambiguous | 0.385 | Likely Benign | -3.88 | Deleterious | 0.998 | Probably Damaging | 0.993 | Probably Damaging | 3.40 | Benign | 0.03 | Affected | 0.0574 | 0.1594 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.151A>T | I51F 2D AIThe SynGAP1 missense variant I51F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus indicates Likely Benign, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for I51F, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.291804 | Structured | 0.454181 | Uncertain | 0.606 | 0.710 | 0.000 | -5.687 | Likely Benign | 0.526 | Ambiguous | Likely Benign | 0.114 | Likely Benign | -0.87 | Neutral | 0.099 | Benign | 0.039 | Benign | 4.13 | Benign | 0.00 | Affected | 0.0565 | 0.3070 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.1528A>T | I510F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I510F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include premPS and AlphaMissense‑Optimized, whereas the majority of tools (SGM‑Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and Foldetta) predict a pathogenic impact; Rosetta remains uncertain. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenicity, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenicity. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.250630 | Uncertain | 0.945 | 0.273 | 0.000 | -8.185 | Likely Pathogenic | 0.713 | Likely Pathogenic | Likely Benign | 4.66 | Destabilizing | 0.7 | 1.38 | Ambiguous | 3.02 | Destabilizing | 0.50 | Likely Benign | 0.692 | Likely Pathogenic | -2.64 | Deleterious | 0.991 | Probably Damaging | 0.854 | Possibly Damaging | -1.14 | Pathogenic | 0.01 | Affected | 0.0552 | 0.1794 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1540A>T | I514F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I514F is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that classify the variant as benign include only FATHMM. All other evaluated algorithms—REVEL, FoldX, Rosetta, Foldetta, premPS (uncertain), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—report it as pathogenic or likely pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, classifies the variant as pathogenic. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, which is consistent with its ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.049374 | Structured | 0.221408 | Uncertain | 0.948 | 0.266 | 0.000 | Uncertain | 1 | -13.383 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 2.35 | Destabilizing | 0.3 | 3.74 | Destabilizing | 3.05 | Destabilizing | 0.93 | Ambiguous | 0.601 | Likely Pathogenic | -3.98 | Deleterious | 0.997 | Probably Damaging | 0.993 | Probably Damaging | 2.89 | Benign | 0.00 | Affected | 3.37 | 35 | 0.0574 | 0.1629 | 0 | 1 | -1.7 | 34.02 | |||||||||||||||||||||||||
| c.1585A>T | I529F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I529F is not reported in ClinVar and is present in gnomAD (variant ID 6‑33438828‑A‑T). Consensus from most in‑silico predictors classifies the change as benign: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic effect. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus is Likely Benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports a benign effect. Overall, the preponderance of evidence indicates that I529F is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.318242 | Structured | 0.019545 | Uncertain | 0.901 | 0.403 | 0.000 | 6-33438828-A-T | 4 | 2.48e-6 | -5.669 | Likely Benign | 0.235 | Likely Benign | Likely Benign | -0.21 | Likely Benign | 0.0 | 0.06 | Likely Benign | -0.08 | Likely Benign | 0.06 | Likely Benign | 0.320 | Likely Benign | -0.97 | Neutral | 0.266 | Benign | 0.054 | Benign | -1.30 | Pathogenic | 0.23 | Tolerated | 3.37 | 35 | 0.0522 | 0.2713 | 0 | 1 | -1.7 | 34.02 | ||||||||||||||||||||||||
| c.1750A>T | I584F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I584F missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only Rosetta, whereas the remaining pathogenic‑predicating tools—REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently classify the variant as deleterious. High‑accuracy assessments further support a pathogenic interpretation: the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Pathogenic,” while AlphaMissense‑Optimized and Foldetta yield uncertain results and are therefore not used as evidence. No other folding‑stability methods provide definitive support. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.059222 | Structured | 0.046673 | Uncertain | 0.846 | 0.244 | 0.000 | -13.582 | Likely Pathogenic | 0.833 | Likely Pathogenic | Ambiguous | 3.20 | Destabilizing | 0.2 | 0.28 | Likely Benign | 1.74 | Ambiguous | 0.66 | Ambiguous | 0.618 | Likely Pathogenic | -3.47 | Deleterious | 0.980 | Probably Damaging | 0.808 | Possibly Damaging | -1.26 | Pathogenic | 0.04 | Affected | 0.0641 | 0.2150 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1765A>T | I589F 2D AIThe SynGAP1 missense variant I589F is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that assess pathogenicity all agree that the variant is deleterious: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify it as pathogenic. No tool predicts a benign effect. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, indicates a pathogenic impact. All available predictions are concordant and supportive. Therefore, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.018415 | Structured | 0.084536 | Uncertain | 0.927 | 0.214 | 0.000 | -15.300 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 7.38 | Destabilizing | 3.4 | 2.05 | Destabilizing | 4.72 | Destabilizing | 1.04 | Destabilizing | 0.905 | Likely Pathogenic | -3.98 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | -1.98 | Pathogenic | 0.00 | Affected | 0.0888 | 0.2574 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1804A>T | I602F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I602F missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a pathogenic effect include SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. No tool predicts a benign outcome. Predictions that are uncertain or inconclusive are Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta as Uncertain. Taken together, the overwhelming majority of evidence points to a pathogenic effect. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.010221 | Structured | 0.186541 | Uncertain | 0.963 | 0.171 | 0.000 | -13.974 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 2.47 | Destabilizing | 1.1 | -0.61 | Ambiguous | 0.93 | Ambiguous | 0.87 | Ambiguous | 0.822 | Likely Pathogenic | -3.98 | Deleterious | 0.999 | Probably Damaging | 0.937 | Probably Damaging | -1.82 | Pathogenic | 0.00 | Affected | 0.0708 | 0.2343 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1876A>T | I626F 2D 3DClick to see structure in 3D Viewer AISynGAP1 I626F is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include only FATHMM, whereas the remaining tools—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS (uncertain), PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—predict it to be pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM‑Consensus (derived from a majority of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. Taken together, the overwhelming majority of evidence indicates that I626F is likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.109221 | Structured | 0.040732 | Uncertain | 0.970 | 0.223 | 0.000 | -14.483 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 4.37 | Destabilizing | 0.3 | 2.12 | Destabilizing | 3.25 | Destabilizing | 0.66 | Ambiguous | 0.631 | Likely Pathogenic | -3.78 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 3.07 | Benign | 0.00 | Affected | 0.0481 | 0.1964 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1888A>T | I630F 2D 3DClick to see structure in 3D Viewer AISynGAP1 I630F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only AlphaMissense‑Optimized. Those that agree on a pathogenic effect comprise SGM‑Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain results are reported by Rosetta, Foldetta, and premPS and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting likely pathogenic, and Foldetta yielding an uncertain stability change. Overall, the preponderance of evidence points to a pathogenic impact for I630F. This conclusion is not contradicted by ClinVar, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.040537 | Structured | 0.036106 | Uncertain | 0.966 | 0.236 | 0.000 | -13.669 | Likely Pathogenic | 0.705 | Likely Pathogenic | Likely Benign | 2.52 | Destabilizing | 0.3 | 0.76 | Ambiguous | 1.64 | Ambiguous | 0.75 | Ambiguous | 0.782 | Likely Pathogenic | -3.12 | Deleterious | 0.935 | Possibly Damaging | 0.473 | Possibly Damaging | -1.46 | Pathogenic | 0.01 | Affected | 0.0443 | 0.1964 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1999A>T | I667F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I667F missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: benign calls are limited to FATHMM, while the remaining 11 predictors—SGM‑Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Foldetta—classify the change as pathogenic. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) reports a pathogenic effect. No prediction is inconclusive. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.142424 | Structured | 0.083597 | Uncertain | 0.927 | 0.379 | 0.000 | -14.389 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 7.55 | Destabilizing | 0.3 | 1.80 | Ambiguous | 4.68 | Destabilizing | 0.86 | Ambiguous | 0.569 | Likely Pathogenic | -3.98 | Deleterious | 0.998 | Probably Damaging | 0.790 | Possibly Damaging | 2.96 | Benign | 0.00 | Affected | 0.0668 | 0.2648 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.2047A>T | I683F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I683F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, and FATHMM, whereas a majority of tools (SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. High‑accuracy assessments show the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, while AlphaMissense‑Optimized and Foldetta are inconclusive and thus treated as unavailable evidence. Overall, the balance of evidence favors a pathogenic classification for I683F, and this assessment does not contradict the current ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.200174 | Structured | 0.143268 | Uncertain | 0.848 | 0.314 | 0.000 | -12.781 | Likely Pathogenic | 0.828 | Likely Pathogenic | Ambiguous | 1.38 | Ambiguous | 0.1 | -0.23 | Likely Benign | 0.58 | Ambiguous | 0.59 | Ambiguous | 0.481 | Likely Benign | -3.99 | Deleterious | 0.971 | Probably Damaging | 0.499 | Possibly Damaging | 3.27 | Benign | 0.01 | Affected | 0.0770 | 0.2307 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.205A>T | I69F 2D AIThe SynGAP1 missense variant I69F is reported in gnomAD (variant ID 6‑33425813‑A‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority‑vote) is also benign. Foldetta results are unavailable. Overall, the preponderance of evidence indicates that I69F is most likely benign, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.575842 | Disordered | 0.466129 | Uncertain | 0.437 | 0.786 | 0.375 | 6-33425813-A-T | 2 | 1.24e-6 | -3.747 | Likely Benign | 0.251 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -0.99 | Neutral | 0.824 | Possibly Damaging | 0.507 | Possibly Damaging | 4.12 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0412 | 0.2743 | 0 | 1 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||
| c.2161A>T | I721F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I721F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated algorithms—FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic or likely pathogenic impact, while premPS remains uncertain. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. Based on the overwhelming agreement among these predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.394753 | Structured | 0.454550 | Uncertain | 0.957 | 0.437 | 0.125 | -12.559 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 4.61 | Destabilizing | 0.1 | 2.74 | Destabilizing | 3.68 | Destabilizing | 0.66 | Ambiguous | 0.295 | Likely Benign | -3.74 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 2.22 | Pathogenic | 0.00 | Affected | 0.0420 | 0.2931 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.2188A>T | I730F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I730F has no ClinVar entry and is not reported in gnomAD. Prediction tools largely agree on a benign effect: SGM‑Consensus (Likely Benign), REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen2_HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign or tolerated. Only polyPhen2_HumDiv classifies the change as pathogenic. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized scores the variant as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates Likely Benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, reports a benign effect. No prediction tool or stability analysis is inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.420109 | Uncertain | 0.591 | 0.619 | 0.750 | -4.953 | Likely Benign | 0.173 | Likely Benign | Likely Benign | -0.54 | Ambiguous | 0.1 | -0.01 | Likely Benign | -0.28 | Likely Benign | -0.01 | Likely Benign | 0.055 | Likely Benign | -1.86 | Neutral | 0.699 | Possibly Damaging | 0.152 | Benign | 3.45 | Benign | 0.08 | Tolerated | 0.0509 | 0.2082 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||
| c.2299A>T | I767F 2D AIThe SynGAP1 missense variant I767F is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.321458 | Structured | 0.927771 | Binding | 0.369 | 0.872 | 0.125 | -3.618 | Likely Benign | 0.162 | Likely Benign | Likely Benign | 0.213 | Likely Benign | -1.37 | Neutral | 0.003 | Benign | 0.002 | Benign | 4.04 | Benign | 0.06 | Tolerated | 0.0643 | 0.3567 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.22A>T | I8F 2D AIThe SynGAP1 missense variant I8F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.490133 | Structured | 0.543080 | Binding | 0.341 | 0.916 | 0.625 | -3.000 | Likely Benign | 0.092 | Likely Benign | Likely Benign | 0.116 | Likely Benign | -0.29 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.02 | Benign | 0.00 | Affected | 0.0483 | 0.2871 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2479A>T | I827F 2D AIThe SynGAP1 missense variant I827F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also reports it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact. The AlphaMissense‑Default score is uncertain, and no Foldetta stability assessment is available. High‑accuracy methods give a benign consensus: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign, with no Foldetta data. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.636272 | Binding | 0.383 | 0.884 | 0.625 | -4.799 | Likely Benign | 0.517 | Ambiguous | Likely Benign | 0.155 | Likely Benign | -1.30 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.65 | Benign | 0.09 | Tolerated | 0.0490 | 0.2306 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2695A>T | I899F 2D AIThe SynGAP1 missense variant I899F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.443727 | Uncertain | 0.292 | 0.928 | 0.375 | -4.049 | Likely Benign | 0.161 | Likely Benign | Likely Benign | 0.060 | Likely Benign | -1.05 | Neutral | 0.906 | Possibly Damaging | 0.473 | Possibly Damaging | 2.69 | Benign | 0.01 | Affected | 0.0593 | 0.2435 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2767A>T | I923F 2D AIThe SynGAP1 missense variant I923F is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.964857 | Binding | 0.292 | 0.852 | 0.250 | -2.967 | Likely Benign | 0.169 | Likely Benign | Likely Benign | 0.112 | Likely Benign | -0.85 | Neutral | 0.007 | Benign | 0.005 | Benign | 2.70 | Benign | 0.11 | Tolerated | 0.0726 | 0.3887 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2998A>T | I1000F 2D AIThe SynGAP1 missense variant I1000F is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of predictions indicates that I1000F is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.957020 | Binding | 0.293 | 0.904 | 0.625 | -3.801 | Likely Benign | 0.201 | Likely Benign | Likely Benign | 0.147 | Likely Benign | -1.02 | Neutral | 0.968 | Probably Damaging | 0.713 | Possibly Damaging | 2.70 | Benign | 0.13 | Tolerated | 0.0594 | 0.3259 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3109A>T | I1037F 2D AIThe SynGAP1 missense variant I1037F is not reported in ClinVar and has no entries in gnomAD. Consensus from multiple in‑silico predictors shows a split: benign predictions come from REVEL, PROVEAN, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy tools further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the majority of evidence points to a benign effect, and this assessment is consistent with the absence of a ClinVar pathogenic claim. Thus, the variant is most likely benign, and this conclusion does not contradict ClinVar, which has no pathogenic assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.939629 | Disordered | 0.986140 | Binding | 0.309 | 0.774 | 0.625 | -3.517 | Likely Benign | 0.653 | Likely Pathogenic | Likely Benign | 0.115 | Likely Benign | -0.97 | Neutral | 0.977 | Probably Damaging | 0.632 | Possibly Damaging | 2.71 | Benign | 0.16 | Tolerated | 0.0653 | 0.3722 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3115A>T | I1039F 2D AIThe SynGAP1 missense variant I1039F is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign interpretation: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all predict benign, while the majority of other predictors (polyPhen‑2 HumDiv and HumVar) indicate pathogenic. When predictions are grouped by agreement, the benign‑oriented tools outnumber the pathogenic ones, and the single uncertain call from AlphaMissense‑Default does not alter this balance. High‑accuracy assessments reinforce the benign trend: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports it as Likely Benign. Foldetta data are unavailable. Overall, the computational evidence overwhelmingly supports a benign effect, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.979204 | Binding | 0.292 | 0.806 | 0.625 | -2.872 | Likely Benign | 0.545 | Ambiguous | Likely Benign | 0.124 | Likely Benign | -1.16 | Neutral | 0.925 | Possibly Damaging | 0.510 | Possibly Damaging | 2.68 | Benign | 0.13 | Tolerated | 0.0710 | 0.4368 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3343A>T | I1115F 2D AIThe SynGAP1 missense variant I1115F is reported in gnomAD (ID 6‑33443895‑A‑T) and has no ClinVar entry. All available in silico predictors classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are not available. Based on the unanimous benign predictions and the lack of a ClinVar pathogenic classification, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.889439 | Disordered | 0.892339 | Binding | 0.308 | 0.912 | 0.750 | 6-33443895-A-T | -3.426 | Likely Benign | 0.095 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -0.98 | Neutral | 0.230 | Benign | 0.098 | Benign | 2.71 | Benign | 0.19 | Tolerated | 4.32 | 2 | 0.0769 | 0.4338 | 0 | 1 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||
| c.3397A>T | I1133F 2D AIThe SynGAP1 missense variant I1133F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect. The variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.832785 | Binding | 0.316 | 0.892 | 0.750 | -2.941 | Likely Benign | 0.164 | Likely Benign | Likely Benign | 0.272 | Likely Benign | -1.21 | Neutral | 0.290 | Benign | 0.124 | Benign | 5.45 | Benign | 0.05 | Affected | 0.0598 | 0.3522 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3502A>T | I1168F 2D AIThe SynGAP1 missense variant I1168F is not reported in ClinVar and is absent from gnomAD. Prediction tools show a mixed signal: benign calls come from REVEL, PROVEAN, ESM1b, and FATHMM, while pathogenic calls come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments are limited: AlphaMissense‑Optimized is uncertain, and Foldetta results are unavailable. Considering the majority of individual predictors and the SGM‑Consensus outcome, the variant is most likely benign. This assessment does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.763262 | Binding | 0.423 | 0.796 | 0.500 | -3.422 | Likely Benign | 0.879 | Likely Pathogenic | Ambiguous | 0.440 | Likely Benign | -1.21 | Neutral | 0.998 | Probably Damaging | 0.958 | Probably Damaging | 5.45 | Benign | 0.04 | Affected | 0.0616 | 0.3524 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3517A>T | I1173F 2D AIThe SynGAP1 missense variant I1173F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (both HumDiv and HumVar) predict a pathogenic outcome. AlphaMissense‑Default remains uncertain, and the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect for I1173F, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.501700 | Disordered | 0.653145 | Binding | 0.521 | 0.756 | 0.375 | -3.635 | Likely Benign | 0.491 | Ambiguous | Likely Benign | 0.365 | Likely Benign | -0.87 | Neutral | 0.934 | Possibly Damaging | 0.636 | Possibly Damaging | 5.39 | Benign | 0.13 | Tolerated | 0.0567 | 0.2108 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3604A>T | I1202F 2D AIThe SynGAP1 missense variant I1202F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) is likely pathogenic. Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.510422 | Binding | 0.874 | 0.593 | 0.250 | -12.304 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.188 | Likely Benign | -3.23 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.81 | Pathogenic | 0.02 | Affected | 0.0583 | 0.2079 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3697A>T | I1233F 2D AIThe SynGAP1 missense variant I1233F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, more tools predict pathogenicity (five) than benignity (three), and no ClinVar evidence contradicts this assessment. Thus, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -8.414 | Likely Pathogenic | 0.861 | Likely Pathogenic | Ambiguous | 0.075 | Likely Benign | -2.34 | Neutral | 0.968 | Probably Damaging | 0.713 | Possibly Damaging | 2.56 | Benign | 0.03 | Affected | 0.0418 | 0.2731 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3775A>T | I1259F 2D AIThe SynGAP1 missense variant I1259F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and FATHMM, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are less decisive: AlphaMissense‑Optimized returns an uncertain result, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta data are unavailable. Overall, the majority of evidence (five pathogenic vs. three benign) points toward a pathogenic effect. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.494003 | Structured | 0.576405 | Binding | 0.885 | 0.574 | 0.250 | -10.666 | Likely Pathogenic | 0.955 | Likely Pathogenic | Ambiguous | 0.301 | Likely Benign | -1.68 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.55 | Benign | 0.03 | Affected | 0.0428 | 0.2563 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3784A>T | I1262F 2D AIThe SynGAP1 missense variant I1262F is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess functional impact uniformly indicate a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic. No tool in the dataset predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the evidence strongly supports that the variant is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -11.343 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.515 | Likely Pathogenic | -3.32 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.81 | Pathogenic | 0.00 | Affected | 0.0532 | 0.2731 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3787A>T | I1263F 2D AIThe SynGAP1 missense variant I1263F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta results are unavailable. Because the majority of evidence points to a deleterious effect and there is no ClinVar annotation to contradict this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -5.887 | Likely Benign | 0.952 | Likely Pathogenic | Ambiguous | 0.335 | Likely Benign | -3.32 | Deleterious | 0.968 | Probably Damaging | 0.637 | Possibly Damaging | 1.81 | Pathogenic | 0.00 | Affected | 0.0494 | 0.2375 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.37A>T | I13F 2D AIThe SynGAP1 missense variant I13F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | -3.359 | Likely Benign | 0.126 | Likely Benign | Likely Benign | 0.134 | Likely Benign | -0.09 | Neutral | 0.107 | Benign | 0.010 | Benign | 4.04 | Benign | 0.00 | Affected | 0.0654 | 0.3972 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3991A>T | I1331F 2D AIThe SynGAP1 missense variant I1331F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain” and the SGM‑Consensus as “Likely Benign”; Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign interpretation, with no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -4.220 | Likely Benign | 0.946 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -2.46 | Neutral | 0.984 | Probably Damaging | 0.969 | Probably Damaging | 3.31 | Benign | 0.00 | Affected | 0.0522 | 0.3225 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.421A>T | I141F 2D AIThe SynGAP1 missense variant I141F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2). Foldetta results are unavailable. Overall, the balance of evidence slightly favors a benign interpretation, with no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.465241 | Structured | 0.577021 | Binding | 0.367 | 0.877 | 0.500 | -9.621 | Likely Pathogenic | 0.980 | Likely Pathogenic | Likely Pathogenic | 0.221 | Likely Benign | -1.82 | Neutral | 0.001 | Benign | 0.002 | Benign | 3.57 | Benign | 0.01 | Affected | 0.0489 | 0.2775 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.475A>T | I159F 2D AIThe SynGAP1 missense variant I159F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence, particularly from the high‑accuracy tools, points to a benign impact. This conclusion is not contradicted by any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -9.530 | Likely Pathogenic | 0.447 | Ambiguous | Likely Benign | 0.200 | Likely Benign | -1.41 | Neutral | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.87 | Benign | 0.00 | Affected | 0.0490 | 0.2446 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.613A>T | I205F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I205F missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: SGM‑Consensus, REVEL, FoldX, PROVEAN, SIFT, AlphaMissense‑Default, AlphaMissense‑Optimized, FATHMM, and polyPhen‑2 HumVar all classify it as benign. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. Uncertain results from Foldetta, premPS, ESM1b, and Rosetta are treated as unavailable. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely benign, while Foldetta remains uncertain. Overall, the consensus of the majority of tools indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.264545 | Structured | 0.409933 | Uncertain | 0.821 | 0.414 | 0.125 | -7.913 | In-Between | 0.271 | Likely Benign | Likely Benign | 0.30 | Likely Benign | 0.3 | 0.89 | Ambiguous | 0.60 | Ambiguous | 0.52 | Ambiguous | 0.113 | Likely Benign | -1.80 | Neutral | 0.838 | Possibly Damaging | 0.368 | Benign | 4.08 | Benign | 0.10 | Tolerated | 0.0461 | 0.2205 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.616A>T | I206F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I206F missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and polyPhen‑2 HumVar. Those that predict a pathogenic effect comprise SGM‑Consensus, Foldetta, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Rosetta. Predictions that are uncertain or inconclusive are FoldX and premPS. High‑accuracy methods all support a deleterious outcome: AlphaMissense‑Optimized reports pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.298791 | Structured | 0.405123 | Uncertain | 0.863 | 0.391 | 0.125 | -12.669 | Likely Pathogenic | 0.961 | Likely Pathogenic | Likely Pathogenic | 1.82 | Ambiguous | 0.4 | 3.39 | Destabilizing | 2.61 | Destabilizing | 0.70 | Ambiguous | 0.133 | Likely Benign | -3.40 | Deleterious | 0.838 | Possibly Damaging | 0.368 | Benign | 3.64 | Benign | 0.01 | Affected | 0.0428 | 0.2721 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.637A>T | I213F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I213F missense variant is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only FATHMM, while the remaining evaluated algorithms (SGM‑Consensus, REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact; FoldX and premPS are uncertain and are treated as unavailable evidence. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain and thus not considered. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which simply lacks an entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.158265 | Structured | 0.372201 | Uncertain | 0.850 | 0.295 | 0.125 | -12.212 | Likely Pathogenic | 0.980 | Likely Pathogenic | Likely Pathogenic | 0.96 | Ambiguous | 0.7 | 2.66 | Destabilizing | 1.81 | Ambiguous | 0.73 | Ambiguous | 0.815 | Likely Pathogenic | -3.30 | Deleterious | 0.995 | Probably Damaging | 0.829 | Possibly Damaging | 5.79 | Benign | 0.01 | Affected | 0.0473 | 0.2560 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.727A>T | I243F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I243F missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give inconclusive results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and thus unavailable; Foldetta is uncertain. Overall, the majority of available predictions (7 pathogenic vs. 3 benign) indicate a likely pathogenic impact. This assessment does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.363090 | Structured | 0.344471 | Uncertain | 0.842 | 0.347 | 0.000 | -12.559 | Likely Pathogenic | 0.934 | Likely Pathogenic | Ambiguous | 2.92 | Destabilizing | 2.5 | 0.53 | Ambiguous | 1.73 | Ambiguous | 0.42 | Likely Benign | 0.793 | Likely Pathogenic | -2.21 | Neutral | 0.985 | Probably Damaging | 0.724 | Possibly Damaging | 5.53 | Benign | 0.02 | Affected | 0.0409 | 0.2205 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||
| c.802A>T | I268F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I268F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that assess pathogenicity uniformly indicate a deleterious effect: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity, while the SGM‑Consensus score is “Likely Pathogenic.” No tool in the dataset predicts a benign outcome; only Rosetta and Foldetta are uncertain. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) remains uncertain. Overall, the consensus of available predictions strongly supports a pathogenic classification for I268F, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.216401 | Structured | 0.314336 | Uncertain | 0.951 | 0.264 | 0.000 | -11.426 | Likely Pathogenic | 0.968 | Likely Pathogenic | Likely Pathogenic | 2.85 | Destabilizing | 0.7 | 0.59 | Ambiguous | 1.72 | Ambiguous | 1.00 | Destabilizing | 0.685 | Likely Pathogenic | -3.68 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.53 | Pathogenic | 0.00 | Affected | 0.0427 | 0.2158 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||
| c.1091C>A | P364H 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant P364H is reported in gnomAD (ID 6‑33437996‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, and AlphaMissense‑Optimized; pathogenic predictions come from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments further support this dichotomy: AlphaMissense‑Optimized classifies the variant as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels it pathogenic. The protein‑folding stability predictor Foldetta, which integrates FoldX‑MD and Rosetta outputs, is inconclusive. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not conflict with ClinVar status, which currently lacks an entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.390993 | Structured | 0.439474 | Uncertain | 0.942 | 0.590 | 0.250 | 6-33437996-C-A | -10.744 | Likely Pathogenic | 0.632 | Likely Pathogenic | Likely Benign | 1.65 | Ambiguous | 0.9 | 0.25 | Likely Benign | 0.95 | Ambiguous | 0.77 | Ambiguous | 0.407 | Likely Benign | -6.96 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.55 | Pathogenic | 0.02 | Affected | 3.39 | 20 | 0.1800 | 0.4584 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||
| c.1238C>A | P413H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P413H is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include only FATHMM, while the remaining tools—SGM‑Consensus, REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic or likely pathogenic impact; Rosetta is uncertain. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No predictions are missing or inconclusive. Based on the overwhelming agreement among pathogenic predictors and the high‑accuracy tools, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.332472 | Uncertain | 0.927 | 0.201 | 0.000 | -13.376 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 4.89 | Destabilizing | 1.0 | 1.85 | Ambiguous | 3.37 | Destabilizing | 1.07 | Destabilizing | 0.546 | Likely Pathogenic | -8.29 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.16 | Benign | 0.00 | Affected | 0.1745 | 0.3480 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.125C>A | P42H 2D AIThe SynGAP1 missense variant P42H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.308712 | Structured | 0.431487 | Uncertain | 0.420 | 0.771 | 0.375 | -3.088 | Likely Benign | 0.167 | Likely Benign | Likely Benign | 0.140 | Likely Benign | -1.95 | Neutral | 0.992 | Probably Damaging | 0.977 | Probably Damaging | 4.16 | Benign | 0.00 | Affected | 0.1995 | 0.3705 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.1310C>A | P437H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P437H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Predictions that are inconclusive or unavailable are FoldX, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show that the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts a likely pathogenic outcome, while AlphaMissense‑Optimized and Foldetta remain uncertain. Overall, the balance of evidence favors a pathogenic classification for P437H, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.306196 | Uncertain | 0.921 | 0.298 | 0.000 | -13.496 | Likely Pathogenic | 0.826 | Likely Pathogenic | Ambiguous | 1.19 | Ambiguous | 0.0 | -3.28 | Stabilizing | -1.05 | Ambiguous | 0.31 | Likely Benign | 0.343 | Likely Benign | -7.75 | Deleterious | 1.000 | Probably Damaging | 0.995 | Probably Damaging | 3.44 | Benign | 0.01 | Affected | 0.1819 | 0.4328 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.137C>A | P46H 2D AIThe SynGAP1 missense variant P46H is evaluated by multiple in silico tools. Consensus from SGM‑Consensus indicates a likely benign effect, and the variant is not reported in ClinVar or gnomAD. Functional predictors that agree on a benign outcome include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Predictors that flag a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also reports likely benign, while Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact for P46H, and this conclusion is not contradicted by any ClinVar annotation. The variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.390993 | Structured | 0.433588 | Uncertain | 0.549 | 0.741 | 0.375 | -4.022 | Likely Benign | 0.428 | Ambiguous | Likely Benign | 0.097 | Likely Benign | -0.43 | Neutral | 0.992 | Probably Damaging | 0.977 | Probably Damaging | 4.10 | Benign | 0.00 | Affected | 0.2360 | 0.5229 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.1583C>A | P528H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P528H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely indicate a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the change as pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also reports it as likely pathogenic. No tool predicts a benign outcome; the remaining predictions (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized) are uncertain or inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta as uncertain. Taken together, the preponderance of evidence supports a pathogenic interpretation, and this is consistent with the absence of a ClinVar entry; there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.225814 | Structured | 0.020396 | Uncertain | 0.909 | 0.403 | 0.000 | -15.365 | Likely Pathogenic | 0.940 | Likely Pathogenic | Ambiguous | 1.93 | Ambiguous | 0.1 | 0.74 | Ambiguous | 1.34 | Ambiguous | 0.61 | Ambiguous | 0.565 | Likely Pathogenic | -8.61 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.47 | Pathogenic | 0.00 | Affected | 0.1434 | 0.3523 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.1814C>A | P605H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P605H is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts a benign outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. All available predictions are consistent and indicate a deleterious effect. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.023087 | Structured | 0.192737 | Uncertain | 0.929 | 0.231 | 0.000 | -13.846 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 8.80 | Destabilizing | 2.7 | 6.69 | Destabilizing | 7.75 | Destabilizing | 0.87 | Ambiguous | 0.828 | Likely Pathogenic | -8.95 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 0.68 | Pathogenic | 0.00 | Affected | 0.1778 | 0.4142 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.197C>A | P66H 2D AIThe SynGAP1 missense variant P66H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and the SGM‑Consensus result is benign; Foldetta predictions are unavailable. Overall, the balance of evidence leans toward a benign interpretation, with no conflict with ClinVar status because no ClinVar assertion exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.394753 | Structured | 0.474132 | Uncertain | 0.455 | 0.762 | 0.125 | -4.134 | Likely Benign | 0.941 | Likely Pathogenic | Ambiguous | 0.239 | Likely Benign | -2.26 | Neutral | 0.992 | Probably Damaging | 0.893 | Possibly Damaging | 3.89 | Benign | 0.00 | Affected | 0.1914 | 0.4675 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2102C>A | P701H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P701H missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Predictions that are uncertain or inconclusive are FoldX, Foldetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict the ClinVar status, which contains no pathogenic assertion for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.404318 | Uncertain | 0.918 | 0.345 | 0.000 | -8.786 | Likely Pathogenic | 0.553 | Ambiguous | Likely Benign | 1.40 | Ambiguous | 0.0 | -0.41 | Likely Benign | 0.50 | Ambiguous | 0.62 | Ambiguous | 0.141 | Likely Benign | -2.12 | Neutral | 0.936 | Possibly Damaging | 0.539 | Possibly Damaging | 3.38 | Benign | 0.03 | Affected | 0.1495 | 0.3495 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||
| c.2144C>A | P715H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P715H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, Rosetta, and FATHMM, whereas a majority of predictors (FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus) indicate a pathogenic impact. Tools with inconclusive results (Foldetta, premPS, AlphaMissense‑Optimized) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta as uncertain. Overall, the consensus of the majority of evidence points to a pathogenic effect for P715H. Based on the aggregate predictions, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.243554 | Structured | 0.409757 | Uncertain | 0.956 | 0.362 | 0.000 | -10.523 | Likely Pathogenic | 0.906 | Likely Pathogenic | Ambiguous | 2.80 | Destabilizing | 0.0 | 0.28 | Likely Benign | 1.54 | Ambiguous | 0.56 | Ambiguous | 0.271 | Likely Benign | -7.73 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.37 | Benign | 0.00 | Affected | 0.1413 | 0.3684 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.2183C>A | P728H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P728H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default, all of which predict a deleterious impact. Predictions that are inconclusive or uncertain are FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show that AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Overall, the majority of evidence points to a pathogenic effect for P728H, and this conclusion does not contradict any ClinVar status because the variant is not yet classified in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.632174 | Disordered | 0.434760 | Uncertain | 0.725 | 0.567 | 0.625 | -8.897 | Likely Pathogenic | 0.945 | Likely Pathogenic | Ambiguous | 0.94 | Ambiguous | 0.0 | 0.86 | Ambiguous | 0.90 | Ambiguous | 0.64 | Ambiguous | 0.402 | Likely Benign | -7.23 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 0.65 | Pathogenic | 0.00 | Affected | 0.1993 | 0.3016 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||
| c.2222C>A | P741H 2D AIThe SynGAP1 missense variant P741H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. The prediction is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.885302 | Disordered | 0.493550 | Uncertain | 0.354 | 0.859 | 0.875 | -5.592 | Likely Benign | 0.085 | Likely Benign | Likely Benign | 0.099 | Likely Benign | -0.99 | Neutral | 0.006 | Benign | 0.007 | Benign | 2.81 | Benign | 0.01 | Affected | 0.1372 | 0.3831 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2228C>A | P743H 2D AIThe SynGAP1 missense variant P743H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available, so it does not influence the overall assessment. Overall, the majority of computational evidence points to a benign effect for P743H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.526809 | Binding | 0.317 | 0.862 | 0.875 | -5.649 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.121 | Likely Benign | -1.99 | Neutral | 0.989 | Probably Damaging | 0.870 | Possibly Damaging | 2.70 | Benign | 0.01 | Affected | 0.1653 | 0.3762 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2234C>A | P745H 2D AIThe SynGAP1 missense variant P745H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.791621 | Disordered | 0.558331 | Binding | 0.341 | 0.860 | 0.875 | -5.569 | Likely Benign | 0.120 | Likely Benign | Likely Benign | 0.200 | Likely Benign | -2.84 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.53 | Benign | 0.00 | Affected | 0.1912 | 0.3403 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2360C>A | P787H 2D AIThe SynGAP1 missense variant P787H has no ClinVar entry and is not listed in gnomAD. Prediction tools that classify the variant as benign include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default—predict it to be pathogenic. The SGM‑Consensus, which is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Pathogenic” (3 pathogenic votes versus 1 benign). High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains pathogenic; Foldetta results are unavailable. Overall, the majority of predictions lean toward pathogenicity, but this is contradicted by the benign call from AlphaMissense‑Optimized. Because ClinVar has no reported status, there is no conflict with existing clinical annotations. Thus, the variant is most likely pathogenic based on the prevailing computational evidence, though one high‑accuracy tool suggests a benign effect. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | SH3-binding motif | 0.901269 | Disordered | 0.613211 | Binding | 0.377 | 0.899 | 0.750 | -5.819 | Likely Benign | 0.691 | Likely Pathogenic | Likely Benign | 0.308 | Likely Benign | -4.96 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.44 | Pathogenic | 0.01 | Affected | 0.1790 | 0.4061 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||
| c.2384C>A | P795H 2D AIThe SynGAP1 missense variant P795H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic outcome. ESM1b is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.972450 | Disordered | 0.410339 | Uncertain | 0.457 | 0.903 | 0.875 | -7.717 | In-Between | 0.121 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -0.61 | Neutral | 0.898 | Possibly Damaging | 0.477 | Possibly Damaging | 4.24 | Benign | 0.05 | Affected | 0.1947 | 0.5184 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||
| c.2396C>A | P799H 2D AIThe SynGAP1 missense variant P799H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree that the substitution is benign: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign effect. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence supports a benign classification for P799H, and this conclusion does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.871313 | Disordered | 0.537892 | Binding | 0.400 | 0.894 | 0.750 | -5.611 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.089 | Likely Benign | -0.80 | Neutral | 0.999 | Probably Damaging | 0.933 | Probably Damaging | 4.20 | Benign | 0.00 | Affected | 0.1684 | 0.4192 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||
| c.2435C>A | P812H 2D AIThe SynGAP1 missense variant P812H is listed in ClinVar with an uncertain significance and is present in the gnomAD database (ID 6‑33442987‑C‑A). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic outcome; ESM1b remains uncertain. High‑accuracy methods give a benign result from AlphaMissense‑Optimized, a pathogenic consensus from the SGM approach (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta data are unavailable. Overall, the balance of evidence points to a pathogenic effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.414856 | Structured | 0.842442 | Binding | 0.388 | 0.901 | 0.125 | Uncertain | 2 | 6-33442987-C-A | 3 | 1.86e-6 | -7.470 | In-Between | 0.698 | Likely Pathogenic | Likely Benign | 0.272 | Likely Benign | -2.81 | Deleterious | 1.000 | Probably Damaging | 0.995 | Probably Damaging | 2.68 | Benign | 0.00 | Affected | 4.32 | 4 | 0.1568 | 0.4923 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||
| c.2552C>A | P851H 2D AIThe SynGAP1 missense variant P851H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict pathogenicity. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.648219 | Disordered | 0.526893 | Binding | 0.347 | 0.819 | 0.625 | -4.730 | Likely Benign | 0.102 | Likely Benign | Likely Benign | 0.184 | Likely Benign | 0.19 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 4.18 | Benign | 0.15 | Tolerated | 0.1577 | 0.5378 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2654C>A | P885H 2D AIThe SynGAP1 missense variant P885H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence, including the high‑accuracy tools, points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.690604 | Disordered | 0.636133 | Binding | 0.344 | 0.917 | 0.250 | -5.239 | Likely Benign | 0.171 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -1.68 | Neutral | 0.938 | Possibly Damaging | 0.749 | Possibly Damaging | 2.74 | Benign | 0.00 | Affected | 0.1552 | 0.5280 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2660C>A | P887H 2D AIThe SynGAP1 missense variant P887H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (both HumDiv and HumVar models) predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect for P887H, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.716283 | Disordered | 0.602269 | Binding | 0.348 | 0.925 | 0.500 | -4.900 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.072 | Likely Benign | -1.71 | Neutral | 0.977 | Probably Damaging | 0.777 | Possibly Damaging | 2.73 | Benign | 0.13 | Tolerated | 0.1236 | 0.3275 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2750C>A | P917H 2D AIThe SynGAP1 missense variant P917H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence—including the high‑accuracy tools—points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.599170 | Disordered | 0.863949 | Binding | 0.314 | 0.862 | 0.375 | -5.395 | Likely Benign | 0.182 | Likely Benign | Likely Benign | 0.122 | Likely Benign | -2.00 | Neutral | 0.975 | Probably Damaging | 0.766 | Possibly Damaging | 2.65 | Benign | 0.00 | Affected | 0.1584 | 0.4410 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2771C>A | P924H 2D AIThe SynGAP1 missense variant P924H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of computational evidence indicates that P924H is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.521092 | Disordered | 0.971858 | Binding | 0.293 | 0.846 | 0.250 | -6.236 | Likely Benign | 0.957 | Likely Pathogenic | Likely Pathogenic | 0.457 | Likely Benign | -5.87 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 0.65 | Pathogenic | 0.00 | Affected | 0.1317 | 0.3281 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2789C>A | P930H 2D AIThe SynGAP1 missense variant P930H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated predictors—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence from multiple independent predictors and high‑accuracy tools indicates that P930H is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.538167 | Disordered | 0.988036 | Binding | 0.304 | 0.855 | 0.375 | -9.824 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.497 | Likely Benign | -6.44 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 0.65 | Pathogenic | 0.00 | Affected | 0.1942 | 0.4392 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.281C>A | P94H 2D AIThe SynGAP1 missense variant P94H is reported in gnomAD (variant ID 6‑33425889‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 HumDiv and SIFT predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. No Foldetta stability analysis is available, so it does not influence the assessment. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that P94H is most likely benign, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.570978 | Binding | 0.350 | 0.869 | 0.625 | 6-33425889-C-A | 1 | 6.20e-7 | -3.708 | Likely Benign | 0.106 | Likely Benign | Likely Benign | 0.077 | Likely Benign | -2.31 | Neutral | 0.637 | Possibly Damaging | 0.102 | Benign | 4.11 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1486 | 0.3913 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||
| c.2822C>A | P941H 2D AIThe SynGAP1 missense variant P941H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates a likely benign outcome. In contrast, two tools—polyPhen‑2 HumDiv and SIFT—classify the change as pathogenic. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; no Foldetta stability data are available. Overall, the majority of evidence points to a benign effect, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.874069 | Disordered | 0.900790 | Binding | 0.403 | 0.906 | 0.625 | -4.439 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.058 | Likely Benign | -0.09 | Neutral | 0.589 | Possibly Damaging | 0.309 | Benign | 2.71 | Benign | 0.00 | Affected | 0.1621 | 0.4826 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2861C>A | P954H 2D AIThe SynGAP1 missense variant P954H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of ClinVar or gnomAD entries—there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984159 | Disordered | 0.932268 | Binding | 0.465 | 0.926 | 0.750 | -3.670 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.098 | Likely Benign | -0.50 | Neutral | 0.041 | Benign | 0.067 | Benign | 2.75 | Benign | 0.04 | Affected | 0.2054 | 0.4756 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2912C>A | P971H 2D AIThe SynGAP1 missense variant P971H is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443464‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; a Foldetta stability prediction is not available. Overall, the majority of evidence points to a benign impact, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.950334 | Disordered | 0.951523 | Binding | 0.545 | 0.905 | 0.625 | Uncertain | 1 | 6-33443464-C-A | 1 | 6.20e-7 | -5.243 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.039 | Likely Benign | -1.11 | Neutral | 0.898 | Possibly Damaging | 0.477 | Possibly Damaging | 3.89 | Benign | 0.00 | Affected | 4.32 | 2 | 0.1584 | 0.4858 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||
| c.2915C>A | P972H 2D AIThe SynGAP1 missense variant P972H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools (polyPhen‑2 HumDiv and SIFT) predict pathogenicity, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign classification, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.954150 | Binding | 0.472 | 0.904 | 0.625 | -4.791 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.049 | Likely Benign | -1.62 | Neutral | 0.589 | Possibly Damaging | 0.229 | Benign | 4.19 | Benign | 0.02 | Affected | 0.1930 | 0.4798 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2978C>A | P993H 2D AIThe SynGAP1 missense variant P993H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P993H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.923979 | Binding | 0.319 | 0.908 | 0.750 | -4.535 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.053 | Likely Benign | -0.93 | Neutral | 0.938 | Possibly Damaging | 0.819 | Possibly Damaging | 4.11 | Benign | 0.00 | Affected | 0.1799 | 0.4591 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2984C>A | P995H 2D AIThe SynGAP1 missense variant P995H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P995H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.935305 | Binding | 0.338 | 0.902 | 0.750 | -5.347 | Likely Benign | 0.107 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.75 | Neutral | 0.832 | Possibly Damaging | 0.600 | Possibly Damaging | 4.16 | Benign | 0.00 | Affected | 0.1618 | 0.4571 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2987C>A | P996H 2D AIThe SynGAP1 missense variant P996H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.775545 | Disordered | 0.942262 | Binding | 0.312 | 0.900 | 0.750 | -5.554 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 0.045 | Likely Benign | -1.19 | Neutral | 0.001 | Benign | 0.003 | Benign | 4.25 | Benign | 0.00 | Affected | 0.1585 | 0.5160 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3035C>A | P1012H 2D AIThe SynGAP1 missense variant P1012H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Benign. Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect for P1012H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.894674 | Binding | 0.319 | 0.866 | 0.625 | -4.877 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.024 | Likely Benign | -0.38 | Neutral | 0.832 | Possibly Damaging | 0.600 | Possibly Damaging | 2.75 | Benign | 0.08 | Tolerated | 0.1553 | 0.4635 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3101C>A | P1034H 2D AIThe SynGAP1 missense variant P1034H is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. three benign) lean toward pathogenicity, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.926919 | Disordered | 0.991713 | Binding | 0.343 | 0.752 | 0.625 | -4.634 | Likely Benign | 0.540 | Ambiguous | Likely Benign | 0.083 | Likely Benign | -3.17 | Deleterious | 0.938 | Possibly Damaging | 0.750 | Possibly Damaging | 2.38 | Pathogenic | 0.02 | Affected | 0.1776 | 0.5451 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3104C>A | P1035H 2D AIThe SynGAP1 missense variant P1035H is not reported in ClinVar and has no entries in gnomAD. Consensus predictions from multiple in‑silico tools are mixed: benign calls come from SGM‑Consensus (Likely Benign), REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls are made by polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates a likely benign outcome. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.945666 | Disordered | 0.989572 | Binding | 0.300 | 0.756 | 0.625 | -5.333 | Likely Benign | 0.608 | Likely Pathogenic | Likely Benign | 0.058 | Likely Benign | -0.76 | Neutral | 0.997 | Probably Damaging | 0.889 | Possibly Damaging | 2.68 | Benign | 0.15 | Tolerated | 0.1893 | 0.5669 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3122C>A | P1041H 2D AIThe SynGAP1 missense variant P1041H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, and Foldetta’s protein‑folding stability result is unavailable. Overall, the majority of high‑confidence tools predict a benign impact, and there is no conflict with ClinVar status because the variant has not been reported there. Thus, based on current predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.962114 | Disordered | 0.967463 | Binding | 0.345 | 0.833 | 0.625 | -5.312 | Likely Benign | 0.377 | Ambiguous | Likely Benign | 0.476 | Likely Benign | -3.32 | Deleterious | 0.999 | Probably Damaging | 0.917 | Probably Damaging | 5.45 | Benign | 0.03 | Affected | 0.1883 | 0.5434 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3137C>A | P1046H 2D AIThe SynGAP1 missense variant P1046H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Benign”). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for P1046H. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.970265 | Disordered | 0.942366 | Binding | 0.364 | 0.898 | 0.750 | -5.715 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -1.64 | Neutral | 0.832 | Possibly Damaging | 0.670 | Possibly Damaging | 2.33 | Pathogenic | 0.07 | Tolerated | 0.1585 | 0.5250 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3146C>A | P1049H 2D AIThe SynGAP1 missense variant P1049H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.917915 | Binding | 0.428 | 0.920 | 0.750 | -5.427 | Likely Benign | 0.115 | Likely Benign | Likely Benign | 0.066 | Likely Benign | -2.06 | Neutral | 0.978 | Probably Damaging | 0.750 | Possibly Damaging | 2.76 | Benign | 0.01 | Affected | 0.1800 | 0.5070 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3197C>A | P1066H 2D AIThe SynGAP1 missense variant P1066H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.967676 | Disordered | 0.968838 | Binding | 0.403 | 0.913 | 0.875 | -6.034 | Likely Benign | 0.185 | Likely Benign | Likely Benign | 0.146 | Likely Benign | -2.90 | Deleterious | 1.000 | Probably Damaging | 0.975 | Probably Damaging | 2.61 | Benign | 0.00 | Affected | 0.1984 | 0.5861 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3251C>A | P1084H 2D AIThe SynGAP1 missense variant P1084H is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443803‑C‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which reports “Likely Benign”). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar designation of uncertainty rather than a definitive pathogenic claim. Thus, the variant is most likely benign, and its prediction profile does not contradict the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.842060 | Disordered | 0.979020 | Binding | 0.348 | 0.889 | 1.000 | Uncertain | 1 | 6-33443803-C-A | 1 | 6.31e-7 | -4.125 | Likely Benign | 0.323 | Likely Benign | Likely Benign | 0.134 | Likely Benign | -3.16 | Deleterious | 0.997 | Probably Damaging | 0.840 | Possibly Damaging | 3.96 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1751 | 0.5523 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||
| c.3302C>A | P1101H 2D AIThe SynGAP1 missense variant P1101H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available. Overall, the majority of evidence points to a benign effect for P1101H, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.874069 | Disordered | 0.968967 | Binding | 0.457 | 0.861 | 0.875 | -5.370 | Likely Benign | 0.149 | Likely Benign | Likely Benign | 0.167 | Likely Benign | -1.87 | Neutral | 0.996 | Probably Damaging | 0.864 | Possibly Damaging | 4.18 | Benign | 0.02 | Affected | 0.2046 | 0.4597 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3392C>A | P1131H 2D AIThe SynGAP1 missense variant P1131H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the balance of evidence leans toward a benign classification, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | -5.207 | Likely Benign | 0.454 | Ambiguous | Likely Benign | 0.357 | Likely Benign | -3.50 | Deleterious | 0.971 | Probably Damaging | 0.750 | Possibly Damaging | 5.24 | Benign | 0.00 | Affected | 0.1519 | 0.5546 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3437C>A | P1146H 2D AIThe SynGAP1 missense variant P1146H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT) predict a pathogenic impact; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign outcome (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | -4.428 | Likely Benign | 0.518 | Ambiguous | Likely Benign | 0.529 | Likely Pathogenic | -4.93 | Deleterious | 1.000 | Probably Damaging | 0.983 | Probably Damaging | 5.46 | Benign | 0.00 | Affected | 0.1625 | 0.4512 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3485C>A | P1162H 2D AIThe SynGAP1 missense variant P1162H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as Pathogenic, whereas the SGM‑Consensus (majority vote) indicates Likely Benign; Foldetta results are unavailable. Overall, the majority of tools (six benign vs. four pathogenic) lean toward a benign interpretation, but the high‑accuracy AlphaMissense‑Optimized prediction conflicts with the consensus. Because ClinVar contains no entry, there is no contradiction with clinical database status. Based on the collective predictions, the variant is most likely benign, though the AlphaMissense‑Optimized result suggests caution. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.599170 | Disordered | 0.858809 | Binding | 0.366 | 0.823 | 0.375 | -3.733 | Likely Benign | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.168 | Likely Benign | -2.46 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.67 | Benign | 0.17 | Tolerated | 0.1481 | 0.5001 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.365C>A | P122H 2D AIThe SynGAP1 missense variant P122H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. Foldetta results are unavailable. Overall, the majority of evidence, including the high‑accuracy tools, points to a benign effect for P122H. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.672358 | Binding | 0.400 | 0.887 | 0.750 | -4.993 | Likely Benign | 0.321 | Likely Benign | Likely Benign | 0.096 | Likely Benign | -1.82 | Neutral | 0.996 | Probably Damaging | 0.750 | Possibly Damaging | 4.14 | Benign | 0.03 | Affected | 0.2306 | 0.4213 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.374C>A | P125H 2D AIThe SynGAP1 missense variant P125H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2). Foldetta results are unavailable. Overall, the majority of conventional predictors (five pathogenic vs four benign) lean toward a pathogenic interpretation, but the single high‑accuracy benign prediction and the inconclusive SGM Consensus temper this view. No ClinVar annotation is present, so there is no reported status to contradict the computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.704227 | Binding | 0.373 | 0.878 | 0.625 | -5.177 | Likely Benign | 0.583 | Likely Pathogenic | Likely Benign | 0.235 | Likely Benign | -4.18 | Deleterious | 0.996 | Probably Damaging | 0.750 | Possibly Damaging | 2.82 | Benign | 0.01 | Affected | 0.1598 | 0.4263 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3833C>A | P1278H 2D AIThe SynGAP1 missense variant P1278H is reported in gnomAD (variant ID 6‑33447881‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict pathogenic. High‑accuracy assessments confirm the benign prediction: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (derived from the same set of benign predictions) is benign; Foldetta results are unavailable. Overall, the majority of evidence supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic designation. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.806955 | Binding | 0.532 | 0.722 | 0.750 | 6-33447881-C-A | 7 | 4.51e-6 | -5.691 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.087 | Likely Benign | -1.69 | Neutral | 0.938 | Possibly Damaging | 0.477 | Possibly Damaging | 2.67 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1997 | 0.3155 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||
| c.383C>A | P128H 2D AIThe SynGAP1 missense variant P128H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic outcome. AlphaMissense‑Default remains uncertain, and the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.713069 | Binding | 0.376 | 0.878 | 0.625 | -4.806 | Likely Benign | 0.564 | Ambiguous | Likely Benign | 0.169 | Likely Benign | -1.90 | Neutral | 0.996 | Probably Damaging | 0.750 | Possibly Damaging | 4.14 | Benign | 0.03 | Affected | 0.2207 | 0.3414 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3860C>A | P1287H 2D AIThe SynGAP1 missense variant P1287H is recorded in gnomAD (6‑33447908‑C‑A) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) is not available for this variant. Based on the aggregate predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.813701 | Binding | 0.538 | 0.777 | 0.750 | 6-33447908-C-A | -3.606 | Likely Benign | 0.135 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -1.36 | Neutral | 0.938 | Possibly Damaging | 0.596 | Possibly Damaging | 2.80 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1306 | 0.3448 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.3899C>A | P1300H 2D AIThe SynGAP1 missense variant P1300H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.885826 | Binding | 0.400 | 0.834 | 0.875 | -5.062 | Likely Benign | 0.140 | Likely Benign | Likely Benign | 0.093 | Likely Benign | -1.48 | Neutral | 0.990 | Probably Damaging | 0.840 | Possibly Damaging | 2.80 | Benign | 0.01 | Affected | 0.1757 | 0.3802 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.3902C>A | P1301H 2D AIThe SynGAP1 missense variant P1301H is listed in ClinVar (ID 212356.0) with an “Uncertain” clinical significance and is present in gnomAD (variant ID 6‑33451776‑C‑A). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The high‑accuracy consensus methods report a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” No Foldetta stability result is available. Overall, the majority of predictions, including the high‑accuracy consensus, support a benign classification, which does not contradict the ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.728858 | Disordered | 0.885064 | Binding | 0.447 | 0.841 | 0.875 | Conflicting | 2 | 6-33451776-C-A | 5 | 3.10e-6 | -5.756 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.232 | Likely Benign | -1.13 | Neutral | 0.642 | Possibly Damaging | 0.378 | Benign | 2.79 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1305 | 0.2965 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||
| c.3959C>A | P1320H 2D AIThe SynGAP1 missense variant P1320H is listed in gnomAD (ID 6‑33451833‑C‑A) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | 6-33451833-C-A | 2 | 1.29e-6 | -6.296 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -1.05 | Neutral | 0.998 | Probably Damaging | 0.990 | Probably Damaging | 4.17 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1955 | 0.5461 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||
| c.3974C>A | P1325H 2D AIThe SynGAP1 missense variant P1325H is reported in gnomAD (variant ID 6‑33451848‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv and SIFT. The high‑accuracy consensus methods reinforce the benign assessment: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” No result is available from the Foldetta stability analysis. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451848-C-A | -6.970 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.056 | Likely Benign | 0.10 | Neutral | 0.704 | Possibly Damaging | 0.187 | Benign | 4.04 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2103 | 0.4364 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.3980C>A | P1327H 2D AIThe SynGAP1 missense variant P1327H is reported in gnomAD (ID 6‑33451854‑C‑A) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus is Likely Benign, and Foldetta results are unavailable. Based on the preponderance of evidence from both general and high‑accuracy predictors, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.896620 | Disordered | 0.900145 | Binding | 0.369 | 0.777 | 0.875 | 6-33451854-C-A | -5.496 | Likely Benign | 0.263 | Likely Benign | Likely Benign | 0.144 | Likely Benign | -0.49 | Neutral | 0.998 | Probably Damaging | 0.953 | Probably Damaging | 4.09 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1559 | 0.3932 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.41C>A | P14H 2D AIThe SynGAP1 missense variant P14H is listed in gnomAD (ID 6‑33420305‑C‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Overall, the majority of predictions—including the high‑accuracy tools—indicate that P14H is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | 6-33420305-C-A | -3.747 | Likely Benign | 0.231 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -0.27 | Neutral | 0.742 | Possibly Damaging | 0.091 | Benign | 4.15 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1999 | 0.5176 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.443C>A | P148H 2D AIThe SynGAP1 missense variant P148H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple in silico predictors points to a pathogenic effect for P148H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.653063 | Disordered | 0.500109 | Binding | 0.372 | 0.837 | 0.625 | -11.034 | Likely Pathogenic | 0.961 | Likely Pathogenic | Likely Pathogenic | 0.169 | Likely Benign | -3.21 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.90 | Benign | 0.00 | Affected | 0.1881 | 0.3847 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.482C>A | P161H 2D AIThe SynGAP1 missense variant P161H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus reports it as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple in silico predictors indicates that P161H is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -12.103 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 0.291 | Likely Benign | -4.22 | Deleterious | 0.964 | Probably Damaging | 0.650 | Possibly Damaging | 3.89 | Benign | 0.00 | Affected | 0.2060 | 0.4039 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.566C>A | P189H 2D AIThe SynGAP1 missense variant P189H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that P189H is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.428590 | Uncertain | 0.331 | 0.602 | 0.250 | -9.633 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.298 | Likely Benign | -6.49 | Deleterious | 0.999 | Probably Damaging | 0.936 | Probably Damaging | 4.00 | Benign | 0.00 | Affected | 0.1620 | 0.5327 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.59C>A | P20H 2D AIThe SynGAP1 missense variant P20H is reported in gnomAD (ID 6‑33420323‑C‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates benign. Foldetta results are unavailable. Overall, the preponderance of evidence from both consensus and high‑accuracy tools points to a benign effect for P20H, and this conclusion does not contradict any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.442804 | Uncertain | 0.448 | 0.899 | 0.500 | 6-33420323-C-A | -4.450 | Likely Benign | 0.352 | Ambiguous | Likely Benign | 0.132 | Likely Benign | -0.69 | Neutral | 0.992 | Probably Damaging | 0.893 | Possibly Damaging | 4.23 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1834 | 0.5066 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.755C>A | P252H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P252H missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include only FATHMM, while the remaining evaluated algorithms (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is uncertain. No other high‑confidence stability predictions are available. Overall, the preponderance of evidence indicates that P252H is most likely pathogenic, and this conclusion is not contradicted by ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.461924 | Structured | 0.211606 | Uncertain | 0.753 | 0.304 | 0.250 | -11.991 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 1.06 | Ambiguous | 0.2 | 1.96 | Ambiguous | 1.51 | Ambiguous | 0.82 | Ambiguous | 0.845 | Likely Pathogenic | -8.27 | Deleterious | 0.999 | Probably Damaging | 0.936 | Probably Damaging | 5.76 | Benign | 0.00 | Affected | 0.1592 | 0.4128 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.80C>A | P27H 2D AIThe SynGAP1 missense variant P27H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P27H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.517562 | Disordered | 0.437871 | Uncertain | 0.430 | 0.881 | 0.375 | -4.116 | Likely Benign | 0.177 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -2.46 | Neutral | 0.992 | Probably Damaging | 0.977 | Probably Damaging | 3.79 | Benign | 0.00 | Affected | 0.2384 | 0.5290 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.824C>A | P275H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P275H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a deleterious effect include SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default, all of which classify the change as pathogenic. Tools that are inconclusive or uncertain are Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta as uncertain. Taken together, the majority of evidence points to a pathogenic impact, and this conclusion is consistent with the lack of ClinVar annotation (no contradiction). Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.059222 | Structured | 0.353469 | Uncertain | 0.811 | 0.208 | 0.250 | -13.157 | Likely Pathogenic | 0.804 | Likely Pathogenic | Ambiguous | 2.34 | Destabilizing | 0.8 | 0.87 | Ambiguous | 1.61 | Ambiguous | 0.88 | Ambiguous | 0.585 | Likely Pathogenic | -6.54 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.73 | Pathogenic | 0.01 | Affected | 0.2069 | 0.3074 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.827C>A | P276H 2D 3DClick to see structure in 3D Viewer AISynGAP1 P276H is reported in gnomAD (ID 6‑33437732‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a deleterious effect: SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the variant as pathogenic, while only AlphaMissense‑Optimized predicts a benign outcome. Uncertain results are provided by FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as inconclusive. Overall, the preponderance of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.037156 | Structured | 0.338937 | Uncertain | 0.724 | 0.230 | 0.250 | 6-33437732-C-A | 1 | 6.20e-7 | -10.469 | Likely Pathogenic | 0.575 | Likely Pathogenic | Likely Benign | 1.98 | Ambiguous | 0.1 | 1.09 | Ambiguous | 1.54 | Ambiguous | 0.85 | Ambiguous | 0.534 | Likely Pathogenic | -4.80 | Deleterious | 1.000 | Probably Damaging | 0.961 | Probably Damaging | 1.84 | Pathogenic | 0.00 | Affected | 3.38 | 19 | 0.1739 | 0.3439 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||
| c.893C>A | P298H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P298H missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Remaining tools (AlphaMissense‑Default, FoldX, Rosetta, Foldetta, premPS) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta is uncertain. Overall, the majority of predictions (five pathogenic vs. three benign) and the SGM Consensus support a pathogenic interpretation. Therefore, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.328603 | Structured | 0.268765 | Uncertain | 0.860 | 0.283 | 0.500 | -9.777 | Likely Pathogenic | 0.443 | Ambiguous | Likely Benign | 1.57 | Ambiguous | 0.2 | 1.49 | Ambiguous | 1.53 | Ambiguous | 0.83 | Ambiguous | 0.313 | Likely Benign | -2.37 | Neutral | 0.999 | Probably Damaging | 0.964 | Probably Damaging | 1.92 | Pathogenic | 0.04 | Affected | 0.1769 | 0.4943 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||
| c.1027G>T | V343F 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V343F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions are made by REVEL and premPS, whereas pathogenic predictions are made by SIFT, polyPhen‑2 (HumDiv and HumVar), PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; Foldetta, which integrates FoldX‑MD and Rosetta outputs, is also uncertain. No evidence from FoldX or Rosetta alone is available. Overall, the majority of predictions support a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.291804 | Structured | 0.383911 | Uncertain | 0.882 | 0.497 | 0.250 | -10.709 | Likely Pathogenic | 0.799 | Likely Pathogenic | Ambiguous | 1.51 | Ambiguous | 0.4 | 1.28 | Ambiguous | 1.40 | Ambiguous | 0.23 | Likely Benign | 0.324 | Likely Benign | -3.37 | Deleterious | 0.976 | Probably Damaging | 0.759 | Possibly Damaging | 1.61 | Pathogenic | 0.01 | Affected | 0.0915 | 0.4552 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.103G>T | V35F 2D AIThe SynGAP1 missense variant V35F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; no Foldetta stability result is available. Overall, the majority of evidence points to a benign impact for V35F, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.613573 | Disordered | 0.434838 | Uncertain | 0.360 | 0.851 | 0.375 | -4.114 | Likely Benign | 0.122 | Likely Benign | Likely Benign | 0.133 | Likely Benign | -0.79 | Neutral | 0.923 | Possibly Damaging | 0.865 | Possibly Damaging | 4.14 | Benign | 0.00 | Affected | 0.0765 | 0.3421 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.1270G>T | V424F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant V424F is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, and FATHMM, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The remaining tools (FoldX, Foldetta, premPS, AlphaMissense‑Optimized) yield uncertain or unavailable results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.050641 | Structured | 0.411431 | Uncertain | 0.973 | 0.248 | 0.000 | -10.947 | Likely Pathogenic | 0.939 | Likely Pathogenic | Ambiguous | 1.99 | Ambiguous | 0.3 | -0.37 | Likely Benign | 0.81 | Ambiguous | 0.55 | Ambiguous | 0.275 | Likely Benign | -3.66 | Deleterious | 0.995 | Probably Damaging | 0.775 | Possibly Damaging | 3.40 | Benign | 0.01 | Affected | 0.0816 | 0.2094 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.1300G>T | V434F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V434F is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that indicate a benign effect include REVEL, premPS, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise SGM‑Consensus, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; SGM‑Consensus predicts pathogenic; Foldetta predicts pathogenic. All predictions are available and none are inconclusive. Based on the overall distribution of predictions, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.158265 | Structured | 0.342846 | Uncertain | 0.954 | 0.306 | 0.000 | -12.553 | Likely Pathogenic | 0.672 | Likely Pathogenic | Likely Benign | 3.64 | Destabilizing | 0.1 | 3.27 | Destabilizing | 3.46 | Destabilizing | 0.27 | Likely Benign | 0.417 | Likely Benign | -3.93 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 3.44 | Benign | 0.04 | Affected | 0.0596 | 0.3391 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.1321G>T | V441F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V441F missense variant is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic outcome are SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default; premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools lean toward a benign interpretation, and the high‑accuracy predictions are split but favor benign. Thus, the variant is most likely benign based on current computational evidence, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | -13.519 | Likely Pathogenic | 0.653 | Likely Pathogenic | Likely Benign | -0.26 | Likely Benign | 0.0 | 0.32 | Likely Benign | 0.03 | Likely Benign | 0.54 | Ambiguous | 0.355 | Likely Benign | -4.22 | Deleterious | 0.992 | Probably Damaging | 0.658 | Possibly Damaging | 3.37 | Benign | 0.08 | Tolerated | 0.0670 | 0.3269 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.1339G>T | V447F 2D 3DClick to see structure in 3D Viewer AISynGAP1 variant V447F is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that classify the variant as benign include REVEL, premPS, FATHMM, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized labeling the variant benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports an uncertain effect on protein folding. Overall, the majority of predictions lean toward pathogenicity, suggesting the variant is most likely pathogenic, a conclusion that does not conflict with the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.250310 | Structured | 0.283801 | Uncertain | 0.970 | 0.243 | 0.000 | Uncertain | 1 | -8.673 | Likely Pathogenic | 0.701 | Likely Pathogenic | Likely Benign | 1.40 | Ambiguous | 0.3 | 0.61 | Ambiguous | 1.01 | Ambiguous | 0.20 | Likely Benign | 0.206 | Likely Benign | -2.62 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 3.44 | Benign | 0.03 | Affected | 0.0551 | 0.3055 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||
| c.1354G>T | V452F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V452F variant is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect are Rosetta and FATHMM, whereas the remaining tools (REVEL, FoldX, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) all predict a pathogenic impact. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. Based on the preponderance of evidence, the variant is most likely pathogenic, a conclusion that contradicts its current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.335645 | Structured | 0.315167 | Uncertain | 0.970 | 0.229 | 0.000 | Uncertain | 1 | -14.769 | Likely Pathogenic | 0.975 | Likely Pathogenic | Likely Pathogenic | 9.21 | Destabilizing | 0.1 | 0.37 | Likely Benign | 4.79 | Destabilizing | 0.61 | Ambiguous | 0.511 | Likely Pathogenic | -4.94 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 3.29 | Benign | 0.00 | Affected | 3.37 | 34 | 0.0564 | 0.3451 | -1 | -1 | -1.4 | 48.04 | 249.4 | -35.7 | 0.0 | 0.0 | 0.4 | 0.1 | X | Potentially Pathogenic | The iso-propyl side chain of Val452, located in the middle of an α helix (res. Val441-Ser457), packs against hydrophobic residues in the inter-helix space at the intersection of three α helices (e.g., Leu500, His453, Leu465). In the variant simulations, the larger side chain of Phe452 cannot pack against the opposing α helix (res. Leu489-Glu519) as efficiently as valine. Due to space restrictions, the phenol ring adjusts to make room by rotating slightly sideways in the inter-helix space. Besides this small and local shift, no large-scale effects on the protein structure are seen based on the simulations. However, the size difference between the swapped residues could affect the protein folding process. | ||||||||||||||||
| c.1663G>T | V555F 2D 3DClick to see structure in 3D Viewer AISynGAP1 V555F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or stability result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.013265 | Structured | 0.008218 | Uncertain | 0.943 | 0.225 | 0.000 | -10.965 | Likely Pathogenic | 0.599 | Likely Pathogenic | Likely Benign | 0.17 | Likely Benign | 0.3 | -2.01 | Stabilizing | -0.92 | Ambiguous | 0.16 | Likely Benign | 0.440 | Likely Benign | -2.85 | Deleterious | 0.011 | Benign | 0.013 | Benign | -1.20 | Pathogenic | 0.16 | Tolerated | 0.0727 | 0.2366 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.1885G>T | V629F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V629F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining tools—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact; premPS is uncertain and is not counted as evidence. High‑accuracy methods further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta is pathogenic. No prediction is missing or inconclusive. Based on the overwhelming consensus of pathogenic predictions and the absence of any ClinVar annotation, the variant is most likely pathogenic, with no contradiction from ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.040537 | Structured | 0.034796 | Uncertain | 0.970 | 0.236 | 0.000 | -13.484 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 3.41 | Destabilizing | 0.5 | 4.40 | Destabilizing | 3.91 | Destabilizing | 0.64 | Ambiguous | 0.642 | Likely Pathogenic | -4.68 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.07 | Benign | 0.00 | Affected | 0.0560 | 0.2915 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.2509G>T | V837F 2D AIThe SynGAP1 missense variant V837F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.525368 | Disordered | 0.626284 | Binding | 0.318 | 0.871 | 0.125 | -5.223 | Likely Benign | 0.322 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -1.41 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.60 | Benign | 0.08 | Tolerated | 0.0749 | 0.3744 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.256G>T | V86F 2D AIThe SynGAP1 missense variant V86F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, while the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome; Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, and this conclusion does not contradict ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.552911 | Binding | 0.295 | 0.887 | 0.500 | -5.011 | Likely Benign | 0.981 | Likely Pathogenic | Likely Pathogenic | 0.126 | Likely Benign | -1.54 | Neutral | 0.824 | Possibly Damaging | 0.403 | Benign | 3.75 | Benign | 0.00 | Affected | 0.0755 | 0.4078 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.2578G>T | V860F 2D AIThe SynGAP1 missense variant V860F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy methods give a benign consensus: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.545602 | Disordered | 0.518121 | Binding | 0.269 | 0.803 | 0.250 | -4.576 | Likely Benign | 0.143 | Likely Benign | Likely Benign | 0.101 | Likely Benign | -2.25 | Neutral | 0.846 | Possibly Damaging | 0.522 | Possibly Damaging | 4.09 | Benign | 0.00 | Affected | 0.0717 | 0.4138 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.2731G>T | V911F 2D AIThe SynGAP1 missense variant V911F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for V911F, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.724137 | Binding | 0.327 | 0.914 | 0.375 | -4.454 | Likely Benign | 0.169 | Likely Benign | Likely Benign | 0.079 | Likely Benign | -1.08 | Neutral | 0.977 | Probably Damaging | 0.721 | Possibly Damaging | 2.68 | Benign | 0.04 | Affected | 0.0749 | 0.4380 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.2746G>T | V916F 2D AIThe SynGAP1 missense variant V916F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.835395 | Binding | 0.308 | 0.879 | 0.250 | -4.011 | Likely Benign | 0.123 | Likely Benign | Likely Benign | 0.121 | Likely Benign | -1.18 | Neutral | 0.642 | Possibly Damaging | 0.265 | Benign | 2.67 | Benign | 0.02 | Affected | 0.0729 | 0.4593 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.2974G>T | V992F 2D AIThe SynGAP1 missense variant V992F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and there is no conflict with ClinVar status, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.921728 | Binding | 0.331 | 0.917 | 0.750 | -3.131 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -1.25 | Neutral | 0.680 | Possibly Damaging | 0.356 | Benign | 4.17 | Benign | 0.04 | Affected | 0.0832 | 0.4083 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3232G>T | V1078F 2D AIThe SynGAP1 missense variant V1078F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect for V1078F, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.986989 | Binding | 0.294 | 0.898 | 0.750 | -3.768 | Likely Benign | 0.570 | Likely Pathogenic | Likely Benign | 0.142 | Likely Benign | -0.97 | Neutral | 0.977 | Probably Damaging | 0.722 | Possibly Damaging | 3.87 | Benign | 0.02 | Affected | 0.0677 | 0.4561 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.328G>T | V110F 2D AIThe SynGAP1 missense variant V110F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, SGM‑Consensus is Likely Benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.622677 | Disordered | 0.665934 | Binding | 0.347 | 0.860 | 0.750 | -4.872 | Likely Benign | 0.397 | Ambiguous | Likely Benign | 0.042 | Likely Benign | -1.63 | Neutral | 0.006 | Benign | 0.003 | Benign | 4.13 | Benign | 0.01 | Affected | 0.0794 | 0.4027 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3472G>T | V1158F 2D AIThe SynGAP1 missense variant V1158F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, while the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is classified as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. Foldetta results are unavailable. Overall, the majority of evidence indicates that V1158F is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar annotation exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.599170 | Disordered | 0.877504 | Binding | 0.369 | 0.847 | 0.250 | -3.888 | Likely Benign | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.264 | Likely Benign | -2.66 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.33 | Pathogenic | 0.02 | Affected | 0.0792 | 0.3577 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.916G>T | V306F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V306F is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL, premPS, and AlphaMissense‑Optimized, whereas the remaining tools (SGM‑Consensus, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) labeling it likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicting a pathogenic effect. Overall, the majority of evidence supports a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.363090 | Structured | 0.315026 | Uncertain | 0.896 | 0.287 | 0.125 | -9.311 | Likely Pathogenic | 0.700 | Likely Pathogenic | Likely Benign | 13.59 | Destabilizing | 1.4 | 6.99 | Destabilizing | 10.29 | Destabilizing | 0.23 | Likely Benign | 0.336 | Likely Benign | -3.28 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.74 | Pathogenic | 0.01 | Affected | 0.0535 | 0.3318 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.958G>T | V320F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant V320F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on benign impact include REVEL, premPS, and SIFT, whereas tools that agree on pathogenic impact include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as inconclusive, SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as inconclusive. Because the majority of available predictions and the SGM‑Consensus favor pathogenicity, the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for V320F. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.185198 | Structured | 0.419626 | Uncertain | 0.905 | 0.266 | 0.125 | -9.958 | Likely Pathogenic | 0.877 | Likely Pathogenic | Ambiguous | 1.49 | Ambiguous | 1.4 | 1.55 | Ambiguous | 1.52 | Ambiguous | 0.44 | Likely Benign | 0.237 | Likely Benign | -3.26 | Deleterious | 0.994 | Probably Damaging | 0.944 | Probably Damaging | 1.79 | Pathogenic | 0.06 | Tolerated | 0.0473 | 0.3064 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.1070A>G | H357R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 H357R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. Two tools remain uncertain: Rosetta and AlphaMissense‑Default. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority; and Foldetta also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -9.212 | Likely Pathogenic | 0.348 | Ambiguous | Likely Benign | -0.10 | Likely Benign | 0.3 | 1.07 | Ambiguous | 0.49 | Likely Benign | 0.32 | Likely Benign | 0.107 | Likely Benign | -1.03 | Neutral | 0.495 | Possibly Damaging | 0.095 | Benign | 4.22 | Benign | 0.59 | Tolerated | 0.2147 | 0.2818 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||
| c.107A>G | H36R 2D AIThe SynGAP1 missense variant H36R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.433974 | Uncertain | 0.334 | 0.834 | 0.375 | -2.513 | Likely Benign | 0.162 | Likely Benign | Likely Benign | 0.034 | Likely Benign | -0.70 | Neutral | 0.084 | Benign | 0.033 | Benign | 4.25 | Benign | 0.00 | Affected | 0.2417 | 0.3590 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.1280A>G | H427R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H427R is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438185‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT, while premPS and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign stability change. No predictions or folding results are missing or inconclusive. Overall, the majority of evidence indicates the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.394261 | Uncertain | 0.962 | 0.287 | 0.000 | 6-33438185-A-G | -3.259 | Likely Benign | 0.439 | Ambiguous | Likely Benign | -0.04 | Likely Benign | 0.1 | 0.48 | Likely Benign | 0.22 | Likely Benign | 0.72 | Ambiguous | 0.168 | Likely Benign | -2.61 | Deleterious | 0.174 | Benign | 0.018 | Benign | 3.51 | Benign | 0.03 | Affected | 3.38 | 25 | 0.2108 | 0.2121 | 0 | 2 | -1.3 | 19.05 | |||||||||||||||||||||||||||
| c.1358A>G | H453R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant H453R is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that indicate a benign effect include REVEL, Rosetta, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools predicting a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Uncertain results come from FoldX and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of standard predictors are split, but the two most reliable methods (AlphaMissense‑Optimized and Foldetta) favor a benign outcome, while the SGM Consensus suggests pathogenicity. Thus, the variant is most likely benign based on the available predictions, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.352862 | Structured | 0.316097 | Uncertain | 0.946 | 0.200 | 0.000 | Uncertain | 1 | -9.239 | Likely Pathogenic | 0.573 | Likely Pathogenic | Likely Benign | -0.52 | Ambiguous | 0.1 | 0.37 | Likely Benign | -0.08 | Likely Benign | 0.56 | Ambiguous | 0.396 | Likely Benign | -7.91 | Deleterious | 0.993 | Probably Damaging | 0.957 | Probably Damaging | 3.53 | Benign | 0.39 | Tolerated | 0.1646 | 0.2031 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||
| c.1442A>G | H481R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H481R is not listed in ClinVar and has no reported allele in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, SIFT, and FATHMM, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Uncertain results are reported by AlphaMissense‑Optimized, premPS, and Rosetta. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized is inconclusive; the SGM‑Consensus majority vote (AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely pathogenic effect; and Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, predicts a benign impact. Overall, the balance of evidence favors a pathogenic interpretation, and this conclusion is not contradicted by the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.257454 | Structured | 0.430977 | Uncertain | 0.764 | 0.247 | 0.000 | -11.753 | Likely Pathogenic | 0.823 | Likely Pathogenic | Ambiguous | -0.45 | Likely Benign | 0.1 | 0.68 | Ambiguous | 0.12 | Likely Benign | 0.59 | Ambiguous | 0.252 | Likely Benign | -4.48 | Deleterious | 0.983 | Probably Damaging | 0.977 | Probably Damaging | 3.47 | Benign | 0.53 | Tolerated | 0.1515 | 0.1690 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||
| c.1616A>G | H539R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H539R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from SIFT and the protein‑folding stability method Foldetta, whereas the remaining tools (SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict pathogenicity. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, while Foldetta (combining FoldX‑MD and Rosetta outputs) suggests a benign impact. Overall, the preponderance of evidence points to a pathogenic effect for H539R, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.031398 | Uncertain | 0.948 | 0.360 | 0.000 | -14.979 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | -0.83 | Ambiguous | 0.1 | 0.66 | Ambiguous | -0.09 | Likely Benign | 1.02 | Destabilizing | 0.832 | Likely Pathogenic | -7.19 | Deleterious | 0.997 | Probably Damaging | 0.989 | Probably Damaging | -1.16 | Pathogenic | 0.07 | Tolerated | 0.1497 | 0.1112 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||
| c.1673A>G | H558R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant H558R is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from AlphaMissense‑Optimized, Rosetta, SIFT, and polyPhen‑2 HumVar, while pathogenic predictions arise from REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and FATHMM. Four tools give inconclusive results: AlphaMissense‑Default, SGM‑Consensus, FoldX, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, remains uncertain. Overall, the majority of evidence points toward a pathogenic impact, which does not conflict with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.033407 | Structured | 0.011039 | Uncertain | 0.897 | 0.200 | 0.000 | Uncertain | 1 | -14.445 | Likely Pathogenic | 0.554 | Ambiguous | Likely Benign | -1.14 | Ambiguous | 0.1 | -0.23 | Likely Benign | -0.69 | Ambiguous | 1.03 | Destabilizing | 0.587 | Likely Pathogenic | -4.94 | Deleterious | 0.677 | Possibly Damaging | 0.239 | Benign | -1.24 | Pathogenic | 0.14 | Tolerated | 3.37 | 35 | 0.1500 | 0.1512 | 0 | 2 | -1.3 | 19.05 | |||||||||||||||||||||||||
| c.194A>G | H65R 2D AIThe SynGAP1 missense variant H65R is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33425802‑A‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, whereas the SGM‑Consensus remains benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | Uncertain | 1 | 6-33425802-A-G | 1 | 6.20e-7 | -1.980 | Likely Benign | 0.967 | Likely Pathogenic | Likely Pathogenic | 0.073 | Likely Benign | -1.60 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.19 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1483 | 0.1671 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||
| c.2078A>G | H693R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H693R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are limited to FATHMM, while the majority of algorithms (SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact; FoldX, Rosetta, and Foldetta are inconclusive. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely pathogenic effect, and Foldetta’s stability analysis is unavailable. Based on the preponderance of pathogenic predictions and the lack of benign evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.073402 | Structured | 0.323991 | Uncertain | 0.964 | 0.260 | 0.000 | -14.326 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 1.39 | Ambiguous | 0.2 | 1.28 | Ambiguous | 1.34 | Ambiguous | 1.03 | Destabilizing | 0.593 | Likely Pathogenic | -7.97 | Deleterious | 0.998 | Probably Damaging | 0.646 | Possibly Damaging | 3.13 | Benign | 0.01 | Affected | 0.1839 | 0.1670 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||
| c.2612A>G | H871R 2D AIThe SynGAP1 missense variant H871R is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. No pathogenic predictions are present among the evaluated tools. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are unavailable. Consequently, the variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.679301 | Binding | 0.279 | 0.858 | 0.250 | -3.894 | Likely Benign | 0.243 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -0.90 | Neutral | 0.144 | Benign | 0.085 | Benign | 2.67 | Benign | 0.23 | Tolerated | 0.1632 | 0.2099 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.26A>G | H9R 2D AIThe SynGAP1 missense variant H9R is reported in gnomAD (variant ID 6‑33420290‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that H9R is most likely benign, and this conclusion does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.528099 | Binding | 0.394 | 0.916 | 0.750 | 6-33420290-A-G | -1.736 | Likely Benign | 0.112 | Likely Benign | Likely Benign | 0.113 | Likely Benign | -0.04 | Neutral | 0.012 | Benign | 0.002 | Benign | 4.25 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2330 | 0.3266 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||
| c.2798A>G | H933R 2D AIThe SynGAP1 missense variant H933R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and AlphaMissense‑Optimized, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Overall, the majority of tools (five pathogenic vs. four benign) predict a pathogenic impact. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.987531 | Binding | 0.305 | 0.862 | 0.625 | -4.410 | Likely Benign | 0.650 | Likely Pathogenic | Likely Benign | 0.393 | Likely Benign | -3.84 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 2.43 | Pathogenic | 0.06 | Tolerated | 0.2074 | 0.2922 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.2834A>G | H945R 2D AIThe SynGAP1 missense variant H945R is reported in gnomAD (variant ID 6‑33443386‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict pathogenicity. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.982235 | Disordered | 0.849210 | Binding | 0.386 | 0.923 | 0.750 | 6-33443386-A-G | 1 | 6.20e-7 | -5.186 | Likely Benign | 0.118 | Likely Benign | Likely Benign | 0.333 | Likely Benign | -0.59 | Neutral | 0.982 | Probably Damaging | 0.903 | Possibly Damaging | 5.05 | Benign | 0.08 | Tolerated | 4.32 | 4 | 0.2764 | 0.3020 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.284A>G | H95R 2D AIThe SynGAP1 missense variant H95R is reported in gnomAD (variant ID 6‑33425892‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that H95R is most likely benign, and this conclusion does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.590542 | Binding | 0.335 | 0.875 | 0.625 | 6-33425892-A-G | 1 | 6.20e-7 | -2.789 | Likely Benign | 0.153 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -1.31 | Neutral | 0.084 | Benign | 0.009 | Benign | 4.20 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1907 | 0.2087 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.2852A>G | H951R 2D AIThe SynGAP1 missense variant H951R is listed in ClinVar as Pathogenic (ClinVar ID 1003739.0) and is not reported in gnomAD. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. The Foldetta protein‑folding stability analysis is unavailable for this variant. Based on the collective predictions, the variant is most likely benign, which contradicts its ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.901477 | Binding | 0.415 | 0.925 | 0.750 | Likely Pathogenic | 1 | -4.964 | Likely Benign | 0.125 | Likely Benign | Likely Benign | 0.185 | Likely Benign | -1.08 | Neutral | 0.048 | Benign | 0.029 | Benign | 5.46 | Benign | 0.24 | Tolerated | 3.77 | 5 | 0.2808 | 0.3220 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||
| c.2870A>G | H957R 2D AIThe SynGAP1 missense variant H957R is catalogued in gnomAD (ID 6‑33443422‑A‑G) but has no ClinVar entry. Functional prediction tools uniformly classify it as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes. No tool predicts pathogenicity. Grouping by consensus, all listed predictors fall into the benign category, with no opposing pathogenic calls. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. Foldetta results are unavailable, so they do not influence the assessment. Overall, the evidence strongly supports a benign classification for H957R, and this conclusion does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.968874 | Binding | 0.362 | 0.915 | 0.750 | 6-33443422-A-G | 1 | 6.20e-7 | -6.723 | Likely Benign | 0.183 | Likely Benign | Likely Benign | 0.105 | Likely Benign | -1.31 | Neutral | 0.144 | Benign | 0.078 | Benign | 2.58 | Benign | 0.32 | Tolerated | 3.77 | 5 | 0.2410 | 0.3610 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.2873A>G | H958R 2D AIThe SynGAP1 missense variant H958R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.976011 | Binding | 0.371 | 0.913 | 0.750 | -9.188 | Likely Pathogenic | 0.169 | Likely Benign | Likely Benign | 0.134 | Likely Benign | -1.29 | Neutral | 0.836 | Possibly Damaging | 0.232 | Benign | 4.17 | Benign | 0.09 | Tolerated | 0.2440 | 0.3410 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.2876A>G | H959R 2D AIThe SynGAP1 missense variant H959R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only ESM1b predicts a pathogenic outcome, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates likely benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.980566 | Binding | 0.333 | 0.905 | 0.750 | -9.459 | Likely Pathogenic | 0.182 | Likely Benign | Likely Benign | 0.162 | Likely Benign | -1.11 | Neutral | 0.144 | Benign | 0.078 | Benign | 4.14 | Benign | 0.15 | Tolerated | 0.2416 | 0.3610 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.2879A>G | H960R 2D AIThe SynGAP1 missense variant H960R is reported in gnomAD (ID 6‑33443431‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only two tools predict pathogenicity—polyPhen‑2 HumDiv and ESM1b—while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also reports likely benign. No Foldetta stability data are available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987911 | Disordered | 0.983385 | Binding | 0.380 | 0.901 | 0.750 | 6-33443431-A-G | 1 | 6.20e-7 | -9.238 | Likely Pathogenic | 0.192 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -1.10 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.19 | Benign | 0.25 | Tolerated | 3.77 | 5 | 0.2237 | 0.3667 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.2882A>G | H961R 2D AIThe SynGAP1 missense variant H961R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign (3 benign vs. 1 pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.989835 | Disordered | 0.984562 | Binding | 0.323 | 0.893 | 0.750 | -9.258 | Likely Pathogenic | 0.189 | Likely Benign | Likely Benign | 0.101 | Likely Benign | -0.90 | Neutral | 0.144 | Benign | 0.078 | Benign | 4.16 | Benign | 0.02 | Affected | 0.2201 | 0.3267 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.2885A>G | H962R 2D AIThe SynGAP1 missense variant H962R is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” status. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of any ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.984483 | Binding | 0.369 | 0.886 | 0.750 | -9.166 | Likely Pathogenic | 0.212 | Likely Benign | Likely Benign | 0.117 | Likely Benign | -1.04 | Neutral | 0.325 | Benign | 0.129 | Benign | 4.18 | Benign | 0.07 | Tolerated | 0.2276 | 0.3337 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.2888A>G | H963R 2D AIThe SynGAP1 missense variant H963R is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443440‑A‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Only ESM1b predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized reports benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, which does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.983973 | Binding | 0.325 | 0.886 | 0.750 | Uncertain | 1 | 6-33443440-A-G | 8 | 4.96e-6 | -8.952 | Likely Pathogenic | 0.169 | Likely Benign | Likely Benign | 0.081 | Likely Benign | -1.28 | Neutral | 0.001 | Benign | 0.003 | Benign | 4.15 | Benign | 0.24 | Tolerated | 3.77 | 5 | 0.2330 | 0.3380 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||
| c.2891A>G | H964R 2D AIThe SynGAP1 missense variant H964R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign (3 benign vs. 1 pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.990547 | Disordered | 0.982486 | Binding | 0.364 | 0.886 | 0.750 | -8.275 | Likely Pathogenic | 0.163 | Likely Benign | Likely Benign | 0.129 | Likely Benign | -0.68 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.19 | Benign | 0.05 | Affected | 0.2237 | 0.3467 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.2894A>G | H965R 2D AIThe SynGAP1 missense variant H965R is catalogued in gnomAD (ID 6‑33443446‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign or likely benign. No tool predicts pathogenicity; ESM1b is uncertain but does not contradict the benign consensus. High‑accuracy assessments confirm this view: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so it does not influence the assessment. Overall, the available predictions strongly suggest that H965R is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988505 | Disordered | 0.978700 | Binding | 0.342 | 0.882 | 0.750 | 6-33443446-A-G | 1 | 6.20e-7 | -7.056 | In-Between | 0.156 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -0.88 | Neutral | 0.065 | Benign | 0.049 | Benign | 4.08 | Benign | 0.38 | Tolerated | 3.77 | 5 | 0.2394 | 0.3610 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.2897A>G | H966R 2D AIThe SynGAP1 missense variant H966R is reported in gnomAD (ID 6‑33443449‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant call. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that H966R is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.976962 | Disordered | 0.974672 | Binding | 0.378 | 0.879 | 0.750 | 6-33443449-A-G | -5.474 | Likely Benign | 0.157 | Likely Benign | Likely Benign | 0.172 | Likely Benign | -0.71 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.06 | Benign | 0.69 | Tolerated | 4.32 | 2 | 0.2198 | 0.3410 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||
| c.2939A>G | H980R 2D AIThe SynGAP1 missense variant H980R is listed in gnomAD (ID 6‑33443491‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.974598 | Binding | 0.309 | 0.892 | 0.625 | 6-33443491-A-G | 1 | 6.20e-7 | -2.736 | Likely Benign | 0.409 | Ambiguous | Likely Benign | 0.095 | Likely Benign | -1.44 | Neutral | 0.802 | Possibly Damaging | 0.354 | Benign | 4.17 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2439 | 0.3810 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.293A>G | H98R 2D AIThe SynGAP1 missense variant H98R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.631713 | Binding | 0.348 | 0.872 | 0.625 | -2.772 | Likely Benign | 0.180 | Likely Benign | Likely Benign | 0.116 | Likely Benign | -0.39 | Neutral | 0.115 | Benign | 0.006 | Benign | 4.27 | Benign | 0.00 | Affected | 0.2271 | 0.3022 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.3005A>G | H1002R 2D AIThe SynGAP1 missense variant H1002R is listed in gnomAD (ID 6‑33443557‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Taken together, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar classification (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.953758 | Binding | 0.285 | 0.900 | 0.500 | 6-33443557-A-G | 1 | 6.20e-7 | -3.624 | Likely Benign | 0.609 | Likely Pathogenic | Likely Benign | 0.082 | Likely Benign | -1.52 | Neutral | 0.012 | Benign | 0.022 | Benign | 2.76 | Benign | 0.25 | Tolerated | 4.32 | 4 | 0.2160 | 0.2978 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.3011A>G | H1004R 2D AIThe SynGAP1 missense variant H1004R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Benign, and no Foldetta stability data is available. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.808535 | Disordered | 0.943707 | Binding | 0.271 | 0.901 | 0.750 | -3.316 | Likely Benign | 0.813 | Likely Pathogenic | Ambiguous | 0.198 | Likely Benign | -1.88 | Neutral | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 2.77 | Benign | 0.50 | Tolerated | 0.2097 | 0.3132 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.302A>G | H101R 2D AIThe SynGAP1 missense variant H101R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for H101R, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.791621 | Disordered | 0.688356 | Binding | 0.370 | 0.884 | 0.625 | -2.685 | Likely Benign | 0.211 | Likely Benign | Likely Benign | 0.141 | Likely Benign | -0.76 | Neutral | 0.824 | Possibly Damaging | 0.840 | Possibly Damaging | 4.21 | Benign | 0.00 | Affected | 0.1891 | 0.2225 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.3089A>G | H1030R 2D AIThe SynGAP1 missense variant H1030R is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. The high‑accuracy consensus methods likewise support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.849326 | Disordered | 0.995856 | Binding | 0.375 | 0.735 | 0.500 | -2.312 | Likely Benign | 0.340 | Likely Benign | Likely Benign | 0.031 | Likely Benign | -1.08 | Neutral | 0.224 | Benign | 0.066 | Benign | 2.85 | Benign | 0.06 | Tolerated | 0.1872 | 0.2955 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.3410A>G | H1137R 2D AIThe SynGAP1 missense variant H1137R is reported in gnomAD (ID 6‑33444445‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | 6-33444445-A-G | 1 | 6.20e-7 | -3.468 | Likely Benign | 0.228 | Likely Benign | Likely Benign | 0.296 | Likely Benign | -1.19 | Neutral | 0.925 | Possibly Damaging | 0.629 | Possibly Damaging | 5.32 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2131 | 0.3449 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.3488A>G | H1163R 2D AIThe SynGAP1 missense variant H1163R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and no Foldetta stability data is available. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.509769 | Disordered | 0.858469 | Binding | 0.328 | 0.825 | 0.375 | -2.505 | Likely Benign | 0.942 | Likely Pathogenic | Ambiguous | 0.472 | Likely Benign | -2.08 | Neutral | 0.991 | Probably Damaging | 0.991 | Probably Damaging | 5.43 | Benign | 0.27 | Tolerated | 0.1863 | 0.2402 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.3515A>G | H1172R 2D AIThe SynGAP1 missense variant H1172R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that H1172R is most likely benign, and this conclusion does not contradict any ClinVar status, as none is assigned to the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.673805 | Binding | 0.465 | 0.758 | 0.625 | -0.848 | Likely Benign | 0.285 | Likely Benign | Likely Benign | 0.173 | Likely Benign | -1.08 | Neutral | 0.001 | Benign | 0.004 | Benign | 5.46 | Benign | 0.05 | Affected | 0.1547 | 0.2219 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||||
| c.3608A>G | H1203R 2D AIThe SynGAP1 missense variant H1203R is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.618285 | Disordered | 0.527023 | Binding | 0.892 | 0.589 | 0.250 | -3.355 | Likely Benign | 0.204 | Likely Benign | Likely Benign | 0.287 | Likely Benign | -1.61 | Neutral | 0.473 | Possibly Damaging | 0.265 | Benign | 5.51 | Benign | 0.20 | Tolerated | 0.1263 | 0.1178 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||||
| c.3629A>G | H1210R 2D AIThe SynGAP1 missense variant H1210R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools uniformly classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity, so the pathogenic‑prediction group is empty. High‑accuracy assessments corroborate this benign consensus: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields “Likely Benign.” Foldetta results are not available, so they do not influence the interpretation. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.587579 | Binding | 0.900 | 0.567 | 0.375 | 0.860 | Likely Benign | 0.173 | Likely Benign | Likely Benign | 0.088 | Likely Benign | 0.89 | Neutral | 0.178 | Benign | 0.089 | Benign | 3.25 | Benign | 1.00 | Tolerated | 0.1535 | 0.2384 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||||
| c.3830A>G | H1277R 2D AIThe SynGAP1 missense variant H1277R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | -2.940 | Likely Benign | 0.306 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -5.60 | Deleterious | 0.259 | Benign | 0.066 | Benign | 2.15 | Pathogenic | 0.00 | Affected | 0.1710 | 0.1562 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||||||
| c.4028A>G | H1343R 2D AIThe SynGAP1 missense variant H1343R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -2.179 | Likely Benign | 0.105 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -1.16 | Neutral | 0.659 | Possibly Damaging | 0.071 | Benign | 4.08 | Benign | 0.00 | Affected | 0.2301 | 0.2602 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.503A>G | H168R 2D AIThe SynGAP1 missense variant H168R is reported in gnomAD (ID 6‑33432800‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized all classify the change as benign, whereas SIFT predicts it to be pathogenic. Two tools (ESM1b and AlphaMissense‑Default) return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it contains two benign and two uncertain calls, and Foldetta data are not available. Overall, the preponderance of evidence points to a benign impact. This conclusion does not contradict ClinVar, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.433034 | Structured | 0.502450 | Binding | 0.402 | 0.678 | 0.125 | 6-33432800-A-G | 1 | 6.20e-7 | -7.334 | In-Between | 0.395 | Ambiguous | Likely Benign | 0.153 | Likely Benign | -1.08 | Neutral | 0.016 | Benign | 0.011 | Benign | 4.26 | Benign | 0.02 | Affected | 4.32 | 3 | 0.1752 | 0.2499 | 0 | 2 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||
| c.542A>G | H181R 2D AIThe SynGAP1 missense variant H181R is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.505461 | Disordered | 0.439530 | Uncertain | 0.294 | 0.616 | 0.500 | -3.668 | Likely Benign | 0.521 | Ambiguous | Likely Benign | 0.174 | Likely Benign | -0.11 | Neutral | 0.818 | Possibly Damaging | 0.188 | Benign | 4.35 | Benign | 1.00 | Tolerated | 0.1585 | 0.1782 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.548A>G | H183R 2D AIThe SynGAP1 H183R missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, SIFT, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus (majority vote) also pathogenic; Foldetta stability analysis is unavailable. Based on the majority of computational evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.432952 | Uncertain | 0.421 | 0.622 | 0.500 | -10.937 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.368 | Likely Benign | -5.43 | Deleterious | 0.596 | Possibly Damaging | 0.142 | Benign | 3.90 | Benign | 0.13 | Tolerated | 0.1915 | 0.2249 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.629A>G | H210R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H210R missense variant is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FATHMM, and FoldX, whereas a majority of tools predict a pathogenic impact: premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). The high‑accuracy AlphaMissense‑Optimized result is pathogenic; the SGM Consensus, based on the same set of predictors, is also pathogenic; Foldetta’s stability assessment is uncertain and therefore not considered evidence. Overall, the balance of evidence points to a pathogenic effect for H210R. This prediction does not contradict the ClinVar “Uncertain” classification, which remains unresolved. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.144935 | Structured | 0.390904 | Uncertain | 0.872 | 0.298 | 0.125 | Uncertain | 1 | -14.254 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.40 | Likely Benign | 0.4 | 3.05 | Destabilizing | 1.73 | Ambiguous | 1.12 | Destabilizing | 0.431 | Likely Benign | -6.74 | Deleterious | 0.808 | Possibly Damaging | 0.452 | Possibly Damaging | 3.09 | Benign | 0.00 | Affected | 0.1427 | 0.1982 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||
| c.89A>G | H30R 2D AIThe SynGAP1 missense variant H30R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | -2.156 | Likely Benign | 0.186 | Likely Benign | Likely Benign | 0.138 | Likely Benign | -2.17 | Neutral | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.94 | Benign | 0.00 | Affected | 0.2762 | 0.3590 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.932A>G | H311R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H311R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, SIFT, and AlphaMissense‑Optimized, whereas a majority of tools (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic outcome. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and premPS. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized indicates benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates pathogenic, and Foldetta remains uncertain. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.229226 | Structured | 0.354792 | Uncertain | 0.902 | 0.314 | 0.125 | -9.825 | Likely Pathogenic | 0.719 | Likely Pathogenic | Likely Benign | 0.43 | Likely Benign | 0.1 | 0.85 | Ambiguous | 0.64 | Ambiguous | 0.70 | Ambiguous | 0.532 | Likely Pathogenic | -5.72 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.94 | Pathogenic | 0.14 | Tolerated | 0.1931 | 0.2490 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||
| c.977A>G | H326R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H326R missense variant is not listed in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include SIFT and Foldetta, whereas the remaining tools (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict a pathogenic impact. High‑accuracy methods give the following results: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports it as likely pathogenic; Foldetta, a protein‑folding stability approach that integrates FoldX‑MD and Rosetta outputs, predicts a benign effect. Because the majority of evidence points to a deleterious change, the variant is most likely pathogenic, and this assessment is not contradicted by the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.342579 | Structured | 0.418150 | Uncertain | 0.944 | 0.455 | 0.000 | -12.369 | Likely Pathogenic | 0.980 | Likely Pathogenic | Likely Pathogenic | -0.65 | Ambiguous | 0.1 | 1.40 | Ambiguous | 0.38 | Likely Benign | 0.83 | Ambiguous | 0.601 | Likely Pathogenic | -6.89 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.97 | Pathogenic | 0.10 | Tolerated | 0.1832 | 0.2413 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||
| c.1082A>G | Q361R 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q361R has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, SIFT, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Rosetta; pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign; the SGM‑Consensus itself is benign; and Foldetta, which integrates FoldX‑MD (Uncertain) and Rosetta (Benign), yields a benign prediction. Overall, the consensus of available computational evidence indicates that the variant is most likely benign, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.305330 | Structured | 0.427593 | Uncertain | 0.945 | 0.534 | 0.250 | -6.898 | Likely Benign | 0.289 | Likely Benign | Likely Benign | -0.88 | Ambiguous | 0.0 | -0.07 | Likely Benign | -0.48 | Likely Benign | -0.20 | Likely Benign | 0.296 | Likely Benign | -0.56 | Neutral | 0.987 | Probably Damaging | 0.953 | Probably Damaging | 1.71 | Pathogenic | 0.41 | Tolerated | 0.1321 | 0.2955 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1220A>G | Q407R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q407R has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default; premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. With two of the three high‑accuracy methods indicating benign and the remaining one pathogenic, the overall evidence leans toward a benign classification. This conclusion does not contradict ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.109221 | Structured | 0.382522 | Uncertain | 0.916 | 0.271 | 0.000 | -13.263 | Likely Pathogenic | 0.693 | Likely Pathogenic | Likely Benign | 0.15 | Likely Benign | 0.2 | 0.09 | Likely Benign | 0.12 | Likely Benign | 0.65 | Ambiguous | 0.340 | Likely Benign | -3.52 | Deleterious | 0.909 | Possibly Damaging | 0.889 | Possibly Damaging | 4.02 | Benign | 0.17 | Tolerated | 0.1347 | 0.1596 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1236G>C | L412F 2D  AIThe SynGAP1 missense variant L412F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, and FATHMM, whereas the majority of tools (SGM‑Consensus, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. Taken together, the consensus of the majority of evidence points to a pathogenic effect for L412F, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.122885 | Structured | 0.331108 | Uncertain | 0.937 | 0.196 | 0.000 | -8.710 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 7.00 | Destabilizing | 3.5 | 3.61 | Destabilizing | 5.31 | Destabilizing | 0.48 | Likely Benign | 0.286 | Likely Benign | -3.69 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | 3.27 | Benign | 0.04 | Affected | 0.0602 | 0.2891 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1236G>T | L412F 2D AIThe SynGAP1 missense variant L412F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, and FATHMM, whereas the majority of tools (SGM‑Consensus, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. Taken together, the consensus of the majority of evidence points to a pathogenic effect for L412F, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.122885 | Structured | 0.331108 | Uncertain | 0.937 | 0.196 | 0.000 | -8.710 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 7.00 | Destabilizing | 3.5 | 3.61 | Destabilizing | 5.31 | Destabilizing | 0.48 | Likely Benign | 0.283 | Likely Benign | -3.69 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | 3.27 | Benign | 0.04 | Affected | 0.0602 | 0.2891 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1305G>C | L435F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L435F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas a majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM Consensus (majority vote). High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also uncertain, providing no definitive evidence for either outcome. Given the preponderance of pathogenic predictions and the lack of any ClinVar annotation to contradict this assessment, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.229226 | Structured | 0.333584 | Uncertain | 0.954 | 0.292 | 0.000 | -11.871 | Likely Pathogenic | 0.932 | Likely Pathogenic | Ambiguous | 0.51 | Ambiguous | 0.1 | 0.95 | Ambiguous | 0.73 | Ambiguous | 0.69 | Ambiguous | 0.222 | Likely Benign | -3.75 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | 3.26 | Benign | 0.01 | Affected | 0.0587 | 0.2723 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1305G>T | L435F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L435F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas a majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM Consensus (Likely Pathogenic). High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also uncertain, providing no definitive evidence for either outcome. Given the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.229226 | Structured | 0.333584 | Uncertain | 0.954 | 0.292 | 0.000 | -11.871 | Likely Pathogenic | 0.932 | Likely Pathogenic | Ambiguous | 0.51 | Ambiguous | 0.1 | 0.95 | Ambiguous | 0.73 | Ambiguous | 0.69 | Ambiguous | 0.222 | Likely Benign | -3.75 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | 3.26 | Benign | 0.01 | Affected | 0.0587 | 0.2723 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1367A>G | Q456R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q456R has no ClinVar entry and is not reported in gnomAD. Prediction tools that classify it as benign include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, and FATHMM. Those that predict pathogenicity are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain; the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic, whereas Foldetta (a folding‑stability method combining FoldX‑MD and Rosetta outputs) predicts benign. No evidence from the data contradicts ClinVar status, which is currently unclassified. Based on the available predictions, the variant is most likely benign, though the evidence is conflicting and does not conflict with the lack of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.170161 | Structured | 0.302348 | Uncertain | 0.939 | 0.164 | 0.000 | -11.326 | Likely Pathogenic | 0.801 | Likely Pathogenic | Ambiguous | 0.21 | Likely Benign | 0.1 | 0.12 | Likely Benign | 0.17 | Likely Benign | 0.33 | Likely Benign | 0.295 | Likely Benign | -3.69 | Deleterious | 0.980 | Probably Damaging | 0.943 | Probably Damaging | 3.37 | Benign | 0.20 | Tolerated | 0.1241 | 0.1124 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1393C>T | L465F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L465F is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL and Rosetta, whereas the majority of tools predict a pathogenic impact: FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Two tools (Foldetta and premPS) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus also pathogenic, and Foldetta remains uncertain. Overall, the consensus of high‑confidence predictors points to a pathogenic effect for L465F. This conclusion is not contradicted by ClinVar status, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.346032 | Structured | 0.319240 | Uncertain | 0.956 | 0.202 | 0.000 | -12.626 | Likely Pathogenic | 0.979 | Likely Pathogenic | Likely Pathogenic | 3.11 | Destabilizing | 0.7 | 0.05 | Likely Benign | 1.58 | Ambiguous | 0.52 | Ambiguous | 0.432 | Likely Benign | -3.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.44 | Pathogenic | 0.01 | Affected | 0.0771 | 0.2759 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1444C>T | L482F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L482F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from FoldX and premPS, while pathogenic calls are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain results are reported by Rosetta, Foldetta, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further reveal AlphaMissense‑Optimized as Uncertain, SGM Consensus as Likely Pathogenic, and Foldetta as Uncertain. Overall, the preponderance of evidence points to a pathogenic effect for L482F. This conclusion is consistent with the absence of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.254060 | Structured | 0.426236 | Uncertain | 0.795 | 0.248 | 0.000 | -11.257 | Likely Pathogenic | 0.951 | Likely Pathogenic | Ambiguous | 0.48 | Likely Benign | 0.0 | 1.09 | Ambiguous | 0.79 | Ambiguous | 0.43 | Likely Benign | 0.724 | Likely Pathogenic | -3.95 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | -1.22 | Pathogenic | 0.01 | Affected | 0.0457 | 0.1814 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1465C>T | L489F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L489F is listed in ClinVar with an uncertain significance (ClinVar ID 522018.0) and is present in the gnomAD database (gnomAD ID 6‑33438497‑C‑T). In silico prediction tools that assess pathogenicity all converge on a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report a pathogenic outcome, while no tool predicts a benign effect. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is inconclusive. No prediction or folding‑stability result is missing or ambiguous. **Thus, the variant is most likely pathogenic based on the collective predictions, and this does not contradict the ClinVar uncertain status.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.326126 | Uncertain | 0.949 | 0.234 | 0.125 | Uncertain | 2 | 6-33438497-C-T | 1 | 6.20e-7 | -12.066 | Likely Pathogenic | 0.965 | Likely Pathogenic | Likely Pathogenic | 1.72 | Ambiguous | 0.5 | 1.14 | Ambiguous | 1.43 | Ambiguous | 0.56 | Ambiguous | 0.724 | Likely Pathogenic | -3.76 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.51 | Pathogenic | 0.01 | Affected | 3.37 | 35 | 0.0791 | 0.3729 | 2 | 0 | -1.0 | 34.02 | 246.4 | -17.8 | 0.0 | 0.0 | 0.6 | 0.1 | X | Potentially Benign | The iso-butyl side chain of Leu489, located in the α-helix (res. Leu489-Glu519) within an inter-helix space of four helices (res. Ala461-Phe476, res. Val441-Ser457, and res. Met414-Glu436), packs with hydrophobic residues (e.g., Cys432, Ala448, Lys444, Ala493, Val447, Met468) in the inter-helix space. In the variant simulations, the phenyl ring of the Phe489 side chain can also pack favorably in the hydrophobic region. However, due to the size difference, the aromatic side chain of Phe489 tends to reposition to escape the tight region to accommodate the larger side chain, stacking with Lys444. Although no apparent negative changes are observed during the variant simulation, the size difference between the swapped residues could affect the protein folding process. | |||||||||||||
| c.1508A>G | Q503R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q503R has no ClinVar entry and is not reported in gnomAD. Prediction tools that classify the variant as benign include FoldX, Foldetta, premPS, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and FATHMM. The high‑accuracy methods give a benign result for AlphaMissense‑Optimized, a pathogenic result for the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and a benign result for Foldetta (combining FoldX‑MD and Rosetta). Overall, the majority of tools and the high‑accuracy methods lean toward a benign effect. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.040537 | Structured | 0.322935 | Uncertain | 0.848 | 0.168 | 0.000 | -11.396 | Likely Pathogenic | 0.232 | Likely Benign | Likely Benign | -0.40 | Likely Benign | 0.3 | 0.60 | Ambiguous | 0.10 | Likely Benign | 0.50 | Likely Benign | 0.640 | Likely Pathogenic | -3.34 | Deleterious | 0.577 | Possibly Damaging | 0.395 | Benign | -1.42 | Pathogenic | 0.06 | Tolerated | 0.1426 | 0.0963 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1516C>T | L506F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L506F is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the majority of tools (FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic impact; premPS and AlphaMissense‑Optimized are inconclusive. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is uncertain, the SGM Consensus (derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, which is consistent with its ClinVar “Uncertain” classification and does not contradict the available data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.034884 | Structured | 0.279180 | Uncertain | 0.924 | 0.196 | 0.000 | Uncertain | 1 | -11.262 | Likely Pathogenic | 0.883 | Likely Pathogenic | Ambiguous | 4.92 | Destabilizing | 0.8 | 5.76 | Destabilizing | 5.34 | Destabilizing | 0.91 | Ambiguous | 0.464 | Likely Benign | -3.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.62 | Pathogenic | 0.01 | Affected | 3.37 | 35 | 0.0566 | 0.1471 | 0 | 2 | -1.0 | 34.02 | |||||||||||||||||||||||||
| c.1608G>C | L536F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L536F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only Rosetta. Those that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus. Predictions that are uncertain or inconclusive are FoldX, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of reliable tools predict a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.137348 | Structured | 0.042188 | Uncertain | 0.931 | 0.341 | 0.000 | -10.316 | Likely Pathogenic | 0.851 | Likely Pathogenic | Ambiguous | 1.31 | Ambiguous | 0.5 | 0.40 | Likely Benign | 0.86 | Ambiguous | 0.70 | Ambiguous | 0.724 | Likely Pathogenic | -3.90 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.36 | Pathogenic | 0.01 | Affected | 0.0874 | 0.2804 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1608G>T | L536F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L536F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only Rosetta. Those that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus. Predictions that are uncertain or inconclusive are FoldX, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of reliable tools predict a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.137348 | Structured | 0.042188 | Uncertain | 0.931 | 0.341 | 0.000 | -10.316 | Likely Pathogenic | 0.851 | Likely Pathogenic | Ambiguous | 1.31 | Ambiguous | 0.5 | 0.40 | Likely Benign | 0.86 | Ambiguous | 0.70 | Ambiguous | 0.722 | Likely Pathogenic | -3.90 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.36 | Pathogenic | 0.01 | Affected | 0.0874 | 0.2804 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1619A>G | Q540R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q540R has no ClinVar entry and is present in gnomAD (ID 6‑33438862‑A‑G). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Two tools report uncertainty: premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) as benign. Overall, the majority of evidence points to a benign impact, with only a minority of tools indicating pathogenicity. This conclusion does not contradict ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.085092 | Structured | 0.029522 | Uncertain | 0.958 | 0.371 | 0.000 | 6-33438862-A-G | 1 | 6.19e-7 | -13.312 | Likely Pathogenic | 0.540 | Ambiguous | Likely Benign | -0.06 | Likely Benign | 0.0 | -0.07 | Likely Benign | -0.07 | Likely Benign | 0.88 | Ambiguous | 0.795 | Likely Pathogenic | -3.98 | Deleterious | 0.991 | Probably Damaging | 0.985 | Probably Damaging | -1.28 | Pathogenic | 0.08 | Tolerated | 3.37 | 35 | 0.1328 | 0.1886 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||
| c.1647G>C | L549F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L549F is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include Rosetta, premPS, and SIFT, whereas a majority of tools predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. FoldX and Foldetta give uncertain results. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely pathogenic, and Foldetta remains uncertain. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, which is consistent with its ClinVar “Uncertain” classification and does not contradict the available data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.007921 | Uncertain | 0.955 | 0.281 | 0.000 | Uncertain | 1 | -11.634 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.80 | Ambiguous | 0.3 | 0.27 | Likely Benign | 0.54 | Ambiguous | 0.49 | Likely Benign | 0.514 | Likely Pathogenic | -3.42 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.29 | Pathogenic | 0.14 | Tolerated | 0.0724 | 0.1859 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||
| c.1647G>T | L549F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L549F is not reported in ClinVar and is absent from gnomAD. Benign predictions come from Rosetta, premPS, and SIFT, whereas pathogenic predictions are made by REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; the SGM‑Consensus score indicates likely pathogenic, while FoldX and Foldetta are uncertain. High‑accuracy tools specifically: AlphaMissense‑Optimized predicts pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. Overall, the majority of evidence points to a pathogenic effect, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.007921 | Uncertain | 0.955 | 0.281 | 0.000 | -11.634 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.80 | Ambiguous | 0.3 | 0.27 | Likely Benign | 0.54 | Ambiguous | 0.49 | Likely Benign | 0.514 | Likely Pathogenic | -3.42 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.29 | Pathogenic | 0.14 | Tolerated | 0.0724 | 0.1859 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.164A>G | Q55R 2D AIThe SynGAP1 missense variant Q55R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.332115 | Structured | 0.470108 | Uncertain | 0.461 | 0.657 | 0.000 | -6.626 | Likely Benign | 0.578 | Likely Pathogenic | Likely Benign | 0.166 | Likely Benign | -1.15 | Neutral | 0.140 | Benign | 0.275 | Benign | 3.87 | Benign | 0.00 | Affected | 0.1607 | 0.1558 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.169C>T | L57F 2D AIThe SynGAP1 missense variant L57F (ClinVar ID 1973575.0) is listed as “Uncertain” in ClinVar and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion aligns with the ClinVar “Uncertain” status, as it does not contradict the current classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.254060 | Structured | 0.481044 | Uncertain | 0.554 | 0.642 | 0.000 | Uncertain | 2 | -5.096 | Likely Benign | 0.459 | Ambiguous | Likely Benign | 0.051 | Likely Benign | -0.78 | Neutral | 0.824 | Possibly Damaging | 0.879 | Possibly Damaging | 3.96 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0679 | 0.3355 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||
| c.1762C>T | L588F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L588F is reported in gnomAD (variant ID 6‑33440814‑C‑T) but has no ClinVar entry. Across the available in‑silico predictors, every tool examined classifies the substitution as pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return pathogenic or likely pathogenic scores. No tool predicts a benign effect. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is pathogenic. Because every available prediction converges on a deleterious effect and there is no ClinVar annotation to contradict this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.038042 | Structured | 0.082229 | Uncertain | 0.887 | 0.214 | 0.000 | 6-33440814-C-T | -14.050 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 2.90 | Destabilizing | 0.2 | 2.55 | Destabilizing | 2.73 | Destabilizing | 1.12 | Destabilizing | 0.736 | Likely Pathogenic | -3.98 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.38 | Pathogenic | 0.04 | Affected | 3.38 | 34 | 0.0816 | 0.2007 | 0 | 2 | -1.0 | 34.02 | ||||||||||||||||||||||||||
| c.1777C>T | L593F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L593F is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, and FATHMM, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Predictions that are inconclusive—FoldX, Foldetta, premPS, and AlphaMissense‑Optimized—are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.009728 | Structured | 0.110534 | Uncertain | 0.941 | 0.151 | 0.000 | -12.723 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 1.46 | Ambiguous | 0.4 | 0.24 | Likely Benign | 0.85 | Ambiguous | 0.62 | Ambiguous | 0.304 | Likely Benign | -3.78 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 2.95 | Benign | 0.01 | Affected | 0.0884 | 0.2631 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1792C>T | L598F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L598F is not reported in ClinVar and is absent from gnomAD. Computational predictors show mixed results: benign calls come from REVEL, Rosetta, premPS, SIFT, and FATHMM; pathogenic calls come from SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. FoldX and AlphaMissense‑Optimized returned uncertain results, which are treated as unavailable evidence. High‑accuracy tools give a split view: AlphaMissense‑Optimized is uncertain; the SGM‑Consensus majority vote (AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts Likely Pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the balance of evidence leans toward pathogenicity, with six pathogenic versus five benign predictions and no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.007259 | Structured | 0.147872 | Uncertain | 0.953 | 0.154 | 0.000 | -10.649 | Likely Pathogenic | 0.820 | Likely Pathogenic | Ambiguous | 1.12 | Ambiguous | 0.7 | -0.24 | Likely Benign | 0.44 | Likely Benign | 0.47 | Likely Benign | 0.283 | Likely Benign | -3.91 | Deleterious | 0.999 | Probably Damaging | 0.946 | Probably Damaging | 3.62 | Benign | 0.22 | Tolerated | 0.0630 | 0.1414 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1819C>T | L607F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L607F is catalogued in gnomAD (6‑33440871‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all report pathogenic or likely pathogenic. Only FoldX predicts a benign outcome, while Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show the SGM‑Consensus as likely pathogenic, AlphaMissense‑Optimized as uncertain, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for L607F, and this conclusion is not contradicted by ClinVar status (none). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.048328 | Structured | 0.194229 | Uncertain | 0.869 | 0.250 | 0.000 | 6-33440871-C-T | 1 | 6.19e-7 | -13.654 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 0.23 | Likely Benign | 0.1 | 1.20 | Ambiguous | 0.72 | Ambiguous | 0.61 | Ambiguous | 0.758 | Likely Pathogenic | -3.98 | Deleterious | 0.998 | Probably Damaging | 0.997 | Probably Damaging | -1.54 | Pathogenic | 0.01 | Affected | 3.37 | 35 | 0.0872 | 0.2816 | 0 | 2 | -1.0 | 34.02 | ||||||||||||||||||||||||
| c.1828C>T | L610F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L610F is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a pathogenic effect include SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—all of which classify the variant as pathogenic. No tools predict a benign outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a pathogenic effect. Based on the consensus of these predictions, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.271506 | Structured | 0.209504 | Uncertain | 0.888 | 0.253 | 0.000 | -13.244 | Likely Pathogenic | 0.808 | Likely Pathogenic | Ambiguous | 6.24 | Destabilizing | 1.1 | 3.40 | Destabilizing | 4.82 | Destabilizing | 0.68 | Ambiguous | 0.782 | Likely Pathogenic | -3.92 | Deleterious | 0.998 | Probably Damaging | 0.997 | Probably Damaging | -1.52 | Pathogenic | 0.01 | Affected | 0.0834 | 0.2816 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1835A>G | Q612R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q612R is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are SIFT and FoldX. Tools that agree on a pathogenic effect are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. Predictions that are uncertain or inconclusive are Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show that the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenicity, while AlphaMissense‑Optimized and Foldetta are uncertain. No high‑accuracy tool provides a benign prediction. Overall, the majority of available evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.203988 | Uncertain | 0.822 | 0.263 | 0.000 | -10.571 | Likely Pathogenic | 0.837 | Likely Pathogenic | Ambiguous | -0.35 | Likely Benign | 0.2 | 1.56 | Ambiguous | 0.61 | Ambiguous | 0.78 | Ambiguous | 0.683 | Likely Pathogenic | -3.85 | Deleterious | 0.956 | Probably Damaging | 0.969 | Probably Damaging | -1.29 | Pathogenic | 0.10 | Tolerated | 0.1480 | 0.2196 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1853A>G | Q618R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q618R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus, REVEL, FoldX, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while Rosetta and premPS are inconclusive. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts benign. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | -2.513 | Likely Benign | 0.207 | Likely Benign | Likely Benign | -0.43 | Likely Benign | 0.1 | 0.60 | Ambiguous | 0.09 | Likely Benign | -0.66 | Ambiguous | 0.285 | Likely Benign | 1.22 | Neutral | 0.005 | Benign | 0.009 | Benign | -1.24 | Pathogenic | 1.00 | Tolerated | 0.1218 | 0.1163 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1867C>T | L623F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L623F is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus predicts likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) yields an uncertain result. No predictions are missing or inconclusive. Based on the overwhelming agreement among pathogenic predictions and the lack of a benign signal, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.060667 | Uncertain | 0.962 | 0.211 | 0.000 | -11.655 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 1.22 | Ambiguous | 0.2 | 0.92 | Ambiguous | 1.07 | Ambiguous | 0.57 | Ambiguous | 0.440 | Likely Benign | -3.98 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 1.65 | Pathogenic | 0.02 | Affected | 0.0838 | 0.3016 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1873C>T | L625F 2D AIThe SynGAP1 missense variant L625F is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM. Tools that predict a pathogenic effect include SGM‑Consensus (Likely Pathogenic), Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Predictions that are uncertain or inconclusive are FoldX, premPS, and Foldetta. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts Pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts Likely Pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is Uncertain. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.229226 | Structured | 0.045896 | Uncertain | 0.966 | 0.215 | 0.000 | -12.989 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 1.70 | Ambiguous | 1.3 | 2.26 | Destabilizing | 1.98 | Ambiguous | 0.74 | Ambiguous | 0.479 | Likely Benign | -3.82 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.01 | Benign | 0.01 | Affected | 0.0586 | 0.2998 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1892A>G | Q631R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q631R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: benign predictions come from FoldX, Foldetta, and FATHMM, while pathogenic predictions are made by SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; Rosetta and premPS are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a pathogenic impact for Q631R, and this conclusion is not contradicted by any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.038963 | Uncertain | 0.948 | 0.230 | 0.000 | -14.881 | Likely Pathogenic | 0.958 | Likely Pathogenic | Likely Pathogenic | -0.34 | Likely Benign | 0.1 | 0.83 | Ambiguous | 0.25 | Likely Benign | 0.77 | Ambiguous | 0.627 | Likely Pathogenic | -3.98 | Deleterious | 0.973 | Probably Damaging | 0.969 | Probably Damaging | 2.77 | Benign | 0.01 | Affected | 0.1193 | 0.0653 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1982A>G | Q661R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q661R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that predict a pathogenic effect are polyPhen2_HumDiv and ESM1b. FoldX, Rosetta, Foldetta, and AlphaMissense‑Default are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign prediction, while Foldetta remains uncertain. Overall, the majority of available evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.048328 | Structured | 0.117089 | Uncertain | 0.924 | 0.309 | 0.000 | -10.386 | Likely Pathogenic | 0.479 | Ambiguous | Likely Benign | -1.12 | Ambiguous | 0.0 | -0.59 | Ambiguous | -0.86 | Ambiguous | -0.15 | Likely Benign | 0.281 | Likely Benign | -2.06 | Neutral | 0.812 | Possibly Damaging | 0.251 | Benign | 3.52 | Benign | 0.18 | Tolerated | 0.1502 | 0.2196 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||
| c.1985A>G | Q662R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q662R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that assess sequence conservation, structural impact, or functional effect all converge on a benign outcome: SIFT, PolyPhen‑2 (HumDiv and HumVar), REVEL, PROVEAN, FATHMM, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, premPS, FoldX, and Rosetta all predict benign. No tool in the dataset indicates pathogenicity. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts benign. Thus, based on the available predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.046336 | Structured | 0.103446 | Uncertain | 0.932 | 0.323 | 0.000 | -6.678 | Likely Benign | 0.300 | Likely Benign | Likely Benign | -0.41 | Likely Benign | 0.0 | 0.10 | Likely Benign | -0.16 | Likely Benign | -0.42 | Likely Benign | 0.159 | Likely Benign | 0.49 | Neutral | 0.428 | Benign | 0.095 | Benign | 3.48 | Benign | 1.00 | Tolerated | 0.2023 | 0.1391 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.1992G>C | L664F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L664F resides in the GAP domain. ClinVar lists no entry for this variant, and it is absent from gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM. Those that predict a pathogenic effect include SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Predictions that are uncertain (FoldX, Rosetta, Foldetta, premPS) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic, SGM‑Consensus as Likely Pathogenic, and Foldetta as Uncertain. Overall, the majority of available predictions support a pathogenic impact. Therefore, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.100716 | Structured | 0.089318 | Uncertain | 0.937 | 0.339 | 0.000 | -12.834 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 1.98 | Ambiguous | 0.3 | 1.03 | Ambiguous | 1.51 | Ambiguous | 0.55 | Ambiguous | 0.355 | Likely Benign | -3.99 | Deleterious | 0.999 | Probably Damaging | 0.895 | Possibly Damaging | 3.05 | Benign | 0.03 | Affected | 0.0537 | 0.2311 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.1992G>T | L664F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L664F resides in the GAP domain. ClinVar lists no entry for this variant, and it is absent from gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM. Those that predict a pathogenic effect include SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Predictions that are uncertain (FoldX, Rosetta, Foldetta, premPS) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic, SGM‑Consensus as Likely Pathogenic, and Foldetta as Uncertain. Overall, the majority of available predictions support a pathogenic impact. Therefore, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.100716 | Structured | 0.089318 | Uncertain | 0.937 | 0.339 | 0.000 | -12.834 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 1.98 | Ambiguous | 0.3 | 1.03 | Ambiguous | 1.51 | Ambiguous | 0.55 | Ambiguous | 0.355 | Likely Benign | -3.99 | Deleterious | 0.999 | Probably Damaging | 0.895 | Possibly Damaging | 3.05 | Benign | 0.03 | Affected | 0.0537 | 0.2311 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2055G>C | L685F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L685F is not reported in ClinVar and has no gnomAD entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, and FATHMM, whereas pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the change as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic, while Foldetta’s stability analysis is inconclusive. FoldX and Rosetta predictions are uncertain and are treated as unavailable. Overall, the preponderance of evidence points to a pathogenic impact for the variant, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.162061 | Uncertain | 0.913 | 0.280 | 0.000 | -12.304 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 1.62 | Ambiguous | 0.2 | 1.80 | Ambiguous | 1.71 | Ambiguous | 0.50 | Likely Benign | 0.300 | Likely Benign | -3.99 | Deleterious | 0.999 | Probably Damaging | 0.895 | Possibly Damaging | 3.33 | Benign | 0.01 | Affected | 0.0724 | 0.2367 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2055G>T | L685F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L685F is not reported in ClinVar and has no gnomAD entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, and FATHMM, whereas pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the change as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic, while Foldetta’s stability analysis is inconclusive. FoldX and Rosetta predictions are uncertain and are treated as unavailable. Overall, the preponderance of evidence points to a pathogenic impact for the variant, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.162061 | Uncertain | 0.913 | 0.280 | 0.000 | -12.304 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 1.62 | Ambiguous | 0.2 | 1.80 | Ambiguous | 1.71 | Ambiguous | 0.50 | Likely Benign | 0.300 | Likely Benign | -3.99 | Deleterious | 0.999 | Probably Damaging | 0.895 | Possibly Damaging | 3.33 | Benign | 0.01 | Affected | 0.0724 | 0.2367 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2065C>T | L689F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L689F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas a majority of tools predict a pathogenic impact: FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus. Two tools (Rosetta and premPS) yield uncertain results. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM Consensus is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No prediction or stability result is missing or inconclusive. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.042364 | Structured | 0.227227 | Uncertain | 0.963 | 0.248 | 0.000 | -9.817 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 2.45 | Destabilizing | 0.2 | 1.95 | Ambiguous | 2.20 | Destabilizing | 0.67 | Ambiguous | 0.286 | Likely Benign | -3.98 | Deleterious | 0.999 | Probably Damaging | 0.860 | Possibly Damaging | 3.18 | Benign | 0.00 | Affected | 0.0608 | 0.2891 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2086C>T | L696F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L696F has no ClinVar entry and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict any ClinVar annotation because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.200174 | Structured | 0.390093 | Uncertain | 0.962 | 0.267 | 0.000 | -9.651 | Likely Pathogenic | 0.897 | Likely Pathogenic | Ambiguous | 0.14 | Likely Benign | 0.1 | 0.74 | Ambiguous | 0.44 | Likely Benign | 0.55 | Ambiguous | 0.422 | Likely Benign | -3.79 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | 3.05 | Benign | 0.00 | Affected | 0.0667 | 0.2008 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2105A>G | Q702R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q702R is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Predictions that remain inconclusive are Rosetta, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to equal benign and pathogenic signals. Overall, the majority of available predictions lean toward a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.397258 | Uncertain | 0.907 | 0.345 | 0.000 | Uncertain | 1 | -7.894 | In-Between | 0.348 | Ambiguous | Likely Benign | -0.31 | Likely Benign | 0.1 | 0.63 | Ambiguous | 0.16 | Likely Benign | 0.13 | Likely Benign | 0.294 | Likely Benign | -3.14 | Deleterious | 0.909 | Possibly Damaging | 0.889 | Possibly Damaging | 3.43 | Benign | 0.02 | Affected | 3.47 | 10 | 0.1225 | 0.0605 | 1 | 1 | -1.0 | 28.06 | 270.3 | -52.9 | 0.0 | 0.0 | 0.0 | 0.1 | X | Potentially Pathogenic | The carboxamide side chain of Gln702 is located at the end and outer surface of an α-helix (res. Leu685-Gln702), where it does not directly form hydrogen bonds with any residues in the WT simulations. In the variant simulations, the positively charged guanidinium group of Arg702 forms a salt bridge with the negatively charged carboxylate group of Glu698 on the same helix and/or hydrogen bonds with the backbone carbonyl group of Ala438 on an opposite α-helix (res. Tyr428-Glu436). Consequently, the residue swap could strengthen the tertiary structure assembly, which could have either positive or negative effects on its function. | |||||||||||||||||
| c.2107C>T | L703F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L703F is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL, premPS, FATHMM, and AlphaMissense‑Optimized, while those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. FoldX, Rosetta, and Foldetta provide uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta remains unavailable. Overall, the majority of reliable predictions lean toward pathogenicity, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.144935 | Structured | 0.388282 | Uncertain | 0.929 | 0.353 | 0.000 | -10.929 | Likely Pathogenic | 0.768 | Likely Pathogenic | Likely Benign | 1.03 | Ambiguous | 0.1 | 0.59 | Ambiguous | 0.81 | Ambiguous | 0.50 | Likely Benign | 0.247 | Likely Benign | -3.25 | Deleterious | 0.994 | Probably Damaging | 0.806 | Possibly Damaging | 3.12 | Benign | 0.00 | Affected | 0.0600 | 0.2555 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2122C>T | L708F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L708F is not reported in ClinVar and is present in gnomAD (ID 6‑33441587‑C‑T). Functional prediction tools that reach consensus classify the change as benign: REVEL, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Pathogenic predictions are limited to polyPhen‑2 HumDiv and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Default) return uncertain or no result. High‑accuracy assessments give AlphaMissense‑Optimized a benign score, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, and Foldetta indicates no significant destabilization (uncertain). Overall, the preponderance of evidence supports a benign effect for L708F, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.250310 | Structured | 0.365875 | Uncertain | 0.931 | 0.378 | 0.000 | 6-33441587-C-T | 2 | 1.24e-6 | -9.154 | Likely Pathogenic | 0.436 | Ambiguous | Likely Benign | 1.48 | Ambiguous | 0.3 | 0.93 | Ambiguous | 1.21 | Ambiguous | 0.37 | Likely Benign | 0.110 | Likely Benign | -2.46 | Neutral | 0.931 | Possibly Damaging | 0.326 | Benign | 3.29 | Benign | 0.07 | Tolerated | 3.50 | 9 | 0.0497 | 0.2366 | 0 | 2 | -1.0 | 34.02 | |||||||||||||||||||||||||
| c.2149C>T | L717F 2D AIThe SynGAP1 missense variant L717F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and FATHMM, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM‑Consensus indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is also uncertain. No folding‑stability metrics (FoldX, Rosetta, premPS) provide decisive evidence. Overall, the majority of predictions lean toward pathogenicity, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.239899 | Structured | 0.429342 | Uncertain | 0.969 | 0.397 | 0.000 | -10.917 | Likely Pathogenic | 0.903 | Likely Pathogenic | Ambiguous | 0.74 | Ambiguous | 0.6 | 0.59 | Ambiguous | 0.67 | Ambiguous | 0.63 | Ambiguous | 0.157 | Likely Benign | -2.56 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | 3.36 | Benign | 0.02 | Affected | 0.0444 | 0.2207 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2152C>T | L718F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L718F lies in the GAP domain. ClinVar has no entry for this variant, and it is not present in gnomAD. Prediction tools cluster into two groups: benign predictions are made only by REVEL, whereas the remaining tools (FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict pathogenicity; premPS is uncertain and is not counted as evidence. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Pathogenic,” and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. Based on the overwhelming consensus of these predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.298791 | Structured | 0.438417 | Uncertain | 0.966 | 0.385 | 0.000 | -11.302 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 3.87 | Destabilizing | 0.2 | 2.64 | Destabilizing | 3.26 | Destabilizing | 0.78 | Ambiguous | 0.347 | Likely Benign | -3.73 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 1.32 | Pathogenic | 0.00 | Affected | 0.0590 | 0.2979 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.2192A>G | Q731R 2D AIThe SynGAP1 missense variant Q731R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.750527 | Disordered | 0.415202 | Uncertain | 0.507 | 0.654 | 0.750 | -5.873 | Likely Benign | 0.267 | Likely Benign | Likely Benign | 0.082 | Likely Benign | -1.63 | Neutral | 0.604 | Possibly Damaging | 0.293 | Benign | 2.66 | Benign | 0.14 | Tolerated | 0.1571 | 0.1775 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2198A>G | Q733R 2D AIThe SynGAP1 missense variant Q733R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for Q733R, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.874069 | Disordered | 0.410831 | Uncertain | 0.331 | 0.686 | 0.875 | -5.986 | Likely Benign | 0.291 | Likely Benign | Likely Benign | 0.117 | Likely Benign | -1.96 | Neutral | 0.950 | Possibly Damaging | 0.612 | Possibly Damaging | 2.57 | Benign | 0.04 | Affected | 0.1419 | 0.1279 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2210A>G | Q737R 2D AIThe SynGAP1 missense variant Q737R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict pathogenicity. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for Q737R, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.889439 | Disordered | 0.425743 | Uncertain | 0.323 | 0.803 | 0.875 | -4.524 | Likely Benign | 0.159 | Likely Benign | Likely Benign | 0.077 | Likely Benign | -1.02 | Neutral | 0.986 | Probably Damaging | 0.793 | Possibly Damaging | 2.74 | Benign | 0.06 | Tolerated | 0.1688 | 0.1852 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2231A>G | Q744R 2D AIThe SynGAP1 missense variant Q744R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta results are unavailable. Overall, the majority of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.540428 | Binding | 0.316 | 0.866 | 0.875 | -2.207 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.062 | Likely Benign | -0.43 | Neutral | 0.001 | Benign | 0.002 | Benign | 2.80 | Benign | 0.05 | Affected | 0.1437 | 0.1588 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2267A>G | Q756R 2D AIThe SynGAP1 missense variant Q756R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for Q756R, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.458154 | Structured | 0.806299 | Binding | 0.340 | 0.866 | 0.250 | -5.044 | Likely Benign | 0.307 | Likely Benign | Likely Benign | 0.245 | Likely Benign | -1.77 | Neutral | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.58 | Pathogenic | 0.14 | Tolerated | 0.1500 | 0.2440 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2287C>T | L763F 2D AIThe SynGAP1 missense variant L763F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for this variant. There is no ClinVar entry to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.380708 | Structured | 0.918636 | Binding | 0.351 | 0.865 | 0.125 | -4.127 | Likely Benign | 0.255 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -0.71 | Neutral | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.39 | Pathogenic | 0.19 | Tolerated | 0.0584 | 0.3140 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2305C>T | L769F 2D AIThe SynGAP1 missense variant L769F is listed in ClinVar (ID 3617309.0) with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority of the high‑accuracy tools) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions—including the high‑accuracy tools—suggest the variant is most likely benign, which is consistent with its ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.928432 | Binding | 0.367 | 0.883 | 0.250 | Uncertain | 1 | -5.044 | Likely Benign | 0.146 | Likely Benign | Likely Benign | 0.060 | Likely Benign | -0.89 | Neutral | 0.925 | Possibly Damaging | 0.510 | Possibly Damaging | 3.94 | Benign | 0.02 | Affected | 0.0554 | 0.2881 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||
| c.2309A>G | Q770R 2D AIThe SynGAP1 missense variant Q770R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable. Overall, the collective evidence strongly suggests that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.923732 | Binding | 0.328 | 0.887 | 0.250 | -3.873 | Likely Benign | 0.344 | Ambiguous | Likely Benign | 0.175 | Likely Benign | -1.38 | Neutral | 0.194 | Benign | 0.071 | Benign | 4.14 | Benign | 0.07 | Tolerated | 0.1610 | 0.2580 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2329C>T | L777F 2D AIThe SynGAP1 missense variant L777F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.408655 | Structured | 0.876129 | Binding | 0.336 | 0.882 | 0.250 | -4.499 | Likely Benign | 0.175 | Likely Benign | Likely Benign | 0.115 | Likely Benign | -1.91 | Neutral | 0.968 | Probably Damaging | 0.966 | Probably Damaging | 3.98 | Benign | 0.01 | Affected | 0.0697 | 0.3576 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2356C>T | L786F 2D AIThe SynGAP1 missense variant L786F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas a majority of tools (SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default) predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome; Foldetta results are unavailable. Overall, the balance of evidence leans toward pathogenicity, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | SH3-binding motif | 0.882776 | Disordered | 0.655253 | Binding | 0.341 | 0.895 | 0.750 | -4.949 | Likely Benign | 0.578 | Likely Pathogenic | Likely Benign | 0.112 | Likely Benign | -2.53 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.83 | Pathogenic | 0.00 | Affected | 0.0765 | 0.3847 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.2495A>G | Q832R 2D AIThe SynGAP1 missense variant Q832R is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.720929 | Disordered | 0.619913 | Binding | 0.290 | 0.877 | 0.375 | -3.680 | Likely Benign | 0.203 | Likely Benign | Likely Benign | 0.087 | Likely Benign | -1.17 | Neutral | 0.912 | Possibly Damaging | 0.424 | Benign | 2.74 | Benign | 0.09 | Tolerated | 0.1494 | 0.1434 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2538A>C | L846F 2D AIThe SynGAP1 missense variant L846F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as Likely Pathogenic. AlphaMissense‑Optimized yields an uncertain result, and no Foldetta (FoldX‑MD/Rosetta) stability assessment is available. Overall, the preponderance of evidence from multiple high‑accuracy predictors indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.653063 | Disordered | 0.589606 | Binding | 0.349 | 0.825 | 0.500 | -10.559 | Likely Pathogenic | 0.862 | Likely Pathogenic | Ambiguous | 0.206 | Likely Benign | -2.88 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.0665 | 0.2912 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2538A>T | L846F 2D AIThe SynGAP1 missense variant L846F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—predict a pathogenic or deleterious impact, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Pathogenic. AlphaMissense‑Optimized is uncertain, providing no definitive direction. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Overall, the majority of predictions indicate a pathogenic effect, and this conclusion does not contradict any ClinVar status because none is available. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.653063 | Disordered | 0.589606 | Binding | 0.349 | 0.825 | 0.500 | -10.559 | Likely Pathogenic | 0.862 | Likely Pathogenic | Ambiguous | 0.206 | Likely Benign | -2.88 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.0665 | 0.2912 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2540A>G | Q847R 2D AIThe SynGAP1 missense variant Q847R has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Optimized; pathogenic calls come from PROVEAN, SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further reveal AlphaMissense‑Optimized as benign, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Thus, the overall prediction leans toward benign based on the majority of tools, but the high‑accuracy SGM‑Consensus contradicts this by indicating likely pathogenic. No ClinVar annotation exists, so there is no conflict with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.653063 | Disordered | 0.577677 | Binding | 0.282 | 0.818 | 0.500 | -3.232 | Likely Benign | 0.662 | Likely Pathogenic | Likely Benign | 0.256 | Likely Benign | -2.63 | Deleterious | 0.014 | Benign | 0.026 | Benign | 2.30 | Pathogenic | 0.00 | Affected | 0.1404 | 0.1979 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2563C>T | L855F 2D AIThe SynGAP1 missense variant L855F is predicted to be benign by all evaluated in‑silico tools. Consensus predictions from **SGM‑Consensus** (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classify the variant as *Likely Benign*. High‑accuracy predictors **AlphaMissense‑Optimized** also report a benign effect. Other pathogenicity predictors—REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default—uniformly predict benign. No tools predict pathogenicity. **Foldetta**, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its stability impact remains unknown. The variant is not listed in ClinVar and has no entry in gnomAD, so no population frequency or clinical annotation is available. Based on the unanimous benign predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.485558 | Uncertain | 0.285 | 0.823 | 0.625 | -4.985 | Likely Benign | 0.106 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -1.86 | Neutral | 0.411 | Benign | 0.187 | Benign | 4.00 | Benign | 0.12 | Tolerated | 0.0700 | 0.3529 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2621A>G | Q874R 2D AIThe SynGAP1 missense variant Q874R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized independently predicts a benign effect. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.490133 | Structured | 0.635258 | Binding | 0.289 | 0.873 | 0.250 | -4.834 | Likely Benign | 0.527 | Ambiguous | Likely Benign | 0.200 | Likely Benign | -2.33 | Neutral | 0.985 | Probably Damaging | 0.982 | Probably Damaging | 2.69 | Benign | 0.00 | Affected | 0.1619 | 0.2924 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2643G>C | L881F 2D AIThe SynGAP1 missense variant L881F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for L881F, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.629350 | Binding | 0.299 | 0.874 | 0.250 | -5.759 | Likely Benign | 0.101 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.87 | Neutral | 0.818 | Possibly Damaging | 0.360 | Benign | 2.48 | Pathogenic | 0.02 | Affected | 0.0711 | 0.3415 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2643G>T | L881F 2D AIThe SynGAP1 missense variant L881F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for L881F, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.629350 | Binding | 0.299 | 0.874 | 0.250 | -5.759 | Likely Benign | 0.101 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.87 | Neutral | 0.818 | Possibly Damaging | 0.360 | Benign | 2.48 | Pathogenic | 0.02 | Affected | 0.0711 | 0.3415 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2671C>T | L891F 2D AIThe SynGAP1 missense variant L891F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available. Overall, the majority of evidence points to a benign effect for L891F, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.712013 | Disordered | 0.505861 | Binding | 0.305 | 0.923 | 0.750 | -5.650 | Likely Benign | 0.126 | Likely Benign | Likely Benign | 0.102 | Likely Benign | -1.80 | Neutral | 0.970 | Probably Damaging | 0.744 | Possibly Damaging | 2.66 | Benign | 0.04 | Affected | 0.0652 | 0.2907 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2678A>G | Q893R 2D AIThe SynGAP1 missense variant Q893R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.447267 | Uncertain | 0.310 | 0.925 | 0.750 | -3.338 | Likely Benign | 0.392 | Ambiguous | Likely Benign | 0.056 | Likely Benign | -1.43 | Neutral | 0.802 | Possibly Damaging | 0.333 | Benign | 2.79 | Benign | 0.09 | Tolerated | 0.1565 | 0.2412 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2716C>T | L906F 2D AIThe SynGAP1 missense variant L906F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for the variant. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.604312 | Disordered | 0.644316 | Binding | 0.315 | 0.920 | 0.250 | -4.700 | Likely Benign | 0.304 | Likely Benign | Likely Benign | 0.113 | Likely Benign | -1.79 | Neutral | 0.999 | Probably Damaging | 0.979 | Probably Damaging | 2.22 | Pathogenic | 0.18 | Tolerated | 0.0753 | 0.3342 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2723A>G | Q908R 2D AIThe SynGAP1 missense variant Q908R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (both HumDiv and HumVar) predict a pathogenic outcome. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.608892 | Disordered | 0.678728 | Binding | 0.275 | 0.917 | 0.250 | -4.284 | Likely Benign | 0.485 | Ambiguous | Likely Benign | 0.170 | Likely Benign | -1.02 | Neutral | 0.985 | Probably Damaging | 0.982 | Probably Damaging | 2.75 | Benign | 0.10 | Tolerated | 0.1391 | 0.1743 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2756A>G | Q919R 2D AIThe SynGAP1 missense variant Q919R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Benign”). In contrast, the two polyPhen‑2 implementations (HumDiv and HumVar) predict a pathogenic effect. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also likely benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.911223 | Binding | 0.299 | 0.841 | 0.250 | -3.636 | Likely Benign | 0.272 | Likely Benign | Likely Benign | 0.105 | Likely Benign | -0.96 | Neutral | 0.961 | Probably Damaging | 0.596 | Possibly Damaging | 2.65 | Benign | 0.85 | Tolerated | 0.1558 | 0.2305 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2759A>G | Q920R 2D AIThe SynGAP1 missense variant Q920R is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors shows a split: benign calls come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic calls arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy tools further support this view: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus confirms Likely Benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.699094 | Disordered | 0.927260 | Binding | 0.306 | 0.845 | 0.250 | -3.170 | Likely Benign | 0.352 | Ambiguous | Likely Benign | 0.168 | Likely Benign | -1.41 | Neutral | 0.961 | Probably Damaging | 0.596 | Possibly Damaging | 2.63 | Benign | 0.00 | Affected | 0.1626 | 0.2369 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.2773C>T | L925F 2D AIThe SynGAP1 missense variant L925F is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify it as benign include REVEL and ESM1b, whereas the majority of tools predict it to be pathogenic: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Pathogenic. Foldetta results are not available for this variant. Overall, the preponderance of computational evidence points to a pathogenic impact for the L925F substitution, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.977963 | Binding | 0.290 | 0.852 | 0.125 | -4.147 | Likely Benign | 0.966 | Likely Pathogenic | Likely Pathogenic | 0.242 | Likely Benign | -3.04 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.32 | Pathogenic | 0.00 | Affected | 0.0897 | 0.2881 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2783A>G | Q928R 2D AIThe SynGAP1 missense variant Q928R is listed in gnomAD (ID 6‑33443335‑A‑G) but has no ClinVar entry. Functional prediction tools fall into two consensus groups: benign predictions come from REVEL and ESM1b, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). AlphaMissense‑Optimized returns an uncertain result, and no Foldetta (FoldX‑MD/Rosetta stability) data are available. High‑accuracy assessments therefore indicate a likely pathogenic outcome (SGM‑Consensus) with no contradictory evidence from ClinVar. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not conflict with the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.986260 | Binding | 0.324 | 0.852 | 0.250 | 6-33443335-A-G | 1 | 6.20e-7 | -4.089 | Likely Benign | 0.826 | Likely Pathogenic | Ambiguous | 0.290 | Likely Benign | -2.91 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.58 | Pathogenic | 0.00 | Affected | 4.32 | 4 | 0.1357 | 0.3453 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||
| c.2791C>T | L931F 2D AIThe SynGAP1 missense variant L931F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and ESM1b, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. A high‑accuracy consensus (SGM) that aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a 2‑to‑2 split, leaving the consensus inconclusive. Foldetta, which assesses protein‑folding stability via FoldX‑MD and Rosetta, has no available result for this variant. Consequently, the overall evidence leans toward pathogenicity, driven by the majority of standard predictors, and does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.549308 | Disordered | 0.989212 | Binding | 0.335 | 0.856 | 0.375 | -5.101 | Likely Benign | 0.790 | Likely Pathogenic | Ambiguous | 0.167 | Likely Benign | -2.00 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.41 | Pathogenic | 0.04 | Affected | 0.0718 | 0.3208 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.2962C>T | L988F 2D AIThe SynGAP1 missense variant L988F is listed in ClinVar (ID 968833.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443514‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 (HumDiv and HumVar) and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is not in conflict with the ClinVar “Uncertain” classification. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.918781 | Binding | 0.360 | 0.913 | 0.750 | Uncertain | 1 | 6-33443514-C-T | 1 | 6.20e-7 | -4.368 | Likely Benign | 0.356 | Ambiguous | Likely Benign | 0.135 | Likely Benign | -1.70 | Neutral | 0.977 | Probably Damaging | 0.900 | Possibly Damaging | 2.69 | Benign | 0.00 | Affected | 4.32 | 2 | 0.0693 | 0.3592 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||
| c.3001C>T | L1001F 2D AIThe SynGAP1 missense variant L1001F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while the SGM‑Consensus (derived from the majority of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized scores benign, and the SGM‑Consensus (majority vote of the four high‑confidence tools) is benign; Foldetta data are unavailable. Overall, the preponderance of evidence supports a benign classification for L1001F, and this conclusion does not conflict with ClinVar, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.958507 | Binding | 0.269 | 0.902 | 0.375 | -4.712 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.026 | Likely Benign | -0.85 | Neutral | 0.934 | Possibly Damaging | 0.617 | Possibly Damaging | 2.65 | Benign | 0.00 | Affected | 0.0703 | 0.2876 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3062A>G | Q1021R 2D AIThe SynGAP1 missense variant Q1021R is catalogued in gnomAD (ID 6‑33443614‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves as “Likely Benign” (three benign votes versus one pathogenic). High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.979641 | Binding | 0.326 | 0.763 | 0.500 | 6-33443614-A-G | 1 | 6.20e-7 | -4.467 | Likely Benign | 0.770 | Likely Pathogenic | Likely Benign | 0.174 | Likely Benign | -1.80 | Neutral | 0.985 | Probably Damaging | 0.982 | Probably Damaging | 2.61 | Benign | 0.03 | Affected | 3.77 | 5 | 0.1257 | 0.2405 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||
| c.3064C>T | L1022F 2D AIThe SynGAP1 missense variant L1022F is reported in gnomAD (variant ID 6‑33443616‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are PolyPhen‑2 HumDiv and PolyPhen‑2 HumVar; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact for this variant, and this conclusion is not contradicted by any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.859585 | Disordered | 0.986981 | Binding | 0.339 | 0.752 | 0.500 | 6-33443616-C-T | 2 | 1.24e-6 | -5.035 | Likely Benign | 0.351 | Ambiguous | Likely Benign | 0.072 | Likely Benign | -1.84 | Neutral | 0.971 | Probably Damaging | 0.801 | Possibly Damaging | 2.51 | Benign | 0.07 | Tolerated | 3.77 | 5 | 0.0730 | 0.4231 | 0 | 2 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||
| c.306G>C | L102F 2D AIThe SynGAP1 missense variant L102F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.696014 | Binding | 0.357 | 0.885 | 0.625 | -4.712 | Likely Benign | 0.094 | Likely Benign | Likely Benign | 0.144 | Likely Benign | -0.80 | Neutral | 0.984 | Probably Damaging | 0.969 | Probably Damaging | 4.13 | Benign | 0.00 | Affected | 0.0863 | 0.3205 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.306G>T | L102F 2D AIThe SynGAP1 missense variant L102F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.696014 | Binding | 0.357 | 0.885 | 0.625 | -4.712 | Likely Benign | 0.094 | Likely Benign | Likely Benign | 0.153 | Likely Benign | -0.80 | Neutral | 0.984 | Probably Damaging | 0.969 | Probably Damaging | 4.13 | Benign | 0.00 | Affected | 0.0863 | 0.3205 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3070C>T | L1024F 2D AIThe SynGAP1 missense variant L1024F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it contains both benign and pathogenic calls. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence favors a benign interpretation, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.862302 | Disordered | 0.992699 | Binding | 0.327 | 0.753 | 0.500 | -5.133 | Likely Benign | 0.544 | Ambiguous | Likely Benign | 0.059 | Likely Benign | -2.08 | Neutral | 0.994 | Probably Damaging | 0.924 | Probably Damaging | 2.40 | Pathogenic | 0.07 | Tolerated | 0.0706 | 0.3708 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3074A>G | Q1025R 2D AIThe SynGAP1 missense variant Q1025R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (both HumDiv and HumVar) predict a pathogenic outcome. AlphaMissense‑Default remains uncertain, and the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.879233 | Disordered | 0.993410 | Binding | 0.363 | 0.746 | 0.500 | -4.435 | Likely Benign | 0.480 | Ambiguous | Likely Benign | 0.101 | Likely Benign | -1.28 | Neutral | 0.818 | Possibly Damaging | 0.453 | Possibly Damaging | 2.72 | Benign | 0.10 | Tolerated | 0.1342 | 0.2656 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3086A>G | Q1029R 2D AIThe SynGAP1 missense variant Q1029R has no ClinVar entry and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (both HumDiv and HumVar) predict a pathogenic outcome. AlphaMissense‑Default remains uncertain, and the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates likely benign; Foldetta results are unavailable. Based on the preponderance of evidence from both general and high‑accuracy predictors, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.995643 | Binding | 0.375 | 0.734 | 0.500 | -3.437 | Likely Benign | 0.420 | Ambiguous | Likely Benign | 0.073 | Likely Benign | -0.72 | Neutral | 0.961 | Probably Damaging | 0.677 | Possibly Damaging | 2.73 | Benign | 0.88 | Tolerated | 0.1377 | 0.2420 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3107A>G | Q1036R 2D AIThe SynGAP1 missense variant Q1036R is listed in ClinVar (ID 4746139) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely benign classification (3 benign vs. 1 pathogenic). High‑accuracy assessments further support benignity: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.987955 | Binding | 0.275 | 0.765 | 0.625 | Uncertain | 1 | -3.816 | Likely Benign | 0.596 | Likely Pathogenic | Likely Benign | 0.083 | Likely Benign | -1.32 | Neutral | 0.292 | Benign | 0.071 | Benign | 2.54 | Benign | 0.04 | Affected | 0.1710 | 0.3071 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||
| c.3125A>G | Q1042R 2D AIThe SynGAP1 missense variant Q1042R is listed in ClinVar (ID 2662705.0) with an “Uncertain” clinical significance and is present in gnomAD (variant ID 6‑33443677‑A‑G). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are unavailable. Overall, the majority of evidence supports a benign impact for Q1042R, and this conclusion does not contradict the ClinVar status, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.959333 | Binding | 0.310 | 0.846 | 0.625 | Uncertain | 2 | 6-33443677-A-G | 2 | 1.24e-6 | -2.928 | Likely Benign | 0.413 | Ambiguous | Likely Benign | 0.300 | Likely Benign | -1.39 | Neutral | 0.586 | Possibly Damaging | 0.120 | Benign | 5.48 | Benign | 0.12 | Tolerated | 3.77 | 5 | 0.1929 | 0.3887 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||
| c.3191A>G | Q1064R 2D AIThe SynGAP1 missense variant Q1064R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and there is no conflict with ClinVar status because the variant is not yet classified there. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.953106 | Binding | 0.378 | 0.914 | 0.875 | -2.981 | Likely Benign | 0.202 | Likely Benign | Likely Benign | 0.143 | Likely Benign | -0.28 | Neutral | 0.586 | Possibly Damaging | 0.159 | Benign | 4.19 | Benign | 0.16 | Tolerated | 0.2257 | 0.3654 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3204G>C | L1068F 2D AIThe SynGAP1 missense variant L1068F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of predictions (six benign vs. four pathogenic) favor a benign classification. Thus, the variant is most likely benign based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.947281 | Disordered | 0.981041 | Binding | 0.362 | 0.907 | 0.875 | -6.338 | Likely Benign | 0.182 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -1.38 | Neutral | 0.934 | Possibly Damaging | 0.537 | Possibly Damaging | 2.49 | Pathogenic | 0.00 | Affected | 0.0754 | 0.4202 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3204G>T | L1068F 2D AIThe SynGAP1 missense variant L1068F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of predictions (six benign vs. four pathogenic) favor a benign classification. Thus, the variant is most likely benign based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.947281 | Disordered | 0.981041 | Binding | 0.362 | 0.907 | 0.875 | -6.338 | Likely Benign | 0.182 | Likely Benign | Likely Benign | 0.106 | Likely Benign | -1.38 | Neutral | 0.934 | Possibly Damaging | 0.537 | Possibly Damaging | 2.49 | Pathogenic | 0.00 | Affected | 0.0754 | 0.4202 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3206A>G | Q1069R 2D AIThe SynGAP1 missense variant Q1069R is not reported in ClinVar (ClinVar ID = None) and has no entries in gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable. Overall, the preponderance of evidence points to the variant being most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.943310 | Disordered | 0.981477 | Binding | 0.333 | 0.906 | 0.875 | -3.257 | Likely Benign | 0.467 | Ambiguous | Likely Benign | 0.094 | Likely Benign | -1.17 | Neutral | 0.666 | Possibly Damaging | 0.355 | Benign | 2.73 | Benign | 0.21 | Tolerated | 0.1557 | 0.3114 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3221A>G | Q1074R 2D AIThe SynGAP1 missense variant Q1074R is listed in gnomAD (ID 6‑33443773‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none reported). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.987006 | Binding | 0.339 | 0.897 | 0.750 | 6-33443773-A-G | -5.710 | Likely Benign | 0.594 | Likely Pathogenic | Likely Benign | 0.124 | Likely Benign | -0.54 | Neutral | 0.292 | Benign | 0.157 | Benign | 2.70 | Benign | 0.28 | Tolerated | 3.77 | 5 | 0.1523 | 0.2643 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||
| c.3224A>G | Q1075R 2D AIThe SynGAP1 missense variant Q1075R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default predict a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates Likely Benign. No Foldetta stability analysis is available, so it does not influence the assessment. Overall, the majority of computational evidence points to a benign effect, and this conclusion does not contradict any ClinVar status, as none is assigned to the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.922952 | Disordered | 0.988305 | Binding | 0.354 | 0.894 | 0.750 | -5.039 | Likely Benign | 0.697 | Likely Pathogenic | Likely Benign | 0.142 | Likely Benign | -0.37 | Neutral | 0.985 | Probably Damaging | 0.973 | Probably Damaging | 2.72 | Benign | 0.93 | Tolerated | 0.1504 | 0.2650 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3228G>C | L1076F 2D AIThe SynGAP1 missense variant L1076F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (5 pathogenic vs. 4 benign) lean toward pathogenicity, and this is consistent with the lack of ClinVar annotation. Thus, the variant is most likely pathogenic, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.922952 | Disordered | 0.989617 | Binding | 0.301 | 0.892 | 0.750 | -4.606 | Likely Benign | 0.675 | Likely Pathogenic | Likely Benign | 0.092 | Likely Benign | -0.92 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.42 | Pathogenic | 0.03 | Affected | 0.0759 | 0.3767 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3228G>T | L1076F 2D AIThe SynGAP1 missense variant L1076F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (5 pathogenic vs. 4 benign) lean toward pathogenicity, and this is consistent with the lack of ClinVar annotation. Thus, the variant is most likely pathogenic, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.922952 | Disordered | 0.989617 | Binding | 0.301 | 0.892 | 0.750 | -4.606 | Likely Benign | 0.675 | Likely Pathogenic | Likely Benign | 0.092 | Likely Benign | -0.92 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.42 | Pathogenic | 0.03 | Affected | 0.0759 | 0.3767 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3245A>G | Q1082R 2D AIThe SynGAP1 missense variant Q1082R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.819762 | Disordered | 0.979325 | Binding | 0.347 | 0.896 | 0.875 | -3.584 | Likely Benign | 0.418 | Ambiguous | Likely Benign | 0.076 | Likely Benign | -0.96 | Neutral | 0.224 | Benign | 0.058 | Benign | 4.14 | Benign | 0.10 | Tolerated | 0.1590 | 0.3014 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3276G>C | L1092F 2D AIThe SynGAP1 missense variant L1092F is reported in gnomAD (6‑33443828‑G‑C) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.924947 | Disordered | 0.985431 | Binding | 0.385 | 0.890 | 1.000 | 6-33443828-G-C | 1 | 6.40e-7 | -5.010 | Likely Benign | 0.623 | Likely Pathogenic | Likely Benign | 0.067 | Likely Benign | -1.40 | Neutral | 0.986 | Probably Damaging | 0.823 | Possibly Damaging | 2.63 | Benign | 0.18 | Tolerated | 3.77 | 5 | 0.0772 | 0.4060 | 0 | 2 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||
| c.3276G>T | L1092F 2D AIThe SynGAP1 missense variant L1092F is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools show a split: benign calls from REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also leans benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the lack of ClinVar annotation and gnomAD absence. Therefore, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.924947 | Disordered | 0.985431 | Binding | 0.385 | 0.890 | 1.000 | -5.010 | Likely Benign | 0.623 | Likely Pathogenic | Likely Benign | 0.067 | Likely Benign | -1.40 | Neutral | 0.986 | Probably Damaging | 0.823 | Possibly Damaging | 2.63 | Benign | 0.18 | Tolerated | 3.77 | 5 | 0.0772 | 0.4060 | 0 | 2 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||
| c.3278A>G | Q1093R 2D AIThe SynGAP1 missense variant Q1093R is reported in gnomAD (ID 6‑33443830‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic prediction comes from SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.983312 | Binding | 0.351 | 0.886 | 1.000 | 6-33443830-A-G | 1 | 6.40e-7 | -3.681 | Likely Benign | 0.483 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -1.06 | Neutral | 0.224 | Benign | 0.091 | Benign | 2.73 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1537 | 0.3904 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||
| c.3317A>G | Q1106R 2D AIThe SynGAP1 missense variant Q1106R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. AlphaMissense‑Default is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it contains two benign, one pathogenic, and one uncertain call. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictors (four benign vs. three pathogenic) lean toward a benign interpretation, and this assessment does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.885302 | Disordered | 0.952043 | Binding | 0.382 | 0.870 | 0.875 | -4.211 | Likely Benign | 0.475 | Ambiguous | Likely Benign | 0.133 | Likely Benign | -2.20 | Neutral | 0.985 | Probably Damaging | 0.973 | Probably Damaging | 1.80 | Pathogenic | 0.09 | Tolerated | 0.1412 | 0.2850 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||||
| c.3320A>G | Q1107R 2D AIThe SynGAP1 missense variant Q1107R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. AlphaMissense‑Default is uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” AlphaMissense‑Optimized also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.951017 | Binding | 0.393 | 0.880 | 0.875 | -2.837 | Likely Benign | 0.394 | Ambiguous | Likely Benign | 0.126 | Likely Benign | -1.76 | Neutral | 0.965 | Probably Damaging | 0.425 | Benign | 2.55 | Benign | 0.04 | Affected | 0.1482 | 0.3319 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3325C>T | L1109F 2D AIThe SynGAP1 missense variant L1109F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to the variant being most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.856457 | Disordered | 0.948334 | Binding | 0.343 | 0.893 | 0.875 | -3.459 | Likely Benign | 0.109 | Likely Benign | Likely Benign | 0.089 | Likely Benign | -1.04 | Neutral | 0.832 | Possibly Damaging | 0.324 | Benign | 2.74 | Benign | 0.12 | Tolerated | 0.0780 | 0.4540 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3407A>G | Q1136R 2D AIThe SynGAP1 missense variant Q1136R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign, while the majority‑vote SGM‑Consensus also reports a likely benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact. The high‑accuracy AlphaMissense‑Optimized score is benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also leans benign; Foldetta data are not available. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a benign classification, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.775584 | Binding | 0.321 | 0.884 | 0.875 | -5.526 | Likely Benign | 0.478 | Ambiguous | Likely Benign | 0.173 | Likely Benign | -1.42 | Neutral | 0.801 | Possibly Damaging | 0.506 | Possibly Damaging | 6.28 | Benign | 0.09 | Tolerated | 0.1527 | 0.3073 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3416A>G | Q1139R 2D AIThe SynGAP1 missense variant Q1139R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of ClinVar annotation—there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -4.249 | Likely Benign | 0.267 | Likely Benign | Likely Benign | 0.315 | Likely Benign | -1.91 | Neutral | 0.126 | Benign | 0.138 | Benign | 5.47 | Benign | 0.00 | Affected | 0.1342 | 0.2405 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3432G>C | L1144F 2D AIThe SynGAP1 missense variant L1144F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for the variant, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.950334 | Disordered | 0.726803 | Binding | 0.277 | 0.840 | 1.000 | -4.375 | Likely Benign | 0.406 | Ambiguous | Likely Benign | 0.415 | Likely Benign | -1.71 | Neutral | 0.999 | Probably Damaging | 0.969 | Probably Damaging | 5.69 | Benign | 0.04 | Affected | 0.0685 | 0.3283 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3432G>T | L1144F 2D AIThe SynGAP1 missense variant L1144F is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar) and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the aggregate evidence, the variant is most likely benign. This assessment does not contradict ClinVar status, as no ClinVar claim exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.950334 | Disordered | 0.726803 | Binding | 0.277 | 0.840 | 1.000 | -4.375 | Likely Benign | 0.406 | Ambiguous | Likely Benign | 0.415 | Likely Benign | -1.71 | Neutral | 0.999 | Probably Damaging | 0.969 | Probably Damaging | 5.69 | Benign | 0.04 | Affected | 0.0685 | 0.3283 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.344A>G | Q115R 2D AIThe SynGAP1 missense variant Q115R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign, while the majority of high‑accuracy predictors—AlphaMissense‑Optimized and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—also support a benign classification. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact, and AlphaMissense‑Default remains uncertain. No Foldetta stability analysis is available. Overall, the preponderance of evidence from both general and high‑accuracy tools points to a benign effect for Q115R. This assessment is consistent with the absence of a ClinVar claim, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.657256 | Binding | 0.327 | 0.878 | 0.750 | -1.429 | Likely Benign | 0.497 | Ambiguous | Likely Benign | 0.109 | Likely Benign | -0.46 | Neutral | 0.967 | Probably Damaging | 0.901 | Possibly Damaging | 4.12 | Benign | 0.24 | Tolerated | 0.1469 | 0.1507 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3538C>T | L1180F 2D AIThe SynGAP1 missense variant L1180F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of high‑accuracy predictions lean toward a benign impact, and there is no conflict with ClinVar status. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.559845 | Binding | 0.591 | 0.672 | 0.250 | -5.370 | Likely Benign | 0.953 | Likely Pathogenic | Ambiguous | 0.077 | Likely Benign | -1.29 | Neutral | 0.749 | Possibly Damaging | 0.444 | Benign | 2.65 | Benign | 0.00 | Affected | 0.0516 | 0.2345 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3677A>G | Q1226R 2D AIThe SynGAP1 missense change Q1226R occurs in a coiled‑coil domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus) all predict a pathogenic impact. The high‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain; the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Pathogenic”; Foldetta data are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -12.260 | Likely Pathogenic | 0.883 | Likely Pathogenic | Ambiguous | 0.301 | Likely Benign | -3.16 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.1166 | 0.1234 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3686A>G | Q1229R 2D AIThe SynGAP1 missense variant Q1229R is not reported in ClinVar and has no gnomAD entry. Consensus from multiple in‑silico predictors shows a split: benign calls come from REVEL, SIFT, and AlphaMissense‑Optimized, whereas pathogenic calls arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic verdict. High‑accuracy assessments further illustrate this divergence: AlphaMissense‑Optimized predicts benign, while the SGM‑Consensus (majority vote) indicates pathogenic; Foldetta data are unavailable. With five tools favoring pathogenicity versus two supporting benign, the overall prediction leans toward pathogenic. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.466729 | Uncertain | 0.865 | 0.544 | 0.375 | -8.998 | Likely Pathogenic | 0.518 | Ambiguous | Likely Benign | 0.282 | Likely Benign | -2.53 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.80 | Pathogenic | 0.16 | Tolerated | 0.1178 | 0.1200 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3707A>G | Q1236R 2D AIThe SynGAP1 missense variant Q1236R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b all predict a pathogenic outcome; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence leans toward a benign classification, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.680603 | Disordered | 0.567914 | Binding | 0.883 | 0.537 | 0.125 | -8.186 | Likely Pathogenic | 0.486 | Ambiguous | Likely Benign | 0.235 | Likely Benign | -2.16 | Neutral | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.69 | Benign | 0.01 | Affected | 0.1110 | 0.1228 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3713A>G | Q1238R 2D AIThe SynGAP1 missense variant Q1238R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Consensus from standard predictors shows a split: benign calls come from REVEL, PROVEAN, and AlphaMissense‑Optimized, whereas pathogenic calls arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessment gives AlphaMissense‑Optimized benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence favors a pathogenic interpretation, and this conclusion does not conflict with the ClinVar status, which currently contains no assertion for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.562014 | Disordered | 0.548882 | Binding | 0.855 | 0.545 | 0.250 | -10.216 | Likely Pathogenic | 0.477 | Ambiguous | Likely Benign | 0.229 | Likely Benign | -2.29 | Neutral | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.40 | Pathogenic | 0.04 | Affected | 0.1165 | 0.2019 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3728A>G | Q1243R 2D AIThe SynGAP1 missense variant Q1243R is catalogued in gnomAD (ID 6‑33446720‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status, as none is reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.545602 | Disordered | 0.433693 | Uncertain | 0.887 | 0.551 | 0.500 | 6-33446720-A-G | 1 | 6.20e-7 | -7.131 | In-Between | 0.225 | Likely Benign | Likely Benign | 0.127 | Likely Benign | -1.99 | Neutral | 0.912 | Possibly Damaging | 0.629 | Possibly Damaging | 2.67 | Benign | 0.02 | Affected | 3.77 | 5 | 0.1181 | 0.1028 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||
| c.3749A>G | Q1250R 2D AIThe SynGAP1 missense variant Q1250R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict pathogenicity. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.759478 | Disordered | 0.360484 | Uncertain | 0.881 | 0.554 | 0.750 | -5.183 | Likely Benign | 0.230 | Likely Benign | Likely Benign | 0.148 | Likely Benign | 0.76 | Neutral | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 3.09 | Benign | 1.00 | Tolerated | 0.1330 | 0.0874 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3752A>G | Q1251R 2D AIThe SynGAP1 missense variant Q1251R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of other in silico predictors (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) indicate a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic; this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | -9.456 | Likely Pathogenic | 0.890 | Likely Pathogenic | Ambiguous | 0.236 | Likely Benign | -2.92 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.1257 | 0.1224 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3755A>G | Q1252R 2D AIThe SynGAP1 missense variant Q1252R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: REVEL scores the variant as benign, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all predict pathogenicity. Grouping by consensus, the majority of tools (seven) support a pathogenic effect, while only one tool (REVEL) indicates benign. High‑accuracy assessments further support a deleterious impact: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic; AlphaMissense‑Optimized remains uncertain, and Foldetta data are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus result, the variant is most likely pathogenic. This conclusion aligns with the lack of ClinVar annotation and gnomAD absence, and there is no contradiction with ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -11.890 | Likely Pathogenic | 0.913 | Likely Pathogenic | Ambiguous | 0.249 | Likely Benign | -3.26 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.00 | Pathogenic | 0.00 | Affected | 0.1284 | 0.0854 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3773A>G | Q1258R 2D AIThe SynGAP1 missense variant Q1258R is listed in ClinVar with an uncertain significance (ClinVar ID 3359527.0) and is not observed in gnomAD. Functional prediction tools largely agree on a deleterious effect: pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default, while only REVEL predicts a benign outcome. The high‑accuracy predictors give the following results: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic classification; Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, has no available output for this variant. Based on the preponderance of pathogenic predictions and the SGM Consensus, the variant is most likely pathogenic, which is consistent with its ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | Uncertain | 1 | -10.971 | Likely Pathogenic | 0.931 | Likely Pathogenic | Ambiguous | 0.316 | Likely Benign | -3.19 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.00 | Pathogenic | 0.00 | Affected | 0.1027 | 0.0991 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||
| c.3874C>T | L1292F 2D AIThe SynGAP1 missense variant L1292F is reported in gnomAD (ID 6‑33447922‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.779859 | Disordered | 0.882643 | Binding | 0.586 | 0.800 | 0.625 | 6-33447922-C-T | -5.759 | Likely Benign | 0.200 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -1.88 | Neutral | 0.611 | Possibly Damaging | 0.329 | Benign | 2.49 | Pathogenic | 0.03 | Affected | 3.77 | 5 | 0.0707 | 0.2799 | 0 | 2 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||
| c.3884A>G | Q1295R 2D AIThe SynGAP1 missense variant Q1295R is catalogued in gnomAD (ID 6‑33447932‑A‑G) but has no ClinVar entry. Functional prediction tools show discordant results: benign calls come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments are mixed: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta data are unavailable. Overall, the balance of evidence slightly favors a pathogenic interpretation, though the presence of several benign predictions and the lack of a ClinVar classification mean the variant’s clinical significance remains uncertain. No contradiction exists with ClinVar status, as no ClinVar claim is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | 6-33447932-A-G | -3.342 | Likely Benign | 0.300 | Likely Benign | Likely Benign | 0.351 | Likely Benign | -3.30 | Deleterious | 0.925 | Possibly Damaging | 0.932 | Probably Damaging | 2.44 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1432 | 0.1348 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||
| c.3893A>G | Q1298R 2D AIThe SynGAP1 missense variant Q1298R is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.895297 | Binding | 0.410 | 0.821 | 0.750 | -2.643 | Likely Benign | 0.174 | Likely Benign | Likely Benign | 0.112 | Likely Benign | 1.38 | Neutral | 0.000 | Benign | 0.001 | Benign | 3.15 | Benign | 1.00 | Tolerated | 0.1142 | 0.1628 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3895C>T | L1299F 2D AIThe SynGAP1 missense variant L1299F is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.771762 | Disordered | 0.896323 | Binding | 0.398 | 0.832 | 0.750 | -5.618 | Likely Benign | 0.171 | Likely Benign | Likely Benign | 0.138 | Likely Benign | -1.80 | Neutral | 0.002 | Benign | 0.005 | Benign | 2.69 | Benign | 0.42 | Tolerated | 0.0707 | 0.3342 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.389A>G | Q130R 2D AIThe SynGAP1 missense variant Q130R is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar) and SIFT. AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as benign, and the SGM‑Consensus also reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a benign impact. This conclusion is consistent with the absence of a ClinVar assertion, so there is no contradiction with existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.509769 | Disordered | 0.718853 | Binding | 0.306 | 0.885 | 0.375 | -4.043 | Likely Benign | 0.478 | Ambiguous | Likely Benign | 0.166 | Likely Benign | -0.32 | Neutral | 0.967 | Probably Damaging | 0.901 | Possibly Damaging | 4.21 | Benign | 0.05 | Affected | 0.1697 | 0.2343 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3906G>C | L1302F 2D AISynGAP1 missense variant L1302F is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments indicate AlphaMissense‑Optimized predicts benign, whereas the SGM consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta stability analysis is unavailable. Overall, the majority of predictions (five pathogenic versus four benign) lean toward a pathogenic effect. Thus, the variant is most likely pathogenic, a conclusion that contrasts with its current ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | Uncertain | 1 | -5.674 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.211 | Likely Benign | -2.70 | Deleterious | 0.960 | Probably Damaging | 0.657 | Possibly Damaging | 1.53 | Pathogenic | 0.00 | Affected | 0.0721 | 0.2648 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3906G>T | L1302F 2D AIThe SynGAP1 missense variant L1302F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools that are available indicate a benign outcome (AlphaMissense‑Optimized) while the consensus and stability analyses are missing. Consequently, the overall prediction is ambiguous, but the most reliable evidence points toward a benign effect, and this does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | -5.674 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.215 | Likely Benign | -2.70 | Deleterious | 0.960 | Probably Damaging | 0.657 | Possibly Damaging | 1.53 | Pathogenic | 0.00 | Affected | 0.0721 | 0.2648 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3989A>G | Q1330R 2D AIThe SynGAP1 missense variant Q1330R is listed in gnomAD (ID 6‑33451863‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.943310 | Disordered | 0.931969 | Binding | 0.369 | 0.752 | 0.875 | 6-33451863-A-G | -3.601 | Likely Benign | 0.472 | Ambiguous | Likely Benign | 0.031 | Likely Benign | -1.65 | Neutral | 0.898 | Possibly Damaging | 0.341 | Benign | 3.95 | Benign | 0.03 | Affected | 3.77 | 5 | 0.1349 | 0.2333 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||
| c.440A>G | Q147R 2D AIThe SynGAP1 missense variant Q147R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a 2‑to‑2 split, and Foldetta results are unavailable. Overall, more tools predict a benign outcome (five vs. three pathogenic predictions, with one uncertain). Therefore, the variant is most likely benign based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.541878 | Disordered | 0.503877 | Binding | 0.349 | 0.840 | 0.625 | -12.301 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 0.166 | Likely Benign | -2.22 | Neutral | 0.079 | Benign | 0.052 | Benign | 3.91 | Benign | 0.02 | Affected | 0.1760 | 0.1042 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||||
| c.448C>T | L150F 2D AIThe SynGAP1 missense variant L150F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta results are not available for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and the SGM‑Consensus suggests that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.494003 | Structured | 0.505752 | Binding | 0.299 | 0.839 | 0.625 | -12.459 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.105 | Likely Benign | -2.56 | Deleterious | 0.998 | Probably Damaging | 0.991 | Probably Damaging | 3.69 | Benign | 0.00 | Affected | 0.0580 | 0.3112 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.473A>G | Q158R 2D AIThe SynGAP1 missense variant Q158R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also reports a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. The predictions do not contradict ClinVar status, as ClinVar contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.527565 | Binding | 0.286 | 0.750 | 0.375 | -6.873 | Likely Benign | 0.438 | Ambiguous | Likely Benign | 0.090 | Likely Benign | -0.85 | Neutral | 0.276 | Benign | 0.121 | Benign | 4.14 | Benign | 0.11 | Tolerated | 0.1527 | 0.1498 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.524A>G | Q175R 2D AIThe SynGAP1 missense variant Q175R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM. The only tool that predicts a pathogenic outcome is AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus remains likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.653063 | Disordered | 0.474689 | Uncertain | 0.367 | 0.618 | 0.375 | -5.785 | Likely Benign | 0.789 | Likely Pathogenic | Ambiguous | 0.194 | Likely Benign | -1.29 | Neutral | 0.012 | Benign | 0.006 | Benign | 4.14 | Benign | 0.26 | Tolerated | 0.1485 | 0.1352 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.558G>C | L186F 2D AIThe SynGAP1 missense variant L186F is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), and FATHMM. In contrast, tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Pathogenic.” High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a pathogenic effect, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.458154 | Structured | 0.428613 | Uncertain | 0.397 | 0.617 | 0.500 | Uncertain | 1 | -11.861 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.132 | Likely Benign | -3.03 | Deleterious | 0.009 | Benign | 0.012 | Benign | 3.50 | Benign | 0.00 | Affected | 0.0524 | 0.3177 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||
| c.558G>T | L186F 2D AIThe SynGAP1 missense variant L186F has no ClinVar entry (ClinVar status: not reported) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Pathogenic.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.458154 | Structured | 0.428613 | Uncertain | 0.397 | 0.617 | 0.500 | -11.861 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.132 | Likely Benign | -3.03 | Deleterious | 0.009 | Benign | 0.012 | Benign | 3.50 | Benign | 0.00 | Affected | 0.0524 | 0.3177 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.588G>C | L196F 2D AIThe SynGAP1 missense variant L196F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.284882 | Structured | 0.429452 | Uncertain | 0.489 | 0.521 | 0.125 | -10.245 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.177 | Likely Benign | -3.17 | Deleterious | 0.917 | Possibly Damaging | 0.502 | Possibly Damaging | 3.64 | Benign | 0.01 | Affected | 0.0557 | 0.2756 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.588G>T | L196F 2D AIThe SynGAP1 missense variant L196F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.284882 | Structured | 0.429452 | Uncertain | 0.489 | 0.521 | 0.125 | -10.245 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.177 | Likely Benign | -3.17 | Deleterious | 0.917 | Possibly Damaging | 0.502 | Possibly Damaging | 3.64 | Benign | 0.01 | Affected | 0.0557 | 0.2756 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.592C>T | L198F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L198F has no ClinVar entry and is absent from gnomAD. Functional prediction tools split into two consensus groups: benign predictions come from REVEL, Rosetta, premPS, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments are mixed: AlphaMissense‑Optimized and Foldetta report uncertain results, whereas the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic effect. No evidence from ClinVar contradicts these findings. Overall, the preponderance of pathogenic predictions and the SGM‑Consensus designation suggest that the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.444081 | Structured | 0.431715 | Uncertain | 0.572 | 0.485 | 0.125 | -10.602 | Likely Pathogenic | 0.930 | Likely Pathogenic | Ambiguous | 1.03 | Ambiguous | 0.2 | 0.27 | Likely Benign | 0.65 | Ambiguous | 0.00 | Likely Benign | 0.282 | Likely Benign | -3.33 | Deleterious | 0.997 | Probably Damaging | 0.916 | Probably Damaging | 3.39 | Benign | 0.00 | Affected | 0.0586 | 0.3112 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||
| c.600G>C | L200F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L200F is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33435242‑G‑C). Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Predictions that are inconclusive are FoldX, Rosetta, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.366687 | Structured | 0.428168 | Uncertain | 0.687 | 0.453 | 0.125 | Uncertain | 1 | 6-33435242-G-C | 2 | 1.24e-6 | -7.606 | In-Between | 0.592 | Likely Pathogenic | Likely Benign | 1.00 | Ambiguous | 0.5 | 1.45 | Ambiguous | 1.23 | Ambiguous | 0.43 | Likely Benign | 0.094 | Likely Benign | -1.97 | Neutral | 0.997 | Probably Damaging | 0.916 | Probably Damaging | 4.02 | Benign | 0.17 | Tolerated | 3.46 | 9 | 0.0640 | 0.3120 | 2 | 0 | -1.0 | 34.02 | 250.4 | -15.1 | 0.6 | 0.2 | 0.5 | 0.0 | X | Uncertain | Leu200, a hydrophobic residue located in the N-terminal loop before the first anti-parallel β sheet strand (res. Ile205-Pro208), is replaced by another hydrophobic residue, phenylalanine. Both the phenyl group of Phe200 and the branched iso-butyl hydrocarbon sidechain of Leu200 occupy an inward hydrophobic niche (e.g., Leu246, Val222, Phe231) during the simulations. However, since the model ends abruptly at the N-terminus, no definite conclusions can be drawn from the simulations. | ||||||||||||||
| c.600G>T | L200F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L200F missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Four tools (FoldX, Rosetta, ESM1b, Foldetta) returned uncertain results and are not considered evidence for either side. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (benign), and PROVEAN (benign)—is benign; Foldetta remains uncertain and is treated as unavailable. Overall, the balance of evidence (six benign vs. three pathogenic predictions, with a benign consensus from high‑accuracy methods) indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.366687 | Structured | 0.428168 | Uncertain | 0.687 | 0.453 | 0.125 | -7.606 | In-Between | 0.592 | Likely Pathogenic | Likely Benign | 1.00 | Ambiguous | 0.5 | 1.45 | Ambiguous | 1.23 | Ambiguous | 0.43 | Likely Benign | 0.094 | Likely Benign | -1.97 | Neutral | 0.997 | Probably Damaging | 0.916 | Probably Damaging | 4.02 | Benign | 0.17 | Tolerated | 3.46 | 9 | 0.0640 | 0.3120 | 2 | 0 | -1.0 | 34.02 | 250.4 | -15.1 | 0.6 | 0.2 | 0.5 | 0.0 | X | Uncertain | Leu200, a hydrophobic residue located in the N-terminal loop before the first anti-parallel β sheet strand (res. Ile205-Pro208), is replaced by another hydrophobic residue, phenylalanine. Both the phenyl group of Phe200 and the branched iso-butyl hydrocarbon sidechain of Leu200 occupy an inward hydrophobic niche (e.g., Leu246, Val222, Phe231) during the simulations. However, since the model ends abruptly at the N-terminus, no definite conclusions can be drawn from the simulations. | |||||||||||||||||||
| c.647A>G | Q216R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q216R is not reported in ClinVar and has no gnomAD entry. Prediction tools that call the variant benign include FoldX, Rosetta, Foldetta, premPS, SIFT, and FATHMM, while pathogenic calls come from REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta as Benign. Overall, the predictions are split, with a slight edge toward pathogenicity. The variant is therefore most likely pathogenic based on the current computational evidence, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.206376 | Structured | 0.396100 | Uncertain | 0.804 | 0.274 | 0.000 | -10.149 | Likely Pathogenic | 0.821 | Likely Pathogenic | Ambiguous | -0.42 | Likely Benign | 0.0 | 0.09 | Likely Benign | -0.17 | Likely Benign | -0.01 | Likely Benign | 0.513 | Likely Pathogenic | -2.70 | Deleterious | 0.963 | Probably Damaging | 0.549 | Possibly Damaging | 6.00 | Benign | 0.19 | Tolerated | 0.1972 | 0.2748 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.740A>G | Q247R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q247R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Uncertain predictions come from premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or folding stability result is missing or inconclusive. Based on the overall consensus of the majority of tools and the high‑accuracy methods, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.490133 | Structured | 0.283012 | Uncertain | 0.822 | 0.339 | 0.250 | -4.022 | Likely Benign | 0.442 | Ambiguous | Likely Benign | -0.38 | Likely Benign | 0.0 | 0.21 | Likely Benign | -0.09 | Likely Benign | -0.91 | Ambiguous | 0.471 | Likely Benign | 1.22 | Neutral | 0.948 | Possibly Damaging | 0.533 | Possibly Damaging | 5.91 | Benign | 1.00 | Tolerated | 0.1196 | 0.1324 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.850C>T | L284F 2D AIThe SynGAP1 missense variant L284F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely converge on a deleterious effect: REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity, while Rosetta, Foldetta, and premPS are inconclusive. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic; the SGM Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains uncertain. Taken together, the overwhelming majority of evidence indicates that the variant is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.094817 | Structured | 0.371601 | Uncertain | 0.950 | 0.255 | 0.000 | -11.656 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | 2.04 | Destabilizing | 1.9 | 1.05 | Ambiguous | 1.55 | Ambiguous | 0.62 | Ambiguous | 0.654 | Likely Pathogenic | -3.51 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.69 | Pathogenic | 0.01 | Affected | 0.0515 | 0.2779 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||
| c.98A>G | Q33R 2D AIThe SynGAP1 missense variant Q33R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also reports a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. The predictions do not contradict ClinVar status, as ClinVar contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.436712 | Uncertain | 0.342 | 0.860 | 0.375 | 0.561 | Likely Benign | 0.171 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -0.82 | Neutral | 0.084 | Benign | 0.046 | Benign | 4.20 | Benign | 0.00 | Affected | 0.1936 | 0.2773 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.1039A>C | T347P 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 T347P variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and FATHMM. Three tools (FoldX, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.268042 | Structured | 0.349915 | Uncertain | 0.951 | 0.434 | 0.000 | -9.323 | Likely Pathogenic | 0.417 | Ambiguous | Likely Benign | 0.55 | Ambiguous | 0.0 | 0.28 | Likely Benign | 0.42 | Likely Benign | 0.65 | Ambiguous | 0.236 | Likely Benign | -2.41 | Neutral | 0.627 | Possibly Damaging | 0.139 | Benign | 1.63 | Pathogenic | 0.12 | Tolerated | 0.2000 | 0.5350 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.1054A>C | T352P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant T352P has no ClinVar entry and is not reported in gnomAD. Prediction tools that converge on a benign outcome include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, FATHMM, and Rosetta. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM‑Consensus as Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Uncertain. Overall, the balance of evidence favors a benign interpretation; this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.137348 | Structured | 0.367886 | Uncertain | 0.926 | 0.329 | 0.000 | -3.562 | Likely Benign | 0.138 | Likely Benign | Likely Benign | 1.04 | Ambiguous | 0.1 | 2.57 | Destabilizing | 1.81 | Ambiguous | 0.51 | Ambiguous | 0.159 | Likely Benign | -2.31 | Neutral | 0.627 | Possibly Damaging | 0.196 | Benign | 1.72 | Pathogenic | 0.20 | Tolerated | 0.2244 | 0.5968 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1075A>C | T359P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T359P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. Uncertain or inconclusive results come from FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.281712 | Structured | 0.414952 | Uncertain | 0.939 | 0.480 | 0.250 | -6.884 | Likely Benign | 0.319 | Likely Benign | Likely Benign | 0.97 | Ambiguous | 0.1 | 1.27 | Ambiguous | 1.12 | Ambiguous | 0.68 | Ambiguous | 0.248 | Likely Benign | -2.36 | Neutral | 0.627 | Possibly Damaging | 0.091 | Benign | 1.78 | Pathogenic | 0.14 | Tolerated | 0.2352 | 0.5611 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1096A>C | T366P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T366P is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors leans toward a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign) all classify the substitution as tolerated. In contrast, polyPhen‑2 HumDiv, FATHMM, Rosetta, and the Foldetta stability analysis predict a damaging or pathogenic outcome. FoldX reports an uncertain effect and is therefore not considered evidence. High‑accuracy tools give mixed results: AlphaMissense‑Optimized and the SGM‑Consensus both indicate benign, whereas Foldetta predicts pathogenic. Overall, the majority of predictors (8 benign vs. 4 pathogenic) support a benign classification, and this is consistent with the lack of ClinVar evidence. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.441902 | Uncertain | 0.897 | 0.642 | 0.250 | -6.483 | Likely Benign | 0.226 | Likely Benign | Likely Benign | 1.75 | Ambiguous | 0.5 | 3.10 | Destabilizing | 2.43 | Destabilizing | 0.47 | Likely Benign | 0.150 | Likely Benign | -2.49 | Neutral | 0.627 | Possibly Damaging | 0.139 | Benign | 1.70 | Pathogenic | 0.24 | Tolerated | 0.2250 | 0.6251 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1105A>C | T369P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T369P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome, while FoldX and Foldetta are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as Likely Benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact for T369P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | -2.743 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 1.20 | Ambiguous | 2.1 | 0.18 | Likely Benign | 0.69 | Ambiguous | 0.17 | Likely Benign | 0.138 | Likely Benign | -2.09 | Neutral | 0.396 | Benign | 0.142 | Benign | 1.83 | Pathogenic | 0.16 | Tolerated | 0.2589 | 0.6046 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1222A>C | T408P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T408P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. FoldX, premPS, and Foldetta are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable; Foldetta remains uncertain. Overall, the majority of available predictions (six pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.161087 | Structured | 0.370935 | Uncertain | 0.907 | 0.239 | 0.000 | -10.384 | Likely Pathogenic | 0.230 | Likely Benign | Likely Benign | 1.08 | Ambiguous | 0.3 | 2.27 | Destabilizing | 1.68 | Ambiguous | 0.73 | Ambiguous | 0.323 | Likely Benign | -4.19 | Deleterious | 0.998 | Probably Damaging | 0.963 | Probably Damaging | 4.07 | Benign | 0.05 | Affected | 0.1985 | 0.5779 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.1273A>C | T425P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T425P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Consensus from multiple in silico predictors indicates a pathogenic effect: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as deleterious, while only FATHMM predicts a benign outcome; premPS is uncertain. High‑accuracy tools reinforce this view: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports it as Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts a pathogenic effect. Taken together, the overwhelming majority of evidence supports a pathogenic classification, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.401218 | Uncertain | 0.964 | 0.280 | 0.000 | -12.318 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 2.81 | Destabilizing | 0.3 | 6.49 | Destabilizing | 4.65 | Destabilizing | 0.77 | Ambiguous | 0.512 | Likely Pathogenic | -5.16 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.37 | Benign | 0.03 | Affected | 0.1815 | 0.3469 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1372A>C | T458P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T458P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are SIFT and FATHMM, while the remaining tools (REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, ESM1b, and the SGM‑Consensus) all predict a pathogenic impact; premPS is uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts a pathogenic effect. No prediction or folding stability result is missing or inconclusive. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.185198 | Structured | 0.294848 | Uncertain | 0.915 | 0.144 | 0.000 | -13.547 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 3.05 | Destabilizing | 0.2 | 5.36 | Destabilizing | 4.21 | Destabilizing | 0.78 | Ambiguous | 0.557 | Likely Pathogenic | -5.33 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.35 | Benign | 0.06 | Tolerated | 0.2098 | 0.5395 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1462A>C | T488P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T488P is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (gnomAD ID: none). Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while the remaining twelve tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict pathogenicity; premPS is uncertain and therefore not counted. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenicity. Based on the overwhelming consensus of these predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.206376 | Structured | 0.332663 | Uncertain | 0.928 | 0.233 | 0.125 | -13.432 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 4.40 | Destabilizing | 0.7 | 5.68 | Destabilizing | 5.04 | Destabilizing | 0.68 | Ambiguous | 0.505 | Likely Pathogenic | -5.70 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.24 | Benign | 0.00 | Affected | 0.1518 | 0.3557 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1471A>C | T491P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T491P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while premPS remains uncertain. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenic. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.064632 | Structured | 0.325158 | Uncertain | 0.929 | 0.188 | 0.125 | -13.603 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | 3.19 | Destabilizing | 0.6 | 4.32 | Destabilizing | 3.76 | Destabilizing | 0.96 | Ambiguous | 0.882 | Likely Pathogenic | -5.89 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.49 | Pathogenic | 0.00 | Affected | 0.2024 | 0.3759 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1594A>C | T532P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T532P is listed in ClinVar as Benign (ClinVar ID 1598909.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments—AlphaMissense‑Optimized, the SGM Consensus, and Foldetta (combining FoldX‑MD and Rosetta outputs)—all indicate a benign impact. No prediction or folding‑stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion is consistent with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.275179 | Structured | 0.021478 | Uncertain | 0.889 | 0.385 | 0.000 | Benign | 1 | -2.143 | Likely Benign | 0.061 | Likely Benign | Likely Benign | -0.30 | Likely Benign | 0.2 | 0.06 | Likely Benign | -0.12 | Likely Benign | 0.08 | Likely Benign | 0.201 | Likely Benign | -0.90 | Neutral | 0.005 | Benign | 0.008 | Benign | -1.28 | Pathogenic | 0.18 | Tolerated | 3.37 | 35 | 0.1850 | 0.3811 | 0 | -1 | -0.9 | -3.99 | 174.2 | 35.1 | 0.4 | 0.0 | 0.1 | 0.0 | X | Potentially Benign | Thr532 is located on an α-α loop between the two α-helices (res. Gly502-Tyr518 and Ala533-Val560) facing the membrane. In the WT simulations, the hydroxyl group of Thr532 occasionally forms hydrogen bonds with the backbone atoms of other loop residues without any specific interaction. In the variant simulations, the Pro532 residue swap does not cause structural changes. Although hydrophilic residues seem more favorable in the loop, the pyrrolidine side chain of proline is well suited for unstructured protein regions such as loops. However, due to its location at the SynGAP-membrane interface, the effect of the residue swap cannot be fully addressed using the SynGAP solvent-only simulations. | ||||||||||||||||
| c.1855A>C | T619P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant T619P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from FoldX and SIFT, while pathogenic predictions are made by REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. Two tools report uncertainty: premPS and AlphaMissense‑Optimized. High‑accuracy assessments further clarify the variant’s impact: AlphaMissense‑Optimized remains uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic effect; Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenicity. Overall, the majority of evidence points to a pathogenic effect for T619P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.219301 | Structured | 0.119723 | Uncertain | 0.929 | 0.237 | 0.000 | -12.879 | Likely Pathogenic | 0.933 | Likely Pathogenic | Ambiguous | 0.43 | Likely Benign | 0.2 | 4.81 | Destabilizing | 2.62 | Destabilizing | 0.82 | Ambiguous | 0.860 | Likely Pathogenic | -5.51 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.39 | Pathogenic | 0.09 | Tolerated | 0.1940 | 0.3806 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1864A>C | T622P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant T622P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess functional impact uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as pathogenic. No tool predicts a benign outcome; the only inconclusive result is premPS, which is listed as uncertain. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta stability outputs) also indicates pathogenic. Consequently, the variant is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.268042 | Structured | 0.071403 | Uncertain | 0.957 | 0.198 | 0.000 | -16.410 | Likely Pathogenic | 0.964 | Likely Pathogenic | Likely Pathogenic | 2.81 | Destabilizing | 0.4 | 8.90 | Destabilizing | 5.86 | Destabilizing | 0.78 | Ambiguous | 0.924 | Likely Pathogenic | -5.57 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.57 | Pathogenic | 0.01 | Affected | 0.1700 | 0.3876 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1870A>C | T624P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T624P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while premPS remains uncertain. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenic. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.137348 | Structured | 0.052894 | Uncertain | 0.962 | 0.217 | 0.000 | -15.681 | Likely Pathogenic | 0.972 | Likely Pathogenic | Likely Pathogenic | 3.64 | Destabilizing | 0.2 | 9.04 | Destabilizing | 6.34 | Destabilizing | 0.83 | Ambiguous | 0.914 | Likely Pathogenic | -5.94 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.55 | Pathogenic | 0.01 | Affected | 0.1500 | 0.3569 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1918A>C | T640P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T640P is not reported in ClinVar and is absent from gnomAD. In silico predictors uniformly favor a benign effect: REVEL, Rosetta, premPS, PROVEAN, polyPhen2_HumDiv, polyPhen2_HumVar, SIFT, FATHMM, AlphaMissense-Default, and AlphaMissense-Optimized all indicate benign. No tool predicts pathogenicity. FoldX and ESM1b were inconclusive. High‑accuracy methods corroborate this: AlphaMissense-Optimized is benign, the SGM Consensus (majority vote of AlphaMissense-Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta (combining FoldX‑MD and Rosetta) also predicts benign. Consequently, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for T640P. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.066181 | Structured | 0.137043 | Uncertain | 0.893 | 0.284 | 0.000 | -7.594 | In-Between | 0.077 | Likely Benign | Likely Benign | 0.98 | Ambiguous | 0.3 | -0.38 | Likely Benign | 0.30 | Likely Benign | 0.31 | Likely Benign | 0.313 | Likely Benign | -0.91 | Neutral | 0.004 | Benign | 0.002 | Benign | 3.47 | Benign | 0.14 | Tolerated | 0.2179 | 0.4722 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.2014A>C | T672P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T672P missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Uncertain results come from premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. No predictions are missing or inconclusive. Overall, the majority of evidence points to a pathogenic impact for T672P. This conclusion is not contradicted by ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.116183 | Structured | 0.102069 | Uncertain | 0.586 | 0.362 | 0.000 | -11.022 | Likely Pathogenic | 0.346 | Ambiguous | Likely Benign | 2.22 | Destabilizing | 0.2 | 5.54 | Destabilizing | 3.88 | Destabilizing | 0.55 | Ambiguous | 0.087 | Likely Benign | -4.20 | Deleterious | 0.968 | Probably Damaging | 0.824 | Possibly Damaging | 3.40 | Benign | 0.01 | Affected | 0.1988 | 0.5532 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.2020A>C | T674P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T674P missense change is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The high‑accuracy AlphaMissense‑Optimized predicts benign, while Foldetta predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. No other high‑accuracy tool provides a definitive call. Consequently, the evidence is evenly split between benign and pathogenic, leaving the variant’s clinical significance uncertain. This uncertainty does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.109297 | Uncertain | 0.521 | 0.349 | 0.000 | -8.661 | Likely Pathogenic | 0.109 | Likely Benign | Likely Benign | 0.91 | Ambiguous | 0.6 | 4.12 | Destabilizing | 2.52 | Destabilizing | 0.12 | Likely Benign | 0.143 | Likely Benign | -2.69 | Deleterious | 0.995 | Probably Damaging | 0.891 | Possibly Damaging | 3.50 | Benign | 0.16 | Tolerated | 0.2039 | 0.6103 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.2071A>C | T691P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T691P is listed in ClinVar (ID 648126.0) as Pathogenic and is not reported in gnomAD. Across the broad panel of in‑silico predictors, three tools (REVEL, SIFT, FATHMM) classify the change as benign, whereas the remaining 11 predictors (FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score) report it as pathogenic. High‑accuracy assessments further support a deleterious effect: the AlphaMissense‑Optimized model is inconclusive, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and the Foldetta stability analysis (combining FoldX‑MD and Rosetta outputs) is pathogenic. Taken together, the preponderance of evidence indicates that T691P is most likely pathogenic, which is consistent with its ClinVar classification and does not contradict the database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.271308 | Uncertain | 0.941 | 0.232 | 0.000 | Likely Pathogenic | 1 | -13.801 | Likely Pathogenic | 0.905 | Likely Pathogenic | Ambiguous | 5.04 | Destabilizing | 0.4 | 6.09 | Destabilizing | 5.57 | Destabilizing | 1.27 | Destabilizing | 0.214 | Likely Benign | -3.43 | Deleterious | 1.000 | Probably Damaging | 0.952 | Probably Damaging | 3.43 | Benign | 0.06 | Tolerated | 3.43 | 14 | 0.1466 | 0.4236 | 0 | -1 | -0.9 | -3.99 | 188.9 | 33.0 | 0.1 | 0.0 | -0.6 | 0.0 | X | X | Potentially Pathogenic | The hydroxyl side chain of Thr691, located in an α-helix (res. Leu696-Leu685), can form hydrogen bonds with the backbone carbonyl and the side chain guanidinium group of Arg687. This interaction facilitates the simultaneous formation of salt bridges between Arg687 and Glu688 on the same α-helix. Additionally, Thr691 occasionally interacts with the thioether side chain of Met409 in an anti-parallel β-sheet of the C2 domain (res. Ile411-Ala399), although this interaction is not consistently maintained throughout the WT simulations. In the variant simulations, the pyrrolidine side chain of Pro691 lacks hydrogen bond donors, making a similar setup impossible. Moreover, proline lacks a free amide group necessary for hydrogen bonding with the carbonyl group of Arg687, introducing a slight bend in the α-helix and compromising its integrity. | |||||||||||||||
| c.2167A>C | T723P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T723P is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FATHMM, and AlphaMissense‑Optimized. Tools that predict pathogenicity are SGM Consensus, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts likely pathogenic; Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. premPS is uncertain and does not influence the overall assessment. Overall, the majority of tools and the high‑accuracy methods support a pathogenic effect. Thus, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.476583 | Structured | 0.458243 | Uncertain | 0.945 | 0.447 | 0.375 | -9.231 | Likely Pathogenic | 0.741 | Likely Pathogenic | Likely Benign | 3.98 | Destabilizing | 0.1 | 6.10 | Destabilizing | 5.04 | Destabilizing | 0.54 | Ambiguous | 0.085 | Likely Benign | -2.51 | Deleterious | 0.995 | Probably Damaging | 0.929 | Probably Damaging | 3.49 | Benign | 0.04 | Affected | 0.1826 | 0.4406 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.2368A>C | T790P 2D AIThe SynGAP1 missense variant T790P has no ClinVar entry (ClinVar status: not reported) but is present in gnomAD (ID 6‑33442920‑A‑C). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of conventional tools (5 pathogenic vs 4 benign) lean toward a pathogenic interpretation, while the single high‑accuracy tool suggests benign. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this does not contradict the ClinVar status, which currently has no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.964893 | Disordered | 0.509280 | Binding | 0.385 | 0.896 | 0.875 | 6-33442920-A-C | -3.564 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.250 | Likely Benign | -3.55 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.25 | Pathogenic | 0.01 | Affected | 3.64 | 6 | 0.2147 | 0.4748 | -1 | 0 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||
| c.2464A>C | T822P 2D AIThe SynGAP1 missense variant T822P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for T822P. This conclusion is not contradicted by ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.651624 | Binding | 0.295 | 0.881 | 0.750 | -3.961 | Likely Benign | 0.737 | Likely Pathogenic | Likely Benign | 0.145 | Likely Benign | -2.29 | Neutral | 0.999 | Probably Damaging | 0.985 | Probably Damaging | 2.50 | Benign | 0.09 | Tolerated | 0.2437 | 0.5044 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2482A>C | T828P 2D AIThe SynGAP1 missense variant T828P is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain. The SGM Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to benign (2 benign vs. 1 pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) is unavailable. Overall, the preponderance of evidence points to a benign impact for T828P, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.631236 | Binding | 0.321 | 0.879 | 0.500 | -3.902 | Likely Benign | 0.466 | Ambiguous | Likely Benign | 0.280 | Likely Benign | -2.53 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.61 | Benign | 0.12 | Tolerated | 0.1898 | 0.4424 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.253A>C | T85P 2D AIThe SynGAP1 missense variant T85P is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments are mixed: AlphaMissense‑Optimized predicts pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields no clear majority and is therefore unavailable; Foldetta results are not provided. Overall, the majority of available predictions (five pathogenic vs. four benign) indicate a pathogenic effect. There is no ClinVar entry to contradict this assessment, so the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.542004 | Binding | 0.288 | 0.888 | 0.500 | -8.406 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.125 | Likely Benign | -2.28 | Neutral | 0.813 | Possibly Damaging | 0.053 | Benign | 3.80 | Benign | 0.00 | Affected | 0.1499 | 0.4485 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.2632A>C | T878P 2D AIThe SynGAP1 missense variant T878P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.622677 | Disordered | 0.628767 | Binding | 0.288 | 0.878 | 0.250 | -3.130 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.136 | Likely Benign | -1.41 | Neutral | 0.761 | Possibly Damaging | 0.478 | Possibly Damaging | 2.96 | Benign | 0.01 | Affected | 0.2026 | 0.5538 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2698A>C | T900P 2D AIThe SynGAP1 missense variant T900P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.465347 | Uncertain | 0.289 | 0.924 | 0.375 | -2.684 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.067 | Likely Benign | -0.40 | Neutral | 0.812 | Possibly Damaging | 0.473 | Possibly Damaging | 2.67 | Benign | 0.22 | Tolerated | 0.1863 | 0.4874 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2734A>C | T912P 2D AIThe SynGAP1 missense variant T912P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.740671 | Binding | 0.285 | 0.909 | 0.250 | -2.955 | Likely Benign | 0.111 | Likely Benign | Likely Benign | 0.087 | Likely Benign | -0.78 | Neutral | 0.217 | Benign | 0.121 | Benign | 4.00 | Benign | 0.00 | Affected | 0.1832 | 0.4608 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2737A>C | T913P 2D AIThe SynGAP1 missense variant T913P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. In contrast, polyPhen‑2 (both HumDiv and HumVar models) predict a pathogenic effect. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact for T913P, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.763517 | Binding | 0.339 | 0.899 | 0.375 | -2.029 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.176 | Likely Benign | -1.43 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.65 | Benign | 0.26 | Tolerated | 0.2082 | 0.5786 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2923A>C | T975P 2D AIThe SynGAP1 missense variant T975P has no ClinVar entry and is not reported in gnomAD. All evaluated in‑silico predictors classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports a benign effect, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence strongly supports a benign classification, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.871313 | Disordered | 0.969331 | Binding | 0.332 | 0.890 | 0.625 | -2.181 | Likely Benign | 0.071 | Likely Benign | Likely Benign | 0.286 | Likely Benign | -1.21 | Neutral | 0.000 | Benign | 0.002 | Benign | 4.14 | Benign | 0.06 | Tolerated | 0.2047 | 0.4758 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3043A>C | T1015P 2D AIThe SynGAP1 missense variant T1015P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. No Foldetta stability data are available, so it does not influence the assessment. Overall, the preponderance of evidence points to a benign impact for T1015P, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.928486 | Binding | 0.295 | 0.823 | 0.625 | -1.796 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.154 | Likely Benign | -0.15 | Neutral | 0.586 | Possibly Damaging | 0.223 | Benign | 2.53 | Benign | 0.18 | Tolerated | 0.2176 | 0.4504 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3052A>C | T1018P 2D AIThe SynGAP1 missense variant T1018P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. There is no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.849326 | Disordered | 0.959985 | Binding | 0.348 | 0.801 | 0.500 | -2.046 | Likely Benign | 0.101 | Likely Benign | Likely Benign | 0.218 | Likely Benign | -1.39 | Neutral | 0.586 | Possibly Damaging | 0.302 | Benign | 2.24 | Pathogenic | 0.02 | Affected | 0.2155 | 0.3960 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3112A>C | T1038P 2D AIThe SynGAP1 missense variant T1038P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (both HumDiv and HumVar) predict a pathogenic outcome. AlphaMissense‑Default remains uncertain, and the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.938133 | Disordered | 0.982911 | Binding | 0.279 | 0.794 | 0.625 | -2.196 | Likely Benign | 0.390 | Ambiguous | Likely Benign | 0.116 | Likely Benign | -1.10 | Neutral | 0.965 | Probably Damaging | 0.611 | Possibly Damaging | 2.66 | Benign | 0.26 | Tolerated | 0.1856 | 0.4680 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3229A>C | T1077P 2D AIThe SynGAP1 missense variant T1077P is reported in ClinVar as “Not submitted” and is not present in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and no Foldetta stability assessment is available. High‑accuracy evidence shows AlphaMissense‑Optimized classifying the variant as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicating a likely benign outcome, and Foldetta results are unavailable. Overall, the majority of predictions and the high‑accuracy consensus point to a benign impact. This conclusion is not contradicted by ClinVar status, which has no pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.988141 | Binding | 0.329 | 0.892 | 0.750 | -2.436 | Likely Benign | 0.345 | Ambiguous | Likely Benign | 0.164 | Likely Benign | -0.91 | Neutral | 0.970 | Probably Damaging | 0.787 | Possibly Damaging | 4.16 | Benign | 0.04 | Affected | 0.1876 | 0.4788 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3400A>C | T1134P 2D AIThe SynGAP1 missense variant T1134P is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores. No tool predicts pathogenicity. The high‑accuracy consensus methods likewise support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are unavailable. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.885302 | Disordered | 0.813034 | Binding | 0.335 | 0.885 | 0.875 | -2.993 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -0.57 | Neutral | 0.013 | Benign | 0.022 | Benign | 5.41 | Benign | 0.07 | Tolerated | 0.2001 | 0.5165 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3418A>C | T1140P 2D AIThe SynGAP1 missense variant T1140P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools, polyPhen‑2 HumDiv and HumVar, predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect for T1140P, and this conclusion is consistent with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.926919 | Disordered | 0.708094 | Binding | 0.293 | 0.854 | 1.000 | -1.921 | Likely Benign | 0.126 | Likely Benign | Likely Benign | 0.058 | Likely Benign | -1.49 | Neutral | 0.761 | Possibly Damaging | 0.478 | Possibly Damaging | 2.61 | Benign | 0.32 | Tolerated | 0.2044 | 0.3837 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3427A>C | T1143P 2D AIThe SynGAP1 missense variant T1143P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic effect. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.722918 | Binding | 0.275 | 0.837 | 1.000 | -2.307 | Likely Benign | 0.155 | Likely Benign | Likely Benign | 0.183 | Likely Benign | 0.80 | Neutral | 0.999 | Probably Damaging | 0.966 | Probably Damaging | 2.69 | Benign | 0.24 | Tolerated | 0.1818 | 0.4108 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3439A>C | T1147P 2D AIThe SynGAP1 missense variant T1147P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.831250 | Disordered | 0.746520 | Binding | 0.345 | 0.839 | 0.875 | -2.849 | Likely Benign | 0.072 | Likely Benign | Likely Benign | 0.425 | Likely Benign | -2.17 | Neutral | 0.000 | Benign | 0.002 | Benign | 5.41 | Benign | 0.02 | Affected | 0.1925 | 0.4490 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3460A>C | T1154P 2D AIThe SynGAP1 missense variant T1154P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that T1154P is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -2.513 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.368 | Likely Benign | -4.42 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.1667 | 0.3732 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3688A>C | T1230P 2D AIThe SynGAP1 missense variant T1230P is not reported in ClinVar and has no entries in gnomAD. Prediction tools largely agree on a deleterious effect: benign predictions are limited to FATHMM, whereas the remaining methods—REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta results are unavailable, so no additional stability evidence is provided. Overall, the consensus of available predictions points to a pathogenic effect for T1230P, with no conflict from ClinVar status (which is currently unreported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.570702 | Disordered | 0.486342 | Uncertain | 0.845 | 0.543 | 0.250 | -13.200 | Likely Pathogenic | 0.979 | Likely Pathogenic | Likely Pathogenic | 0.588 | Likely Pathogenic | -2.86 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 5.54 | Benign | 0.01 | Affected | 0.1529 | 0.3710 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||
| c.3913A>C | T1305P 2D AIThe SynGAP1 missense variant T1305P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.791621 | Disordered | 0.894658 | Binding | 0.390 | 0.873 | 0.875 | -1.882 | Likely Benign | 0.058 | Likely Benign | Likely Benign | 0.204 | Likely Benign | -1.41 | Neutral | 0.027 | Benign | 0.114 | Benign | 2.86 | Benign | 0.03 | Affected | 0.2157 | 0.5386 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3928A>C | T1310P 2D AIThe SynGAP1 missense variant T1310P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta results are unavailable, so no additional stability evidence is present. Overall, the consensus of available predictions indicates that T1310P is most likely benign, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.959076 | Binding | 0.398 | 0.904 | 0.750 | -1.807 | Likely Benign | 0.058 | Likely Benign | Likely Benign | 0.209 | Likely Benign | -1.48 | Neutral | 0.594 | Possibly Damaging | 0.298 | Benign | 2.75 | Benign | 0.04 | Affected | 0.1667 | 0.3604 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3994A>C | T1332P 2D AIThe SynGAP1 missense variant T1332P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is tied 2‑2 and thus unavailable, and Foldetta results are not provided. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any existing ClinVar status because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | -3.404 | Likely Benign | 0.918 | Likely Pathogenic | Ambiguous | 0.238 | Likely Benign | -3.12 | Deleterious | 0.994 | Probably Damaging | 0.981 | Probably Damaging | 2.95 | Benign | 0.00 | Affected | 0.2185 | 0.5308 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.4018A>C | T1340P 2D AIThe SynGAP1 missense variant T1340P is catalogued in gnomAD (ID 6‑33451892‑A‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes, while only SIFT predicts a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that T1340P is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.977899 | Binding | 0.444 | 0.697 | 0.750 | 6-33451892-A-C | -2.681 | Likely Benign | 0.102 | Likely Benign | Likely Benign | 0.190 | Likely Benign | -1.81 | Neutral | 0.334 | Benign | 0.099 | Benign | 4.08 | Benign | 0.01 | Affected | 3.77 | 5 | 0.2268 | 0.4983 | -1 | 0 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||
| c.430A>C | T144P 2D AIThe SynGAP1 missense variant T144P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict pathogenicity are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely pathogenic, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of predictions and the consensus call indicate a pathogenic effect. This conclusion is not contradicted by ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | -11.920 | Likely Pathogenic | 0.775 | Likely Pathogenic | Likely Benign | 0.159 | Likely Benign | -2.66 | Deleterious | 0.000 | Benign | 0.000 | Benign | 3.76 | Benign | 0.00 | Affected | 0.2508 | 0.5271 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.457A>C | T153P 2D AIThe SynGAP1 missense variant T153P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it contains two benign and two uncertain calls, and Foldetta data are unavailable. Overall, the balance of evidence leans toward a benign interpretation. This conclusion does not conflict with ClinVar, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.501700 | Disordered | 0.502105 | Binding | 0.297 | 0.818 | 0.625 | -7.092 | In-Between | 0.374 | Ambiguous | Likely Benign | 0.269 | Likely Benign | -1.67 | Neutral | 0.983 | Probably Damaging | 0.725 | Possibly Damaging | 4.09 | Benign | 0.03 | Affected | 0.2270 | 0.3660 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.667A>C | T223P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T223P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include FoldX, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise SGM‑Consensus (Likely Pathogenic), REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic, SGM‑Consensus as Likely Pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as Uncertain. No other high‑accuracy tools provide a definitive prediction. Overall, the majority of evidence points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.070400 | Structured | 0.382605 | Uncertain | 0.867 | 0.316 | 0.125 | -13.707 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | 0.42 | Likely Benign | 0.3 | 3.45 | Destabilizing | 1.94 | Ambiguous | 0.67 | Ambiguous | 0.898 | Likely Pathogenic | -4.54 | Deleterious | 0.838 | Possibly Damaging | 0.367 | Benign | 5.72 | Benign | 0.01 | Affected | 0.1464 | 0.3728 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.670A>C | T224P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T224P is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are SIFT and FATHMM, while the remaining tools (REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b) all predict a pathogenic impact; premPS is uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts pathogenic. No prediction or folding stability result is missing or inconclusive. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.104810 | Structured | 0.360921 | Uncertain | 0.848 | 0.315 | 0.125 | -11.812 | Likely Pathogenic | 0.960 | Likely Pathogenic | Likely Pathogenic | 3.63 | Destabilizing | 0.4 | 2.68 | Destabilizing | 3.16 | Destabilizing | 0.57 | Ambiguous | 0.765 | Likely Pathogenic | -3.65 | Deleterious | 0.971 | Probably Damaging | 0.543 | Possibly Damaging | 5.56 | Benign | 0.23 | Tolerated | 0.2355 | 0.6063 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.682A>C | T228P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T228P is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include FATHMM, Rosetta, and premPS, whereas the majority of tools predict a pathogenic impact: REVEL, SIFT, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized is inconclusive, but the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also inconclusive. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.104810 | Structured | 0.321733 | Uncertain | 0.829 | 0.316 | 0.125 | -8.559 | Likely Pathogenic | 0.939 | Likely Pathogenic | Ambiguous | 1.65 | Ambiguous | 0.6 | 0.47 | Likely Benign | 1.06 | Ambiguous | 0.21 | Likely Benign | 0.690 | Likely Pathogenic | -3.54 | Deleterious | 0.984 | Probably Damaging | 0.690 | Possibly Damaging | 5.64 | Benign | 0.02 | Affected | 0.2207 | 0.6360 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.880A>C | T294P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T294P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX (Uncertain), Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity. No tool reports a benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is pathogenic; SGM‑Consensus, derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.328603 | Structured | 0.316932 | Uncertain | 0.919 | 0.267 | 0.125 | -17.477 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 1.97 | Ambiguous | 0.3 | 5.18 | Destabilizing | 3.58 | Destabilizing | 1.15 | Destabilizing | 0.741 | Likely Pathogenic | -5.52 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -0.19 | Pathogenic | 0.01 | Affected | 0.1906 | 0.4479 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.883A>C | T295P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T295P is not reported in ClinVar and has no entries in gnomAD. Prediction tools that indicate a benign effect include FoldX and AlphaMissense‑Optimized, whereas the remaining evaluated algorithms (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic outcome. High‑accuracy assessments further show that AlphaMissense‑Optimized classifies the variant as benign, the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labels it as likely pathogenic, and Foldetta reports an uncertain stability change. With the majority of evidence pointing to deleterious effects, the variant is most likely pathogenic, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.401658 | Structured | 0.295548 | Uncertain | 0.881 | 0.288 | 0.125 | -9.941 | Likely Pathogenic | 0.688 | Likely Pathogenic | Likely Benign | 0.26 | Likely Benign | 0.2 | 3.30 | Destabilizing | 1.78 | Ambiguous | 0.60 | Ambiguous | 0.531 | Likely Pathogenic | -4.65 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.90 | Pathogenic | 0.02 | Affected | 0.2442 | 0.5327 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.913A>C | T305P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T305P missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM; premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of tools (7 pathogenic vs. 5 benign) lean toward a pathogenic impact, and this assessment does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current predictive evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.359901 | Structured | 0.299706 | Uncertain | 0.872 | 0.274 | 0.125 | -5.203 | Likely Benign | 0.177 | Likely Benign | Likely Benign | 3.64 | Destabilizing | 1.1 | 6.02 | Destabilizing | 4.83 | Destabilizing | 0.71 | Ambiguous | 0.293 | Likely Benign | -2.83 | Deleterious | 0.997 | Probably Damaging | 0.929 | Probably Damaging | 1.72 | Pathogenic | 0.06 | Tolerated | 0.2206 | 0.5333 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.94A>C | T32P 2D AIThe SynGAP1 missense variant T32P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. In contrast, two tools—polyPhen‑2 HumDiv and SIFT—classify the change as pathogenic. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; no Foldetta stability data are available. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.437154 | Uncertain | 0.349 | 0.879 | 0.375 | -2.958 | Likely Benign | 0.055 | Likely Benign | Likely Benign | 0.102 | Likely Benign | -0.94 | Neutral | 0.604 | Possibly Damaging | 0.185 | Benign | 4.14 | Benign | 0.00 | Affected | 0.2327 | 0.5818 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.109T>C | S37P 2D  AIThe SynGAP1 missense variant S37P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for S37P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.433492 | Uncertain | 0.317 | 0.806 | 0.500 | -3.788 | Likely Benign | 0.149 | Likely Benign | Likely Benign | 0.156 | Likely Benign | -1.29 | Neutral | 0.676 | Possibly Damaging | 0.693 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.2657 | 0.5327 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.10T>C | S4P 2D AIThe SynGAP1 missense variant S4P is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools largely support a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, two tools—polyPhen‑2 HumDiv and SIFT—predict a pathogenic impact. High‑accuracy methods give a consistent benign signal: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.608892 | Disordered | 0.547364 | Binding | 0.390 | 0.924 | 0.750 | -4.131 | Likely Benign | 0.114 | Likely Benign | Likely Benign | 0.153 | Likely Benign | -0.33 | Neutral | 0.676 | Possibly Damaging | 0.307 | Benign | 4.12 | Benign | 0.00 | Affected | 0.2043 | 0.6112 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.1120T>C | S374P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S374P is reported in gnomAD (6‑33438025‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and Rosetta; FoldX and Foldetta are inconclusive. The high‑accuracy consensus (SGM‑Consensus) is “Likely Benign,” derived from the unanimous benign calls of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN. AlphaMissense‑Optimized also predicts benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact. There is no ClinVar status to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.642678 | Disordered | 0.428948 | Uncertain | 0.333 | 0.812 | 0.625 | 6-33438025-T-C | 1 | 7.85e-7 | -4.849 | Likely Benign | 0.125 | Likely Benign | Likely Benign | 0.66 | Ambiguous | 0.6 | 2.22 | Destabilizing | 1.44 | Ambiguous | 0.34 | Likely Benign | 0.388 | Likely Benign | -0.89 | Neutral | 0.396 | Benign | 0.099 | Benign | 5.30 | Benign | 0.02 | Affected | 4.32 | 13 | 0.3012 | 0.6813 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||
| c.1135T>C | S379P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S379P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, Rosetta, and Foldetta; FoldX is uncertain. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, whereas Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. Overall, the majority of evidence—including the high‑accuracy benign predictions—suggests that the variant is most likely benign. This conclusion is not contradicted by ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.728858 | Disordered | 0.433206 | Uncertain | 0.327 | 0.931 | 0.625 | -5.007 | Likely Benign | 0.118 | Likely Benign | Likely Benign | 1.10 | Ambiguous | 0.8 | 2.92 | Destabilizing | 2.01 | Destabilizing | 0.17 | Likely Benign | 0.430 | Likely Benign | -0.41 | Neutral | 0.808 | Possibly Damaging | 0.212 | Benign | 3.83 | Benign | 0.10 | Tolerated | 0.3035 | 0.6594 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.1153T>C | S385P 2D 3DClick to see structure in 3D Viewer AISynGAP1 variant S385P is listed in ClinVar with an uncertain significance and is present in gnomAD (variant ID 6-33438058‑T‑C). Prediction tools that classify the variant as benign include REVEL, Foldetta, premPS, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Predictions from FoldX and Rosetta are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of computational evidence supports a benign effect, which is consistent with the ClinVar uncertain status and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.733139 | Disordered | 0.425480 | Uncertain | 0.341 | 0.925 | 0.750 | Uncertain | 1 | 6-33438058-T-C | -5.431 | Likely Benign | 0.123 | Likely Benign | Likely Benign | 0.91 | Ambiguous | 0.6 | -0.90 | Ambiguous | 0.01 | Likely Benign | 0.19 | Likely Benign | 0.385 | Likely Benign | -0.26 | Neutral | 0.676 | Possibly Damaging | 0.693 | Possibly Damaging | 4.63 | Benign | 0.04 | Affected | 4.32 | 3 | 0.2925 | 0.6805 | 1 | -1 | -0.8 | 10.04 | 210.3 | 18.5 | 1.8 | 0.9 | 0.3 | 0.0 | Uncertain | Ser385 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because the Ω loop is assumed to directly interact with the membrane, it moves arbitrarily throughout the WT solvent simulations. The Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play major roles in protein functions that require flexibility, and so they are rich in glycine residues, prolines, and, to a lesser extent, small hydrophilic residues to ensure maximum flexibility. Thus, the variant’s Pro385 is potentially tolerated in the Ω loop. However, since the effects on Gly-rich Ω loop dynamics can only be well studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||||
| c.1165T>C | S389P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S389P is listed in gnomAD (variant ID 6‑33438070‑T‑C) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, Foldetta, premPS, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts benign. No prediction or stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.703578 | Disordered | 0.417444 | Uncertain | 0.306 | 0.803 | 0.875 | 6-33438070-T-C | -5.286 | Likely Benign | 0.130 | Likely Benign | Likely Benign | 1.85 | Ambiguous | 2.0 | -0.99 | Ambiguous | 0.43 | Likely Benign | 0.16 | Likely Benign | 0.460 | Likely Benign | -0.44 | Neutral | 0.676 | Possibly Damaging | 0.693 | Possibly Damaging | 5.05 | Benign | 0.01 | Affected | 4.32 | 8 | 0.3098 | 0.7002 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||
| c.1396T>C | S466P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S466P is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that assess pathogenicity unanimously classify the variant as deleterious: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect; the only inconclusive result is from premPS (Uncertain). High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts Pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts Pathogenic. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.311707 | Structured | 0.322353 | Uncertain | 0.933 | 0.227 | 0.000 | -13.107 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 2.67 | Destabilizing | 0.2 | 2.83 | Destabilizing | 2.75 | Destabilizing | 0.95 | Ambiguous | 0.811 | Likely Pathogenic | -3.02 | Deleterious | 0.995 | Probably Damaging | 0.986 | Probably Damaging | -1.55 | Pathogenic | 0.04 | Affected | 0.1617 | 0.4915 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.1411T>C | S471P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S471P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only polyPhen‑2 HumVar, whereas the remaining evaluated algorithms (REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No predictions are missing or inconclusive. Based on the overwhelming agreement among the majority of tools, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.305330 | Structured | 0.355411 | Uncertain | 0.888 | 0.261 | 0.000 | -12.379 | Likely Pathogenic | 0.980 | Likely Pathogenic | Likely Pathogenic | 3.65 | Destabilizing | 0.2 | 9.24 | Destabilizing | 6.45 | Destabilizing | 0.84 | Ambiguous | 0.530 | Likely Pathogenic | -4.03 | Deleterious | 0.552 | Possibly Damaging | 0.141 | Benign | -1.31 | Pathogenic | 0.05 | Affected | 0.2007 | 0.4769 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.154T>C | S52P 2D AIThe SynGAP1 missense variant S52P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default all predict a pathogenic outcome. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign votes); and Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact for S52P. This conclusion does not contradict ClinVar, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.291804 | Structured | 0.457753 | Uncertain | 0.499 | 0.677 | 0.000 | -10.007 | Likely Pathogenic | 0.797 | Likely Pathogenic | Ambiguous | 0.144 | Likely Benign | -1.28 | Neutral | 0.676 | Possibly Damaging | 0.693 | Possibly Damaging | 4.07 | Benign | 0.00 | Affected | 0.2276 | 0.5909 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||||
| c.1558T>C | S520P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S520P is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that assess pathogenicity all converge on a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report a pathogenic outcome, while premPS remains inconclusive. No tool predicts a benign effect. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, is pathogenic. Overall, the evidence strongly supports a pathogenic impact for S520P, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.094817 | Structured | 0.084894 | Uncertain | 0.887 | 0.337 | 0.000 | Uncertain | 1 | -12.707 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 3.72 | Destabilizing | 0.8 | 8.86 | Destabilizing | 6.29 | Destabilizing | 0.83 | Ambiguous | 0.855 | Likely Pathogenic | -4.57 | Deleterious | 0.997 | Probably Damaging | 0.986 | Probably Damaging | -1.32 | Pathogenic | 0.01 | Affected | 0.2154 | 0.4776 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||
| c.1600T>C | S534P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S534P is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33438843‑T‑C). Functional prediction tools that report a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The high‑accuracy assessments are consistent with a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts benign. Based on the aggregate predictions, the variant is most likely benign, which does not contradict the ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.167087 | Structured | 0.032173 | Uncertain | 0.860 | 0.362 | 0.000 | Uncertain | 1 | 6-33438843-T-C | 3 | 1.86e-6 | -5.056 | Likely Benign | 0.265 | Likely Benign | Likely Benign | -0.40 | Likely Benign | 0.2 | 0.35 | Likely Benign | -0.03 | Likely Benign | 0.47 | Likely Benign | 0.203 | Likely Benign | -3.81 | Deleterious | 0.993 | Probably Damaging | 0.993 | Probably Damaging | 3.32 | Benign | 0.05 | Affected | 3.37 | 35 | 0.2071 | 0.4650 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||
| c.1669T>C | S557P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S557P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity all converge on a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a pathogenic or likely pathogenic outcome. No tool in the dataset predicts a benign effect. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, also labels it pathogenic. Based on the uniform predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.028107 | Structured | 0.010261 | Uncertain | 0.924 | 0.215 | 0.000 | -15.383 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 6.91 | Destabilizing | 0.4 | 8.79 | Destabilizing | 7.85 | Destabilizing | 1.09 | Destabilizing | 0.934 | Likely Pathogenic | -4.60 | Deleterious | 0.997 | Probably Damaging | 0.986 | Probably Damaging | -1.76 | Pathogenic | 0.01 | Affected | 0.2222 | 0.6239 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.1711T>C | S571P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S571P is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while premPS remains inconclusive. High‑accuracy assessments corroborate this trend: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenic. Consequently, the aggregate evidence strongly supports a pathogenic effect for S571P, and this conclusion does not conflict with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.069024 | Structured | 0.045569 | Uncertain | 0.928 | 0.270 | 0.000 | Uncertain | 1 | -14.701 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 3.18 | Destabilizing | 0.2 | 4.89 | Destabilizing | 4.04 | Destabilizing | 0.87 | Ambiguous | 0.814 | Likely Pathogenic | -4.68 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | -1.30 | Pathogenic | 0.02 | Affected | 0.2195 | 0.3760 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||
| c.1774T>C | S592P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S592P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include only SIFT, whereas the remaining tools—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. No predictions are inconclusive or missing. Based on the overwhelming agreement among pathogenic predictors and the absence of benign evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.012270 | Structured | 0.100070 | Uncertain | 0.913 | 0.182 | 0.000 | -14.621 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 4.26 | Destabilizing | 0.1 | 2.36 | Destabilizing | 3.31 | Destabilizing | 1.24 | Destabilizing | 0.909 | Likely Pathogenic | -4.77 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | -1.29 | Pathogenic | 0.11 | Tolerated | 0.2412 | 0.4259 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.1810T>C | S604P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S604P is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include premPS and FATHMM, whereas the remaining tools—REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict it to be pathogenic. High‑accuracy methods give consistent results: AlphaMissense‑Optimized indicates pathogenicity; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also labels the variant as likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a destabilizing, pathogenic effect. Taken together, the overwhelming majority of computational evidence supports a pathogenic classification for S604P, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.010926 | Structured | 0.192527 | Uncertain | 0.911 | 0.195 | 0.000 | -10.953 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 3.76 | Destabilizing | 0.3 | 8.40 | Destabilizing | 6.08 | Destabilizing | 0.04 | Likely Benign | 0.512 | Likely Pathogenic | -4.98 | Deleterious | 0.997 | Probably Damaging | 0.986 | Probably Damaging | 3.10 | Benign | 0.01 | Affected | 0.2524 | 0.4829 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.1858T>C | S620P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S620P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity uniformly favor a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity, while premPS remains uncertain. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is pathogenic. Consequently, the variant is most likely pathogenic, and this prediction is consistent with the absence of a ClinVar entry (no contradiction). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.139895 | Structured | 0.100377 | Uncertain | 0.936 | 0.219 | 0.000 | -12.208 | Likely Pathogenic | 0.979 | Likely Pathogenic | Likely Pathogenic | 4.89 | Destabilizing | 0.5 | 12.23 | Destabilizing | 8.56 | Destabilizing | 0.73 | Ambiguous | 0.834 | Likely Pathogenic | -3.62 | Deleterious | 0.998 | Probably Damaging | 0.993 | Probably Damaging | -1.35 | Pathogenic | 0.02 | Affected | 0.2147 | 0.4776 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.1909T>C | S637P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant S637P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, premPS, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls arise from FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments give a pathogenic signal: the SGM Consensus predicts likely pathogenic, Foldetta predicts destabilizing pathogenic effects, whereas AlphaMissense‑Optimized remains uncertain. Overall, the balance of evidence favors a pathogenic interpretation, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.076542 | Structured | 0.083482 | Uncertain | 0.920 | 0.253 | 0.000 | -11.455 | Likely Pathogenic | 0.793 | Likely Pathogenic | Ambiguous | 6.73 | Destabilizing | 0.1 | 6.36 | Destabilizing | 6.55 | Destabilizing | 0.44 | Likely Benign | 0.192 | Likely Benign | -3.12 | Deleterious | 0.946 | Possibly Damaging | 0.360 | Benign | 3.36 | Benign | 0.03 | Affected | 0.2374 | 0.4094 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.1921T>C | S641P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S641P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and SIFT. The remaining tools (FoldX, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward benign, and Foldetta as uncertain. Overall, the majority of reliable predictions indicate a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.125101 | Structured | 0.157322 | Uncertain | 0.786 | 0.270 | 0.000 | -6.907 | Likely Benign | 0.378 | Ambiguous | Likely Benign | 1.73 | Ambiguous | 1.3 | 0.07 | Likely Benign | 0.90 | Ambiguous | 0.59 | Ambiguous | 0.205 | Likely Benign | -3.05 | Deleterious | 0.000 | Benign | 0.000 | Benign | 3.34 | Benign | 0.04 | Affected | 0.2289 | 0.5791 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||
| c.1975T>C | S659P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S659P is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are SGM‑Consensus, Rosetta, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. Tools with uncertain or inconclusive results—FoldX, premPS, and Foldetta—are treated as unavailable. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized reports a benign change, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic, and Foldetta’s stability assessment is uncertain. Overall, the balance of evidence leans toward a pathogenic effect, but the presence of a strong benign prediction from AlphaMissense‑Optimized and the lack of ClinVar annotation means the variant’s clinical significance remains uncertain and does not contradict existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.154597 | Uncertain | 0.954 | 0.283 | 0.000 | -10.461 | Likely Pathogenic | 0.609 | Likely Pathogenic | Likely Benign | -0.73 | Ambiguous | 0.3 | 2.98 | Destabilizing | 1.13 | Ambiguous | 0.73 | Ambiguous | 0.224 | Likely Benign | -3.26 | Deleterious | 0.932 | Possibly Damaging | 0.245 | Benign | 3.39 | Benign | 0.18 | Tolerated | 0.2207 | 0.5553 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.19T>C | S7P 2D AIThe SynGAP1 missense variant S7P is reported in gnomAD (ID 6‑33420283‑T‑C) but has no ClinVar entry. In silico prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority‑vote) is benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools, including the high‑accuracy methods, points to a benign effect. This prediction is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.548467 | Binding | 0.386 | 0.922 | 0.750 | 6-33420283-T-C | -3.798 | Likely Benign | 0.111 | Likely Benign | Likely Benign | 0.210 | Likely Benign | -0.23 | Neutral | 0.267 | Benign | 0.029 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2474 | 0.4925 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||
| c.2002T>C | S668P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S668P has no ClinVar entry and is not reported in gnomAD. Prediction tools largely agree on a deleterious effect: FATHMM is the only benign predictor, while the remaining 13 tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) all classify it as pathogenic or likely pathogenic. The premPS score is uncertain and therefore not used as evidence. High‑accuracy methods give consistent results: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports likely pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No prediction or folding stability result is missing or inconclusive. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.247041 | Structured | 0.084935 | Uncertain | 0.922 | 0.370 | 0.000 | -13.452 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 6.40 | Destabilizing | 1.1 | 10.90 | Destabilizing | 8.65 | Destabilizing | 0.61 | Ambiguous | 0.612 | Likely Pathogenic | -4.82 | Deleterious | 0.998 | Probably Damaging | 0.790 | Possibly Damaging | 3.16 | Benign | 0.02 | Affected | 0.1979 | 0.5137 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.2068T>C | S690P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S690P is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that indicate a benign effect are REVEL and FATHMM, whereas the remaining tools (FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) all predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, also classifies the variant as pathogenic. Overall, the preponderance of evidence from multiple independent predictors indicates that the variant is most likely pathogenic, a conclusion that does not contradict the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.055536 | Structured | 0.247926 | Uncertain | 0.944 | 0.253 | 0.000 | Uncertain | 1 | -14.568 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 4.84 | Destabilizing | 0.3 | 4.40 | Destabilizing | 4.62 | Destabilizing | 1.42 | Destabilizing | 0.431 | Likely Benign | -4.77 | Deleterious | 0.998 | Probably Damaging | 0.790 | Possibly Damaging | 3.44 | Benign | 0.01 | Affected | 3.42 | 17 | 0.1787 | 0.4050 | 1 | -1 | -0.8 | 10.04 | 207.5 | 15.1 | 0.1 | 0.0 | -0.1 | 0.2 | X | X | Potentially Pathogenic | The hydroxyl side chain of Ser690, located in an α-helix (res. Leu696-Leu685), forms a hydrogen bond with the backbone carbonyl group of Ser410 in an anti-parallel β-sheet of the C2 domain (res. Ile411-Ala399). In the variant simulations, the pyrrolidine side chain of Pro690 cannot form hydrogen bonds with the C2 domain residue, resulting in the loss of this inter-domain connection. Additionally, prolines lack a free amide group necessary for hydrogen bonding with the carbonyl group of Gly686, introducing a slight bend in the α-helix and compromising its integrity. | |||||||||||||||
| c.2254T>C | S752P 2D AIThe SynGAP1 missense variant S752P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact for S752P, and this conclusion does not contradict the ClinVar status, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.604312 | Disordered | 0.690594 | Binding | 0.365 | 0.877 | 0.625 | -3.491 | Likely Benign | 0.158 | Likely Benign | Likely Benign | 0.183 | Likely Benign | -1.09 | Neutral | 0.998 | Probably Damaging | 0.912 | Probably Damaging | 1.51 | Pathogenic | 0.02 | Affected | 0.2288 | 0.5882 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.226T>C | S76P 2D AIThe SynGAP1 missense variant S76P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect for S76P, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.517562 | Disordered | 0.444487 | Uncertain | 0.279 | 0.826 | 0.500 | -3.833 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.058 | Likely Benign | -1.64 | Neutral | 0.909 | Possibly Damaging | 0.665 | Possibly Damaging | 3.77 | Benign | 0.00 | Affected | 0.1790 | 0.4138 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2296T>C | S766P 2D AIThe SynGAP1 missense variant S766P is reported in gnomAD (ID 6‑33442454‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while SIFT uniquely predicts it as pathogenic. The remaining tools, ESM1b and AlphaMissense‑Default, are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.405110 | Structured | 0.923125 | Binding | 0.338 | 0.874 | 0.250 | 6-33442454-T-C | 1 | 1.28e-6 | -7.343 | In-Between | 0.374 | Ambiguous | Likely Benign | 0.193 | Likely Benign | -0.91 | Neutral | 0.006 | Benign | 0.013 | Benign | 4.07 | Benign | 0.01 | Affected | 3.64 | 6 | 0.2216 | 0.5862 | -1 | 1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||
| c.2311T>C | S771P 2D AIThe SynGAP1 missense variant S771P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.922503 | Binding | 0.306 | 0.883 | 0.250 | -5.045 | Likely Benign | 0.112 | Likely Benign | Likely Benign | 0.180 | Likely Benign | -1.32 | Neutral | 0.901 | Possibly Damaging | 0.692 | Possibly Damaging | 4.03 | Benign | 0.19 | Tolerated | 0.2194 | 0.5515 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2338T>C | S780P 2D AIThe SynGAP1 missense variant S780P is reported in gnomAD (ID 6‑33442890‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.599170 | Disordered | 0.812415 | Binding | 0.283 | 0.883 | 0.500 | 6-33442890-T-C | 1 | 6.22e-7 | -6.055 | Likely Benign | 0.234 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -1.11 | Neutral | 0.995 | Probably Damaging | 0.892 | Possibly Damaging | 2.64 | Benign | 0.30 | Tolerated | 3.64 | 6 | 0.2086 | 0.6171 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.2362T>C | S788P 2D AIThe SynGAP1 missense variant S788P is catalogued in gnomAD (ID 6‑33442914‑T‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The AlphaMissense‑Default score is uncertain. A consensus derived from the SGM framework (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) is unavailable for this residue. Taken together, the preponderance of evidence from standard predictors and the SGM consensus points to a likely pathogenic effect, which does not contradict any existing ClinVar annotation because none is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.956248 | Disordered | 0.573557 | Binding | 0.349 | 0.895 | 0.750 | 6-33442914-T-C | -1.857 | Likely Benign | 0.435 | Ambiguous | Likely Benign | 0.236 | Likely Benign | -3.91 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 1.51 | Pathogenic | 0.01 | Affected | 3.64 | 6 | 0.2338 | 0.5537 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||
| c.2446T>C | S816P 2D AIThe SynGAP1 missense variant S816P is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign, while the majority of other predictors (polyPhen‑2 HumDiv and HumVar) suggest pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign and the SGM‑Consensus likewise indicates Likely Benign; Foldetta data are unavailable. Overall, the preponderance of evidence points to a benign effect for S816P, and this conclusion is not in conflict with ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.476583 | Structured | 0.747189 | Binding | 0.347 | 0.898 | 0.375 | -3.662 | Likely Benign | 0.510 | Ambiguous | Likely Benign | 0.160 | Likely Benign | -0.26 | Neutral | 0.999 | Probably Damaging | 0.966 | Probably Damaging | 2.66 | Benign | 0.45 | Tolerated | 0.2298 | 0.5102 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2449T>C | S817P 2D AIThe SynGAP1 missense variant S817P is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Overall, the majority of tools and the consensus score indicate a pathogenic effect, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.490133 | Structured | 0.727082 | Binding | 0.314 | 0.901 | 0.625 | -4.633 | Likely Benign | 0.724 | Likely Pathogenic | Likely Benign | 0.201 | Likely Benign | -3.26 | Deleterious | 0.999 | Probably Damaging | 0.966 | Probably Damaging | 2.39 | Pathogenic | 0.00 | Affected | 0.2272 | 0.6023 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2473T>C | S825P 2D AIThe SynGAP1 missense variant S825P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. Foldetta results are not available for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that S825P is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.675549 | Disordered | 0.618614 | Binding | 0.384 | 0.886 | 0.750 | -3.227 | Likely Benign | 0.959 | Likely Pathogenic | Likely Pathogenic | 0.285 | Likely Benign | -3.12 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.94 | Pathogenic | 0.02 | Affected | 0.2060 | 0.5998 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2524T>C | S842P 2D AIThe SynGAP1 missense variant S842P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that assess pathogenicity largely agree: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all predict a deleterious effect. Only REVEL classifies the variant as benign. High‑accuracy assessments further support a pathogenic interpretation: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus indicates “Likely Pathogenic.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the consensus of the majority of tools, including the high‑accuracy predictors, indicates that S842P is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.604312 | Disordered | 0.617281 | Binding | 0.274 | 0.861 | 0.250 | -13.890 | Likely Pathogenic | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.309 | Likely Benign | -3.43 | Deleterious | 0.995 | Probably Damaging | 0.892 | Possibly Damaging | 1.99 | Pathogenic | 0.00 | Affected | 0.1494 | 0.5342 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2581T>C | S861P 2D AIThe SynGAP1 missense variant S861P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). In contrast, polyPhen‑2 (both HumDiv and HumVar models) predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect for S861P, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.557691 | Disordered | 0.540903 | Binding | 0.285 | 0.797 | 0.250 | -4.256 | Likely Benign | 0.107 | Likely Benign | Likely Benign | 0.116 | Likely Benign | -1.65 | Neutral | 0.960 | Probably Damaging | 0.761 | Possibly Damaging | 3.93 | Benign | 0.13 | Tolerated | 0.1830 | 0.6011 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.259T>C | S87P 2D AIThe SynGAP1 missense variant S87P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Based on the available predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.550904 | Binding | 0.302 | 0.878 | 0.500 | -8.566 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.073 | Likely Benign | -2.04 | Neutral | 0.676 | Possibly Damaging | 0.307 | Benign | 3.75 | Benign | 0.00 | Affected | 0.1639 | 0.4260 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2614T>C | S872P 2D AIThe SynGAP1 missense variant S872P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The only tools that predict a pathogenic outcome are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also predicts Likely Benign. Foldetta results are not available for this variant. Overall, the majority of evidence indicates that S872P is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.662664 | Binding | 0.262 | 0.865 | 0.125 | -4.291 | Likely Benign | 0.273 | Likely Benign | Likely Benign | 0.187 | Likely Benign | -1.91 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.59 | Benign | 0.13 | Tolerated | 0.2321 | 0.5581 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2626T>C | S876P 2D AIThe SynGAP1 missense variant S876P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only two tools, polyPhen‑2 HumDiv and HumVar, predict a pathogenic outcome. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the consensus of the available predictions indicates that S876P is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.549308 | Disordered | 0.631130 | Binding | 0.280 | 0.872 | 0.250 | -5.002 | Likely Benign | 0.229 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -2.04 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.56 | Benign | 0.35 | Tolerated | 0.1979 | 0.6928 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2674T>C | S892P 2D AIThe SynGAP1 missense variant S892P is reported in gnomAD (ID 6‑33443226‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign impact. This conclusion is not contradicted by ClinVar, which contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.473390 | Uncertain | 0.319 | 0.926 | 0.875 | 6-33443226-T-C | 1 | 6.20e-7 | -3.247 | Likely Benign | 0.222 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -0.87 | Neutral | 0.057 | Benign | 0.047 | Benign | 2.60 | Benign | 0.01 | Affected | 4.32 | 4 | 0.2230 | 0.5251 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.2689T>C | S897P 2D AIThe SynGAP1 missense variant S897P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.699094 | Disordered | 0.418474 | Uncertain | 0.292 | 0.928 | 0.500 | -4.088 | Likely Benign | 0.216 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -0.21 | Neutral | 0.990 | Probably Damaging | 0.856 | Possibly Damaging | 2.64 | Benign | 0.04 | Affected | 0.2014 | 0.5933 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2692T>C | S898P 2D AIThe SynGAP1 missense variant S898P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are not available. Overall, the majority of predictions (six benign vs. four pathogenic) support a benign classification. There is no ClinVar entry to contradict this assessment, so the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.690604 | Disordered | 0.426070 | Uncertain | 0.305 | 0.922 | 0.500 | -4.192 | Likely Benign | 0.307 | Likely Benign | Likely Benign | 0.149 | Likely Benign | 0.44 | Neutral | 0.998 | Probably Damaging | 0.939 | Probably Damaging | 2.45 | Pathogenic | 0.01 | Affected | 0.2400 | 0.6555 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2776T>C | S926P 2D AIThe SynGAP1 missense variant S926P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default all indicate pathogenicity. The high‑accuracy consensus, SGM‑Consensus, is derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (benign), FATHMM (pathogenic), and PROVEAN (pathogenic), yielding a Likely Pathogenic classification. AlphaMissense‑Optimized is uncertain, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for S926P. This conclusion is not contradicted by ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.981753 | Binding | 0.295 | 0.854 | 0.250 | -3.873 | Likely Benign | 0.916 | Likely Pathogenic | Ambiguous | 0.424 | Likely Benign | -3.79 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.51 | Pathogenic | 0.00 | Affected | 0.1738 | 0.5058 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2863T>C | S955P 2D AIThe SynGAP1 missense variant S955P is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443415‑T‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are SIFT and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984871 | Disordered | 0.945325 | Binding | 0.350 | 0.924 | 0.750 | Uncertain | 1 | 6-33443415-T-C | 3 | 1.86e-6 | -2.584 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.098 | Likely Benign | -0.75 | Neutral | 0.001 | Benign | 0.004 | Benign | 2.33 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2629 | 0.5537 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||
| c.2866T>C | S956P 2D AIThe SynGAP1 missense variant S956P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984871 | Disordered | 0.957345 | Binding | 0.364 | 0.917 | 0.750 | -3.526 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.157 | Likely Benign | -0.55 | Neutral | 0.000 | Benign | 0.001 | Benign | 1.95 | Pathogenic | 0.05 | Affected | 0.2670 | 0.5337 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2965T>C | S989P 2D AIThe SynGAP1 missense variant S989P is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign; Foldetta results are unavailable. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.908835 | Binding | 0.296 | 0.911 | 0.750 | -2.688 | Likely Benign | 0.209 | Likely Benign | Likely Benign | 0.147 | Likely Benign | -2.48 | Neutral | 0.010 | Benign | 0.015 | Benign | 2.67 | Benign | 0.00 | Affected | 0.1690 | 0.4941 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2968T>C | S990P 2D AIThe SynGAP1 missense variant S990P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of any ClinVar pathogenic annotation. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.902387 | Binding | 0.301 | 0.919 | 0.750 | -3.150 | Likely Benign | 0.122 | Likely Benign | Likely Benign | 0.117 | Likely Benign | -1.95 | Neutral | 0.586 | Possibly Damaging | 0.377 | Benign | 2.74 | Benign | 0.01 | Affected | 0.1910 | 0.5345 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2995T>C | S999P 2D AIThe SynGAP1 missense variant S999P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact for S999P, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.950682 | Binding | 0.262 | 0.897 | 0.625 | -2.279 | Likely Benign | 0.095 | Likely Benign | Likely Benign | 0.111 | Likely Benign | -1.05 | Neutral | 0.966 | Probably Damaging | 0.773 | Possibly Damaging | 2.65 | Benign | 0.04 | Affected | 0.1963 | 0.6268 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3037T>C | S1013P 2D AIThe SynGAP1 missense variant S1013P is reported in gnomAD (ID 6‑33443589‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the consensus score from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.899570 | Binding | 0.308 | 0.846 | 0.625 | 6-33443589-T-C | 2 | 1.24e-6 | -2.563 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 0.103 | Likely Benign | -1.27 | Neutral | 0.453 | Possibly Damaging | 0.150 | Benign | 2.66 | Benign | 0.15 | Tolerated | 3.77 | 5 | 0.2338 | 0.5430 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.3067T>C | S1023P 2D AIThe SynGAP1 missense variant S1023P is reported in gnomAD (ID 6‑33443619‑T‑C) but has no ClinVar entry (ClinVar status: not reported). Functional prediction tools are split: benign calls come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic calls come from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, SGM Consensus remains inconclusive, and Foldetta (combining FoldX‑MD and Rosetta) has no available result. Overall, the majority of standard predictors lean toward pathogenicity, whereas the few high‑accuracy tools do not support a pathogenic verdict. Thus, the variant is most likely pathogenic based on the aggregate predictions, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.831250 | Disordered | 0.990262 | Binding | 0.322 | 0.750 | 0.500 | 6-33443619-T-C | 2 | 1.24e-6 | -5.634 | Likely Benign | 0.679 | Likely Pathogenic | Likely Benign | 0.146 | Likely Benign | -2.11 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.43 | Pathogenic | 0.02 | Affected | 3.77 | 5 | 0.1818 | 0.4616 | -1 | 1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||
| c.3097T>C | S1033P 2D AIThe SynGAP1 missense variant S1033P is not reported in ClinVar and is absent from gnomAD, indicating no documented population frequency. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are not available, so they do not influence the overall assessment. Based on the consensus of all available predictions, the variant is most likely benign, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.993473 | Binding | 0.294 | 0.737 | 0.625 | -2.046 | Likely Benign | 0.268 | Likely Benign | Likely Benign | 0.068 | Likely Benign | 0.05 | Neutral | 0.002 | Benign | 0.005 | Benign | 2.68 | Benign | 0.27 | Tolerated | 0.1889 | 0.5486 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3139T>C | S1047P 2D AIThe SynGAP1 missense variant S1047P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only SIFT indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions points to a benign impact, and this is consistent with the lack of any ClinVar pathogenic annotation. Thus, the variant is most likely benign, and this does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.963420 | Disordered | 0.933764 | Binding | 0.409 | 0.909 | 0.750 | -2.415 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.064 | Likely Benign | 0.52 | Neutral | 0.001 | Benign | 0.001 | Benign | 2.62 | Benign | 0.03 | Affected | 0.2373 | 0.5656 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.313T>C | S105P 2D AIThe SynGAP1 missense variant S105P is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only two tools—polyPhen‑2 HumDiv and SIFT—predict a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as likely benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates likely benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign effect, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.669201 | Binding | 0.364 | 0.870 | 0.625 | Uncertain | 1 | -3.631 | Likely Benign | 0.166 | Likely Benign | Likely Benign | 0.204 | Likely Benign | 0.03 | Neutral | 0.808 | Possibly Damaging | 0.212 | Benign | 4.00 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2236 | 0.4584 | -1 | 1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||
| c.3217T>C | S1073P 2D AIThe SynGAP1 missense variant S1073P is reported in gnomAD (variant ID 6‑33443769‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for S1073P, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.916840 | Disordered | 0.985818 | Binding | 0.313 | 0.905 | 0.750 | 6-33443769-T-C | 1 | 6.23e-7 | -4.520 | Likely Benign | 0.338 | Likely Benign | Likely Benign | 0.082 | Likely Benign | -0.76 | Neutral | 0.006 | Benign | 0.008 | Benign | 3.85 | Benign | 0.01 | Affected | 3.77 | 5 | 0.2243 | 0.5900 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.3259T>C | S1087P 2D AIThe SynGAP1 missense variant S1087P is reported in gnomAD (ID 6‑33443811‑T‑C) and has no ClinVar entry. All available in silico predictors classify it as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Based on the unanimous benign predictions and lack of ClinVar pathogenic annotation, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.974805 | Binding | 0.357 | 0.891 | 1.000 | 6-33443811-T-C | 1 | 6.34e-7 | -2.946 | Likely Benign | 0.133 | Likely Benign | Likely Benign | 0.135 | Likely Benign | -1.92 | Neutral | 0.006 | Benign | 0.008 | Benign | 2.56 | Benign | 0.12 | Tolerated | 3.77 | 5 | 0.1803 | 0.5700 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.3280T>C | S1094P 2D AIThe SynGAP1 missense variant S1094P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for S1094P, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.938133 | Disordered | 0.981352 | Binding | 0.358 | 0.877 | 1.000 | -2.430 | Likely Benign | 0.285 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -1.31 | Neutral | 0.990 | Probably Damaging | 0.798 | Possibly Damaging | 2.46 | Pathogenic | 0.13 | Tolerated | 0.2322 | 0.6099 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3394T>C | S1132P 2D AIThe SynGAP1 missense variant S1132P is listed in ClinVar with an uncertain significance (ClinVar ID 1341927.0) and is present in the gnomAD database (gnomAD ID 6‑33443946‑T‑C). All available in‑silico predictors uniformly classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or likely benign outcomes. No tool in the dataset predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status is unavailable. Overall, the computational evidence strongly supports a benign effect, which is consistent with the ClinVar uncertain classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.845506 | Binding | 0.289 | 0.894 | 0.750 | Conflicting | 3 | 6-33443946-T-C | 1 | 6.74e-7 | -1.423 | Likely Benign | 0.144 | Likely Benign | Likely Benign | 0.301 | Likely Benign | 0.38 | Neutral | 0.003 | Benign | 0.006 | Benign | 5.40 | Benign | 0.28 | Tolerated | 4.32 | 4 | 0.2081 | 0.5700 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||
| c.3412T>C | S1138P 2D AIThe SynGAP1 missense variant S1138P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (both HumDiv and HumVar models) predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.738250 | Binding | 0.346 | 0.869 | 1.000 | -5.023 | Likely Benign | 0.187 | Likely Benign | Likely Benign | 0.456 | Likely Benign | -1.21 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 5.41 | Benign | 0.10 | Tolerated | 0.2297 | 0.5856 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3424T>C | S1142P 2D AIThe SynGAP1 missense variant S1142P is listed in ClinVar (ID 2747352.0) as Benign and is present in gnomAD (variant ID 6‑33444459‑T‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, aligning with the ClinVar classification and not contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | Likely Benign | 1 | 6-33444459-T-C | 1 | 6.20e-7 | -2.713 | Likely Benign | 0.222 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -2.19 | Neutral | 0.918 | Possibly Damaging | 0.761 | Possibly Damaging | 2.64 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2080 | 0.5618 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||
| c.3451T>C | S1151P 2D AIThe SynGAP1 missense variant S1151P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only two tools, polyPhen‑2 HumDiv and HumVar, predict a pathogenic outcome. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy methods further support this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign; Foldetta results are not available. Overall, the consensus of the available predictions indicates that the variant is most likely benign, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.741537 | Disordered | 0.805072 | Binding | 0.394 | 0.839 | 0.625 | -3.031 | Likely Benign | 0.298 | Likely Benign | Likely Benign | 0.123 | Likely Benign | -1.47 | Neutral | 0.995 | Probably Damaging | 0.892 | Possibly Damaging | 2.67 | Benign | 0.11 | Tolerated | 0.1953 | 0.5320 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3475T>C | S1159P 2D AIThe SynGAP1 missense variant S1159P is not reported in ClinVar and has no gnomAD entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and the SGM‑Consensus score, which is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus (majority vote) is benign. No Foldetta stability prediction is available for this residue. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.626927 | Disordered | 0.867068 | Binding | 0.343 | 0.846 | 0.375 | -3.656 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.188 | Likely Benign | -2.06 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.64 | Benign | 0.14 | Tolerated | 0.1807 | 0.5111 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3493T>C | S1165P 2D AIThe SynGAP1 missense variant S1165P is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic, whereas the SGM‑Consensus remains Likely Benign; Foldetta results are unavailable. Overall, the majority of tools (six benign vs. four pathogenic) indicate that the variant is most likely benign. This conclusion is not contradicted by ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.509769 | Disordered | 0.835017 | Binding | 0.308 | 0.807 | 0.375 | -3.476 | Likely Benign | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.147 | Likely Benign | -1.87 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.55 | Benign | 0.23 | Tolerated | 0.2295 | 0.4930 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3550T>C | S1184P 2D AIThe SynGAP1 missense variant S1184P is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains likely benign; Foldetta results are unavailable. Overall, the majority of tools and the consensus prediction lean toward a benign interpretation, and this assessment does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.648219 | Disordered | 0.514669 | Binding | 0.624 | 0.642 | 0.500 | -4.829 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.110 | Likely Benign | -1.38 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.66 | Benign | 0.16 | Tolerated | 0.1772 | 0.4569 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||
| c.3556T>C | S1186P 2D AIThe SynGAP1 missense variant S1186P lies in a coiled‑coil domain. ClinVar has no entry for this variant, and it is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM, while those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the balance of evidence leans toward pathogenicity, with no ClinVar status to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.562014 | Disordered | 0.506433 | Binding | 0.634 | 0.636 | 0.625 | -6.365 | Likely Benign | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.198 | Likely Benign | -2.51 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.64 | Benign | 0.09 | Tolerated | 0.1784 | 0.4006 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3610T>C | S1204P 2D AIThe SynGAP1 missense variant S1204P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include PROVEAN, SIFT, and FATHMM, while those that agree on a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive, and Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact for S1204P. This prediction does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.595080 | Disordered | 0.541098 | Binding | 0.887 | 0.587 | 0.375 | -14.031 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.502 | Likely Pathogenic | -1.80 | Neutral | 0.988 | Probably Damaging | 0.856 | Possibly Damaging | 5.39 | Benign | 0.31 | Tolerated | 0.2139 | 0.3886 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3634T>C | S1212P 2D AIThe SynGAP1 missense variant S1212P is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Prediction tools that classify the variant as benign include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic effect. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus likewise reports likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of computational evidence points to the variant being most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | -13.336 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.239 | Likely Benign | -3.79 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.05 | Pathogenic | 0.00 | Affected | 0.1585 | 0.4370 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||
| c.3673T>C | S1225P 2D AIThe SynGAP1 missense variant S1225P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy predictors (five pathogenic vs. four benign) indicate a pathogenic impact. This conclusion is not contradicted by ClinVar status, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.513880 | Disordered | 0.441915 | Uncertain | 0.891 | 0.544 | 0.500 | -12.278 | Likely Pathogenic | 0.973 | Likely Pathogenic | Likely Pathogenic | 0.489 | Likely Benign | -2.07 | Neutral | 0.784 | Possibly Damaging | 0.575 | Possibly Damaging | 5.36 | Benign | 0.25 | Tolerated | 0.1499 | 0.4570 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3769T>C | S1257P 2D AIThe SynGAP1 missense variant S1257P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, and FATHMM, whereas those that predict a pathogenic impact are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as likely pathogenic; Foldetta results are unavailable. Overall, the majority of computational evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.541878 | Disordered | 0.482380 | Uncertain | 0.889 | 0.572 | 0.375 | -11.985 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.150 | Likely Benign | -2.54 | Deleterious | 0.966 | Probably Damaging | 0.773 | Possibly Damaging | 2.55 | Benign | 0.06 | Tolerated | 0.1656 | 0.4519 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||
| c.385T>C | S129P 2D AIThe SynGAP1 missense variant S129P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of tools, including the high‑accuracy predictors, indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.517562 | Disordered | 0.713635 | Binding | 0.311 | 0.880 | 0.625 | -2.469 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.220 | Likely Benign | -0.09 | Neutral | 0.454 | Possibly Damaging | 0.133 | Benign | 4.11 | Benign | 0.32 | Tolerated | 0.1998 | 0.4989 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.418T>C | S140P 2D AIThe SynGAP1 missense variant S140P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools specifically show AlphaMissense‑Optimized as pathogenic, while SGM Consensus and Foldetta are unavailable. Overall, the majority of evidence points to a pathogenic impact for S140P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.476583 | Structured | 0.587898 | Binding | 0.321 | 0.896 | 0.625 | -4.089 | Likely Benign | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.287 | Likely Benign | -2.59 | Deleterious | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.52 | Benign | 0.08 | Tolerated | 0.1726 | 0.5117 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||||
| c.436T>C | S146P 2D AIThe SynGAP1 missense variant S146P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions arise from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates likely pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for S146P, and this conclusion is not contradicted by ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.541878 | Disordered | 0.508612 | Binding | 0.349 | 0.837 | 0.625 | -13.292 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.215 | Likely Benign | -3.11 | Deleterious | 0.392 | Benign | 0.230 | Benign | 3.60 | Benign | 0.00 | Affected | 0.1934 | 0.4627 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.49T>C | S17P 2D AIThe SynGAP1 missense variant S17P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a benign effect. AlphaMissense‑Optimized independently scores the variant as benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | -3.251 | Likely Benign | 0.256 | Likely Benign | Likely Benign | 0.210 | Likely Benign | -0.65 | Neutral | 0.212 | Benign | 0.010 | Benign | 4.01 | Benign | 0.00 | Affected | 0.2406 | 0.6473 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.538T>C | S180P 2D AIThe SynGAP1 missense variant S180P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; the Foldetta protein‑folding stability analysis is unavailable. Based on the preponderance of pathogenic predictions and the high‑accuracy consensus, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.442877 | Uncertain | 0.320 | 0.616 | 0.500 | -14.574 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.225 | Likely Benign | -3.19 | Deleterious | 0.917 | Possibly Damaging | 0.446 | Benign | 3.81 | Benign | 0.01 | Affected | 0.1916 | 0.4735 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.544T>C | S182P 2D AIThe SynGAP1 missense variant S182P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority of the four high‑accuracy tools) also indicates pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple independent predictors points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.501700 | Disordered | 0.436016 | Uncertain | 0.368 | 0.619 | 0.625 | -13.362 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.279 | Likely Benign | -3.71 | Deleterious | 0.838 | Possibly Damaging | 0.367 | Benign | 3.67 | Benign | 0.00 | Affected | 0.2438 | 0.5030 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.553T>C | S185P 2D AIThe SynGAP1 missense variant S185P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default—predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the S185P variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.545602 | Disordered | 0.430485 | Uncertain | 0.365 | 0.623 | 0.500 | -15.129 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.285 | Likely Benign | -4.02 | Deleterious | 0.940 | Possibly Damaging | 0.462 | Possibly Damaging | 3.56 | Benign | 0.00 | Affected | 0.2156 | 0.5242 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.610T>C | S204P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 S204P variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and polyPhen‑2 HumVar, whereas a separate group predicts a pathogenic effect: FoldX, Rosetta, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The remaining tools (premPS) are inconclusive and are not counted in either group. High‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain (treated as unavailable), the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and therefore unavailable, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a pathogenic effect. Overall, the majority of consensus predictions lean toward pathogenicity, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.268042 | Structured | 0.420667 | Uncertain | 0.816 | 0.405 | 0.125 | -8.855 | Likely Pathogenic | 0.820 | Likely Pathogenic | Ambiguous | 3.71 | Destabilizing | 0.4 | 4.61 | Destabilizing | 4.16 | Destabilizing | 0.61 | Ambiguous | 0.137 | Likely Benign | -1.28 | Neutral | 0.808 | Possibly Damaging | 0.382 | Benign | 4.13 | Benign | 0.12 | Tolerated | 0.1430 | 0.4815 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||
| c.634T>C | S212P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S212P is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include FoldX and FATHMM, whereas the remaining tools—REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict it to be pathogenic. High‑accuracy methods give consistent pathogenic results: AlphaMissense‑Optimized scores it as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a destabilizing, pathogenic effect. With the overwhelming majority of tools supporting pathogenicity and no ClinVar entry to contradict, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.158265 | Structured | 0.381517 | Uncertain | 0.846 | 0.278 | 0.125 | -13.051 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.02 | Likely Benign | 0.9 | 6.30 | Destabilizing | 3.16 | Destabilizing | 0.90 | Ambiguous | 0.809 | Likely Pathogenic | -4.13 | Deleterious | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 5.75 | Benign | 0.01 | Affected | 0.1859 | 0.5679 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.673T>C | S225P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S225P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign calls come from premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic calls arise from REVEL, FoldX, PROVEAN, and ESM1b. Two tools—Rosetta and Foldetta—return uncertain results and are treated as unavailable. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts benign, whereas the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta remains unavailable. Overall, the majority of evidence leans toward a benign effect, and this conclusion does not conflict with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.150080 | Structured | 0.344191 | Uncertain | 0.817 | 0.319 | 0.250 | -8.084 | Likely Pathogenic | 0.156 | Likely Benign | Likely Benign | 2.22 | Destabilizing | 1.8 | 1.14 | Ambiguous | 1.68 | Ambiguous | 0.15 | Likely Benign | 0.503 | Likely Pathogenic | -3.31 | Deleterious | 0.396 | Benign | 0.133 | Benign | 5.91 | Benign | 0.17 | Tolerated | 0.2128 | 0.6946 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||
| c.676T>C | S226P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S226P is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining tools (REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus) all predict a pathogenic or likely pathogenic outcome. Uncertain results are reported by FoldX, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is inconclusive, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a pathogenic effect. Overall, the preponderance of evidence indicates that S226P is most likely pathogenic, and this conclusion is not contradicted by ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.129801 | Structured | 0.334959 | Uncertain | 0.800 | 0.324 | 0.250 | -11.030 | Likely Pathogenic | 0.933 | Likely Pathogenic | Ambiguous | 1.51 | Ambiguous | 1.1 | 5.29 | Destabilizing | 3.40 | Destabilizing | 0.57 | Ambiguous | 0.776 | Likely Pathogenic | -3.22 | Deleterious | 0.971 | Probably Damaging | 0.543 | Possibly Damaging | 5.74 | Benign | 0.04 | Affected | 0.2837 | 0.7038 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.703T>C | S235P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S235P is listed in ClinVar as Pathogenic (ClinVar ID 1067856.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect are polyPhen‑2 HumVar and FATHMM; all other evaluated algorithms—including REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic verdict; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports pathogenic. No predictions or stability results are missing or inconclusive. **Based on the collective predictions, the variant is most likely pathogenic, and this conclusion aligns with its ClinVar status.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.250310 | Structured | 0.319150 | Uncertain | 0.743 | 0.331 | 0.000 | Likely Pathogenic | 1 | -14.857 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 4.02 | Destabilizing | 0.1 | 6.91 | Destabilizing | 5.47 | Destabilizing | 1.23 | Destabilizing | 0.870 | Likely Pathogenic | -4.24 | Deleterious | 0.917 | Possibly Damaging | 0.446 | Benign | 5.47 | Benign | 0.01 | Affected | 3.40 | 14 | 0.2432 | 0.5307 | 1 | -1 | -0.8 | 10.04 | 201.5 | 17.0 | 0.1 | 0.0 | -0.6 | 0.0 | X | Potentially Pathogenic | In the WT, the hydroxyl group of Ser235, located in a β-α loop between an anti-parallel β sheet strand (res. Gly227-Phe231) and an α helix (residues Ala236-Val250), forms hydrogen bonds with the GAP domain loop residue Glu680 and with the backbone amide groups of Ala237 and Glu238 from the α helix. In the variant simulations, the pyrrolidine ring of Pro235 cannot stabilize the α helix end or maintain tertiary bonding interactions between the PH and GAP domains via hydrogen bonding as effectively as serine. | ||||||||||||||||
| c.82T>C | S28P 2D AIThe SynGAP1 missense variant S28P is listed in ClinVar (ID 1500161.0) with an uncertain significance designation and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while only SIFT indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta stability analysis is unavailable. Overall, the collective evidence points to a benign classification for S28P, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.545602 | Disordered | 0.438157 | Uncertain | 0.354 | 0.884 | 0.125 | Uncertain | 1 | -3.309 | Likely Benign | 0.051 | Likely Benign | Likely Benign | 0.047 | Likely Benign | 1.37 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.53 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2432 | 0.5499 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||
| c.886T>C | S296P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S296P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL, FoldX, Rosetta, and SIFT, whereas pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. When predictions are grouped by agreement, four tools predict benign and seven predict pathogenic, with premPS remaining uncertain. High‑accuracy methods give a mixed signal: AlphaMissense‑Optimized predicts pathogenic; the SGM‑Consensus, derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports a benign effect. Thus, the majority of evidence leans toward pathogenicity, and this conclusion is consistent with the lack of ClinVar annotation and gnomAD absence, which do not contradict the prediction. Overall, the variant is most likely pathogenic based on the current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.444081 | Structured | 0.282669 | Uncertain | 0.887 | 0.284 | 0.250 | -8.302 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.04 | Likely Benign | 0.4 | 0.18 | Likely Benign | 0.11 | Likely Benign | 0.72 | Ambiguous | 0.439 | Likely Benign | -3.74 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.92 | Pathogenic | 0.12 | Tolerated | 0.2273 | 0.6320 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.898T>C | S300P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S300P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. The remaining tools—FoldX, Foldetta, and ESM1b—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.356642 | Structured | 0.256848 | Uncertain | 0.742 | 0.280 | 0.375 | -7.379 | In-Between | 0.110 | Likely Benign | Likely Benign | 1.24 | Ambiguous | 1.0 | 0.06 | Likely Benign | 0.65 | Ambiguous | 0.50 | Likely Benign | 0.086 | Likely Benign | -2.46 | Neutral | 0.784 | Possibly Damaging | 0.390 | Benign | 1.54 | Pathogenic | 0.02 | Affected | 0.2192 | 0.5585 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||
| c.904T>C | S302P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S302P is not reported in ClinVar and is absent from gnomAD. Prediction tools that uniformly indicate a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only Rosetta predicts a pathogenic outcome, while FoldX and Foldetta are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM‑Consensus, derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also supports a benign classification. Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains inconclusive. Overall, the preponderance of evidence points to a benign impact for S302P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.263489 | Uncertain | 0.616 | 0.258 | 0.375 | -2.485 | Likely Benign | 0.121 | Likely Benign | Likely Benign | 1.19 | Ambiguous | 0.4 | 2.74 | Destabilizing | 1.97 | Ambiguous | 0.14 | Likely Benign | 0.101 | Likely Benign | -0.89 | Neutral | 0.157 | Benign | 0.153 | Benign | 4.11 | Benign | 0.20 | Tolerated | 0.2522 | 0.6243 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||
| c.991T>C | S331P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S331P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, ESM1b, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are Rosetta, polyPhen‑2 HumDiv, and FATHMM. Two tools give inconclusive results: AlphaMissense‑Default and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.433034 | Structured | 0.346458 | Uncertain | 0.658 | 0.475 | 0.250 | -5.104 | Likely Benign | 0.409 | Ambiguous | Likely Benign | 0.33 | Likely Benign | 0.2 | 2.35 | Destabilizing | 1.34 | Ambiguous | 0.44 | Likely Benign | 0.186 | Likely Benign | -1.81 | Neutral | 0.784 | Possibly Damaging | 0.390 | Benign | 1.83 | Pathogenic | 0.26 | Tolerated | 0.2306 | 0.4086 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||
| c.1231A>C | I411L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I411L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas a majority (premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. Tools with uncertain or mixed outputs are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, yielding no definitive call; and Foldetta also reports an uncertain stability change. Overall, the majority of standard predictors lean toward pathogenicity, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current computational evidence, though high‑accuracy tools remain inconclusive. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.116183 | Structured | 0.339366 | Uncertain | 0.927 | 0.198 | 0.000 | -10.723 | Likely Pathogenic | 0.819 | Likely Pathogenic | Ambiguous | 1.02 | Ambiguous | 0.3 | 1.89 | Ambiguous | 1.46 | Ambiguous | 1.17 | Destabilizing | 0.285 | Likely Benign | -1.84 | Neutral | 0.908 | Possibly Damaging | 0.943 | Probably Damaging | 3.36 | Benign | 0.01 | Affected | 0.0876 | 0.3539 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1360A>C | I454L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I454L is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, while polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default predict a pathogenic impact. The remaining tools—FoldX, Rosetta, Foldetta, premPS, ESM1b, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward benign, and Foldetta also uncertain. Overall, the majority of reliable predictors and the SGM Consensus favor a benign classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.254060 | Structured | 0.312811 | Uncertain | 0.965 | 0.182 | 0.000 | -7.852 | In-Between | 0.786 | Likely Pathogenic | Ambiguous | 0.57 | Ambiguous | 0.1 | 1.36 | Ambiguous | 0.97 | Ambiguous | 0.80 | Ambiguous | 0.246 | Likely Benign | -1.98 | Neutral | 0.908 | Possibly Damaging | 0.943 | Probably Damaging | 3.51 | Benign | 0.26 | Tolerated | 0.0653 | 0.2857 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1447A>C | I483L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I483L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree that the substitution is benign: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a neutral effect, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Benign.” In contrast, polyPhen‑2 (HumDiv and HumVar) and ESM1b predict a pathogenic impact, and premPS remains uncertain. High‑accuracy assessments are uniformly benign: AlphaMissense‑Optimized is benign, the SGM‑Consensus is “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign effect. Taken together, the majority of evidence, including the high‑accuracy tools, supports a benign classification for I483L. This conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.206376 | Structured | 0.415850 | Uncertain | 0.798 | 0.254 | 0.000 | -10.258 | Likely Pathogenic | 0.332 | Likely Benign | Likely Benign | 0.31 | Likely Benign | 0.1 | 0.38 | Likely Benign | 0.35 | Likely Benign | 0.67 | Ambiguous | 0.341 | Likely Benign | -1.86 | Neutral | 0.879 | Possibly Damaging | 0.970 | Probably Damaging | 4.09 | Benign | 0.31 | Tolerated | 0.0664 | 0.2461 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||
| c.1447A>T | I483L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I483L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree that the substitution is benign: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a neutral effect, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Benign.” In contrast, polyPhen‑2 (HumDiv and HumVar) and ESM1b predict a pathogenic impact, and premPS remains uncertain. High‑accuracy assessments are uniformly benign: AlphaMissense‑Optimized is benign, the SGM‑Consensus is “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign effect. Taken together, the majority of evidence, including the high‑accuracy tools, supports a benign classification for I483L. This conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.206376 | Structured | 0.415850 | Uncertain | 0.798 | 0.254 | 0.000 | -10.258 | Likely Pathogenic | 0.332 | Likely Benign | Likely Benign | 0.31 | Likely Benign | 0.1 | 0.38 | Likely Benign | 0.35 | Likely Benign | 0.67 | Ambiguous | 0.342 | Likely Benign | -1.86 | Neutral | 0.879 | Possibly Damaging | 0.970 | Probably Damaging | 4.09 | Benign | 0.31 | Tolerated | 0.0664 | 0.2461 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||
| c.1480A>C | I494L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I494L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the balance of evidence (five pathogenic versus four benign predictions, with several uncertain calls) leans toward a pathogenic interpretation. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | -10.175 | Likely Pathogenic | 0.478 | Ambiguous | Likely Benign | 0.28 | Likely Benign | 0.1 | 1.11 | Ambiguous | 0.70 | Ambiguous | 0.91 | Ambiguous | 0.513 | Likely Pathogenic | -1.69 | Neutral | 0.645 | Possibly Damaging | 0.718 | Possibly Damaging | -0.88 | Pathogenic | 0.11 | Tolerated | 0.0816 | 0.2851 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1480A>T | I494L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I494L variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta indicating uncertain stability change. Overall, the balance of evidence leans toward pathogenicity, with a majority of tools and the SGM Consensus supporting a deleterious effect. This conclusion does not contradict ClinVar status, as the variant is currently unreported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | -10.175 | Likely Pathogenic | 0.478 | Ambiguous | Likely Benign | 0.28 | Likely Benign | 0.1 | 1.11 | Ambiguous | 0.70 | Ambiguous | 0.91 | Ambiguous | 0.513 | Likely Pathogenic | -1.69 | Neutral | 0.645 | Possibly Damaging | 0.718 | Possibly Damaging | -0.88 | Pathogenic | 0.11 | Tolerated | 0.0816 | 0.2851 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.148A>C | I50L 2D AIThe SynGAP1 missense variant I50L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is benign; Foldetta results are not available. Overall, the consensus of the available predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.295083 | Structured | 0.449965 | Uncertain | 0.545 | 0.708 | 0.000 | -3.509 | Likely Benign | 0.300 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -0.73 | Neutral | 0.010 | Benign | 0.004 | Benign | 3.91 | Benign | 0.00 | Affected | 0.0749 | 0.3247 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.1501A>C | I501L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I501L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, Rosetta, Foldetta, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar) and SIFT. Uncertain or inconclusive results are reported for FoldX, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM‑Consensus as Likely Benign, and Foldetta as Benign, all supporting a non‑deleterious effect. Based on the preponderance of benign predictions and the concordant high‑accuracy tools, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.079919 | Structured | 0.366596 | Uncertain | 0.886 | 0.153 | 0.000 | -7.348 | In-Between | 0.282 | Likely Benign | Likely Benign | 0.78 | Ambiguous | 0.2 | -0.34 | Likely Benign | 0.22 | Likely Benign | 0.86 | Ambiguous | 0.219 | Likely Benign | -1.92 | Neutral | 0.930 | Possibly Damaging | 0.985 | Probably Damaging | 3.51 | Benign | 0.05 | Affected | 0.0817 | 0.2261 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||
| c.151A>C | I51L 2D AIThe SynGAP1 missense variant I51L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the consensus of the majority of tools and the high‑accuracy predictions point to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.291804 | Structured | 0.454181 | Uncertain | 0.606 | 0.710 | 0.000 | 0.408 | Likely Benign | 0.147 | Likely Benign | Likely Benign | 0.080 | Likely Benign | 0.14 | Neutral | 0.000 | Benign | 0.001 | Benign | 4.35 | Benign | 0.00 | Affected | 0.0924 | 0.3993 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.1528A>C | I510L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I510L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments—AlphaMissense‑Optimized, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta (combining FoldX‑MD and Rosetta)—all indicate a benign effect. No prediction or folding‑stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.025762 | Structured | 0.250630 | Uncertain | 0.945 | 0.273 | 0.000 | -2.362 | Likely Benign | 0.116 | Likely Benign | Likely Benign | 0.31 | Likely Benign | 0.2 | -0.05 | Likely Benign | 0.13 | Likely Benign | -1.01 | Stabilizing | 0.338 | Likely Benign | 0.69 | Neutral | 0.016 | Benign | 0.130 | Benign | -0.74 | Pathogenic | 1.00 | Tolerated | 0.0819 | 0.2461 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||
| c.1540A>C | I514L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I514L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools (premPS and AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta also predicts a benign outcome. No prediction or folding stability result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation—there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.049374 | Structured | 0.221408 | Uncertain | 0.948 | 0.266 | 0.000 | -10.239 | Likely Pathogenic | 0.490 | Ambiguous | Likely Benign | -0.05 | Likely Benign | 0.2 | 0.34 | Likely Benign | 0.15 | Likely Benign | 0.81 | Ambiguous | 0.310 | Likely Benign | -1.99 | Neutral | 0.879 | Possibly Damaging | 0.985 | Probably Damaging | 3.31 | Benign | 0.12 | Tolerated | 0.0767 | 0.2678 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1585A>C | I529L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I529L is listed in gnomAD (variant ID 6‑33438828‑A‑C) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support benignity: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. No prediction or folding result is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.318242 | Structured | 0.019545 | Uncertain | 0.901 | 0.403 | 0.000 | 6-33438828-A-C | 1 | 6.20e-7 | 0.920 | Likely Benign | 0.066 | Likely Benign | Likely Benign | -0.13 | Likely Benign | 0.0 | -0.11 | Likely Benign | -0.12 | Likely Benign | -0.21 | Likely Benign | 0.309 | Likely Benign | 0.00 | Neutral | 0.001 | Benign | 0.022 | Benign | -1.24 | Pathogenic | 0.48 | Tolerated | 3.37 | 35 | 0.0819 | 0.3789 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||
| c.1750A>C | I584L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I584L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM, while premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the preponderance of evidence points to a benign impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.046673 | Uncertain | 0.846 | 0.244 | 0.000 | -8.266 | Likely Pathogenic | 0.285 | Likely Benign | Likely Benign | -0.18 | Likely Benign | 0.1 | -0.30 | Likely Benign | -0.24 | Likely Benign | 0.84 | Ambiguous | 0.420 | Likely Benign | -1.74 | Neutral | 0.008 | Benign | 0.046 | Benign | -1.23 | Pathogenic | 0.18 | Tolerated | 0.0927 | 0.2817 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1765A>C | I589L 2D AIThe SynGAP1 missense variant I589L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only PROVEAN, whereas the remaining tools (REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus) all predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Pathogenic”; AlphaMissense‑Optimized and Foldetta are inconclusive and therefore not considered evidence. Taken together, the preponderance of evidence points to a pathogenic effect for I589L. This conclusion is not contradicted by ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.018415 | Structured | 0.084536 | Uncertain | 0.927 | 0.214 | 0.000 | -11.337 | Likely Pathogenic | 0.850 | Likely Pathogenic | Ambiguous | 0.95 | Ambiguous | 1.1 | 1.44 | Ambiguous | 1.20 | Ambiguous | 0.95 | Ambiguous | 0.728 | Likely Pathogenic | -1.99 | Neutral | 0.955 | Possibly Damaging | 0.985 | Probably Damaging | -1.76 | Pathogenic | 0.02 | Affected | 0.1243 | 0.3430 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||
| c.1804A>C | I602L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I602L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, PROVEAN, and AlphaMissense‑Optimized, whereas a majority of tools predict pathogenicity: REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Three tools (Rosetta, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the balance of evidence leans toward pathogenicity, with no conflict with the ClinVar status because the variant is not yet classified in that database. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.010221 | Structured | 0.186541 | Uncertain | 0.963 | 0.171 | 0.000 | -9.660 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | -0.15 | Likely Benign | 0.1 | 1.12 | Ambiguous | 0.49 | Likely Benign | 0.91 | Ambiguous | 0.631 | Likely Pathogenic | -1.99 | Neutral | 0.645 | Possibly Damaging | 0.718 | Possibly Damaging | -1.54 | Pathogenic | 0.04 | Affected | 0.1044 | 0.3199 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1876A>C | I626L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I626L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default predict a pathogenic outcome. Three tools—Foldetta, premPS, and Rosetta—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta also yields an uncertain stability prediction. Overall, the balance of evidence leans toward a benign interpretation, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.109221 | Structured | 0.040732 | Uncertain | 0.970 | 0.223 | 0.000 | -10.696 | Likely Pathogenic | 0.622 | Likely Pathogenic | Likely Benign | 0.32 | Likely Benign | 0.1 | 1.00 | Ambiguous | 0.66 | Ambiguous | 0.83 | Ambiguous | 0.311 | Likely Benign | -1.86 | Neutral | 0.955 | Possibly Damaging | 0.985 | Probably Damaging | 3.52 | Benign | 0.12 | Tolerated | 0.0741 | 0.2461 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1888A>C | I630L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I630L is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33440940‑A‑C). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign; the SGM Consensus remains unavailable. Based on the overall predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.036106 | Uncertain | 0.966 | 0.236 | 0.000 | 6-33440940-A-C | -8.949 | Likely Pathogenic | 0.277 | Likely Benign | Likely Benign | -0.39 | Likely Benign | 0.0 | 0.23 | Likely Benign | -0.08 | Likely Benign | 0.33 | Likely Benign | 0.165 | Likely Benign | -1.30 | Neutral | 0.102 | Benign | 0.108 | Benign | -0.81 | Pathogenic | 0.27 | Tolerated | 3.37 | 34 | 0.0688 | 0.2461 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||
| c.190A>C | I64L 2D AIThe SynGAP1 missense variant I64L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus indicates Likely Benign, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact for I64L, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.422041 | Structured | 0.475481 | Uncertain | 0.478 | 0.747 | 0.125 | -2.498 | Likely Benign | 0.437 | Ambiguous | Likely Benign | 0.087 | Likely Benign | -0.27 | Neutral | 0.010 | Benign | 0.001 | Benign | 4.13 | Benign | 0.00 | Affected | 0.0628 | 0.3030 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.190A>T | I64L 2D AIThe SynGAP1 missense variant I64L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus indicates Likely Benign, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.422041 | Structured | 0.475481 | Uncertain | 0.478 | 0.747 | 0.125 | -2.498 | Likely Benign | 0.437 | Ambiguous | Likely Benign | 0.087 | Likely Benign | -0.27 | Neutral | 0.010 | Benign | 0.001 | Benign | 4.13 | Benign | 0.00 | Affected | 0.0628 | 0.3030 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.1999A>C | I667L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I667L is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, indicating no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.142424 | Structured | 0.083597 | Uncertain | 0.927 | 0.379 | 0.000 | -10.452 | Likely Pathogenic | 0.459 | Ambiguous | Likely Benign | 0.93 | Ambiguous | 0.2 | 0.72 | Ambiguous | 0.83 | Ambiguous | 0.75 | Ambiguous | 0.300 | Likely Benign | -1.97 | Neutral | 0.457 | Possibly Damaging | 0.392 | Benign | 3.36 | Benign | 0.13 | Tolerated | 0.0980 | 0.3145 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2047A>C | I683L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I683L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b. Remaining methods (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, while Foldetta remains uncertain. Overall, the balance of evidence favors a benign impact for I683L, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.143268 | Uncertain | 0.848 | 0.314 | 0.000 | -9.988 | Likely Pathogenic | 0.488 | Ambiguous | Likely Benign | 0.63 | Ambiguous | 0.1 | 0.76 | Ambiguous | 0.70 | Ambiguous | 0.69 | Ambiguous | 0.286 | Likely Benign | -2.00 | Neutral | 0.011 | Benign | 0.056 | Benign | 3.43 | Benign | 0.04 | Affected | 0.0957 | 0.3185 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.205A>C | I69L 2D AIThe SynGAP1 missense variant I69L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic call comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign; Foldetta results are not available. Overall, the majority of evidence supports a benign impact for I69L, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.575842 | Disordered | 0.466129 | Uncertain | 0.437 | 0.786 | 0.375 | -2.065 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.067 | Likely Benign | -0.32 | Neutral | 0.267 | Benign | 0.141 | Benign | 4.27 | Benign | 0.00 | Affected | 0.0636 | 0.3447 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.2161A>C | I721L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I721L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. Two tools (premPS and ESM1b) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the predictions are split, but the majority of high‑confidence tools lean toward a benign interpretation. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.394753 | Structured | 0.454550 | Uncertain | 0.957 | 0.437 | 0.125 | -7.843 | In-Between | 0.757 | Likely Pathogenic | Likely Benign | 0.35 | Likely Benign | 0.0 | 0.34 | Likely Benign | 0.35 | Likely Benign | 0.65 | Ambiguous | 0.188 | Likely Benign | -1.83 | Neutral | 0.955 | Possibly Damaging | 0.985 | Probably Damaging | 2.37 | Pathogenic | 0.02 | Affected | 0.0627 | 0.3446 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2188A>C | I730L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I730L is reported as “Likely Benign” in ClinVar and is not present in gnomAD. All available in‑silico predictors classify the change as benign: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments concur: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also reports a benign effect. Consequently, the variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.420109 | Uncertain | 0.591 | 0.619 | 0.750 | -1.681 | Likely Benign | 0.069 | Likely Benign | Likely Benign | -0.05 | Likely Benign | 0.0 | -0.31 | Likely Benign | -0.18 | Likely Benign | -0.17 | Likely Benign | 0.028 | Likely Benign | -0.63 | Neutral | 0.000 | Benign | 0.005 | Benign | 3.53 | Benign | 0.47 | Tolerated | 0.0739 | 0.2869 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2299A>C | I767L 2D AIThe SynGAP1 missense variant I767L is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Based on the collective predictions, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.321458 | Structured | 0.927771 | Binding | 0.369 | 0.872 | 0.125 | -1.881 | Likely Benign | 0.112 | Likely Benign | Likely Benign | 0.159 | Likely Benign | -0.73 | Neutral | 0.001 | Benign | 0.002 | Benign | 4.13 | Benign | 0.34 | Tolerated | 0.1020 | 0.4317 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.22A>C | I8L 2D AIThe SynGAP1 missense variant I8L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote from the four high‑accuracy tools) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the I8L variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.490133 | Structured | 0.543080 | Binding | 0.341 | 0.916 | 0.625 | -1.752 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.124 | Likely Benign | 0.08 | Neutral | 0.002 | Benign | 0.000 | Benign | 4.20 | Benign | 0.00 | Affected | 0.0707 | 0.3776 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.2479A>C | I827L 2D AIThe SynGAP1 missense variant I827L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Benign”). In contrast, the two polyPhen‑2 implementations (HumDiv and HumVar) predict a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also likely benign. No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the majority of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.636272 | Binding | 0.383 | 0.884 | 0.625 | -2.535 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.120 | Likely Benign | -0.64 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 2.71 | Benign | 0.43 | Tolerated | 0.0711 | 0.2901 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.2695A>C | I899L 2D AIThe SynGAP1 missense variant I899L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.443727 | Uncertain | 0.292 | 0.928 | 0.375 | -1.137 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.055 | Likely Benign | -0.36 | Neutral | 0.220 | Benign | 0.078 | Benign | 2.74 | Benign | 0.04 | Affected | 0.0863 | 0.2840 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.2767A>C | I923L 2D AIThe SynGAP1 missense variant I923L is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors—including REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods likewise support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.964857 | Binding | 0.292 | 0.852 | 0.250 | -1.637 | Likely Benign | 0.157 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -0.03 | Neutral | 0.166 | Benign | 0.101 | Benign | 2.82 | Benign | 0.56 | Tolerated | 0.1133 | 0.4528 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.2998A>C | I1000L 2D AIThe SynGAP1 missense variant I1000L is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.957020 | Binding | 0.293 | 0.904 | 0.625 | -2.103 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -0.34 | Neutral | 0.211 | Benign | 0.108 | Benign | 2.76 | Benign | 0.70 | Tolerated | 0.0916 | 0.4034 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3109A>C | I1037L 2D AIThe SynGAP1 missense variant I1037L is not reported in ClinVar and is absent from gnomAD, indicating no known clinical or population evidence. Functional prediction tools uniformly favor a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity, and the only uncertain call comes from AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as “Likely Benign.” High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that I1037L is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.939629 | Disordered | 0.986140 | Binding | 0.309 | 0.774 | 0.625 | -2.563 | Likely Benign | 0.448 | Ambiguous | Likely Benign | 0.074 | Likely Benign | -0.46 | Neutral | 0.421 | Benign | 0.128 | Benign | 2.82 | Benign | 0.81 | Tolerated | 0.0939 | 0.4302 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3115A>C | I1039L 2D AIThe SynGAP1 missense variant I1039L is not reported in ClinVar and is absent from gnomAD, indicating no documented clinical or population evidence. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the consensus of all available predictions strongly supports a benign classification, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.979204 | Binding | 0.292 | 0.806 | 0.625 | -2.035 | Likely Benign | 0.269 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -0.14 | Neutral | 0.264 | Benign | 0.048 | Benign | 2.82 | Benign | 0.98 | Tolerated | 0.1057 | 0.4605 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3343A>C | I1115L 2D AIThe SynGAP1 missense variant I1115L is listed in ClinVar (ID 4178654) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443895‑A‑C). All evaluated in‑silico predictors classify the substitution as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool reports a pathogenic effect, so the pathogenic‑prediction group is empty. High‑accuracy assessments reinforce this benign consensus: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status is unavailable. Overall, the computational evidence strongly supports a benign impact, and this does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.889439 | Disordered | 0.892339 | Binding | 0.308 | 0.912 | 0.750 | Uncertain | 1 | 6-33443895-A-C | 1 | 7.56e-7 | -2.308 | Likely Benign | 0.067 | Likely Benign | Likely Benign | 0.120 | Likely Benign | -0.46 | Neutral | 0.004 | Benign | 0.007 | Benign | 2.82 | Benign | 1.00 | Tolerated | 4.32 | 2 | 0.1154 | 0.4575 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||||
| c.3397A>C | I1133L 2D AIThe SynGAP1 missense variant I1133L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of all available predictions is that I1133L is most likely benign, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.832785 | Binding | 0.316 | 0.892 | 0.750 | -1.147 | Likely Benign | 0.107 | Likely Benign | Likely Benign | 0.202 | Likely Benign | -0.26 | Neutral | 0.000 | Benign | 0.001 | Benign | 5.48 | Benign | 0.23 | Tolerated | 0.0911 | 0.4302 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3502A>C | I1168L 2D AIThe SynGAP1 missense variant I1168L is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors indicates a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the substitution as tolerated, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also classifies it as likely benign. Only AlphaMissense‑Default predicts a pathogenic outcome, but this is an outlier relative to the other tools. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus (majority vote) concurs. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so it does not influence the overall assessment. Overall, the computational evidence overwhelmingly favors a benign classification, and this is consistent with the absence of a ClinVar pathogenic report. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.763262 | Binding | 0.423 | 0.796 | 0.500 | -1.855 | Likely Benign | 0.604 | Likely Pathogenic | Likely Benign | 0.361 | Likely Benign | -0.60 | Neutral | 0.241 | Benign | 0.286 | Benign | 5.48 | Benign | 0.20 | Tolerated | 0.0956 | 0.4465 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3517A>C | I1173L 2D AIThe SynGAP1 missense variant I1173L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly classify the change as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a benign effect. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the evidence strongly supports a benign classification, and there is no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.501700 | Disordered | 0.653145 | Binding | 0.521 | 0.756 | 0.375 | -1.792 | Likely Benign | 0.202 | Likely Benign | Likely Benign | 0.218 | Likely Benign | -0.30 | Neutral | 0.152 | Benign | 0.102 | Benign | 5.41 | Benign | 0.97 | Tolerated | 0.0745 | 0.3004 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||
| c.3604A>C | I1202L 2D AIThe SynGAP1 I1202L missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, and SIFT, whereas those that predict a pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) remains Likely Pathogenic; a Foldetta stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation (none exists). Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.510422 | Binding | 0.874 | 0.593 | 0.250 | -8.026 | Likely Pathogenic | 0.953 | Likely Pathogenic | Ambiguous | 0.119 | Likely Benign | -1.27 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 1.94 | Pathogenic | 0.59 | Tolerated | 0.0788 | 0.2874 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||
| c.3697A>C | I1233L 2D AIThe SynGAP1 missense variant I1233L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the substitution as benign, and AlphaMissense‑Optimized also predicts a benign outcome. No tool predicts pathogenicity; AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also indicates likely benign; Foldetta data are not available. Overall, the consensus of available predictions points to a benign effect, and this is consistent with the lack of ClinVar evidence or gnomAD observation. Thus, the variant is most likely benign, and this conclusion does not contradict ClinVar status, which has no entry for it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -3.031 | Likely Benign | 0.382 | Ambiguous | Likely Benign | 0.113 | Likely Benign | -0.01 | Neutral | 0.211 | Benign | 0.108 | Benign | 2.95 | Benign | 1.00 | Tolerated | 0.0631 | 0.3246 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||
| c.3775A>C | I1259L 2D AIThe SynGAP1 missense variant I1259L is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default predict a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for I1259L, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.494003 | Structured | 0.576405 | Binding | 0.885 | 0.574 | 0.250 | -5.822 | Likely Benign | 0.770 | Likely Pathogenic | Likely Benign | 0.152 | Likely Benign | -0.01 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 2.80 | Benign | 0.18 | Tolerated | 0.0639 | 0.3078 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||
| c.3784A>C | I1262L 2D AIThe SynGAP1 missense variant I1262L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and PROVEAN, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b remains uncertain. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (pathogenic), and PROVEAN (benign)—also indicates pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic impact for I1262L, and this conclusion does not contradict the current ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -7.334 | In-Between | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.269 | Likely Benign | -1.66 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 1.94 | Pathogenic | 0.00 | Affected | 0.0735 | 0.3246 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3787A>C | I1263L 2D AIThe SynGAP1 missense variant I1263L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign, two pathogenic). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -1.210 | Likely Benign | 0.699 | Likely Pathogenic | Likely Benign | 0.194 | Likely Benign | -1.66 | Neutral | 0.011 | Benign | 0.022 | Benign | 1.94 | Pathogenic | 0.00 | Affected | 0.0717 | 0.2890 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.37A>C | I13L 2D AIThe SynGAP1 missense variant I13L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | -2.144 | Likely Benign | 0.100 | Likely Benign | Likely Benign | 0.095 | Likely Benign | 0.07 | Neutral | 0.002 | Benign | 0.001 | Benign | 4.19 | Benign | 0.00 | Affected | 0.1055 | 0.4923 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3991A>C | I1331L 2D AIThe SynGAP1 missense variant I1331L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), which collectively classify the variant as likely benign. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized result is uncertain, and Foldetta stability analysis is unavailable. Overall, the balance of evidence leans toward a benign interpretation, with no conflict with ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -2.450 | Likely Benign | 0.892 | Likely Pathogenic | Ambiguous | 0.180 | Likely Benign | -1.04 | Neutral | 0.762 | Possibly Damaging | 0.785 | Possibly Damaging | 3.45 | Benign | 0.00 | Affected | 0.0752 | 0.3616 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.421A>C | I141L 2D AIThe SynGAP1 missense variant I141L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Only AlphaMissense‑Default predicts a pathogenic outcome, while AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) is not available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.465241 | Structured | 0.577021 | Binding | 0.367 | 0.877 | 0.500 | -6.563 | Likely Benign | 0.787 | Likely Pathogenic | Ambiguous | 0.156 | Likely Benign | -1.00 | Neutral | 0.016 | Benign | 0.013 | Benign | 3.84 | Benign | 0.10 | Tolerated | 0.0742 | 0.3180 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.475A>C | I159L 2D AIThe SynGAP1 missense variant I159L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. ESM1b is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for I159L, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -7.936 | In-Between | 0.195 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -0.39 | Neutral | 0.904 | Possibly Damaging | 0.847 | Possibly Damaging | 4.04 | Benign | 0.00 | Affected | 0.0776 | 0.2652 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.613A>C | I205L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I205L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools uniformly indicate a benign effect: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the variant as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, the SGM Consensus predicts likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. No contradictory evidence exists. Based on the collective predictions, the variant is most likely benign, and this assessment does not conflict with the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.264545 | Structured | 0.409933 | Uncertain | 0.821 | 0.414 | 0.125 | -5.474 | Likely Benign | 0.105 | Likely Benign | Likely Benign | 0.07 | Likely Benign | 0.2 | 0.02 | Likely Benign | 0.05 | Likely Benign | 0.30 | Likely Benign | 0.112 | Likely Benign | -0.86 | Neutral | 0.004 | Benign | 0.012 | Benign | 4.17 | Benign | 0.34 | Tolerated | 0.0682 | 0.2702 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||
| c.616A>C | I206L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I206L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and Foldetta. Those that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. Uncertain or inconclusive results come from premPS, AlphaMissense‑Optimized, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta predicts a benign outcome. Overall, the majority of tools (seven benign vs two pathogenic) support a benign classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.298791 | Structured | 0.405123 | Uncertain | 0.863 | 0.391 | 0.125 | -9.488 | Likely Pathogenic | 0.790 | Likely Pathogenic | Ambiguous | 0.05 | Likely Benign | 0.1 | 0.75 | Ambiguous | 0.40 | Likely Benign | 0.78 | Ambiguous | 0.080 | Likely Benign | -1.72 | Neutral | 0.004 | Benign | 0.012 | Benign | 3.78 | Benign | 0.08 | Tolerated | 0.0632 | 0.3608 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.637A>C | I213L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I213L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM all classify the variant as benign, while AlphaMissense‑Default predicts it as pathogenic and ESM1b remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores the variant benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign outcome; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, also reports a benign effect. No prediction or stability result is missing or inconclusive. Overall, the variant is most likely benign based on the collective evidence, and this benign prediction does not contradict the ClinVar status, which simply lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.158265 | Structured | 0.372201 | Uncertain | 0.850 | 0.295 | 0.125 | -7.673 | In-Between | 0.577 | Likely Pathogenic | Likely Benign | 0.29 | Likely Benign | 0.4 | 0.20 | Likely Benign | 0.25 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -1.45 | Neutral | 0.447 | Benign | 0.177 | Benign | 5.89 | Benign | 0.18 | Tolerated | 0.0771 | 0.2857 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.727A>C | I243L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I243L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; the only inconclusive results come from AlphaMissense‑Default and ESM1b, which are treated as unavailable. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields benign; and Foldetta also predicts benign. Overall, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.363090 | Structured | 0.344471 | Uncertain | 0.842 | 0.347 | 0.000 | -7.890 | In-Between | 0.524 | Ambiguous | Likely Benign | -0.03 | Likely Benign | 0.2 | -0.35 | Likely Benign | -0.19 | Likely Benign | 0.32 | Likely Benign | 0.380 | Likely Benign | -0.33 | Neutral | 0.048 | Benign | 0.039 | Benign | 5.74 | Benign | 0.19 | Tolerated | 0.0626 | 0.2891 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.802A>C | I268L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I268L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Two tools give uncertain results: premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, and Foldetta indicates a benign folding‑stability outcome. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.216401 | Structured | 0.314336 | Uncertain | 0.951 | 0.264 | 0.000 | -6.544 | Likely Benign | 0.402 | Ambiguous | Likely Benign | 0.37 | Likely Benign | 0.1 | -0.07 | Likely Benign | 0.15 | Likely Benign | 0.96 | Ambiguous | 0.390 | Likely Benign | -1.84 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 1.65 | Pathogenic | 0.03 | Affected | 0.0635 | 0.3037 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.805A>C | I269L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I269L missense variant is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in silico predictors shows a predominance of benign calls: REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized all predict benign. Pathogenicity is suggested by polyPhen‑2 (HumDiv and HumVar) and FATHMM, while ESM1b and AlphaMissense‑Default remain uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, Foldetta predicts benign stability, and the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to an equal split between benign and pathogenic signals. Overall, the balance of evidence favors a benign effect for I269L, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.216401 | Structured | 0.343787 | Uncertain | 0.937 | 0.244 | 0.125 | -7.588 | In-Between | 0.470 | Ambiguous | Likely Benign | 0.00 | Likely Benign | 0.0 | 0.19 | Likely Benign | 0.10 | Likely Benign | 0.47 | Likely Benign | 0.348 | Likely Benign | -1.47 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 1.88 | Pathogenic | 0.28 | Tolerated | 0.0662 | 0.2817 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.805A>T | I269L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I269L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two consensus groups: eight tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, AlphaMissense‑Optimized) predict a benign effect, while three tools (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM) predict pathogenicity. Two tools (ESM1b, AlphaMissense‑Default) return uncertain results. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized classifies the variant as benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, also predicts a benign impact. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields no clear majority and is therefore unavailable as evidence. Overall, the preponderance of evidence supports a benign classification, which is consistent with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.216401 | Structured | 0.343787 | Uncertain | 0.937 | 0.244 | 0.125 | -7.588 | In-Between | 0.470 | Ambiguous | Likely Benign | 0.00 | Likely Benign | 0.0 | 0.19 | Likely Benign | 0.10 | Likely Benign | 0.47 | Likely Benign | 0.348 | Likely Benign | -1.47 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 1.88 | Pathogenic | 0.28 | Tolerated | 0.0662 | 0.2817 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1001A>G | K334R 2D 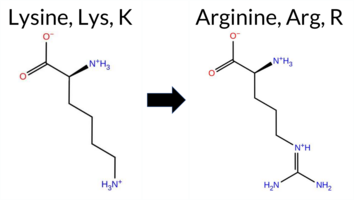 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K334R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Foldetta. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.377384 | Structured | 0.325972 | Uncertain | 0.544 | 0.414 | 0.500 | -5.384 | Likely Benign | 0.162 | Likely Benign | Likely Benign | -0.37 | Likely Benign | 0.1 | -0.53 | Ambiguous | -0.45 | Likely Benign | 0.39 | Likely Benign | 0.247 | Likely Benign | -2.62 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.76 | Pathogenic | 0.07 | Tolerated | 0.4647 | 0.0976 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.1007A>G | K336R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K336R is not reported in ClinVar (ClinVar status: not listed) but is present in the gnomAD database (gnomAD ID: 6‑33437912‑A‑G). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; the only tool predicting a pathogenic outcome is FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, the SGM‑Consensus is Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts Benign. No predictions or folding‑stability results are missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.318242 | Structured | 0.338219 | Uncertain | 0.396 | 0.428 | 0.500 | 6-33437912-A-G | 1 | 6.20e-7 | -5.897 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.00 | Likely Benign | 0.0 | -0.15 | Likely Benign | -0.08 | Likely Benign | 0.52 | Ambiguous | 0.038 | Likely Benign | -2.01 | Neutral | 0.002 | Benign | 0.005 | Benign | 1.69 | Pathogenic | 0.12 | Tolerated | 3.38 | 22 | 0.4899 | 0.1240 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||
| c.1010A>G | K337R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K337R has no ClinVar entry and is not reported in gnomAD. Prediction tools cluster into benign and pathogenic groups: benign predictions come from REVEL, FoldX, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Foldetta; pathogenic predictions come from polyPhen2_HumDiv, polyPhen2_HumVar, and FATHMM, while Rosetta remains uncertain. High‑accuracy methods reinforce the benign assessment: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts benign. Consequently, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.321458 | Structured | 0.348540 | Uncertain | 0.449 | 0.438 | 0.500 | -6.302 | Likely Benign | 0.249 | Likely Benign | Likely Benign | -0.30 | Likely Benign | 0.2 | 1.07 | Ambiguous | 0.39 | Likely Benign | 0.12 | Likely Benign | 0.142 | Likely Benign | -2.36 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 1.70 | Pathogenic | 0.07 | Tolerated | 0.3913 | 0.1134 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.1016A>G | K339R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K339R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. No predictions or stability results are missing or inconclusive. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.447574 | Structured | 0.384153 | Uncertain | 0.535 | 0.465 | 0.250 | -5.773 | Likely Benign | 0.120 | Likely Benign | Likely Benign | -0.15 | Likely Benign | 0.1 | 0.46 | Likely Benign | 0.16 | Likely Benign | 0.08 | Likely Benign | 0.134 | Likely Benign | -2.05 | Neutral | 0.046 | Benign | 0.040 | Benign | 1.94 | Pathogenic | 0.48 | Tolerated | 0.4197 | 0.0976 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.1175A>G | K392R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). All available in silico predictors classify the change as benign: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign effect. The high‑accuracy folding‑stability tool Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts a benign impact. No tool predicts pathogenicity. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.006 | Likely Benign | 0.091 | Likely Benign | Likely Benign | -0.03 | Likely Benign | 0.0 | 0.44 | Likely Benign | 0.21 | Likely Benign | 0.23 | Likely Benign | 0.131 | Likely Benign | -1.44 | Neutral | 0.436 | Benign | 0.112 | Benign | 7.12 | Benign | 0.08 | Tolerated | 0.5686 | 0.1875 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||
| c.1181A>G | K394R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K394R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate benign. Only SIFT predicts a pathogenic outcome, while Rosetta and Foldetta are uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, SGM‑Consensus is Likely Benign, and Foldetta remains uncertain. Overall, the preponderance of evidence supports a benign classification for K394R, and this conclusion does not contradict the absence of a ClinVar report. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | -4.902 | Likely Benign | 0.097 | Likely Benign | Likely Benign | -0.01 | Likely Benign | 0.1 | 1.19 | Ambiguous | 0.59 | Ambiguous | 0.42 | Likely Benign | 0.335 | Likely Benign | -1.97 | Neutral | 0.141 | Benign | 0.091 | Benign | 5.11 | Benign | 0.03 | Affected | 0.5488 | 0.2075 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||
| c.1208A>G | K403R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K403R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. The remaining tools—FoldX, ESM1b, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a tie, and Foldetta also predicts benign. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.275179 | Structured | 0.424920 | Uncertain | 0.960 | 0.372 | 0.000 | -7.362 | In-Between | 0.355 | Ambiguous | Likely Benign | -0.78 | Ambiguous | 0.1 | 0.23 | Likely Benign | -0.28 | Likely Benign | 0.41 | Likely Benign | 0.335 | Likely Benign | -2.56 | Deleterious | 0.983 | Probably Damaging | 0.926 | Probably Damaging | 3.91 | Benign | 0.15 | Tolerated | 0.4834 | 0.1861 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.1253A>G | K418R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K418R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, Foldetta, ESM1b, FATHMM, PROVEAN, and AlphaMissense‑Optimized. Tools that predict pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy consensus methods give a benign verdict: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign (3 benign vs. 1 pathogenic); and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. premPS is uncertain and therefore not considered evidence. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.104810 | Structured | 0.360134 | Uncertain | 0.948 | 0.263 | 0.000 | -6.809 | Likely Benign | 0.635 | Likely Pathogenic | Likely Benign | -0.18 | Likely Benign | 0.1 | 0.12 | Likely Benign | -0.03 | Likely Benign | 0.56 | Ambiguous | 0.229 | Likely Benign | -2.46 | Neutral | 0.994 | Probably Damaging | 0.962 | Probably Damaging | 3.37 | Benign | 0.04 | Affected | 0.4384 | 0.1151 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.1325A>G | K442R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K442R is catalogued in gnomAD (ID 6‑33438230‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; the only tools predicting a pathogenic outcome are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) classifies the variant as benign. No prediction or folding stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.170161 | Structured | 0.255766 | Uncertain | 0.912 | 0.225 | 0.000 | 6-33438230-A-G | 1 | 6.20e-7 | -5.761 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.13 | Likely Benign | 0.1 | 0.29 | Likely Benign | 0.21 | Likely Benign | 0.51 | Ambiguous | 0.098 | Likely Benign | -1.42 | Neutral | 0.972 | Probably Damaging | 0.875 | Possibly Damaging | 3.46 | Benign | 0.35 | Tolerated | 3.37 | 29 | 0.3885 | 0.0778 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||
| c.1331A>G | K444R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K444R is not reported in ClinVar and has no gnomAD allele. Prediction tools cluster into benign (REVEL, FoldX, SIFT, FATHMM, AlphaMissense‑Optimized) and pathogenic (SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default). Two tools (Rosetta, premPS) give uncertain results. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta predicts benign. No prediction or folding stability result is missing. Overall, six tools favor pathogenicity while five favor benignity, and the high‑accuracy consensus leans toward benign. Thus the variant is most likely pathogenic based on the broader set of predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.203355 | Structured | 0.262172 | Uncertain | 0.955 | 0.213 | 0.000 | -10.753 | Likely Pathogenic | 0.693 | Likely Pathogenic | Likely Benign | -0.12 | Likely Benign | 0.1 | -0.61 | Ambiguous | -0.37 | Likely Benign | 0.77 | Ambiguous | 0.213 | Likely Benign | -2.86 | Deleterious | 0.972 | Probably Damaging | 0.926 | Probably Damaging | 3.45 | Benign | 0.12 | Tolerated | 0.4597 | 0.0972 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.1379A>G | K460R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K460R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Foldetta, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate benign or likely benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict pathogenicity, while Rosetta and premPS are inconclusive. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, the SGM‑Consensus is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign stability. Taken together, the overwhelming majority of evidence supports a benign classification, and this is consistent with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.155435 | Structured | 0.289547 | Uncertain | 0.938 | 0.150 | 0.125 | -5.349 | Likely Benign | 0.175 | Likely Benign | Likely Benign | -0.41 | Likely Benign | 0.0 | 0.63 | Ambiguous | 0.11 | Likely Benign | 0.55 | Ambiguous | 0.103 | Likely Benign | -1.52 | Neutral | 0.991 | Probably Damaging | 0.962 | Probably Damaging | 3.46 | Benign | 0.63 | Tolerated | 0.5093 | 0.1169 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||
| c.1385A>G | K462R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K462R is catalogued in gnomAD (ID 6‑33438290‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Three tools (Rosetta, Foldetta, premPS) yield uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta is uncertain, so neither provides decisive evidence. Overall, the majority of reliable predictors lean toward a benign effect. This consensus does not contradict ClinVar status, which is currently absent. Thus, based on available predictions, K462R is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.264545 | Structured | 0.297737 | Uncertain | 0.921 | 0.159 | 0.125 | 6-33438290-A-G | 1 | 6.20e-7 | -9.980 | Likely Pathogenic | 0.184 | Likely Benign | Likely Benign | 0.13 | Likely Benign | 0.1 | 0.90 | Ambiguous | 0.52 | Ambiguous | 0.84 | Ambiguous | 0.246 | Likely Benign | -2.75 | Deleterious | 0.983 | Probably Damaging | 0.962 | Probably Damaging | 3.43 | Benign | 0.09 | Tolerated | 3.37 | 34 | 0.4995 | 0.1201 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||
| c.1475A>G | K492R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K492R is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta predicts benign. No prediction or folding stability result is missing or inconclusive. Overall, the majority of tools (seven pathogenic vs. six benign) lean toward a pathogenic interpretation, and this does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.061840 | Structured | 0.327121 | Uncertain | 0.947 | 0.192 | 0.000 | -11.290 | Likely Pathogenic | 0.599 | Likely Pathogenic | Likely Benign | -0.20 | Likely Benign | 0.1 | 0.45 | Likely Benign | 0.13 | Likely Benign | 0.84 | Ambiguous | 0.487 | Likely Benign | -2.98 | Deleterious | 0.997 | Probably Damaging | 0.990 | Probably Damaging | 3.25 | Benign | 0.05 | Affected | 0.3911 | 0.0835 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.1511A>G | K504R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K504R is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33438543‑A‑G). Consensus from most in‑silico predictors is benign: REVEL, FoldX, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all report a benign effect, while only FATHMM predicts pathogenicity. Uncertain calls come from Rosetta and premPS. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts benign. Overall, the preponderance of evidence indicates the variant is most likely benign, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.028107 | Structured | 0.304984 | Uncertain | 0.850 | 0.189 | 0.000 | Uncertain | 1 | 6-33438543-A-G | 2 | 1.24e-6 | -4.365 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.13 | Likely Benign | 0.1 | 0.51 | Ambiguous | 0.32 | Likely Benign | 0.94 | Ambiguous | 0.238 | Likely Benign | -2.16 | Neutral | 0.002 | Benign | 0.015 | Benign | -1.41 | Pathogenic | 0.11 | Tolerated | 3.37 | 35 | 0.3363 | 0.0647 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||
| c.1520A>G | K507R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K507R is not listed in ClinVar (ClinVar status: not reported) but is present in gnomAD (gnomAD ID 6-33438552-A-G). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No prediction or folding stability result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no pathogenic annotation for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.019401 | Structured | 0.262601 | Uncertain | 0.885 | 0.222 | 0.000 | 6-33438552-A-G | 1 | 6.20e-7 | -7.363 | In-Between | 0.085 | Likely Benign | Likely Benign | -0.08 | Likely Benign | 0.1 | 0.38 | Likely Benign | 0.15 | Likely Benign | 0.36 | Likely Benign | 0.503 | Likely Pathogenic | -0.68 | Neutral | 0.992 | Probably Damaging | 0.980 | Probably Damaging | -1.48 | Pathogenic | 0.27 | Tolerated | 3.37 | 35 | 0.3155 | 0.0579 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||
| c.1589A>G | K530R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K530R is catalogued in gnomAD (6‑33438832‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support benignity: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.308712 | Structured | 0.018455 | Uncertain | 0.891 | 0.409 | 0.000 | 6-33438832-A-G | 1 | 6.19e-7 | -3.836 | Likely Benign | 0.081 | Likely Benign | Likely Benign | -0.02 | Likely Benign | 0.1 | 0.18 | Likely Benign | 0.08 | Likely Benign | 0.33 | Likely Benign | 0.234 | Likely Benign | -1.39 | Neutral | 0.000 | Benign | 0.002 | Benign | -1.54 | Pathogenic | 0.08 | Tolerated | 3.37 | 35 | 0.3297 | 0.0716 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||
| c.1658A>G | K553R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K553R has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include FoldX, premPS, SIFT, AlphaMissense‑Optimized, and Foldetta. Tools that predict a pathogenic effect comprise SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. Rosetta’s output is uncertain and is treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicating likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta stability calculations) predicting a benign outcome. Overall, the majority of tools (nine pathogenic vs. five benign) lean toward a pathogenic interpretation, while the high‑accuracy consensus is split. Because ClinVar has no classification, there is no contradiction with existing clinical data. Based on the preponderance of predictions, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.012270 | Structured | 0.006539 | Uncertain | 0.949 | 0.246 | 0.000 | -8.182 | Likely Pathogenic | 0.644 | Likely Pathogenic | Likely Benign | -0.14 | Likely Benign | 0.2 | 0.63 | Ambiguous | 0.25 | Likely Benign | 0.38 | Likely Benign | 0.641 | Likely Pathogenic | -2.58 | Deleterious | 0.996 | Probably Damaging | 0.990 | Probably Damaging | -1.31 | Pathogenic | 0.13 | Tolerated | 0.3762 | 0.0835 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.1697A>G | K566R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K566R missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include premPS, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. FoldX, Rosetta, and Foldetta give uncertain or inconclusive results. High‑accuracy methods show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is unavailable. Overall, predictions are split evenly between benign and pathogenic, with no clear consensus. The variant is therefore most likely of uncertain significance; this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.027463 | Structured | 0.047887 | Uncertain | 0.924 | 0.219 | 0.000 | -8.493 | Likely Pathogenic | 0.279 | Likely Benign | Likely Benign | 1.26 | Ambiguous | 0.5 | 1.04 | Ambiguous | 1.15 | Ambiguous | 0.29 | Likely Benign | 0.555 | Likely Pathogenic | -2.39 | Neutral | 0.996 | Probably Damaging | 0.990 | Probably Damaging | -0.79 | Pathogenic | 0.43 | Tolerated | 0.4093 | 0.1099 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.1883A>G | K628R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K628R is reported in gnomAD (variant ID 6‑33440935‑A‑G) but has no ClinVar entry. Functional prediction tools show a split assessment: benign calls come from FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized, whereas pathogenic calls are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Two tools remain inconclusive (premPS and AlphaMissense‑Default). The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a likely pathogenic verdict, while the high‑accuracy AlphaMissense‑Optimized predicts benign. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, also predicts benign. Overall, the majority of evidence leans toward pathogenicity, and this conclusion does not conflict with ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.088832 | Structured | 0.035486 | Uncertain | 0.957 | 0.229 | 0.000 | 6-33440935-A-G | 1 | 6.20e-7 | -11.324 | Likely Pathogenic | 0.476 | Ambiguous | Likely Benign | 0.22 | Likely Benign | 0.1 | -0.11 | Likely Benign | 0.06 | Likely Benign | 0.94 | Ambiguous | 0.592 | Likely Pathogenic | -2.99 | Deleterious | 0.996 | Probably Damaging | 0.990 | Probably Damaging | 2.49 | Pathogenic | 0.00 | Affected | 3.37 | 34 | 0.3873 | 0.0873 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||
| c.1913A>G | K638R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K638R is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that uniformly indicate a benign effect include REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). In contrast, PROVEAN and polyPhen‑2 HumDiv predict a pathogenic impact, while premPS remains inconclusive. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, is benign. Overall, the majority of evidence points to a benign effect, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.045352 | Structured | 0.098064 | Uncertain | 0.937 | 0.260 | 0.000 | Uncertain | 1 | -2.700 | Likely Benign | 0.110 | Likely Benign | Likely Benign | 0.09 | Likely Benign | 0.1 | -0.04 | Likely Benign | 0.03 | Likely Benign | 0.53 | Ambiguous | 0.216 | Likely Benign | -2.55 | Deleterious | 0.649 | Possibly Damaging | 0.240 | Benign | 3.41 | Benign | 0.13 | Tolerated | 3.37 | 31 | 0.4026 | 0.0975 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||
| c.1925A>G | K642R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K642R is not reported in ClinVar and has no entries in gnomAD. Prediction tools that assess pathogenicity largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign. In contrast, PROVEAN, SIFT, and ESM1b predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, Foldetta is benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the balance of evidence favors a benign classification, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.061840 | Structured | 0.181468 | Uncertain | 0.806 | 0.289 | 0.000 | -10.562 | Likely Pathogenic | 0.192 | Likely Benign | Likely Benign | -0.13 | Likely Benign | 0.1 | 0.39 | Likely Benign | 0.13 | Likely Benign | 0.04 | Likely Benign | 0.319 | Likely Benign | -2.79 | Deleterious | 0.001 | Benign | 0.001 | Benign | 2.91 | Benign | 0.02 | Affected | 0.4962 | 0.1262 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.2114A>G | K705R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K705R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) all indicate a benign or likely benign outcome. Only two tools, polyPhen‑2 HumDiv and HumVar, predict a pathogenic effect. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign stability. Thus, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.134866 | Structured | 0.379324 | Uncertain | 0.922 | 0.364 | 0.000 | -4.679 | Likely Benign | 0.126 | Likely Benign | Likely Benign | 0.10 | Likely Benign | 0.0 | 0.35 | Likely Benign | 0.23 | Likely Benign | 0.18 | Likely Benign | 0.094 | Likely Benign | -1.66 | Neutral | 0.960 | Probably Damaging | 0.545 | Possibly Damaging | 3.33 | Benign | 0.06 | Tolerated | 0.3456 | 0.0789 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.2129A>G | K710R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K710R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments—all of which are available—show benign predictions: AlphaMissense‑Optimized is benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.321458 | Structured | 0.370438 | Uncertain | 0.949 | 0.368 | 0.000 | -6.295 | Likely Benign | 0.141 | Likely Benign | Likely Benign | 0.04 | Likely Benign | 0.0 | -0.14 | Likely Benign | -0.05 | Likely Benign | 0.29 | Likely Benign | 0.209 | Likely Benign | -2.78 | Deleterious | 0.983 | Probably Damaging | 0.962 | Probably Damaging | 3.44 | Benign | 0.02 | Affected | 0.3351 | 0.0789 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.2372A>G | K791R 2D AIThe SynGAP1 missense variant K791R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags the variant as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Benign. High‑accuracy assessments confirm this trend: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the preponderance of evidence supports a benign classification for K791R, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.966441 | Disordered | 0.478670 | Uncertain | 0.356 | 0.896 | 0.875 | -2.359 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.028 | Likely Benign | -0.96 | Neutral | 0.802 | Possibly Damaging | 0.249 | Benign | 4.14 | Benign | 0.50 | Tolerated | 0.5520 | 0.1223 | Weaken | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||
| c.2378A>G | K793R 2D AIThe SynGAP1 missense variant K793R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that assess sequence conservation and functional impact all converge on a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. No tool in the dataset indicates pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign,” while Foldetta results are unavailable. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.971072 | Disordered | 0.426071 | Uncertain | 0.344 | 0.901 | 0.875 | -2.789 | Likely Benign | 0.113 | Likely Benign | Likely Benign | 0.026 | Likely Benign | -0.29 | Neutral | 0.001 | Benign | 0.003 | Benign | 4.18 | Benign | 0.22 | Tolerated | 0.5270 | 0.1657 | Weaken | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||
| c.239A>G | K80R 2D AIThe SynGAP1 missense variant K80R is reported in gnomAD (ID 6‑33425847‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.477530 | Uncertain | 0.331 | 0.873 | 0.500 | 6-33425847-A-G | 1 | 6.20e-7 | -2.919 | Likely Benign | 0.146 | Likely Benign | Likely Benign | 0.077 | Likely Benign | 0.11 | Neutral | 0.001 | Benign | 0.001 | Benign | 4.33 | Benign | 0.00 | Affected | 4.32 | 1 | 0.4481 | 0.0858 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||
| c.2408A>G | K803R 2D AIThe SynGAP1 missense variant K803R is listed in ClinVar (ID 834618.0) with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—SIFT and FATHMM—predict pathogenicity. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates the variant is most likely benign, which does not contradict the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.827927 | Disordered | 0.733908 | Binding | 0.349 | 0.900 | 0.625 | Uncertain | 1 | -2.281 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.018 | Likely Benign | -1.52 | Neutral | 0.103 | Benign | 0.038 | Benign | 2.38 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.4857 | 0.1715 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||
| c.2498A>G | K833R 2D AIThe SynGAP1 missense variant K833R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Benign”). In contrast, the two polyPhen‑2 implementations (HumDiv and HumVar) predict a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also likely benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence indicates that K833R is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.625797 | Binding | 0.315 | 0.863 | 0.375 | -2.158 | Likely Benign | 0.114 | Likely Benign | Likely Benign | 0.094 | Likely Benign | -0.83 | Neutral | 0.997 | Probably Damaging | 0.925 | Probably Damaging | 2.61 | Benign | 0.11 | Tolerated | 0.3864 | 0.0945 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.2516A>G | K839R 2D AIThe SynGAP1 missense variant K839R is catalogued in gnomAD (ID 6‑33443068‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact, while ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect for K839R, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.611185 | Binding | 0.282 | 0.865 | 0.375 | 6-33443068-A-G | 1 | 6.20e-7 | -7.111 | In-Between | 0.162 | Likely Benign | Likely Benign | 0.133 | Likely Benign | -0.88 | Neutral | 0.972 | Probably Damaging | 0.860 | Possibly Damaging | 2.72 | Benign | 0.31 | Tolerated | 3.77 | 5 | 0.4823 | 0.1345 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||
| c.2951A>G | K984R 2D AIThe SynGAP1 missense variant K984R is reported in gnomAD (ID 6‑33443503‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of tools indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.951648 | Binding | 0.288 | 0.895 | 0.750 | 6-33443503-A-G | 2 | 1.24e-6 | -2.044 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.082 | Likely Benign | -0.41 | Neutral | 0.012 | Benign | 0.012 | Benign | 2.67 | Benign | 0.00 | Affected | 4.32 | 1 | 0.5098 | 0.1558 | Weaken | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||
| c.2981A>G | K994R 2D AIThe SynGAP1 missense variant K994R is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.930054 | Binding | 0.289 | 0.912 | 0.750 | -1.990 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.071 | Likely Benign | 0.02 | Neutral | 0.001 | Benign | 0.001 | Benign | 4.21 | Benign | 0.47 | Tolerated | 0.5064 | 0.1657 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3215A>G | K1072R 2D AIThe SynGAP1 missense variant K1072R is reported in gnomAD (ID 6‑33443767‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 (HumDiv and HumVar) predict it to be pathogenic. When predictions are grouped by consensus, the benign group contains seven tools, whereas the pathogenic group contains two. High‑accuracy assessments reinforce the benign view: AlphaMissense‑Optimized reports a benign outcome, and the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign effect. No Foldetta stability data are available, so folding‑stability evidence is unavailable. Overall, the majority of evidence points to a benign impact, and this is not in conflict with ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.922952 | Disordered | 0.984675 | Binding | 0.307 | 0.907 | 0.750 | 6-33443767-A-G | 1 | 6.23e-7 | -2.458 | Likely Benign | 0.100 | Likely Benign | Likely Benign | 0.116 | Likely Benign | -0.15 | Neutral | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 4.16 | Benign | 0.88 | Tolerated | 3.77 | 5 | 0.4655 | 0.2061 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||
| c.323A>G | K108R 2D AIThe SynGAP1 missense variant K108R is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33432188‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments further support benignity: AlphaMissense‑Optimized is benign, and the SGM‑Consensus itself is benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.626927 | Disordered | 0.673331 | Binding | 0.338 | 0.858 | 0.875 | Uncertain | 1 | 6-33432188-A-G | 6 | 3.72e-6 | -2.892 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.184 | Likely Benign | 0.37 | Neutral | 0.993 | Probably Damaging | 0.956 | Probably Damaging | 4.22 | Benign | 1.00 | Tolerated | 3.61 | 5 | 0.5286 | 0.1229 | Weaken | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||
| c.3248A>G | K1083R 2D AIThe SynGAP1 missense variant K1083R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. The variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.837511 | Disordered | 0.978906 | Binding | 0.302 | 0.893 | 1.000 | -2.212 | Likely Benign | 0.132 | Likely Benign | Likely Benign | 0.119 | Likely Benign | -0.53 | Neutral | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 4.06 | Benign | 0.84 | Tolerated | 0.5053 | 0.1903 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3332A>G | K1111R 2D AIThe SynGAP1 missense variant K1111R is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.921455 | Binding | 0.300 | 0.902 | 0.875 | -2.495 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.059 | Likely Benign | -0.30 | Neutral | 0.006 | Benign | 0.006 | Benign | 2.74 | Benign | 0.71 | Tolerated | 0.4664 | 0.2031 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.3389A>G | K1130R 2D AIThe SynGAP1 missense variant K1130R is reported in gnomAD (variant ID 6‑33443941‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority‑vote) is benign; Foldetta results are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.863782 | Binding | 0.350 | 0.904 | 0.750 | 6-33443941-A-G | -2.163 | Likely Benign | 0.155 | Likely Benign | Likely Benign | 0.187 | Likely Benign | -0.94 | Neutral | 0.014 | Benign | 0.008 | Benign | 5.44 | Benign | 0.00 | Affected | 4.32 | 4 | 0.5096 | 0.2074 | Weaken | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||
| c.3404A>G | K1135R 2D AIThe SynGAP1 missense variant K1135R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also reports Likely Benign. No Foldetta stability data are available, so it does not influence the assessment. Overall, the preponderance of evidence points to a benign impact for K1135R, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.790969 | Binding | 0.303 | 0.889 | 0.875 | -2.286 | Likely Benign | 0.122 | Likely Benign | Likely Benign | 0.209 | Likely Benign | -0.82 | Neutral | 0.586 | Possibly Damaging | 0.321 | Benign | 5.44 | Benign | 0.15 | Tolerated | 0.5095 | 0.1674 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.341A>G | K114R 2D AIThe SynGAP1 missense variant K114R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect; there is no conflict with ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -1.861 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.02 | Neutral | 0.005 | Benign | 0.003 | Benign | 4.09 | Benign | 0.00 | Affected | 0.5668 | 0.1498 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3536A>G | K1179R 2D AIThe SynGAP1 missense variant K1179R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available. Overall, the majority of evidence points to a benign effect for K1179R, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | -2.677 | Likely Benign | 0.178 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -0.92 | Neutral | 0.951 | Possibly Damaging | 0.628 | Possibly Damaging | 2.66 | Benign | 0.00 | Affected | 0.4010 | 0.0782 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3542A>G | K1181R 2D AIThe SynGAP1 missense variant K1181R is reported in gnomAD (variant ID 6‑33444577‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, representing the sole discordant prediction. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the preponderance of evidence indicates the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.622677 | Disordered | 0.539278 | Binding | 0.625 | 0.660 | 0.375 | 6-33444577-A-G | 2 | 1.24e-6 | -2.786 | Likely Benign | 0.185 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.90 | Neutral | 0.573 | Possibly Damaging | 0.429 | Benign | 2.67 | Benign | 0.07 | Tolerated | 4.32 | 3 | 0.3533 | 0.0878 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||
| c.3554A>G | K1185R 2D AIThe SynGAP1 missense variant K1185R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict pathogenicity. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for K1185R, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.566480 | Disordered | 0.510264 | Binding | 0.642 | 0.638 | 0.625 | -2.898 | Likely Benign | 0.224 | Likely Benign | Likely Benign | 0.130 | Likely Benign | -0.48 | Neutral | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 2.67 | Benign | 0.66 | Tolerated | 0.4476 | 0.0909 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3587A>G | K1196R 2D AIThe SynGAP1 missense variant K1196R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a neutral effect, and the SGM‑Consensus score is “Likely Benign.” No tool in the dataset predicts pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign effect. Foldetta results are unavailable, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.435699 | Uncertain | 0.851 | 0.595 | 0.250 | -2.077 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -0.78 | Neutral | 0.031 | Benign | 0.047 | Benign | 5.39 | Benign | 0.53 | Tolerated | 0.3970 | 0.0721 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3617A>G | K1206R 2D AIThe SynGAP1 missense variant K1206R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. AlphaMissense‑Default is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it receives two benign, one pathogenic, and one uncertain signal. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy AlphaMissense‑Optimized predicts a benign effect, while Foldetta data are missing. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.585406 | Disordered | 0.555819 | Binding | 0.893 | 0.569 | 0.375 | -6.357 | Likely Benign | 0.371 | Ambiguous | Likely Benign | 0.126 | Likely Benign | -1.64 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.49 | Pathogenic | 0.07 | Tolerated | 0.3897 | 0.0945 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.3644A>G | K1215R 2D AIThe SynGAP1 missense variant K1215R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the substitution as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact. The AlphaMissense‑Default score is uncertain, and no Foldetta stability assessment is available. High‑accuracy methods reinforce the benign prediction: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign, with no Foldetta data to contradict. Overall, the majority of evidence points to a benign effect, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.497853 | Structured | 0.503613 | Binding | 0.888 | 0.568 | 0.375 | -5.652 | Likely Benign | 0.360 | Ambiguous | Likely Benign | 0.106 | Likely Benign | -0.57 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.85 | Benign | 0.28 | Tolerated | 0.4051 | 0.0582 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3695A>G | K1232R 2D AIThe SynGAP1 missense variant K1232R is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of evidence—including the high‑accuracy tools—points to a benign impact. This conclusion is consistent with the absence of ClinVar annotation and gnomAD data, so there is no contradiction with existing clinical databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -3.146 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.077 | Likely Benign | -1.83 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.3515 | 0.0945 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3737A>G | K1246R 2D AIThe SynGAP1 missense variant K1246R is reported in gnomAD (ID 6‑33446729‑A‑G) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and the Foldetta stability analysis is unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.750527 | Disordered | 0.375382 | Uncertain | 0.887 | 0.564 | 0.625 | 6-33446729-A-G | 1 | 6.20e-7 | -2.444 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.026 | Likely Benign | -0.86 | Neutral | 0.031 | Benign | 0.017 | Benign | 2.66 | Benign | 0.04 | Affected | 3.77 | 5 | 0.3790 | 0.0515 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||
| c.3764A>G | K1255R 2D AIThe SynGAP1 K1255R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the change as damaging. The high‑accuracy consensus approach (SGM‑Consensus) – a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN – yields a pathogenic verdict (3 pathogenic vs. 1 benign). AlphaMissense‑Optimized is uncertain, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for K1255R. This prediction is consistent with the lack of ClinVar annotation; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -9.687 | Likely Pathogenic | 0.866 | Likely Pathogenic | Ambiguous | 0.171 | Likely Benign | -2.43 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.93 | Pathogenic | 0.00 | Affected | 0.3727 | 0.0878 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3779A>G | K1260R 2D AIThe SynGAP1 missense variant K1260R is catalogued in gnomAD (ID 6‑33446771‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; those that predict pathogenicity are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the collective predictions point to a benign impact, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | 6-33446771-A-G | 1 | 6.20e-7 | -6.033 | Likely Benign | 0.135 | Likely Benign | Likely Benign | 0.254 | Likely Benign | -2.29 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.40 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3655 | 0.0878 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||
| c.3863A>G | K1288R 2D AIThe SynGAP1 missense variant K1288R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | -2.367 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 0.141 | Likely Benign | -2.22 | Neutral | 0.979 | Probably Damaging | 0.973 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.4189 | 0.0782 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.3866A>G | K1289R 2D AIThe SynGAP1 missense variant K1289R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. Foldetta results are unavailable. Overall, the computational evidence strongly suggests the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.828700 | Binding | 0.548 | 0.787 | 0.625 | -2.127 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.090 | Likely Benign | -0.62 | Neutral | 0.001 | Benign | 0.003 | Benign | 2.66 | Benign | 0.11 | Tolerated | 0.4046 | 0.0715 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.413A>G | K138R 2D AIThe SynGAP1 missense variant K138R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta results are unavailable. Taken together, the preponderance of evidence points to a benign impact for K138R, and this conclusion does not conflict with the absence of a ClinVar assertion. The variant is most likely benign, and this assessment does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.619482 | Binding | 0.349 | 0.901 | 0.375 | -5.135 | Likely Benign | 0.406 | Ambiguous | Likely Benign | 0.115 | Likely Benign | -1.58 | Neutral | 0.247 | Benign | 0.163 | Benign | 3.66 | Benign | 0.02 | Affected | 0.4555 | 0.1036 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.425A>G | K142R 2D AIThe SynGAP1 missense variant K142R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors benign (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -9.327 | Likely Pathogenic | 0.361 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -1.87 | Neutral | 0.399 | Benign | 0.212 | Benign | 3.65 | Benign | 0.00 | Affected | 0.4459 | 0.0858 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||||
| c.434A>G | K145R 2D AIThe SynGAP1 missense variant K145R is listed in gnomAD (ID 6‑33432731‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign or likely benign. Only SIFT predicts a pathogenic outcome, while ESM1b remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status, as none is currently assigned to the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | 6-33432731-A-G | 1 | 6.20e-7 | -7.685 | In-Between | 0.214 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -1.58 | Neutral | 0.399 | Benign | 0.212 | Benign | 3.71 | Benign | 0.00 | Affected | 3.61 | 5 | 0.4661 | 0.1242 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||
| c.446A>G | K149R 2D AIThe SynGAP1 missense variant K149R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that K149R is most likely benign, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -8.237 | Likely Pathogenic | 0.406 | Ambiguous | Likely Benign | 0.143 | Likely Benign | -1.70 | Neutral | 0.247 | Benign | 0.125 | Benign | 3.70 | Benign | 0.00 | Affected | 0.5639 | 0.1169 | Weaken | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.533A>G | K178R 2D AIThe SynGAP1 missense variant K178R is reported in gnomAD (ID 6‑33435175‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the consensus score from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus also indicates likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact for K178R, and this conclusion is not contradicted by any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.455271 | Uncertain | 0.354 | 0.622 | 0.375 | 6-33435175-A-G | 1 | 6.20e-7 | -4.398 | Likely Benign | 0.281 | Likely Benign | Likely Benign | 0.117 | Likely Benign | -1.62 | Neutral | 0.905 | Possibly Damaging | 0.393 | Benign | 3.98 | Benign | 0.14 | Tolerated | 3.54 | 6 | 0.5403 | 0.1131 | Weaken | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||
| c.620A>G | K207R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense change K207R is not listed in ClinVar and has no reported allele in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign or likely benign outcome. In contrast, polyPhen‑2 HumDiv and HumVar classify the variant as pathogenic. Stability‑based methods (FoldX, Rosetta, Foldetta) are inconclusive, providing no definitive support for either outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta as uncertain. Taken together, the majority of evidence points to a benign effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.374039 | Structured | 0.406823 | Uncertain | 0.847 | 0.359 | 0.125 | -5.157 | Likely Benign | 0.217 | Likely Benign | Likely Benign | -0.58 | Ambiguous | 0.1 | -1.07 | Ambiguous | -0.83 | Ambiguous | 0.37 | Likely Benign | 0.140 | Likely Benign | -1.77 | Neutral | 0.982 | Probably Damaging | 0.679 | Possibly Damaging | 4.09 | Benign | 0.20 | Tolerated | 0.4640 | 0.1704 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.686A>G | K229R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K229R missense variant is catalogued in gnomAD (ID 6‑33435537‑A‑G) but has no entry in ClinVar. Functional prediction tools largely agree on a benign effect: FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all predict benign. In contrast, REVEL, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar predict pathogenic. AlphaMissense‑Default is uncertain and is treated as unavailable. The high‑accuracy consensus methods reinforce the benign assessment: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. Overall, the majority of evidence supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic report. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.179055 | Structured | 0.310912 | Uncertain | 0.843 | 0.306 | 0.000 | 6-33435537-A-G | 1 | 6.20e-7 | -5.651 | Likely Benign | 0.437 | Ambiguous | Likely Benign | 0.09 | Likely Benign | 0.0 | 0.02 | Likely Benign | 0.06 | Likely Benign | 0.40 | Likely Benign | 0.628 | Likely Pathogenic | -2.00 | Neutral | 0.993 | Probably Damaging | 0.971 | Probably Damaging | 5.84 | Benign | 0.19 | Tolerated | 3.43 | 12 | 0.4921 | 0.0972 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||
| c.722A>G | K241R 2D AIThe SynGAP1 missense variant K241R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote) is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. No prediction or folding stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.196879 | Structured | 0.349250 | Uncertain | 0.797 | 0.347 | 0.000 | -4.615 | Likely Benign | 0.444 | Ambiguous | Likely Benign | -0.23 | Likely Benign | 0.5 | -0.03 | Likely Benign | -0.13 | Likely Benign | 0.43 | Likely Benign | 0.602 | Likely Pathogenic | -2.12 | Neutral | 0.982 | Probably Damaging | 0.679 | Possibly Damaging | 5.89 | Benign | 0.14 | Tolerated | 0.4571 | 0.1242 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.752A>G | K251R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K251R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include FoldX, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Rosetta and Foldetta give uncertain results. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, SGM‑Consensus as Likely Benign, and Foldetta as Uncertain. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.447574 | Structured | 0.226632 | Uncertain | 0.758 | 0.312 | 0.125 | -4.355 | Likely Benign | 0.123 | Likely Benign | Likely Benign | -0.37 | Likely Benign | 0.0 | -0.64 | Ambiguous | -0.51 | Ambiguous | 0.14 | Likely Benign | 0.531 | Likely Pathogenic | -0.61 | Neutral | 0.970 | Probably Damaging | 0.681 | Possibly Damaging | 5.92 | Benign | 0.34 | Tolerated | 0.4756 | 0.1049 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.761A>G | K254R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K254R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority (2 benign vs. 1 pathogenic, 1 uncertain); Foldetta is uncertain. Taken together, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.513880 | Disordered | 0.207751 | Uncertain | 0.799 | 0.285 | 0.375 | -11.064 | Likely Pathogenic | 0.528 | Ambiguous | Likely Benign | -0.44 | Likely Benign | 0.1 | -1.13 | Ambiguous | -0.79 | Ambiguous | 0.76 | Ambiguous | 0.604 | Likely Pathogenic | -2.36 | Neutral | 0.970 | Probably Damaging | 0.681 | Possibly Damaging | 5.81 | Benign | 0.09 | Tolerated | 0.4070 | 0.1658 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.794A>G | K265R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K265R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster into two groups: benign predictions include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also reports benign. No prediction or stability result is missing or inconclusive. Based on the aggregate evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.209395 | Structured | 0.309758 | Uncertain | 0.936 | 0.275 | 0.000 | -3.366 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.04 | Likely Benign | 0.1 | -0.44 | Likely Benign | -0.20 | Likely Benign | 0.30 | Likely Benign | 0.223 | Likely Benign | -1.08 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.90 | Pathogenic | 0.36 | Tolerated | 0.4713 | 0.0976 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.830A>G | K277R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K277R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, premPS, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Rosetta gives an uncertain result. High‑accuracy methods give a benign prediction from AlphaMissense‑Optimized and from Foldetta; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it is a 2‑vs‑2 split. Overall, the majority of evidence (7 benign vs. 5 pathogenic) supports a benign classification. This consensus does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.061840 | Structured | 0.321811 | Uncertain | 0.649 | 0.247 | 0.250 | -6.652 | Likely Benign | 0.156 | Likely Benign | Likely Benign | -0.01 | Likely Benign | 0.1 | 0.53 | Ambiguous | 0.26 | Likely Benign | 0.06 | Likely Benign | 0.518 | Likely Pathogenic | -2.76 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.83 | Pathogenic | 0.06 | Tolerated | 0.4232 | 0.0678 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.833A>G | K278R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K278R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields an equal split of benign and pathogenic calls. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.120615 | Structured | 0.310130 | Uncertain | 0.748 | 0.253 | 0.125 | -5.313 | Likely Benign | 0.216 | Likely Benign | Likely Benign | -0.05 | Likely Benign | 0.0 | 0.18 | Likely Benign | 0.07 | Likely Benign | 0.13 | Likely Benign | 0.282 | Likely Benign | -2.66 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.73 | Pathogenic | 0.10 | Tolerated | 0.3926 | 0.0489 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.890A>G | K297R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K297R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar) and FATHMM. Predictions that are uncertain or inconclusive are FoldX, Rosetta, premPS, and ESM1b. High‑accuracy methods give a consistent benign signal: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign; and Foldetta, a protein‑folding stability approach combining FoldX‑MD and Rosetta, predicts benign. Based on the aggregate of these predictions, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.422041 | Structured | 0.272593 | Uncertain | 0.880 | 0.285 | 0.375 | -7.877 | In-Between | 0.314 | Likely Benign | Likely Benign | -0.86 | Ambiguous | 0.3 | 0.67 | Ambiguous | -0.10 | Likely Benign | 0.66 | Ambiguous | 0.257 | Likely Benign | -2.36 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.66 | Pathogenic | 0.19 | Tolerated | 0.4828 | 0.1506 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.998A>G | K333R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K333R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster into two groups: benign (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus “Likely Benign”) and pathogenic (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM). High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign”, and Foldetta (combining FoldX‑MD and Rosetta outputs) also indicates benign stability. No prediction or folding result is missing or inconclusive. Overall, the majority of evidence points to a benign effect for K333R, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.311707 | Structured | 0.330781 | Uncertain | 0.537 | 0.447 | 0.500 | -4.897 | Likely Benign | 0.120 | Likely Benign | Likely Benign | -0.23 | Likely Benign | 0.0 | 0.15 | Likely Benign | -0.04 | Likely Benign | 0.33 | Likely Benign | 0.232 | Likely Benign | -2.02 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.01 | Pathogenic | 0.14 | Tolerated | 0.4294 | 0.1134 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||
| c.1063G>T | G355W 2D 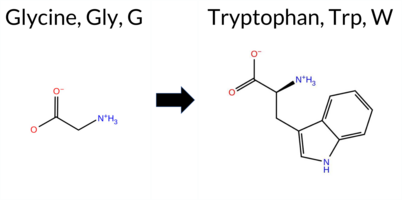 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G355W is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL and premPS, and pathogenic predictions from SIFT, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain results are reported by FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as inconclusive, SGM‑Consensus (derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as inconclusive. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not conflict with ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.155435 | Structured | 0.388832 | Uncertain | 0.810 | 0.354 | 0.125 | -12.316 | Likely Pathogenic | 0.938 | Likely Pathogenic | Ambiguous | 1.16 | Ambiguous | 0.5 | 0.88 | Ambiguous | 1.02 | Ambiguous | 0.18 | Likely Benign | 0.455 | Likely Benign | -6.92 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.73 | Pathogenic | 0.00 | Affected | 0.0796 | 0.4360 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||
| c.110C>A | S37Y 2D 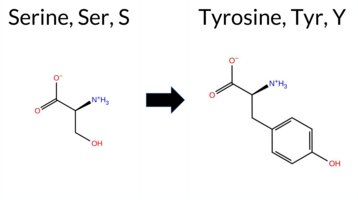 AIThe SynGAP1 missense variant S37Y is listed in gnomAD (ID 6‑33423519‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and Foldetta (FoldX‑MD/Rosetta stability assessment) has no available result. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta data is missing. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.433492 | Uncertain | 0.317 | 0.806 | 0.500 | 6-33423519-C-A | 1 | 6.20e-7 | -4.447 | Likely Benign | 0.370 | Ambiguous | Likely Benign | 0.132 | Likely Benign | -1.61 | Neutral | 0.880 | Possibly Damaging | 0.888 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1247 | 0.5256 | -2 | -3 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||
| c.1114G>T | G372W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G372W has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, FoldX, polyPhen‑2 HumDiv, SIFT, ESM1b, and FATHMM. The remaining tools (Rosetta, Foldetta, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.433034 | Structured | 0.430335 | Uncertain | 0.322 | 0.774 | 0.375 | -8.262 | Likely Pathogenic | 0.478 | Ambiguous | Likely Benign | 2.28 | Destabilizing | 0.5 | 1.14 | Ambiguous | 1.71 | Ambiguous | 0.21 | Likely Benign | 0.649 | Likely Pathogenic | -1.25 | Neutral | 0.657 | Possibly Damaging | 0.075 | Benign | -0.74 | Pathogenic | 0.00 | Affected | 0.1057 | 0.4368 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||
| c.1121C>A | S374Y 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant S374Y is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, whereas polyPhen‑2 HumDiv and SIFT predict a pathogenic impact. Uncertain calls come from FoldX, Rosetta, Foldetta, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome; the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive, and Foldetta likewise yields an uncertain stability change. Overall, the majority of available predictions favor a benign effect, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.642678 | Disordered | 0.428948 | Uncertain | 0.333 | 0.812 | 0.625 | Uncertain | 1 | -7.774 | In-Between | 0.344 | Ambiguous | Likely Benign | 0.71 | Ambiguous | 1.2 | 0.66 | Ambiguous | 0.69 | Ambiguous | -0.02 | Likely Benign | 0.310 | Likely Benign | -1.18 | Neutral | 0.875 | Possibly Damaging | 0.271 | Benign | 5.41 | Benign | 0.01 | Affected | 4.32 | 13 | 0.1175 | 0.6705 | -3 | -2 | -0.5 | 76.10 | 237.3 | -76.9 | 0.5 | 0.4 | 0.5 | 0.3 | Uncertain | Ser374 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because the Ω loop is assumed to directly interact with the membrane, it moves arbitrarily throughout the WT solvent simulations. The Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play major roles in protein functions that require flexibility, and thus, large and relatively hydrophobic residues like tyrosine are rarely tolerated. Additionally, the hydroxyl group of Tyr374 frequently forms various hydrogen bonds with other loop residues in the variant simulations. Although no negative structural effects are observed in the variant simulations, Tyr374 may exert drastic effects on the SynGAP-membrane complex dynamics and stability. However, since the effect on Gly-rich Ω loop dynamics can only be studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||||||
| c.1123G>T | G375W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G375W is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438028‑G‑T). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of evidence, including the high‑accuracy tools, points to a pathogenic effect for G375W. This conclusion is consistent with the absence of a ClinVar classification, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.604312 | Disordered | 0.428340 | Uncertain | 0.301 | 0.836 | 0.625 | 6-33438028-G-T | -9.654 | Likely Pathogenic | 0.464 | Ambiguous | Likely Benign | 4.33 | Destabilizing | 2.0 | 7.01 | Destabilizing | 5.67 | Destabilizing | 0.22 | Likely Benign | 0.450 | Likely Benign | -1.26 | Neutral | 0.992 | Probably Damaging | 0.869 | Possibly Damaging | 1.31 | Pathogenic | 0.01 | Affected | 4.32 | 12 | 0.0992 | 0.4368 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||||||
| c.1141G>T | G381W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G381W is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438046‑G‑T). Prediction tools that agree on a benign effect include premPS, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments give AlphaMissense‑Optimized a benign call, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) a pathogenic call, and Foldetta (combining FoldX‑MD and Rosetta outputs) a pathogenic call. No prediction or folding result is missing or inconclusive. Overall, the majority of tools and the high‑accuracy methods indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.431692 | Uncertain | 0.301 | 0.951 | 0.750 | 6-33438046-G-T | 1 | 6.24e-7 | -10.173 | Likely Pathogenic | 0.438 | Ambiguous | Likely Benign | 8.81 | Destabilizing | 2.0 | 3.17 | Destabilizing | 5.99 | Destabilizing | 0.02 | Likely Benign | 0.576 | Likely Pathogenic | -1.04 | Neutral | 0.996 | Probably Damaging | 0.920 | Probably Damaging | 1.31 | Pathogenic | 0.01 | Affected | 4.32 | 9 | 0.0993 | 0.4000 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||||
| c.1147G>T | G383W 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G383W is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438052‑G‑T). Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as benign, and Foldetta as pathogenic. Because the majority of conventional predictors favor pathogenicity while the high‑accuracy subset is split, the overall evidence leans toward a pathogenic interpretation. This conclusion does not conflict with the ClinVar uncertain status, which reflects the current lack of definitive clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | Uncertain | 1 | 6-33438052-G-T | 1 | 6.22e-7 | -10.161 | Likely Pathogenic | 0.439 | Ambiguous | Likely Benign | 5.81 | Destabilizing | 3.6 | 4.44 | Destabilizing | 5.13 | Destabilizing | 0.08 | Likely Benign | 0.469 | Likely Benign | -1.01 | Neutral | 0.959 | Probably Damaging | 0.704 | Possibly Damaging | 4.09 | Benign | 0.00 | Affected | 4.32 | 7 | 0.0972 | 0.3785 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||
| c.1156G>T | G386W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G386W is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Computational predictors that classify the change as benign include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. High‑accuracy assessments give a benign verdict from AlphaMissense‑Optimized, a benign consensus from the SGM method (majority of the four contributing tools are benign), and a pathogenic result from Foldetta. Uncertain calls from AlphaMissense‑Default and Rosetta are treated as unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not conflict with the lack of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.733139 | Disordered | 0.424156 | Uncertain | 0.334 | 0.898 | 0.750 | -10.389 | Likely Pathogenic | 0.519 | Ambiguous | Likely Benign | 6.07 | Destabilizing | 5.5 | 1.28 | Ambiguous | 3.68 | Destabilizing | -0.23 | Likely Benign | 0.471 | Likely Benign | -0.85 | Neutral | 0.996 | Probably Damaging | 0.920 | Probably Damaging | 3.90 | Benign | 0.00 | Affected | 0.1058 | 0.4676 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||
| c.1168G>T | G390W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G390W is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include AlphaMissense‑Optimized, premPS, and PROVEAN, whereas the remaining tools—REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts benign, but the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic verdict, and Foldetta also indicates pathogenic. With the majority of evidence pointing to a deleterious effect, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | -9.405 | Likely Pathogenic | 0.487 | Ambiguous | Likely Benign | 3.69 | Destabilizing | 1.9 | 5.31 | Destabilizing | 4.50 | Destabilizing | 0.14 | Likely Benign | 0.639 | Likely Pathogenic | -1.11 | Neutral | 0.987 | Probably Damaging | 0.744 | Possibly Damaging | 1.31 | Pathogenic | 0.00 | Affected | 0.0917 | 0.4368 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||
| c.11C>A | S4Y 2D AIThe SynGAP1 missense variant S4Y is reported in gnomAD (ID 6‑33420275‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign; Foldetta results are unavailable. Taken together, the majority of evidence points to a benign impact. There is no ClinVar classification to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.608892 | Disordered | 0.547364 | Binding | 0.390 | 0.924 | 0.750 | 6-33420275-C-A | -5.156 | Likely Benign | 0.209 | Likely Benign | Likely Benign | 0.099 | Likely Benign | -0.34 | Neutral | 0.880 | Possibly Damaging | 0.608 | Possibly Damaging | 4.12 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0748 | 0.6181 | -2 | -3 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||
| c.1412C>A | S471Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S471Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL, FoldX, premPS, and AlphaMissense‑Optimized; pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The high‑accuracy AlphaMissense‑Optimized scores the variant as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels it as likely pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, returns an uncertain stability change. No evidence from ClinVar contradicts these computational assessments. Overall, the preponderance of pathogenic predictions and the SGM Consensus suggest the variant is most likely pathogenic, consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.305330 | Structured | 0.355411 | Uncertain | 0.888 | 0.261 | 0.000 | -11.257 | Likely Pathogenic | 0.493 | Ambiguous | Likely Benign | -0.33 | Likely Benign | 0.1 | -1.31 | Ambiguous | -0.82 | Ambiguous | 0.19 | Likely Benign | 0.446 | Likely Benign | -4.79 | Deleterious | 0.980 | Probably Damaging | 0.584 | Possibly Damaging | -1.27 | Pathogenic | 0.03 | Affected | 0.0607 | 0.4417 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.1559C>A | S520Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S520Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include Rosetta and Foldetta, whereas the majority of algorithms predict pathogenicity: SGM‑Consensus (Likely Pathogenic), REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Two tools give inconclusive results: FoldX (Uncertain) and premPS (Uncertain). High‑accuracy assessments further support a mixed signal: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus indicates likely pathogenic, and Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts benign. Overall, the preponderance of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.094817 | Structured | 0.084894 | Uncertain | 0.887 | 0.337 | 0.000 | -13.124 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | -0.93 | Ambiguous | 0.4 | 0.16 | Likely Benign | -0.39 | Likely Benign | 0.59 | Ambiguous | 0.814 | Likely Pathogenic | -5.57 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | -1.36 | Pathogenic | 0.00 | Affected | 0.0743 | 0.4563 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.157G>T | G53W 2D AIThe SynGAP1 missense variant G53W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of available predictions (six pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.268042 | Structured | 0.460894 | Uncertain | 0.386 | 0.666 | 0.000 | -11.012 | Likely Pathogenic | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.243 | Likely Benign | -1.69 | Neutral | 0.998 | Probably Damaging | 0.993 | Probably Damaging | 4.08 | Benign | 0.00 | Affected | 0.0678 | 0.5071 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.1601C>A | S534Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S534Y has no ClinVar entry and is not reported in gnomAD. Prediction tools that classify it as benign include REVEL, FoldX, FATHMM, premPS, AlphaMissense‑Optimized, and Foldetta, while those that predict pathogenicity are SGM‑Consensus, PROVEAN, polyPhen2_HumDiv, polyPhen2_HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta as benign. Overall, the majority of tools (seven versus six) lean toward a pathogenic effect, and the high‑accuracy consensus is split, leaving the variant’s impact uncertain. Based on the aggregate predictions, the variant is most likely pathogenic, which does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.167087 | Structured | 0.032173 | Uncertain | 0.860 | 0.362 | 0.000 | -11.540 | Likely Pathogenic | 0.575 | Likely Pathogenic | Likely Benign | -0.01 | Likely Benign | 0.1 | 0.66 | Ambiguous | 0.33 | Likely Benign | 0.46 | Likely Benign | 0.314 | Likely Benign | -5.02 | Deleterious | 0.998 | Probably Damaging | 0.998 | Probably Damaging | 3.27 | Benign | 0.00 | Affected | 0.0629 | 0.5073 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.1670C>A | S557Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S557Y is not reported in ClinVar and has no gnomAD entry. Prediction tools largely agree on a deleterious effect: all except premPS (which predicts benign) return pathogenic or likely pathogenic. The high‑accuracy methods reinforce this: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels it likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenicity. No tool indicates a benign outcome. Consequently, the variant is most likely pathogenic according to the available computational evidence, and this assessment does not conflict with ClinVar status, which currently contains no entry for S557Y. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.028107 | Structured | 0.010261 | Uncertain | 0.924 | 0.215 | 0.000 | -13.792 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 18.89 | Destabilizing | 1.1 | 3.48 | Destabilizing | 11.19 | Destabilizing | -0.20 | Likely Benign | 0.965 | Likely Pathogenic | -5.45 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | -1.77 | Pathogenic | 0.00 | Affected | 0.1157 | 0.6087 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.1825G>T | G609W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G609W (ClinVar ID 4327030) is not found in gnomAD. Prediction tools that classify the variant as benign include premPS and AlphaMissense‑Optimized, whereas the majority of other in silico predictors (SGM‑Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, ESM1b, FATHMM) predict it to be pathogenic; Rosetta, Foldetta, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence from pathogenic‑oriented tools and the SGM Consensus supports a pathogenic effect, which is consistent with the ClinVar classification and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.179055 | Structured | 0.203786 | Uncertain | 0.851 | 0.252 | 0.000 | 1 | -13.074 | Likely Pathogenic | 0.525 | Ambiguous | Likely Benign | 3.14 | Destabilizing | 0.7 | -0.87 | Ambiguous | 1.14 | Ambiguous | 0.28 | Likely Benign | 0.566 | Likely Pathogenic | -4.70 | Deleterious | 0.999 | Probably Damaging | 0.976 | Probably Damaging | -1.47 | Pathogenic | 0.01 | Affected | 0.0935 | 0.3181 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||
| c.1910C>A | S637Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S637Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include REVEL, FoldX, premPS, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta as uncertain. Overall, the majority of individual predictors (seven versus five) lean toward pathogenicity, and the consensus‑based SGM‑Consensus also supports a likely pathogenic classification. Therefore, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.076542 | Structured | 0.083482 | Uncertain | 0.920 | 0.253 | 0.000 | -12.633 | Likely Pathogenic | 0.770 | Likely Pathogenic | Likely Benign | 0.13 | Likely Benign | 0.1 | 1.52 | Ambiguous | 0.83 | Ambiguous | 0.50 | Likely Benign | 0.209 | Likely Benign | -3.78 | Deleterious | 0.985 | Probably Damaging | 0.681 | Possibly Damaging | 3.35 | Benign | 0.00 | Affected | 0.0968 | 0.4071 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.1976C>A | S659Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S659Y is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default; FoldX is uncertain and therefore not considered evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of individual predictors lean toward a benign impact, and there is no ClinVar entry to contradict this assessment. Thus, based on the available predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.154597 | Uncertain | 0.954 | 0.283 | 0.000 | -10.832 | Likely Pathogenic | 0.600 | Likely Pathogenic | Likely Benign | -0.64 | Ambiguous | 0.1 | 0.00 | Likely Benign | -0.32 | Likely Benign | 0.29 | Likely Benign | 0.173 | Likely Benign | -4.24 | Deleterious | 0.084 | Benign | 0.014 | Benign | 3.38 | Benign | 0.04 | Affected | 0.0993 | 0.5962 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.2003C>A | S668Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S668Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include premPS and FATHMM, whereas the remaining 12 tools (REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) predict it to be pathogenic or likely pathogenic. High‑accuracy methods give consistent results: AlphaMissense‑Optimized scores it as pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenicity. No predictions are missing or inconclusive. Based on the overwhelming majority of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.247041 | Structured | 0.084935 | Uncertain | 0.922 | 0.370 | 0.000 | -14.833 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 18.34 | Destabilizing | 6.2 | 12.69 | Destabilizing | 15.52 | Destabilizing | 0.06 | Likely Benign | 0.641 | Likely Pathogenic | -5.95 | Deleterious | 0.999 | Probably Damaging | 0.935 | Probably Damaging | 3.18 | Benign | 0.00 | Affected | 0.0647 | 0.5693 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.2069C>A | S690Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S690Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL, premPS, and FATHMM, whereas the majority of other in silico predictors (FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) classify the variant as pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus confirms a Likely Pathogenic status, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a pathogenic effect. Based on the preponderance of pathogenic predictions and the absence of benign consensus, the variant is most likely pathogenic, with no contradiction to ClinVar status (which has no entry for this variant). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.055536 | Structured | 0.247926 | Uncertain | 0.944 | 0.253 | 0.000 | -14.051 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 11.45 | Destabilizing | 3.1 | 3.02 | Destabilizing | 7.24 | Destabilizing | 0.16 | Likely Benign | 0.381 | Likely Benign | -5.76 | Deleterious | 0.999 | Probably Damaging | 0.935 | Probably Damaging | 3.39 | Benign | 0.00 | Affected | 0.0512 | 0.4643 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.20C>A | S7Y 2D AIThe SynGAP1 missense variant S7Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. The predictions do not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.548467 | Binding | 0.386 | 0.922 | 0.750 | -5.240 | Likely Benign | 0.230 | Likely Benign | Likely Benign | 0.193 | Likely Benign | -0.36 | Neutral | 0.561 | Possibly Damaging | 0.047 | Benign | 4.06 | Benign | 0.00 | Affected | 0.0708 | 0.5053 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2248G>T | G750W 2D AIThe SynGAP1 missense variant G750W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, while the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all classify the variant as damaging. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as pathogenic, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) is unavailable for this variant. Overall, the preponderance of evidence from multiple in silico predictors indicates that G750W is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.618285 | Disordered | 0.646832 | Binding | 0.348 | 0.866 | 0.625 | -8.433 | Likely Pathogenic | 0.427 | Ambiguous | Likely Benign | 0.135 | Likely Benign | -3.30 | Deleterious | 1.000 | Probably Damaging | 0.994 | Probably Damaging | 2.41 | Pathogenic | 0.00 | Affected | 0.0738 | 0.4009 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.227C>A | S76Y 2D AIThe SynGAP1 missense variant S76Y is listed in ClinVar (ID 4801334) with an uncertain significance status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely benign. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence points to a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.517562 | Disordered | 0.444487 | Uncertain | 0.279 | 0.826 | 0.500 | Uncertain | 1 | -4.243 | Likely Benign | 0.209 | Likely Benign | Likely Benign | 0.119 | Likely Benign | -2.35 | Neutral | 0.972 | Probably Damaging | 0.831 | Possibly Damaging | 3.71 | Benign | 0.00 | Affected | 0.0540 | 0.4753 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||
| c.2297C>A | S766Y 2D AIThe SynGAP1 missense variant S766Y is reported in gnomAD (ID 6‑33442455‑C‑A) but has no ClinVar entry. Functional prediction tools show mixed results: benign calls come from REVEL, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts benign, but the SGM‑Consensus (majority vote) predicts pathogenic, and no Foldetta stability data are available. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.405110 | Structured | 0.923125 | Binding | 0.338 | 0.874 | 0.250 | 6-33442455-C-A | -8.636 | Likely Pathogenic | 0.641 | Likely Pathogenic | Likely Benign | 0.222 | Likely Benign | -2.67 | Deleterious | 0.990 | Probably Damaging | 0.856 | Possibly Damaging | 4.09 | Benign | 0.00 | Affected | 3.64 | 6 | 0.0794 | 0.5609 | -2 | -3 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||
| c.2312C>A | S771Y 2D AIThe SynGAP1 missense variant S771Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools, ESM1b and AlphaMissense‑Default, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it contains two benign and two uncertain calls, and Foldetta data are unavailable. Overall, the balance of evidence favors a benign interpretation, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.436924 | Structured | 0.922503 | Binding | 0.306 | 0.883 | 0.250 | -7.041 | In-Between | 0.397 | Ambiguous | Likely Benign | 0.176 | Likely Benign | -2.13 | Neutral | 0.990 | Probably Damaging | 0.892 | Possibly Damaging | 4.02 | Benign | 0.05 | Affected | 0.0707 | 0.5155 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2339C>A | S780Y 2D AIThe SynGAP1 missense variant S780Y is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.812415 | Binding | 0.283 | 0.883 | 0.500 | -7.682 | In-Between | 0.656 | Likely Pathogenic | Likely Benign | 0.091 | Likely Benign | -1.71 | Neutral | 0.995 | Probably Damaging | 0.925 | Probably Damaging | 2.61 | Benign | 0.11 | Tolerated | 0.0810 | 0.6428 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2363C>A | S788Y 2D AIThe SynGAP1 missense variant S788Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score—consistently predict a pathogenic or likely pathogenic impact. High‑accuracy assessments show that the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic effect, AlphaMissense‑Optimized is uncertain (treated as unavailable), and Foldetta results are not provided. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | SH3-binding motif | 0.956248 | Disordered | 0.573557 | Binding | 0.349 | 0.895 | 0.750 | -8.745 | Likely Pathogenic | 0.795 | Likely Pathogenic | Ambiguous | 0.251 | Likely Benign | -4.56 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.0844 | 0.5551 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||
| c.2447C>A | S816Y 2D AIThe SynGAP1 missense variant S816Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default, all of which classify the substitution as deleterious. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized yields an uncertain result, while Foldetta data are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus, the variant is most likely pathogenic; this assessment does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.747189 | Binding | 0.347 | 0.898 | 0.375 | -8.369 | Likely Pathogenic | 0.880 | Likely Pathogenic | Ambiguous | 0.222 | Likely Benign | -2.83 | Deleterious | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.60 | Benign | 0.03 | Affected | 0.0668 | 0.4741 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2525C>A | S842Y 2D AIThe SynGAP1 missense variant S842Y is listed in ClinVar as Pathogenic (ClinVar ID 624244.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized returns a pathogenic score, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, in agreement with its ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.604312 | Disordered | 0.617281 | Binding | 0.274 | 0.861 | 0.250 | Likely Pathogenic | 1 | -16.124 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.191 | Likely Benign | -4.28 | Deleterious | 0.944 | Possibly Damaging | 0.676 | Possibly Damaging | 1.97 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0576 | 0.5403 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||
| c.2548G>T | G850W 2D AIThe SynGAP1 missense variant G850W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign, two uncertain), and Foldetta data are unavailable. Overall, the balance of evidence (four benign vs. three pathogenic predictions, with a benign high‑accuracy result and no contradictory ClinVar annotation) indicates that the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.648219 | Disordered | 0.540897 | Binding | 0.312 | 0.820 | 0.500 | -7.826 | In-Between | 0.351 | Ambiguous | Likely Benign | 0.170 | Likely Benign | -1.94 | Neutral | 0.996 | Probably Damaging | 0.933 | Probably Damaging | 4.16 | Benign | 0.00 | Affected | 0.0734 | 0.4002 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.2599G>T | G867W 2D AIThe SynGAP1 missense variant G867W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains pathogenic; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.517562 | Disordered | 0.657954 | Binding | 0.285 | 0.801 | 0.250 | -8.983 | Likely Pathogenic | 0.776 | Likely Pathogenic | Likely Benign | 0.163 | Likely Benign | -2.95 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.64 | Benign | 0.01 | Affected | 0.0726 | 0.4584 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.260C>A | S87Y 2D AIThe SynGAP1 missense variant S87Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy predictors (six out of nine) indicate a pathogenic impact, whereas three predict benign. Therefore, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.550904 | Binding | 0.302 | 0.878 | 0.500 | -10.410 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.052 | Likely Benign | -2.20 | Neutral | 0.880 | Possibly Damaging | 0.608 | Possibly Damaging | 3.75 | Benign | 0.00 | Affected | 0.0522 | 0.4798 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2644G>T | G882W 2D AIThe SynGAP1 missense variant G882W has no ClinVar entry and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains pathogenic; Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not conflict with the ClinVar status, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.690604 | Disordered | 0.632888 | Binding | 0.306 | 0.878 | 0.250 | -8.637 | Likely Pathogenic | 0.606 | Likely Pathogenic | Likely Benign | 0.166 | Likely Benign | -2.35 | Neutral | 0.983 | Probably Damaging | 0.813 | Possibly Damaging | 2.01 | Pathogenic | 0.00 | Affected | 0.0829 | 0.4616 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.2675C>A | S892Y 2D AIThe SynGAP1 missense variant S892Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote) also predicts benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.473390 | Uncertain | 0.319 | 0.926 | 0.875 | -4.696 | Likely Benign | 0.645 | Likely Pathogenic | Likely Benign | 0.113 | Likely Benign | -2.29 | Neutral | 0.998 | Probably Damaging | 0.959 | Probably Damaging | 2.55 | Benign | 0.00 | Affected | 0.0814 | 0.5051 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2680G>T | G894W 2D AIThe SynGAP1 missense variant G894W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs. two benign votes). Foldetta, which assesses protein‑folding stability, has no available output for this variant. Overall, the majority of evaluated predictors (five pathogenic vs. three benign) lean toward a pathogenic interpretation. Because there is no ClinVar annotation to contradict this assessment, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.788093 | Disordered | 0.425700 | Uncertain | 0.310 | 0.925 | 0.750 | -6.927 | Likely Benign | 0.814 | Likely Pathogenic | Ambiguous | 0.177 | Likely Benign | -2.70 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.59 | Benign | 0.00 | Affected | 0.0698 | 0.4184 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.2693C>A | S898Y 2D AISynGAP1 missense variant S898Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Overall, the majority of tools and the consensus prediction indicate a pathogenic effect, and this conclusion does not contradict any ClinVar annotation because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.690604 | Disordered | 0.426070 | Uncertain | 0.305 | 0.922 | 0.500 | -5.927 | Likely Benign | 0.712 | Likely Pathogenic | Likely Benign | 0.182 | Likely Benign | -2.69 | Deleterious | 0.998 | Probably Damaging | 0.959 | Probably Damaging | 2.44 | Pathogenic | 0.00 | Affected | 0.1244 | 0.6838 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.274G>T | G92W 2D AIThe SynGAP1 missense variant G92W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) suggest a pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.575842 | Disordered | 0.537848 | Binding | 0.337 | 0.874 | 0.625 | -6.502 | Likely Benign | 0.789 | Likely Pathogenic | Ambiguous | 0.224 | Likely Benign | -2.61 | Deleterious | 1.000 | Probably Damaging | 0.979 | Probably Damaging | 3.95 | Benign | 0.00 | Affected | 0.0640 | 0.4546 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.2777C>A | S926Y 2D AIThe SynGAP1 missense variant S926Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic outcome. High‑accuracy assessments reinforce this trend: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports it as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for S926Y, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.981753 | Binding | 0.295 | 0.854 | 0.250 | -6.602 | Likely Benign | 0.968 | Likely Pathogenic | Likely Pathogenic | 0.444 | Likely Benign | -4.53 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.0602 | 0.4932 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2812G>T | G938W 2D AIThe SynGAP1 missense variant G938W is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar annotation exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.905695 | Disordered | 0.949795 | Binding | 0.318 | 0.883 | 0.625 | -8.763 | Likely Pathogenic | 0.604 | Likely Pathogenic | Likely Benign | 0.194 | Likely Benign | -1.71 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.70 | Benign | 0.04 | Affected | 0.0762 | 0.4008 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.2839G>T | G947W 2D AIThe SynGAP1 missense variant G947W has no ClinVar record and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988695 | Disordered | 0.850554 | Binding | 0.358 | 0.919 | 0.750 | -10.819 | Likely Pathogenic | 0.285 | Likely Benign | Likely Benign | 0.349 | Likely Benign | -1.21 | Neutral | 0.983 | Probably Damaging | 0.868 | Possibly Damaging | 4.90 | Benign | 0.01 | Affected | 0.0914 | 0.4059 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.2864C>A | S955Y 2D AIThe SynGAP1 missense variant S955Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of predictions (six benign vs. four pathogenic) support a benign classification. There is no ClinVar entry to contradict this assessment, so the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984871 | Disordered | 0.945325 | Binding | 0.350 | 0.924 | 0.750 | -6.212 | Likely Benign | 0.216 | Likely Benign | Likely Benign | 0.077 | Likely Benign | -1.62 | Neutral | 0.977 | Probably Damaging | 0.721 | Possibly Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.1321 | 0.4932 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2867C>A | S956Y 2D AIThe SynGAP1 missense variant S956Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Thus, the variant is most likely benign, with no contradiction from ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984871 | Disordered | 0.957345 | Binding | 0.364 | 0.917 | 0.750 | -4.920 | Likely Benign | 0.233 | Likely Benign | Likely Benign | 0.085 | Likely Benign | -0.90 | Neutral | 0.411 | Benign | 0.097 | Benign | 1.93 | Pathogenic | 0.22 | Tolerated | 0.1449 | 0.4932 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2917G>T | G973W 2D AIThe SynGAP1 missense variant G973W is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized reports Benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for G973W, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.959498 | Binding | 0.390 | 0.897 | 0.625 | -6.896 | Likely Benign | 0.329 | Likely Benign | Likely Benign | 0.105 | Likely Benign | -1.53 | Neutral | 0.983 | Probably Damaging | 0.813 | Possibly Damaging | 4.10 | Benign | 0.00 | Affected | 0.0846 | 0.4046 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.2966C>A | S989Y 2D AIThe SynGAP1 missense variant S989Y is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. High‑accuracy predictions therefore indicate a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus is benign, and Foldetta data are unavailable. Consequently, the variant is most likely benign based on the current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.795062 | Disordered | 0.908835 | Binding | 0.296 | 0.911 | 0.750 | -4.774 | Likely Benign | 0.471 | Ambiguous | Likely Benign | 0.103 | Likely Benign | -3.32 | Deleterious | 0.986 | Probably Damaging | 0.876 | Possibly Damaging | 2.60 | Benign | 0.00 | Affected | 0.0611 | 0.4815 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2969C>A | S990Y 2D AIThe SynGAP1 missense variant S990Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, SGM‑Consensus as Likely Benign, and Foldetta results are unavailable. Taken together, the majority of evidence points to a benign impact. The predictions do not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.902387 | Binding | 0.301 | 0.919 | 0.750 | -4.272 | Likely Benign | 0.314 | Likely Benign | Likely Benign | 0.131 | Likely Benign | -2.52 | Deleterious | 0.832 | Possibly Damaging | 0.500 | Possibly Damaging | 2.74 | Benign | 0.00 | Affected | 0.0893 | 0.5832 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2971G>T | G991W 2D AIThe SynGAP1 missense variant G991W is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the overall assessment. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.911393 | Binding | 0.286 | 0.920 | 0.750 | -6.281 | Likely Benign | 0.336 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -2.32 | Neutral | 0.997 | Probably Damaging | 0.975 | Probably Damaging | 4.07 | Benign | 0.00 | Affected | 0.0745 | 0.3654 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.2996C>A | S999Y 2D AIThe SynGAP1 missense variant S999Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for S999Y, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.950682 | Binding | 0.262 | 0.897 | 0.625 | -6.446 | Likely Benign | 0.346 | Ambiguous | Likely Benign | 0.069 | Likely Benign | -1.74 | Neutral | 0.934 | Possibly Damaging | 0.559 | Possibly Damaging | 2.64 | Benign | 0.00 | Affected | 0.0881 | 0.6249 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.3038C>A | S1013Y 2D AIThe SynGAP1 missense variant S1013Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.899570 | Binding | 0.308 | 0.846 | 0.625 | -4.882 | Likely Benign | 0.338 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.78 | Neutral | 0.290 | Benign | 0.124 | Benign | 2.66 | Benign | 0.03 | Affected | 0.0954 | 0.5629 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.3098C>A | S1033Y 2D AIThe SynGAP1 missense variant S1033Y is reported in gnomAD (ID 6‑33443650‑C‑A) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.993473 | Binding | 0.294 | 0.737 | 0.625 | 6-33443650-C-A | 1 | 6.19e-7 | -4.857 | Likely Benign | 0.564 | Ambiguous | Likely Benign | 0.034 | Likely Benign | -1.01 | Neutral | 0.021 | Benign | 0.008 | Benign | 2.69 | Benign | 0.04 | Affected | 3.77 | 5 | 0.0708 | 0.5262 | -2 | -3 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||
| c.3142G>T | G1048W 2D AIThe SynGAP1 missense variant G1048W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Benign”). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.977651 | Disordered | 0.923876 | Binding | 0.346 | 0.916 | 0.750 | -8.803 | Likely Pathogenic | 0.340 | Likely Benign | Likely Benign | 0.498 | Likely Benign | -1.52 | Neutral | 0.996 | Probably Damaging | 0.961 | Probably Damaging | 2.54 | Benign | 0.02 | Affected | 0.0852 | 0.4046 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.3154G>T | G1052W 2D AIThe SynGAP1 missense variant G1052W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984420 | Disordered | 0.892068 | Binding | 0.367 | 0.938 | 0.875 | -11.322 | Likely Pathogenic | 0.309 | Likely Benign | Likely Benign | 0.457 | Likely Benign | -0.90 | Neutral | 0.997 | Probably Damaging | 0.946 | Probably Damaging | 3.90 | Benign | 0.02 | Affected | 0.0872 | 0.4046 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.3163G>T | G1055W 2D AIThe SynGAP1 missense variant G1055W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.982235 | Disordered | 0.872113 | Binding | 0.379 | 0.935 | 0.875 | -11.436 | Likely Pathogenic | 0.317 | Likely Benign | Likely Benign | 0.367 | Likely Benign | -0.77 | Neutral | 0.997 | Probably Damaging | 0.946 | Probably Damaging | 3.28 | Benign | 0.01 | Affected | 0.0899 | 0.4246 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.3175G>T | G1059W 2D AIThe SynGAP1 missense variant G1059W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.983019 | Disordered | 0.898939 | Binding | 0.399 | 0.926 | 0.875 | -11.549 | Likely Pathogenic | 0.312 | Likely Benign | Likely Benign | 0.446 | Likely Benign | -1.18 | Neutral | 0.983 | Probably Damaging | 0.813 | Possibly Damaging | 2.53 | Benign | 0.00 | Affected | 0.0925 | 0.4046 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.3184G>T | G1062W 2D AIThe SynGAP1 missense variant G1062W is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which labels the variant as “Likely Benign.” Pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b, all of which classify the change as damaging. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.976962 | Disordered | 0.936972 | Binding | 0.368 | 0.917 | 0.875 | -9.667 | Likely Pathogenic | 0.315 | Likely Benign | Likely Benign | 0.401 | Likely Benign | -1.38 | Neutral | 0.993 | Probably Damaging | 0.890 | Possibly Damaging | 4.09 | Benign | 0.00 | Affected | 0.0908 | 0.4246 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.31G>T | G11W 2D AIThe SynGAP1 missense variant G11W is catalogued in gnomAD (ID 6‑33420295‑G‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Pathogenic predictions come from polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.444081 | Structured | 0.501027 | Binding | 0.348 | 0.915 | 0.375 | 6-33420295-G-T | -5.819 | Likely Benign | 0.403 | Ambiguous | Likely Benign | 0.096 | Likely Benign | -0.67 | Neutral | 0.959 | Probably Damaging | 0.318 | Benign | 3.87 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0747 | 0.4731 | -2 | -7 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||
| c.3218C>A | S1073Y 2D AIThe SynGAP1 missense change S1073Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.916840 | Disordered | 0.985818 | Binding | 0.313 | 0.905 | 0.750 | -6.768 | Likely Benign | 0.752 | Likely Pathogenic | Likely Benign | 0.165 | Likely Benign | -2.43 | Neutral | 0.990 | Probably Damaging | 0.796 | Possibly Damaging | 3.81 | Benign | 0.00 | Affected | 0.0977 | 0.5684 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.3260C>A | S1087Y 2D AIThe SynGAP1 missense variant S1087Y is catalogued in gnomAD (ID 6‑33443812‑C‑A) and has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as benign, and no Foldetta stability result is available. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.974805 | Binding | 0.357 | 0.891 | 1.000 | 6-33443812-C-A | -4.194 | Likely Benign | 0.587 | Likely Pathogenic | Likely Benign | 0.107 | Likely Benign | -2.41 | Neutral | 0.990 | Probably Damaging | 0.796 | Possibly Damaging | 2.56 | Benign | 0.02 | Affected | 3.77 | 5 | 0.0671 | 0.5845 | -2 | -3 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||
| c.3265G>T | G1089W 2D AIThe SynGAP1 missense variant G1089W is not reported in ClinVar and has no allele in gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the consensus SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments give an uncertain result from AlphaMissense‑Optimized, a Likely Pathogenic verdict from the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and no available Foldetta stability data. Overall, the majority of evidence points to a deleterious effect, so the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.976771 | Binding | 0.366 | 0.890 | 1.000 | -6.561 | Likely Benign | 0.863 | Likely Pathogenic | Ambiguous | 0.236 | Likely Benign | -3.45 | Deleterious | 1.000 | Probably Damaging | 0.988 | Probably Damaging | 2.37 | Pathogenic | 0.00 | Affected | 0.0840 | 0.4610 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.3281C>A | S1094Y 2D AIThe SynGAP1 missense variant S1094Y is not reported in ClinVar (status: none) but is present in gnomAD (ID 6‑33443833‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic). Foldetta, a protein‑folding‑stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions therefore show a benign call from AlphaMissense‑Optimized, an inconclusive SGM Consensus, and no Foldetta data. Overall, the balance of evidence leans toward a pathogenic interpretation (five pathogenic versus four benign calls), and this does not contradict the ClinVar status, which currently has no assertion for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.938133 | Disordered | 0.981352 | Binding | 0.358 | 0.877 | 1.000 | 6-33443833-C-A | -5.609 | Likely Benign | 0.631 | Likely Pathogenic | Likely Benign | 0.155 | Likely Benign | -2.07 | Neutral | 0.990 | Probably Damaging | 0.856 | Possibly Damaging | 2.45 | Pathogenic | 0.02 | Affected | 3.77 | 5 | 0.1007 | 0.5877 | -2 | -3 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||
| c.3346G>T | G1116W 2D AIThe SynGAP1 missense variant G1116W is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443898‑G‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b all predict a pathogenic outcome. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta’s protein‑folding stability result is unavailable. Taken together, the majority of reliable predictors and the consensus high‑accuracy tools indicate a benign effect. This conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.889439 | Disordered | 0.873279 | Binding | 0.320 | 0.909 | 0.750 | 6-33443898-G-T | -9.448 | Likely Pathogenic | 0.356 | Ambiguous | Likely Benign | 0.405 | Likely Benign | -1.27 | Neutral | 0.996 | Probably Damaging | 0.946 | Probably Damaging | 4.04 | Benign | 0.01 | Affected | 4.32 | 2 | 0.0776 | 0.4046 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||
| c.334G>T | G112W 2D AIThe SynGAP1 missense variant G112W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic interpretation. This conclusion is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.728858 | Disordered | 0.640153 | Binding | 0.332 | 0.867 | 0.750 | -6.382 | Likely Benign | 0.720 | Likely Pathogenic | Likely Benign | 0.190 | Likely Benign | -3.98 | Deleterious | 0.983 | Probably Damaging | 0.778 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.0631 | 0.4212 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.3355G>T | G1119W 2D AIThe SynGAP1 missense variant G1119W is reported in gnomAD (ID 6‑33443907‑G‑T) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign (three benign votes versus one pathogenic). High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta results are unavailable. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.818538 | Binding | 0.339 | 0.928 | 0.875 | 6-33443907-G-T | -10.904 | Likely Pathogenic | 0.336 | Likely Benign | Likely Benign | 0.386 | Likely Benign | -1.19 | Neutral | 0.997 | Probably Damaging | 0.949 | Probably Damaging | 3.90 | Benign | 0.02 | Affected | 4.32 | 2 | 0.0948 | 0.4059 | -2 | -7 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||
| c.3373G>T | G1125W 2D AIThe SynGAP1 missense variant G1125W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence, especially from the high‑accuracy tools, points to a benign classification. This conclusion is not contradicted by any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.835839 | Binding | 0.339 | 0.923 | 0.875 | -11.684 | Likely Pathogenic | 0.372 | Ambiguous | Likely Benign | 0.402 | Likely Benign | -1.10 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 4.50 | Benign | 0.00 | Affected | 0.0924 | 0.4046 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.3379G>T | G1127W 2D AIThe SynGAP1 missense variant G1127W is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443931‑G‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b all predict a pathogenic outcome; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta’s protein‑folding stability result is unavailable. Overall, the balance of evidence, particularly from the high‑accuracy tools, indicates that the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.852422 | Binding | 0.344 | 0.915 | 0.875 | 6-33443931-G-T | 3 | 2.01e-6 | -9.850 | Likely Pathogenic | 0.418 | Ambiguous | Likely Benign | 0.394 | Likely Benign | -1.36 | Neutral | 0.983 | Probably Damaging | 0.665 | Possibly Damaging | 4.79 | Benign | 0.03 | Affected | 4.32 | 4 | 0.0782 | 0.4032 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||
| c.337G>T | G113W 2D AIThe SynGAP1 missense variant G113W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence (six benign versus four pathogenic predictions) supports a benign classification. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.639486 | Binding | 0.350 | 0.870 | 0.750 | -5.635 | Likely Benign | 0.620 | Likely Pathogenic | Likely Benign | 0.162 | Likely Benign | -2.44 | Neutral | 0.983 | Probably Damaging | 0.717 | Possibly Damaging | 4.10 | Benign | 0.01 | Affected | 0.0700 | 0.4462 | -7 | -2 | -0.5 | 129.16 | |||||||||||||||||||||||||||||||||||||||
| c.3382G>T | G1128W 2D AIThe SynGAP1 missense variant G1128W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields an inconclusive result (two benign, two uncertain), and Foldetta data are unavailable. Overall, the balance of evidence favors a benign classification. This conclusion does not contradict ClinVar status, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.896620 | Disordered | 0.865136 | Binding | 0.309 | 0.911 | 0.875 | -7.341 | In-Between | 0.402 | Ambiguous | Likely Benign | 0.464 | Likely Benign | -1.36 | Neutral | 0.011 | Benign | 0.008 | Benign | 4.36 | Benign | 0.02 | Affected | 0.0796 | 0.4540 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.3395C>A | S1132Y 2D AIThe SynGAP1 missense variant S1132Y is listed in ClinVar as a benign alteration (ClinVar ID 845357.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence supports a benign classification, which aligns with the ClinVar status and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.845506 | Binding | 0.289 | 0.894 | 0.750 | Likely Benign | 1 | -5.894 | Likely Benign | 0.392 | Ambiguous | Likely Benign | 0.401 | Likely Benign | -1.76 | Neutral | 0.500 | Possibly Damaging | 0.208 | Benign | 5.40 | Benign | 0.09 | Tolerated | 4.32 | 4 | 0.0889 | 0.5233 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||
| c.3413C>A | S1138Y 2D AISynGAP1 missense variant S1138Y is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33444448‑C‑A). Prediction tools that agree on benign impact include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. AlphaMissense‑Default remains uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is ambiguous but leans toward benign (2 benign vs 1 pathogenic, 1 uncertain). High‑accuracy methods further support a benign interpretation: AlphaMissense‑Optimized predicts benign, while Foldetta results are unavailable. Overall, the balance of evidence, especially from high‑accuracy tools, suggests the variant is most likely benign, which does not contradict the current ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.891961 | Disordered | 0.738250 | Binding | 0.346 | 0.869 | 1.000 | Uncertain | 2 | 6-33444448-C-A | 3 | 1.86e-6 | -6.610 | Likely Benign | 0.449 | Ambiguous | Likely Benign | 0.391 | Likely Benign | -2.51 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 5.41 | Benign | 0.05 | Affected | 4.32 | 4 | 0.1034 | 0.5449 | -2 | -3 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||
| c.3425C>A | S1142Y 2D AIThe SynGAP1 missense variant S1142Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of conventional predictors lean toward pathogenicity, but the single high‑accuracy benign prediction and the inconclusive consensus temper this view. Thus, the variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | -6.146 | Likely Benign | 0.569 | Likely Pathogenic | Likely Benign | 0.200 | Likely Benign | -3.39 | Deleterious | 0.971 | Probably Damaging | 0.876 | Possibly Damaging | 2.64 | Benign | 0.00 | Affected | 0.0849 | 0.5429 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.3452C>A | S1151Y 2D AIThe SynGAP1 missense variant S1151Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. AlphaMissense‑Optimized also predicts a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence tools and the consensus prediction favor a benign impact, and there is no ClinVar entry to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.741537 | Disordered | 0.805072 | Binding | 0.394 | 0.839 | 0.625 | -4.456 | Likely Benign | 0.642 | Likely Pathogenic | Likely Benign | 0.166 | Likely Benign | -0.87 | Neutral | 0.995 | Probably Damaging | 0.925 | Probably Damaging | 2.68 | Benign | 0.05 | Affected | 0.0778 | 0.5287 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.3476C>A | S1159Y 2D AIThe SynGAP1 missense variant S1159Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome; Foldetta results are unavailable. Overall, the majority of evidence (six benign predictions versus five pathogenic, plus a benign consensus) points to a likely benign impact for S1159Y. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.626927 | Disordered | 0.867068 | Binding | 0.343 | 0.846 | 0.375 | -5.665 | Likely Benign | 0.957 | Likely Pathogenic | Likely Pathogenic | 0.195 | Likely Benign | -1.57 | Neutral | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 2.65 | Benign | 0.07 | Tolerated | 0.0567 | 0.4923 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.3635C>A | S1212Y 2D AIThe SynGAP1 missense variant S1212Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the convergence of multiple prediction algorithms, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | -12.186 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 0.304 | Likely Benign | -4.55 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.03 | Pathogenic | 0.00 | Affected | 0.0546 | 0.4291 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||
| c.3674C>A | S1225Y 2D AIThe SynGAP1 missense variant S1225Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also leans toward benign (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.513880 | Disordered | 0.441915 | Uncertain | 0.891 | 0.544 | 0.500 | -8.360 | Likely Pathogenic | 0.359 | Ambiguous | Likely Benign | 0.419 | Likely Benign | -2.27 | Neutral | 0.975 | Probably Damaging | 0.767 | Possibly Damaging | 5.34 | Benign | 0.04 | Affected | 0.0547 | 0.4691 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.3770C>A | S1257Y 2D AIThe SynGAP1 missense variant S1257Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) leans pathogenic (2 pathogenic vs 1 benign). Foldetta results are unavailable. Overall, the majority of evaluated predictors (five pathogenic vs. three benign) indicate a pathogenic impact. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.541878 | Disordered | 0.482380 | Uncertain | 0.889 | 0.572 | 0.375 | -8.632 | Likely Pathogenic | 0.399 | Ambiguous | Likely Benign | 0.120 | Likely Benign | -2.65 | Deleterious | 0.989 | Probably Damaging | 0.883 | Possibly Damaging | 2.55 | Benign | 0.03 | Affected | 0.0480 | 0.4463 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.419C>A | S140Y 2D AIThe SynGAP1 missense variant S140Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default—predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the S140Y variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.587898 | Binding | 0.321 | 0.896 | 0.625 | -13.071 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.252 | Likely Benign | -3.88 | Deleterious | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 3.48 | Benign | 0.00 | Affected | 0.0597 | 0.5849 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.50C>A | S17Y 2D AIThe SynGAP1 missense variant S17Y is listed in gnomAD (ID 6‑33420314‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Pathogenic predictions come from polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign trend: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors benign. No Foldetta stability data are available, so it does not influence the conclusion. Overall, the majority of evidence indicates that S17Y is most likely benign, and this assessment does not contradict any ClinVar status, as none is reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | 6-33420314-C-A | -4.492 | Likely Benign | 0.551 | Ambiguous | Likely Benign | 0.050 | Likely Benign | -1.06 | Neutral | 0.742 | Possibly Damaging | 0.047 | Benign | 3.99 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0818 | 0.6366 | -2 | -3 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||
| c.545C>A | S182Y 2D AIThe SynGAP1 missense variant S182Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: pathogenic calls come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, while benign calls are made only by REVEL and FATHMM. High‑accuracy assessments reinforce the pathogenic prediction: AlphaMissense‑Optimized indicates a pathogenic effect, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. A protein‑folding stability analysis with Foldetta is unavailable, so it does not influence the overall assessment. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.501700 | Disordered | 0.436016 | Uncertain | 0.368 | 0.619 | 0.625 | -15.951 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.274 | Likely Benign | -4.40 | Deleterious | 0.940 | Possibly Damaging | 0.564 | Possibly Damaging | 3.64 | Benign | 0.00 | Affected | 0.0656 | 0.5158 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.554C>A | S185Y 2D AIThe SynGAP1 missense variant S185Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.545602 | Disordered | 0.430485 | Uncertain | 0.365 | 0.623 | 0.500 | -13.633 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.316 | Likely Benign | -4.93 | Deleterious | 0.718 | Possibly Damaging | 0.178 | Benign | 3.56 | Benign | 0.00 | Affected | 0.0736 | 0.5439 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.611C>A | S204Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S204Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions arise from FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy consensus (SGM Consensus) is inconclusive because it receives an equal split between pathogenic and benign calls. AlphaMissense‑Optimized, a high‑accuracy model, predicts benign, whereas Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts pathogenic. Overall, the majority of evidence points to a pathogenic effect, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.268042 | Structured | 0.420667 | Uncertain | 0.816 | 0.405 | 0.125 | -10.518 | Likely Pathogenic | 0.782 | Likely Pathogenic | Likely Benign | 8.99 | Destabilizing | 3.8 | 5.20 | Destabilizing | 7.10 | Destabilizing | -0.03 | Likely Benign | 0.170 | Likely Benign | -0.96 | Neutral | 0.978 | Probably Damaging | 0.694 | Possibly Damaging | 4.14 | Benign | 0.03 | Affected | 0.0452 | 0.5625 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||
| c.635C>A | S212Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S212Y is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include FoldX, FATHMM, and premPS. In contrast, a majority of tools predict a pathogenic impact: REVEL, SGM‑Consensus, AlphaMissense‑Default, AlphaMissense‑Optimized, ESM1b, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, is inconclusive for this variant. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as likely pathogenic; Foldetta remains uncertain. Overall, the preponderance of evidence from multiple in silico predictors indicates that S212Y is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.158265 | Structured | 0.381517 | Uncertain | 0.846 | 0.278 | 0.125 | -14.812 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.43 | Likely Benign | 0.5 | 2.32 | Destabilizing | 1.38 | Ambiguous | 0.22 | Likely Benign | 0.819 | Likely Pathogenic | -5.15 | Deleterious | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 5.76 | Benign | 0.00 | Affected | 0.0555 | 0.6120 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.704C>A | S235Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S235Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include FATHMM and Rosetta, whereas the majority of other in silico predictors (REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) classify the variant as pathogenic. Two tools give inconclusive results: Foldetta (protein‑folding stability) and premPS. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports “Likely Pathogenic,” and Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for S235Y, and this conclusion is not contradicted by any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.250310 | Structured | 0.319150 | Uncertain | 0.743 | 0.331 | 0.000 | -15.842 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 3.53 | Destabilizing | 1.8 | 0.43 | Likely Benign | 1.98 | Ambiguous | 0.66 | Ambiguous | 0.887 | Likely Pathogenic | -5.14 | Deleterious | 0.971 | Probably Damaging | 0.641 | Possibly Damaging | 5.45 | Benign | 0.00 | Affected | 0.0733 | 0.5269 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.83C>A | S28Y 2D AIThe SynGAP1 missense variant S28Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. There is no conflict with ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.545602 | Disordered | 0.438157 | Uncertain | 0.354 | 0.884 | 0.125 | -4.521 | Likely Benign | 0.270 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -0.94 | Neutral | 0.009 | Benign | 0.001 | Benign | 4.13 | Benign | 0.05 | Affected | 0.0895 | 0.5067 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.887C>A | S296Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S296Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, and premPS, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. FoldX and Foldetta give uncertain results. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic, and Foldetta remains uncertain. Overall, the majority of evidence points to a pathogenic impact. The variant is most likely pathogenic based on predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.444081 | Structured | 0.282669 | Uncertain | 0.887 | 0.284 | 0.250 | -12.148 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 1.46 | Ambiguous | 0.2 | 0.48 | Likely Benign | 0.97 | Ambiguous | 0.15 | Likely Benign | 0.486 | Likely Benign | -4.34 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.93 | Pathogenic | 0.05 | Affected | 0.0800 | 0.5610 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.899C>A | S300Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S300Y has no ClinVar entry and is not reported in gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Foldetta, premPS, and AlphaMissense‑Optimized; pathogenic predictions come from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Two tools (AlphaMissense‑Default and Rosetta) are uncertain and do not influence the consensus. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts likely pathogenic; Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of evidence points toward a pathogenic effect, and this assessment does not contradict any ClinVar status (none is available). Thus, based on current predictions, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.356642 | Structured | 0.256848 | Uncertain | 0.742 | 0.280 | 0.375 | -10.464 | Likely Pathogenic | 0.422 | Ambiguous | Likely Benign | -0.08 | Likely Benign | 0.4 | 0.55 | Ambiguous | 0.24 | Likely Benign | 0.28 | Likely Benign | 0.116 | Likely Benign | -2.52 | Deleterious | 0.975 | Probably Damaging | 0.686 | Possibly Damaging | 1.52 | Pathogenic | 0.01 | Affected | 0.0678 | 0.5395 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||
| c.905C>A | S302Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S302Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized, and Foldetta. Tools that predict a pathogenic effect are SIFT, polyPhen‑2 HumDiv, and ESM1b. Two tools give uncertain results: AlphaMissense‑Default and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.414856 | Structured | 0.263489 | Uncertain | 0.616 | 0.258 | 0.375 | -9.674 | Likely Pathogenic | 0.355 | Ambiguous | Likely Benign | -0.02 | Likely Benign | 0.1 | 0.56 | Ambiguous | 0.27 | Likely Benign | -0.17 | Likely Benign | 0.070 | Likely Benign | -1.03 | Neutral | 0.801 | Possibly Damaging | 0.383 | Benign | 4.07 | Benign | 0.01 | Affected | 0.0882 | 0.5990 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||
| c.907G>T | G303W 2D 3DClick to see structure in 3D Viewer AIClinVar reports no entry for this SynGAP1 G303W variant, and it is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta’s stability prediction is also unavailable. Overall, the evidence is evenly split between benign and pathogenic predictions, providing no clear bias toward either outcome. This balanced prediction does not contradict ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.450668 | Structured | 0.271087 | Uncertain | 0.630 | 0.254 | 0.250 | -9.041 | Likely Pathogenic | 0.579 | Likely Pathogenic | Likely Benign | 1.69 | Ambiguous | 0.3 | 1.11 | Ambiguous | 1.40 | Ambiguous | 0.28 | Likely Benign | 0.157 | Likely Benign | -1.63 | Neutral | 0.983 | Probably Damaging | 0.813 | Possibly Damaging | 3.93 | Benign | 0.00 | Affected | 0.0663 | 0.4553 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||
| c.1021G>A | G341S 2D 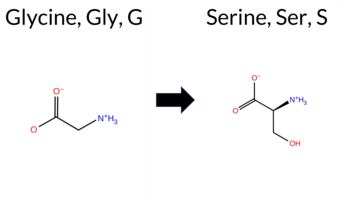 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G341S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, polyPhen‑2 HumVar, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. Predictions that are inconclusive or uncertain are Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM‑Consensus as Likely Benign, and Foldetta as Uncertain. Overall, the majority of evidence points to a benign impact for G341S, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.356642 | Structured | 0.431164 | Uncertain | 0.745 | 0.479 | 0.250 | -3.206 | Likely Benign | 0.090 | Likely Benign | Likely Benign | 0.41 | Likely Benign | 0.3 | -1.46 | Ambiguous | -0.53 | Ambiguous | -0.67 | Ambiguous | 0.343 | Likely Benign | 0.73 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 0.37 | Pathogenic | 0.72 | Tolerated | 0.2416 | 0.4215 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1027G>C | V343L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V343L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) all predict benign or likely benign. Only FATHMM predicts a pathogenic outcome, while FoldX and Rosetta provide uncertain results and are therefore not considered decisive. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also indicates benign. Overall, the majority of evidence supports a benign classification, and this is consistent with the absence of ClinVar reporting. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.291804 | Structured | 0.383911 | Uncertain | 0.882 | 0.497 | 0.250 | -6.268 | Likely Benign | 0.310 | Likely Benign | Likely Benign | -0.93 | Ambiguous | 0.2 | 0.66 | Ambiguous | -0.14 | Likely Benign | 0.16 | Likely Benign | 0.033 | Likely Benign | -1.09 | Neutral | 0.005 | Benign | 0.013 | Benign | 2.12 | Pathogenic | 0.64 | Tolerated | 0.1361 | 0.5008 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1030G>A | G344S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G344S is listed in ClinVar (ID 981240.0) as Pathogenic and is not reported in gnomAD. Prediction tools that assess pathogenicity all agree: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic; the only inconclusive result is premPS, which is marked Uncertain. No tool predicts a benign effect. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. Thus, the variant is most likely pathogenic, and this prediction aligns with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.264545 | Structured | 0.368110 | Uncertain | 0.913 | 0.485 | 0.250 | Pathogenic | 5 | -11.254 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 9.02 | Destabilizing | 0.7 | 6.08 | Destabilizing | 7.55 | Destabilizing | 0.98 | Ambiguous | 0.790 | Likely Pathogenic | -5.28 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -0.45 | Pathogenic | 0.04 | Affected | 3.37 | 25 | 0.2735 | 0.5324 | 1 | 0 | -0.4 | 30.03 | 217.3 | -51.7 | 0.0 | 0.1 | 0.2 | 0.1 | X | X | Potentially Pathogenic | Because Gly344 lacks a proper side chain, it allows the anti-parallel β sheet strand (res. Gly341-Pro349) to have a slight twist. Within a β strand, side chains normally alternate between outward and inward positions, but glycine is an exception as it allows the alternating pattern to skip a residue. Introducing serine or any other residue with a side chain at position 344 prevents this unique skip in the alternating pattern, causing structural strain or likely preventing correct folding altogether. Additionally, Tyr342 shields Gly344 from the solvent, contributing to twist formation in the β sheet and stabilizing the β-strand.In the variant simulations, the side chain of Ser344 assumes the inward position. However, the hydrophobic niche formed by multiple C2 domain residues (e.g., Val365, Val343, Leu327) is not accommodating for its hydroxyl group. The outward position, not seen in the simulations, would be equally disadvantageous due to the presence of hydrophobic residues on that side as well (e.g., Leu345, Tyr342). Serine is also not well-suited for twist formation, as it tends to suppress twisting and bending in β sheets. At this position, the hydroxyl group of Ser344 could also form hydrogen bonds with the backbone atoms of the Gly-rich Ω loop in the C2 domain (e.g., Thr366, Leu367, Gly378; res. Pro364-Pro398), potentially adversely affecting membrane-loop dynamics and ultimately compromising the stability of the SynGAP-membrane association. | |||||||||||||||
| c.1036G>C | V346L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V346L is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, and SIFT, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. Four tools (Foldetta, premPS, AlphaMissense‑Optimized, and Rosetta) give uncertain or inconclusive results and are not used as evidence. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of reliable predictors and the SGM‑Consensus support a pathogenic effect. Therefore, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.260850 | Structured | 0.350921 | Uncertain | 0.949 | 0.461 | 0.000 | -8.178 | Likely Pathogenic | 0.834 | Likely Pathogenic | Ambiguous | 0.05 | Likely Benign | 0.2 | 1.10 | Ambiguous | 0.58 | Ambiguous | 0.60 | Ambiguous | 0.384 | Likely Benign | -2.68 | Deleterious | 0.994 | Probably Damaging | 0.970 | Probably Damaging | 1.58 | Pathogenic | 0.09 | Tolerated | 0.1237 | 0.5348 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1036G>T | V346L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V346L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, and SIFT, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. Four tools (Foldetta, premPS, AlphaMissense‑Optimized, and Rosetta) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points toward a deleterious effect. Therefore, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.260850 | Structured | 0.350921 | Uncertain | 0.949 | 0.461 | 0.000 | -8.178 | Likely Pathogenic | 0.834 | Likely Pathogenic | Ambiguous | 0.05 | Likely Benign | 0.2 | 1.10 | Ambiguous | 0.58 | Ambiguous | 0.60 | Ambiguous | 0.386 | Likely Benign | -2.68 | Deleterious | 0.994 | Probably Damaging | 0.970 | Probably Damaging | 1.58 | Pathogenic | 0.09 | Tolerated | 0.1237 | 0.5348 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.103G>C | V35L 2D AIThe SynGAP1 missense variant V35L is reported in gnomAD (variant ID 6-33423512‑G‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence, including the high‑accuracy tools, points to a benign effect. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.613573 | Disordered | 0.434838 | Uncertain | 0.360 | 0.851 | 0.375 | 6-33423512-G-C | 4 | 2.48e-6 | -2.893 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.039 | Likely Benign | -0.58 | Neutral | 0.481 | Possibly Damaging | 0.506 | Possibly Damaging | 4.18 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1026 | 0.4025 | 1 | 2 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.1042G>C | V348L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V348L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign; and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No other high‑confidence tools provide conflicting evidence. Based on the preponderance of predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.170161 | Structured | 0.346556 | Uncertain | 0.951 | 0.414 | 0.000 | -4.842 | Likely Benign | 0.270 | Likely Benign | Likely Benign | -0.58 | Ambiguous | 0.0 | 0.04 | Likely Benign | -0.27 | Likely Benign | 0.24 | Likely Benign | 0.072 | Likely Benign | -0.75 | Neutral | 0.264 | Benign | 0.030 | Benign | 1.89 | Pathogenic | 0.62 | Tolerated | 0.1088 | 0.4953 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1042G>T | V348L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V348L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign; and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No other high‑confidence tools provide conflicting evidence. Based on the preponderance of predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.170161 | Structured | 0.346556 | Uncertain | 0.951 | 0.414 | 0.000 | -4.842 | Likely Benign | 0.270 | Likely Benign | Likely Benign | -0.58 | Ambiguous | 0.0 | 0.04 | Likely Benign | -0.27 | Likely Benign | 0.24 | Likely Benign | 0.072 | Likely Benign | -0.75 | Neutral | 0.264 | Benign | 0.030 | Benign | 1.89 | Pathogenic | 0.62 | Tolerated | 0.1088 | 0.4953 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1048G>C | V350L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V350L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. Only FATHMM predicts a pathogenic outcome, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign status. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta (combining FoldX‑MD and Rosetta stability outputs) as uncertain. No other tools provide conflicting evidence. Based on the preponderance of benign predictions and the lack of pathogenic support, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.179055 | Structured | 0.353227 | Uncertain | 0.931 | 0.371 | 0.000 | -5.941 | Likely Benign | 0.238 | Likely Benign | Likely Benign | -1.31 | Ambiguous | 0.1 | -0.34 | Likely Benign | -0.83 | Ambiguous | 0.36 | Likely Benign | 0.042 | Likely Benign | -1.63 | Neutral | 0.068 | Benign | 0.022 | Benign | 2.37 | Pathogenic | 0.27 | Tolerated | 0.1173 | 0.4958 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1048G>T | V350L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V350L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only FATHMM predicts a pathogenic outcome, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign status. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is inconclusive. No other tools provide conflicting evidence. Based on the preponderance of predictions, V350L is most likely benign, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.179055 | Structured | 0.353227 | Uncertain | 0.931 | 0.371 | 0.000 | -5.941 | Likely Benign | 0.238 | Likely Benign | Likely Benign | -1.31 | Ambiguous | 0.1 | -0.34 | Likely Benign | -0.83 | Ambiguous | 0.36 | Likely Benign | 0.042 | Likely Benign | -1.63 | Neutral | 0.068 | Benign | 0.022 | Benign | 2.37 | Pathogenic | 0.27 | Tolerated | 0.1173 | 0.4958 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1078G>A | E360K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E360K is reported in gnomAD (variant ID 6-33437983‑G‑A) but has no ClinVar entry. Prediction tools that agree on a benign effect are limited to FoldX, which scores the variant as benign. In contrast, the majority of algorithms predict a pathogenic impact: REVEL, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). Tools with inconclusive results (Foldetta and premPS) are noted as unavailable. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus indicates likely pathogenic, while Foldetta remains uncertain. Overall, the consensus of high‑confidence predictors points to a pathogenic effect for E360K. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.250310 | Structured | 0.421183 | Uncertain | 0.955 | 0.498 | 0.250 | 6-33437983-G-A | -16.006 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.27 | Likely Benign | 0.0 | 2.21 | Destabilizing | 1.24 | Ambiguous | 0.55 | Ambiguous | 0.526 | Likely Pathogenic | -3.68 | Deleterious | 0.997 | Probably Damaging | 0.980 | Probably Damaging | 1.68 | Pathogenic | 0.04 | Affected | 3.37 | 25 | 0.3106 | 0.8594 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||||
| c.1081C>A | Q361K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q361K is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors indicates a benign effect: REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the substitution as tolerated, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also classifies it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and FATHMM predict a pathogenic outcome. High‑accuracy tools that integrate structural and evolutionary information—AlphaMissense‑Optimized, the SGM‑Consensus, and Foldetta (combining FoldX‑MD and Rosetta outputs)—all return benign predictions. No prediction or folding‑stability result is missing or inconclusive. Based on the aggregate evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.305330 | Structured | 0.427593 | Uncertain | 0.945 | 0.534 | 0.250 | -5.147 | Likely Benign | 0.286 | Likely Benign | Likely Benign | 0.17 | Likely Benign | 0.0 | 0.36 | Likely Benign | 0.27 | Likely Benign | -0.44 | Likely Benign | 0.217 | Likely Benign | 1.29 | Neutral | 0.969 | Probably Damaging | 0.930 | Probably Damaging | 1.73 | Pathogenic | 1.00 | Tolerated | 0.1739 | 0.4701 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||
| c.1093G>C | V365L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V365L is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess sequence conservation and functional impact uniformly classify the substitution as benign: REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity; only FoldX and Rosetta report uncertain stability changes, which are treated as unavailable evidence. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta, which integrates FoldX‑MD and Rosetta outputs, reports a benign effect on protein folding stability. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.441505 | Uncertain | 0.923 | 0.608 | 0.250 | -6.141 | Likely Benign | 0.265 | Likely Benign | Likely Benign | -0.72 | Ambiguous | 0.2 | 0.74 | Ambiguous | 0.01 | Likely Benign | 0.35 | Likely Benign | 0.065 | Likely Benign | -1.71 | Neutral | 0.005 | Benign | 0.003 | Benign | 2.50 | Benign | 0.25 | Tolerated | 3.38 | 21 | 0.0852 | 0.5309 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||||
| c.1093G>T | V365L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V365L is catalogued in gnomAD (ID 6‑33437998‑G‑T) but has no ClinVar entry. Across the available in‑silico predictors, the majority (REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly classify the change as benign; only FoldX and Rosetta report uncertain stability effects, which are treated as unavailable evidence. High‑accuracy assessments reinforce this benign prediction: AlphaMissense‑Optimized is benign, the SGM Consensus (derived from the unanimous benign vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts a benign impact. Consequently, the variant is most likely benign based on current computational evidence, and this assessment does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.441505 | Uncertain | 0.923 | 0.608 | 0.250 | 6-33437998-G-T | 5 | 3.31e-6 | -6.141 | Likely Benign | 0.265 | Likely Benign | Likely Benign | -0.72 | Ambiguous | 0.2 | 0.74 | Ambiguous | 0.01 | Likely Benign | 0.35 | Likely Benign | 0.065 | Likely Benign | -1.71 | Neutral | 0.005 | Benign | 0.003 | Benign | 2.50 | Benign | 0.25 | Tolerated | 3.38 | 21 | 0.0852 | 0.5309 | 1 | 2 | -0.4 | 14.03 | ||||||||||||||||||||||||
| c.1108G>A | G370S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G370S is listed in ClinVar with an “Uncertain” status and is present in gnomAD (variant ID 6‑33438013‑G‑A). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Benign”). Tools that predict a pathogenic effect are FoldX and FATHMM. Predictions that are inconclusive are Rosetta and Foldetta. High‑accuracy methods give a benign signal: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is also benign; Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.461924 | Structured | 0.434325 | Uncertain | 0.359 | 0.720 | 0.500 | Uncertain | 2 | 6-33438013-G-A | 15 | 9.31e-6 | -3.533 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 2.83 | Destabilizing | 2.0 | 1.05 | Ambiguous | 1.94 | Ambiguous | -0.02 | Likely Benign | 0.282 | Likely Benign | 0.47 | Neutral | 0.000 | Benign | 0.000 | Benign | 1.33 | Pathogenic | 0.77 | Tolerated | 3.42 | 19 | 0.2664 | 0.5086 | 1 | 0 | -0.4 | 30.03 | 196.6 | -49.6 | 0.9 | 2.2 | -0.1 | 0.4 | Uncertain | Gly370 is located in the Gly-rich Ω loop (res. Pro364- Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because, the Ω loop is assumed to be directly interacting with the membrane, it is only seen to move arbitrarily throughout the WT solvent simulations. The Ω loop is potentially playing a crucial loop in the SynGAP-membrane complex association, stability and dynamics, regardless, this aspect cannot be addressed through the solvent simulations only. The Ω-loops are known to have a major role in protein functions that requires flexibility and thus, they are rich in glycines, prolines and to a lesser extent, hydrophilic residues to ensure maximum flexibility. Thus, Ser370 in the variant is potentially tolerated in the Ω loop. However, since the effect on the Gly-rich Ω loop dynamics can only be well-studied through the SynGAP-membrane complex, no definite conclusions can be withdrawn. | ||||||||||||||
| c.1126G>A | G376S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G376S is reported in gnomAD (ID 6‑33438031‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Pathogenic predictions arise from FoldX, polyPhen‑2 (HumDiv and HumVar), and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates benign, while Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. Overall, the majority of evidence points to a benign effect; this conclusion is not contradicted by ClinVar, which contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.680603 | Disordered | 0.428979 | Uncertain | 0.326 | 0.869 | 0.625 | 6-33438031-G-A | 1 | 6.21e-7 | -4.913 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 2.30 | Destabilizing | 0.5 | -0.45 | Likely Benign | 0.93 | Ambiguous | 0.32 | Likely Benign | 0.471 | Likely Benign | -0.73 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.33 | Pathogenic | 0.22 | Tolerated | 4.32 | 12 | 0.2831 | 0.4801 | 0 | 1 | -0.4 | 30.03 | ||||||||||||||||||||||||
| c.1132G>A | G378S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G378S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta as pathogenic, giving a mixed outcome. Because the predictions are evenly split and the high‑accuracy methods are contradictory, the variant’s functional impact is uncertain. Based on the available evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.767246 | Disordered | 0.432858 | Uncertain | 0.341 | 0.915 | 0.625 | -5.331 | Likely Benign | 0.115 | Likely Benign | Likely Benign | 6.75 | Destabilizing | 2.2 | 7.81 | Destabilizing | 7.28 | Destabilizing | 0.12 | Likely Benign | 0.565 | Likely Pathogenic | -0.63 | Neutral | 1.000 | Probably Damaging | 0.986 | Probably Damaging | 1.33 | Pathogenic | 0.08 | Tolerated | 0.3013 | 0.4724 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1150G>A | G384S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G384S (gnomAD ID 6-33438055‑G‑A) is listed in ClinVar with an uncertain significance. Functional prediction tools cluster into two groups: benign predictions from REVEL, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments further support benignity: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote) is likely benign, and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is inconclusive. No evidence from FoldX or Rosetta alone is available. Overall, the preponderance of evidence points to a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.728858 | Disordered | 0.427831 | Uncertain | 0.323 | 0.934 | 0.750 | Uncertain | 1 | 6-33438055-G-A | 1 | 6.22e-7 | -5.243 | Likely Benign | 0.090 | Likely Benign | Likely Benign | 1.92 | Ambiguous | 0.2 | 1.66 | Ambiguous | 1.79 | Ambiguous | 0.19 | Likely Benign | 0.315 | Likely Benign | -0.67 | Neutral | 0.980 | Probably Damaging | 0.968 | Probably Damaging | 1.33 | Pathogenic | 0.04 | Affected | 4.32 | 2 | 0.2905 | 0.4924 | 1 | 0 | -0.4 | 30.03 | 202.4 | -49.8 | 0.5 | 1.0 | -0.2 | 0.0 | Uncertain | Gly384 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because the Ω loop is assumed to directly interact with the membrane, it moves arbitrarily throughout the WT solvent simulations. The Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play major roles in protein functions that require flexibility, and so they are rich in glycines, prolines, and, to a lesser extent, small hydrophilic residues to ensure maximum flexibility. Thus, the variant’s Ser384 is potentially tolerated in the Ω loop, although the hydroxyl group of Ser384 forms various hydrogen bonds with several other loop residues in the variant simulations. However, since the effects on Gly-rich Ω loop dynamics can only be studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||
| c.1159G>A | G387S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 G387S missense variant is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (variant ID 6‑33438064‑G‑A). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX and FATHMM; Rosetta is uncertain. The high‑accuracy consensus (SGM‑Consensus) is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN and is therefore likely benign. AlphaMissense‑Optimized itself predicts benign. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts a pathogenic effect. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar classification (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.642678 | Disordered | 0.422910 | Uncertain | 0.293 | 0.861 | 0.750 | 6-33438064-G-A | -4.674 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 2.37 | Destabilizing | 0.5 | 1.94 | Ambiguous | 2.16 | Destabilizing | 0.12 | Likely Benign | 0.359 | Likely Benign | -0.12 | Neutral | 0.000 | Benign | 0.002 | Benign | 1.33 | Pathogenic | 0.13 | Tolerated | 4.32 | 3 | 0.3051 | 0.4724 | 0 | 1 | -0.4 | 30.03 | ||||||||||||||||||||||||||
| c.1162G>A | G388S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G388S is catalogued in gnomAD (ID 6‑33438067‑G‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that converge on a pathogenic interpretation are FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, while the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. In contrast, the protein‑folding stability predictor Foldetta, which integrates FoldX‑MD and Rosetta outputs, indicates a pathogenic effect. No prediction is available from Rosetta alone, and the SGM‑Consensus result aligns with the benign consensus. Overall, the majority of evidence points to a benign impact, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.736850 | Disordered | 0.420316 | Uncertain | 0.319 | 0.827 | 0.750 | 6-33438067-G-A | 1 | 6.21e-7 | -5.036 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 3.42 | Destabilizing | 3.5 | 1.69 | Ambiguous | 2.56 | Destabilizing | 0.18 | Likely Benign | 0.446 | Likely Benign | -0.52 | Neutral | 0.980 | Probably Damaging | 0.968 | Probably Damaging | 1.33 | Pathogenic | 0.05 | Affected | 4.32 | 3 | 0.2901 | 0.4985 | 0 | 1 | -0.4 | 30.03 | ||||||||||||||||||||||||
| c.1171G>A | G391S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G391S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and FoldX. Rosetta and Foldetta give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus also as likely benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact for G391S, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.637480 | Disordered | 0.409509 | Uncertain | 0.279 | 0.741 | 0.750 | -5.531 | Likely Benign | 0.096 | Likely Benign | Likely Benign | 2.14 | Destabilizing | 0.6 | 1.01 | Ambiguous | 1.58 | Ambiguous | 0.06 | Likely Benign | 0.462 | Likely Benign | -0.54 | Neutral | 0.978 | Probably Damaging | 0.777 | Possibly Damaging | 1.33 | Pathogenic | 0.35 | Tolerated | 0.2820 | 0.5097 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1177G>A | G393S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G393S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from FoldX, polyPhen‑2 HumDiv, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta (combining FoldX‑MD and Rosetta) as uncertain. No other tools provide decisive evidence. Overall, the majority of reliable predictors lean toward a benign effect, and this consensus does not conflict with the absence of ClinVar annotation. Therefore, G393S is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.538167 | Disordered | 0.402365 | Uncertain | 0.333 | 0.670 | 0.625 | -5.207 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 2.43 | Destabilizing | 0.7 | -0.78 | Ambiguous | 0.83 | Ambiguous | 0.24 | Likely Benign | 0.466 | Likely Benign | -1.76 | Neutral | 0.889 | Possibly Damaging | 0.444 | Benign | 1.33 | Pathogenic | 0.07 | Tolerated | 0.3011 | 0.5232 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1186G>A | G396S 2D AIThe SynGAP1 missense variant G396S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, the SGM‑Consensus is benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) is inconclusive. No tool predicts pathogenicity. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.394626 | Uncertain | 0.640 | 0.584 | 0.500 | -3.934 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 1.76 | Ambiguous | 1.6 | 1.12 | Ambiguous | 1.44 | Ambiguous | 0.10 | Likely Benign | 0.223 | Likely Benign | -0.99 | Neutral | 0.004 | Benign | 0.003 | Benign | 3.93 | Benign | 0.56 | Tolerated | 0.2668 | 0.4894 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1198G>C | V400L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V400L is listed in ClinVar as Benign (ClinVar ID 1166313) and is present in gnomAD (6‑33438103‑G‑C). Prediction tools that agree on a benign effect include REVEL, FoldX‑MD (uncertain), Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. Based on the collective predictions, the variant is most likely benign, and this conclusion is consistent with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.398279 | Structured | 0.415488 | Uncertain | 0.951 | 0.451 | 0.000 | Benign/Likely benign | 2 | 6-33438103-G-C | 22 | 1.36e-5 | -1.000 | Likely Benign | 0.137 | Likely Benign | Likely Benign | -0.71 | Ambiguous | 0.2 | 0.39 | Likely Benign | -0.16 | Likely Benign | -0.29 | Likely Benign | 0.325 | Likely Benign | -0.60 | Neutral | 0.001 | Benign | 0.001 | Benign | 5.33 | Benign | 0.64 | Tolerated | 3.38 | 27 | 0.1006 | 0.5242 | 2 | 1 | -0.4 | 14.03 | 251.0 | -30.1 | 0.0 | 0.0 | 0.7 | 0.1 | X | Potentially Benign | The iso-propyl side chain of Val400, located in an anti-parallel β sheet strand (res. Ala399-Ile411), hydrophobically packs against hydrophobic residues within the anti-parallel β sheet of the C2 domain (e.g., Ile268, Ala404, Leu325, Leu402). Val400 is swapped for another hydrophobic residue, leucine, whose branched hydrocarbon side chain is of a comparable size and thus packs favorably within the C2 domain. In short, the residue swap has no apparent negative effect on the structure based on the variant simulations. | 10.1016/j.ajhg.2020.11.011 | ||||||||||||
| c.1198G>T | V400L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V400L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Functional prediction tools that agree on a benign effect include REVEL, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, PROVEAN, premPS, AlphaMissense‑Default, AlphaMissense‑Optimized, and Rosetta. No tool predicts a pathogenic outcome; the only inconclusive result is from FoldX (Uncertain). High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts Benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields Likely Benign; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts Benign. Based on the collective evidence, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.398279 | Structured | 0.415488 | Uncertain | 0.951 | 0.451 | 0.000 | -1.000 | Likely Benign | 0.137 | Likely Benign | Likely Benign | -0.71 | Ambiguous | 0.2 | 0.39 | Likely Benign | -0.16 | Likely Benign | -0.29 | Likely Benign | 0.325 | Likely Benign | -0.60 | Neutral | 0.001 | Benign | 0.001 | Benign | 5.33 | Benign | 0.64 | Tolerated | 3.38 | 27 | 0.1006 | 0.5242 | 2 | 1 | -0.4 | 14.03 | 251.0 | -30.1 | 0.0 | 0.0 | 0.7 | 0.1 | X | Potentially Benign | The iso-propyl side chain of Val400, located in an anti-parallel β sheet strand (res. Ala399-Ile411), hydrophobically packs against hydrophobic residues within the anti-parallel β sheet of the C2 domain (e.g., Ile268, Ala404, Leu325, Leu402). Val400 is swapped for another hydrophobic residue, leucine, whose branched hydrocarbon side chain is of a comparable size and thus packs favorably within the C2 domain. In short, the residue swap has no apparent negative effect on the structure based on the variant simulations. | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||
| c.1219C>A | Q407K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q407K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default. Two tools, FoldX and premPS, give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, is unavailable because FoldX is uncertain and Rosetta alone is benign. Overall, the majority of evidence points to a pathogenic impact for Q407K. This conclusion is not contradicted by ClinVar status, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.109221 | Structured | 0.382522 | Uncertain | 0.916 | 0.271 | 0.000 | -14.893 | Likely Pathogenic | 0.765 | Likely Pathogenic | Likely Benign | 0.61 | Ambiguous | 0.1 | 0.19 | Likely Benign | 0.40 | Likely Benign | 0.93 | Ambiguous | 0.246 | Likely Benign | -3.42 | Deleterious | 0.863 | Possibly Damaging | 0.773 | Possibly Damaging | 3.96 | Benign | 0.07 | Tolerated | 0.1656 | 0.3504 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||
| c.1243G>A | E415K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E415K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus shows AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Based on the overall balance of predictions—particularly the two high‑accuracy pathogenic calls versus one benign—the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.100716 | Structured | 0.330366 | Uncertain | 0.915 | 0.236 | 0.000 | -11.433 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | -0.07 | Likely Benign | 0.3 | -0.13 | Likely Benign | -0.10 | Likely Benign | 0.46 | Likely Benign | 0.326 | Likely Benign | -3.69 | Deleterious | 0.998 | Probably Damaging | 0.975 | Probably Damaging | 3.22 | Benign | 0.03 | Affected | 0.2174 | 0.3860 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1255G>A | E419K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E419K missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, and FATHMM. Tools that agree on a pathogenic effect include SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Foldetta and Rosetta give uncertain results and are not counted in either group. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus predicts likely pathogenic, and Foldetta remains uncertain. Overall, the majority of evidence points to a pathogenic impact for E419K. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.371949 | Uncertain | 0.961 | 0.261 | 0.000 | -12.257 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.01 | Likely Benign | 0.1 | 1.30 | Ambiguous | 0.66 | Ambiguous | -0.03 | Likely Benign | 0.399 | Likely Benign | -3.75 | Deleterious | 0.998 | Probably Damaging | 0.975 | Probably Damaging | 3.36 | Benign | 0.07 | Tolerated | 0.2759 | 0.6899 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1264G>A | E422K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E422K missense variant is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show a split opinion: benign predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, and FATHMM, while pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus also indicates Likely Pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts a benign effect. Overall, the balance of evidence leans toward pathogenicity, and this assessment does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.426709 | Uncertain | 0.965 | 0.255 | 0.000 | -13.042 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.30 | Likely Benign | 0.0 | 0.10 | Likely Benign | 0.20 | Likely Benign | 0.32 | Likely Benign | 0.346 | Likely Benign | -3.52 | Deleterious | 0.998 | Probably Damaging | 0.975 | Probably Damaging | 3.39 | Benign | 0.04 | Affected | 0.1995 | 0.5129 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1270G>C | V424L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V424L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome, while SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports a likely benign classification. Stability‑based methods are inconclusive: FoldX is uncertain, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.050641 | Structured | 0.411431 | Uncertain | 0.973 | 0.248 | 0.000 | -4.941 | Likely Benign | 0.742 | Likely Pathogenic | Likely Benign | -0.90 | Ambiguous | 0.2 | -0.41 | Likely Benign | -0.66 | Ambiguous | 0.43 | Likely Benign | 0.159 | Likely Benign | -1.68 | Neutral | 0.327 | Benign | 0.026 | Benign | 4.50 | Benign | 0.59 | Tolerated | 0.1018 | 0.2680 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1278C>A | N426K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N426K resides in the GAP domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, and FATHMM. Those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Two tools give inconclusive results: AlphaMissense‑Optimized and premPS. High‑accuracy assessments show that AlphaMissense‑Optimized is uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign effect. Overall, the majority of predictions lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which is currently unclassified. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.042364 | Structured | 0.394941 | Uncertain | 0.959 | 0.287 | 0.000 | -11.659 | Likely Pathogenic | 0.867 | Likely Pathogenic | Ambiguous | 0.03 | Likely Benign | 0.0 | 0.00 | Likely Benign | 0.02 | Likely Benign | 0.54 | Ambiguous | 0.197 | Likely Benign | -3.86 | Deleterious | 0.998 | Probably Damaging | 0.978 | Probably Damaging | 3.38 | Benign | 0.12 | Tolerated | 0.1906 | 0.2822 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1278C>G | N426K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant N426K is located in the GAP domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, SIFT, and FATHMM, while those that predict pathogenicity are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Two tools report uncertainty: AlphaMissense‑Optimized and premPS. High‑accuracy assessments show that AlphaMissense‑Optimized is uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign effect on protein stability. Overall, the majority of predictions lean toward pathogenicity, with a split in the most reliable methods. Therefore, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.042364 | Structured | 0.394941 | Uncertain | 0.959 | 0.287 | 0.000 | -11.659 | Likely Pathogenic | 0.867 | Likely Pathogenic | Ambiguous | 0.03 | Likely Benign | 0.0 | 0.00 | Likely Benign | 0.02 | Likely Benign | 0.54 | Ambiguous | 0.197 | Likely Benign | -3.86 | Deleterious | 0.998 | Probably Damaging | 0.978 | Probably Damaging | 3.38 | Benign | 0.12 | Tolerated | 0.1906 | 0.2822 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.127G>A | G43S 2D AIThe SynGAP1 missense variant G43S is listed in ClinVar with an “Uncertain” status and is present in gnomAD (6‑33423536‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (derived from the same four high‑accuracy tools) also as benign; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.308712 | Structured | 0.431462 | Uncertain | 0.396 | 0.762 | 0.375 | Uncertain | 2 | 6-33423536-G-A | 1 | 6.20e-7 | -3.301 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.057 | Likely Benign | -0.30 | Neutral | 0.162 | Benign | 0.096 | Benign | 4.29 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2559 | 0.4064 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||
| c.1300G>C | V434L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V434L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools, Rosetta and AlphaMissense‑Default, give uncertain results. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta, a protein‑folding stability method, also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the preponderance of evidence indicates that V434L is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.158265 | Structured | 0.342846 | Uncertain | 0.954 | 0.306 | 0.000 | -8.697 | Likely Pathogenic | 0.477 | Ambiguous | Likely Benign | -0.05 | Likely Benign | 0.0 | 1.02 | Ambiguous | 0.49 | Likely Benign | 0.20 | Likely Benign | 0.225 | Likely Benign | -2.30 | Neutral | 0.947 | Possibly Damaging | 0.851 | Possibly Damaging | 3.64 | Benign | 0.27 | Tolerated | 0.0895 | 0.3813 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1306G>A | E436K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E436K is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining evaluated algorithms (REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) uniformly predict a pathogenic impact; FoldX, Foldetta, and premPS are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for E436K, which does not conflict with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.239899 | Structured | 0.321046 | Uncertain | 0.934 | 0.289 | 0.000 | Uncertain | 1 | -13.869 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.56 | Ambiguous | 0.1 | 2.86 | Destabilizing | 1.71 | Ambiguous | 0.82 | Ambiguous | 0.829 | Likely Pathogenic | -3.77 | Deleterious | 0.994 | Probably Damaging | 0.951 | Probably Damaging | 4.71 | Benign | 0.02 | Affected | 3.37 | 29 | 0.2332 | 0.5995 | 0 | 1 | -0.4 | -0.94 | 186.8 | 39.8 | 0.0 | 0.0 | -0.2 | 0.0 | X | X | X | Potentially Pathogenic | The carboxylate group of Glu436, located on the α helix (res. Met414-Glu436), forms a salt bridge with the amino group of the Lys444 side chain on an opposing α helix (res. Val441-Ser457). The backbone carbonyl of Glu436 also H-bonds with the Lys444 side chain, which helps keep the ends of the two α helices tightly connected. In contrast, in the variant simulations, the salt bridge formation with Lys444 is not possible. Instead, the repelled Lys436 side chain rotates outward, causing a change in the α helix backbone H-bonding: the amide group of Lys444 H-bonds with the carbonyl of Ala433 instead of the carbonyl of Cys432. | ||||||||||||||
| c.1320T>A | N440K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N440K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. The remaining methods—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Based on the overall distribution of predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -10.114 | Likely Pathogenic | 0.895 | Likely Pathogenic | Ambiguous | 0.92 | Ambiguous | 0.1 | 1.04 | Ambiguous | 0.98 | Ambiguous | 0.40 | Likely Benign | 0.058 | Likely Benign | -1.97 | Neutral | 0.206 | Benign | 0.021 | Benign | 3.50 | Benign | 0.19 | Tolerated | 0.1660 | 0.2550 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||
| c.1320T>G | N440K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N440K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. The remaining methods—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Based on the overall distribution of predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -10.114 | Likely Pathogenic | 0.895 | Likely Pathogenic | Ambiguous | 0.92 | Ambiguous | 0.1 | 1.04 | Ambiguous | 0.98 | Ambiguous | 0.40 | Likely Benign | 0.057 | Likely Benign | -1.97 | Neutral | 0.206 | Benign | 0.021 | Benign | 3.50 | Benign | 0.19 | Tolerated | 0.1660 | 0.2550 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||
| c.1321G>C | V441L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V441L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while Rosetta and AlphaMissense‑Default are uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts benign. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the preponderance of evidence, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | -9.546 | Likely Pathogenic | 0.395 | Ambiguous | Likely Benign | -0.43 | Likely Benign | 0.0 | 0.59 | Ambiguous | 0.08 | Likely Benign | 0.45 | Likely Benign | 0.135 | Likely Benign | -2.27 | Neutral | 0.165 | Benign | 0.028 | Benign | 3.43 | Benign | 0.11 | Tolerated | 0.0961 | 0.3881 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1327G>A | G443S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G443S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree that the substitution is benign: REVEL, FoldX, PROVEAN, polyPhen‑2 (both HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify it as benign. Only Rosetta and premPS yield uncertain results, which are treated as unavailable. Grouping by consensus, the benign‑predicting tools outnumber any pathogenic calls (none). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also reports benign. Therefore, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.250310 | Structured | 0.258623 | Uncertain | 0.935 | 0.206 | 0.000 | -1.258 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.25 | Likely Benign | 0.1 | -0.72 | Ambiguous | -0.24 | Likely Benign | -0.65 | Ambiguous | 0.087 | Likely Benign | 0.40 | Neutral | 0.000 | Benign | 0.001 | Benign | 3.51 | Benign | 0.34 | Tolerated | 0.2409 | 0.3416 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1333G>A | E445K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E445K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from FoldX, Rosetta, Foldetta, premPS, and FATHMM, whereas pathogenic calls arise from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. When high‑accuracy methods are considered separately, AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of evidence—including the high‑accuracy tools—supports a pathogenic effect. This conclusion does not conflict with ClinVar, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.270205 | Uncertain | 0.947 | 0.228 | 0.000 | -15.371 | Likely Pathogenic | 0.958 | Likely Pathogenic | Likely Pathogenic | 0.02 | Likely Benign | 0.0 | 0.02 | Likely Benign | 0.02 | Likely Benign | 0.46 | Likely Benign | 0.524 | Likely Pathogenic | -3.82 | Deleterious | 0.991 | Probably Damaging | 0.951 | Probably Damaging | 3.35 | Benign | 0.02 | Affected | 0.1784 | 0.5311 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1336G>A | E446K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E446K is not reported in ClinVar (ClinVar status: not present) and is found in gnomAD (ID 6‑33438241‑G‑A). Prediction tools that agree on a benign effect include only FATHMM. Tools that agree on a pathogenic effect comprise REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (majority vote). Predictions that are uncertain or inconclusive are FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy methods give the following: AlphaMissense‑Optimized is uncertain; SGM‑Consensus indicates likely pathogenic; Foldetta is uncertain. Overall, the majority of evidence points to a pathogenic impact. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.271506 | Structured | 0.276479 | Uncertain | 0.940 | 0.216 | 0.000 | 6-33438241-G-A | 1 | 6.19e-7 | -14.140 | Likely Pathogenic | 0.953 | Likely Pathogenic | Ambiguous | 0.80 | Ambiguous | 0.4 | 1.57 | Ambiguous | 1.19 | Ambiguous | 0.81 | Ambiguous | 0.518 | Likely Pathogenic | -3.75 | Deleterious | 0.994 | Probably Damaging | 0.975 | Probably Damaging | 3.36 | Benign | 0.01 | Affected | 3.38 | 31 | 0.2141 | 0.6511 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||
| c.1339G>C | V447L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V447L is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that classify the variant as benign include REVEL, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict pathogenicity are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Uncertain results are reported by FoldX, Rosetta, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM‑Consensus as Likely Benign, and Foldetta as Benign. Overall, the majority of evidence points to a benign effect, and this consensus does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.250310 | Structured | 0.283801 | Uncertain | 0.970 | 0.243 | 0.000 | Uncertain | 1 | -5.136 | Likely Benign | 0.491 | Ambiguous | Likely Benign | -1.13 | Ambiguous | 0.1 | 0.54 | Ambiguous | -0.30 | Likely Benign | 0.03 | Likely Benign | 0.180 | Likely Benign | -0.29 | Neutral | 0.947 | Possibly Damaging | 0.851 | Possibly Damaging | 3.61 | Benign | 0.90 | Tolerated | 3.37 | 32 | 0.0757 | 0.3477 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||
| c.1354G>C | V452L 2D 3DClick to see structure in 3D Viewer AISynGAP1 V452L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM, while those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Benign. Because the majority of tools (six) predict benign and the high‑accuracy Foldetta also supports benign, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.335645 | Structured | 0.315167 | Uncertain | 0.970 | 0.229 | 0.000 | -11.285 | Likely Pathogenic | 0.929 | Likely Pathogenic | Ambiguous | 0.29 | Likely Benign | 0.1 | -0.25 | Likely Benign | 0.02 | Likely Benign | 0.34 | Likely Benign | 0.316 | Likely Benign | -2.96 | Deleterious | 0.947 | Possibly Damaging | 0.851 | Possibly Damaging | 3.54 | Benign | 0.11 | Tolerated | 0.0777 | 0.4061 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.135C>A | N45K 2D AISynGAP1 missense variant N45K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a benign outcome; Foldetta results are unavailable. Overall, the balance of evidence—five benign versus four pathogenic predictions, with two high‑accuracy tools supporting benign—suggests that the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.401658 | Structured | 0.431853 | Uncertain | 0.498 | 0.741 | 0.375 | -1.711 | Likely Benign | 0.697 | Likely Pathogenic | Likely Benign | 0.082 | Likely Benign | -0.58 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.13 | Benign | 0.00 | Affected | 0.2409 | 0.6724 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||||
| c.135C>G | N45K 2D AISynGAP1 missense variant N45K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a benign outcome; Foldetta results are unavailable. Overall, the balance of evidence—five benign versus four pathogenic predictions, with two high‑accuracy tools supporting benign—suggests that the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.401658 | Structured | 0.431853 | Uncertain | 0.498 | 0.741 | 0.375 | -1.711 | Likely Benign | 0.697 | Likely Pathogenic | Likely Benign | 0.082 | Likely Benign | -0.58 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.13 | Benign | 0.00 | Affected | 0.2409 | 0.6724 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||||
| c.1366C>A | Q456K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q456K is not reported in ClinVar and has no gnomAD entry. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, and FATHMM, whereas those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default; premPS and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Benign. Overall, the majority of tools (six pathogenic vs five benign) and the SGM‑Consensus result point toward a pathogenic interpretation, while Foldetta suggests stability‑preserving benignity. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.170161 | Structured | 0.302348 | Uncertain | 0.939 | 0.164 | 0.000 | -13.768 | Likely Pathogenic | 0.806 | Likely Pathogenic | Ambiguous | 0.21 | Likely Benign | 0.1 | 0.23 | Likely Benign | 0.22 | Likely Benign | 0.78 | Ambiguous | 0.391 | Likely Benign | -3.75 | Deleterious | 0.969 | Probably Damaging | 0.875 | Possibly Damaging | 3.36 | Benign | 0.13 | Tolerated | 0.1321 | 0.2547 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||
| c.1375G>A | G459S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G459S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: benign calls come from REVEL and FATHMM, while pathogenic predictions are made by FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). Uncertain results are reported by Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as inconclusive, SGM‑Consensus as Likely Pathogenic, and Foldetta as inconclusive. Overall, the majority of evidence points to a pathogenic impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.185198 | Structured | 0.289888 | Uncertain | 0.903 | 0.150 | 0.125 | -10.979 | Likely Pathogenic | 0.873 | Likely Pathogenic | Ambiguous | 2.34 | Destabilizing | 0.1 | 0.77 | Ambiguous | 1.56 | Ambiguous | 0.60 | Ambiguous | 0.414 | Likely Benign | -5.46 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.09 | Benign | 0.05 | Affected | 0.2340 | 0.5197 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1414G>A | E472K 2D AIThe SynGAP1 missense variant E472K is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that assess pathogenicity unanimously classify the variant as deleterious: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect; the only inconclusive result is from premPS, which is listed as uncertain. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a destabilizing, pathogenic effect. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.359300 | Uncertain | 0.878 | 0.231 | 0.000 | -15.214 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 2.01 | Destabilizing | 1.2 | 3.23 | Destabilizing | 2.62 | Destabilizing | 0.78 | Ambiguous | 0.670 | Likely Pathogenic | -3.95 | Deleterious | 0.996 | Probably Damaging | 0.987 | Probably Damaging | 2.33 | Pathogenic | 0.03 | Affected | 0.3077 | 0.5651 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1417G>C | V473L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V473L is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, and Foldetta, while those that predict a pathogenic effect comprise SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as benign. Overall, the majority of tools (seven pathogenic vs. six benign) indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this conclusion is not contradicted by the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.362529 | Uncertain | 0.884 | 0.239 | 0.000 | -9.073 | Likely Pathogenic | 0.966 | Likely Pathogenic | Likely Pathogenic | -0.29 | Likely Benign | 0.1 | -0.52 | Ambiguous | -0.41 | Likely Benign | 0.35 | Likely Benign | 0.336 | Likely Benign | -2.85 | Deleterious | 0.929 | Possibly Damaging | 0.917 | Probably Damaging | 3.43 | Benign | 0.21 | Tolerated | 0.0708 | 0.3921 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1417G>T | V473L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V473L is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, and Foldetta, while those that predict a pathogenic effect comprise SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as benign. Overall, the majority of tools (seven pathogenic vs. six benign) indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this conclusion is not contradicted by the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.362529 | Uncertain | 0.884 | 0.239 | 0.000 | -9.073 | Likely Pathogenic | 0.966 | Likely Pathogenic | Likely Pathogenic | -0.29 | Likely Benign | 0.1 | -0.52 | Ambiguous | -0.41 | Likely Benign | 0.35 | Likely Benign | 0.335 | Likely Benign | -2.85 | Deleterious | 0.929 | Possibly Damaging | 0.917 | Probably Damaging | 3.43 | Benign | 0.21 | Tolerated | 0.0708 | 0.3921 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1432G>A | E478K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E478K is not reported in ClinVar (ClinVar ID: None) and has no entries in gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SGM‑Consensus (Likely Pathogenic), PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta as benign. Overall, the majority of evidence (eight benign versus five pathogenic predictions) points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | -12.654 | Likely Pathogenic | 0.899 | Likely Pathogenic | Ambiguous | 0.23 | Likely Benign | 0.0 | 0.12 | Likely Benign | 0.18 | Likely Benign | -0.02 | Likely Benign | 0.309 | Likely Benign | -3.45 | Deleterious | 0.320 | Benign | 0.117 | Benign | 3.49 | Benign | 0.05 | Affected | 0.2066 | 0.6192 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1438G>A | E480K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E480K is catalogued in gnomAD (ID 6‑33438470‑G‑A) but has no entry in ClinVar. Functional prediction tools cluster into two groups: benign predictions come from FoldX and SIFT, while pathogenic predictions are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain or inconclusive results are reported by Rosetta, Foldetta, and premPS. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled Likely Pathogenic, and Foldetta remains uncertain. Overall, the preponderance of evidence indicates that E480K is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.216401 | Structured | 0.426867 | Uncertain | 0.798 | 0.250 | 0.000 | 6-33438470-G-A | 1 | 6.20e-7 | -14.059 | Likely Pathogenic | 0.961 | Likely Pathogenic | Likely Pathogenic | 0.40 | Likely Benign | 0.1 | 1.08 | Ambiguous | 0.74 | Ambiguous | 0.83 | Ambiguous | 0.768 | Likely Pathogenic | -3.45 | Deleterious | 0.996 | Probably Damaging | 0.987 | Probably Damaging | -1.26 | Pathogenic | 0.11 | Tolerated | 3.37 | 34 | 0.1828 | 0.6813 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||
| c.1456G>A | E486K 2D 3DClick to see structure in 3D Viewer AISynGAP1 E486K is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, and FATHMM. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus as pathogenic, while Foldetta (combining FoldX‑MD and Rosetta outputs) indicates a benign effect. Because the predictions are split evenly and the high‑accuracy tools are contradictory, the variant’s impact remains uncertain; thus, the variant is most likely pathogenic based on the high‑accuracy predictions, a conclusion that contradicts its ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.196879 | Structured | 0.358545 | Uncertain | 0.833 | 0.245 | 0.125 | Uncertain | 2 | -14.545 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.06 | Likely Benign | 0.1 | 0.37 | Likely Benign | 0.22 | Likely Benign | 0.41 | Likely Benign | 0.435 | Likely Benign | -3.58 | Deleterious | 1.000 | Probably Damaging | 0.988 | Probably Damaging | 3.40 | Benign | 0.12 | Tolerated | 3.37 | 35 | 0.1940 | 0.6392 | 0 | 1 | -0.4 | -0.94 | 206.8 | 52.1 | -0.3 | 0.1 | 0.2 | 0.0 | X | X | Uncertain | Glu486 is located in an α-α loop connecting the two α-helices (res. Ala461-Phe476 and Leu489-Glu519) at the GAP-Ras interface. It is adjacent to the arginine finger (Arg485) and is expected to closely interact with Ras. The residue swap could affect complex formation with the GTPase and its activation. In the WT simulations, the carboxylate group of Glu486 forms salt bridges with Arg485 and Arg475 on the preceding α-helix (res. Ala461-Phe476). In the variant simulations, Lys486 does not form any specific interactions. Although the amino group of the Lys486 side chain cannot form these salt bridges, no negative effects on the protein structure are observed. Nevertheless, the potential role of Glu486 in SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations, and no definite conclusions can be drawn. | |||||||||||||||
| c.1461C>A | N487K 2D AIThe SynGAP1 missense variant N487K lies in the GAP domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM. The remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled “Likely Pathogenic.” High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. No other stability predictions are available. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.209395 | Structured | 0.338511 | Uncertain | 0.890 | 0.243 | 0.125 | -13.520 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 1.10 | Ambiguous | 0.9 | 1.28 | Ambiguous | 1.19 | Ambiguous | 0.80 | Ambiguous | 0.489 | Likely Benign | -5.97 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | 2.72 | Benign | 0.01 | Affected | 0.1953 | 0.2721 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1461C>G | N487K 2D AIThe SynGAP1 missense variant N487K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely disagree: benign predictions come from REVEL and FATHMM, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Stability‑related methods (FoldX, Rosetta, premPS, Foldetta) yield uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as unavailable. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none is available). Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.209395 | Structured | 0.338511 | Uncertain | 0.890 | 0.243 | 0.125 | -13.520 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 1.10 | Ambiguous | 0.9 | 1.28 | Ambiguous | 1.19 | Ambiguous | 0.80 | Ambiguous | 0.488 | Likely Benign | -5.97 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | 2.72 | Benign | 0.01 | Affected | 0.1953 | 0.2721 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1483G>A | E495K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E495K is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include FoldX and Foldetta, whereas the majority of tools predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools with inconclusive results (Rosetta and premPS) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus also as pathogenic, while Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) indicates a benign effect. Overall, the preponderance of evidence from standard and high‑accuracy predictors points to a pathogenic effect for E495K, which is not in conflict with the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.164327 | Structured | 0.364496 | Uncertain | 0.933 | 0.161 | 0.000 | Uncertain | 1 | -11.478 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.15 | Likely Benign | 0.2 | 0.66 | Ambiguous | 0.41 | Likely Benign | 0.70 | Ambiguous | 0.869 | Likely Pathogenic | -3.91 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | -1.29 | Pathogenic | 0.01 | Affected | 3.37 | 35 | 0.1974 | 0.5039 | 1 | 0 | -0.4 | -0.94 | |||||||||||||||||||||||||
| c.1486G>A | E496K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E496K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FoldX, whereas the majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) predict a pathogenic impact. Uncertain predictions come from Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as inconclusive. Overall, the evidence strongly favors a pathogenic classification for E496K, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.383296 | Uncertain | 0.945 | 0.179 | 0.000 | -15.795 | Likely Pathogenic | 0.961 | Likely Pathogenic | Likely Pathogenic | 0.38 | Likely Benign | 0.1 | 1.77 | Ambiguous | 1.08 | Ambiguous | 0.76 | Ambiguous | 0.743 | Likely Pathogenic | -3.58 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | -1.40 | Pathogenic | 0.04 | Affected | 0.1810 | 0.3528 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1504G>A | G502S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G502S is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include only AlphaMissense‑Optimized. All other evaluated predictors—SGM Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently classify the variant as pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts a benign outcome, whereas the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, yields an uncertain result and is treated as unavailable evidence. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.083462 | Structured | 0.340113 | Uncertain | 0.882 | 0.152 | 0.000 | -11.357 | Likely Pathogenic | 0.616 | Likely Pathogenic | Likely Benign | 2.09 | Destabilizing | 0.6 | 0.71 | Ambiguous | 1.40 | Ambiguous | 0.96 | Ambiguous | 0.839 | Likely Pathogenic | -5.74 | Deleterious | 0.975 | Probably Damaging | 0.862 | Possibly Damaging | -1.64 | Pathogenic | 0.01 | Affected | 0.2429 | 0.3217 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1507C>A | Q503K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q503K (ClinVar ID 4327028) is not reported in gnomAD. Prediction tools that classify the variant as benign include FoldX, Rosetta, Foldetta, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and FATHMM. The high‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts benign; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, predicts pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts benign. premPS is inconclusive and therefore not considered. Overall, the predictions are split, with a slight bias toward benign. Thus, the variant is most likely benign according to the computational evidence, which contradicts the ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.040537 | Structured | 0.322935 | Uncertain | 0.848 | 0.168 | 0.000 | 1 | -12.276 | Likely Pathogenic | 0.217 | Likely Benign | Likely Benign | -0.01 | Likely Benign | 0.1 | -0.26 | Likely Benign | -0.14 | Likely Benign | 0.70 | Ambiguous | 0.603 | Likely Pathogenic | -3.37 | Deleterious | 0.676 | Possibly Damaging | 0.297 | Benign | -1.42 | Pathogenic | 0.12 | Tolerated | 0.1601 | 0.2419 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||
| c.1534G>A | E512K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E512K missense variant is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, FATHMM, premPS, and Foldetta, while those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta as benign. Overall, seven tools predict pathogenicity versus six predicting benignity, with one uncertain result. Thus, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.247079 | Uncertain | 0.923 | 0.273 | 0.000 | -13.927 | Likely Pathogenic | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.17 | Likely Benign | 0.1 | 0.63 | Ambiguous | 0.40 | Likely Benign | -0.03 | Likely Benign | 0.344 | Likely Benign | -3.85 | Deleterious | 0.962 | Probably Damaging | 0.658 | Possibly Damaging | 3.32 | Benign | 0.06 | Tolerated | 0.2844 | 0.4776 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1555G>A | E519K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E519K missense variant is listed in gnomAD (ID 6‑33438798‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, and FATHMM, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). Uncertain results are reported by FoldX, Rosetta, and Foldetta. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts Pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta remains Uncertain. Overall, the preponderance of evidence points to a pathogenic effect for E519K, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.078022 | Structured | 0.104514 | Uncertain | 0.899 | 0.328 | 0.000 | 6-33438798-G-A | 1 | 6.20e-7 | -13.532 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | -0.55 | Ambiguous | 0.0 | -0.60 | Ambiguous | -0.58 | Ambiguous | 0.06 | Likely Benign | 0.328 | Likely Benign | -3.48 | Deleterious | 0.996 | Probably Damaging | 0.987 | Probably Damaging | 3.28 | Benign | 0.03 | Affected | 3.37 | 35 | 0.2545 | 0.3379 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||
| c.1561G>A | E521K 2D 3DClick to see structure in 3D Viewer AISynGAP1 E521K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments give mixed results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. Overall, the majority of conventional tools lean toward a benign interpretation, while the high‑accuracy methods are split. Thus, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.086953 | Structured | 0.062387 | Uncertain | 0.865 | 0.349 | 0.000 | -9.596 | Likely Pathogenic | 0.911 | Likely Pathogenic | Ambiguous | -0.48 | Likely Benign | 0.2 | 0.14 | Likely Benign | -0.17 | Likely Benign | -0.10 | Likely Benign | 0.379 | Likely Benign | -3.05 | Deleterious | 0.994 | Probably Damaging | 0.994 | Probably Damaging | 3.57 | Benign | 0.45 | Tolerated | 0.2895 | 0.6513 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1564G>A | E522K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E522K has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from Rosetta, Foldetta, premPS, and SIFT, while pathogenic calls are made by REVEL, PROVEAN, both polyPhen‑2 versions, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further indicate AlphaMissense‑Optimized predicts Pathogenic, SGM‑Consensus also predicts Likely Pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts Benign. No prediction is available from FoldX (Uncertain). Overall, the majority of high‑confidence tools lean toward pathogenicity, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.046216 | Uncertain | 0.823 | 0.376 | 0.000 | -12.637 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | -0.69 | Ambiguous | 0.1 | 0.14 | Likely Benign | -0.28 | Likely Benign | -0.13 | Likely Benign | 0.631 | Likely Pathogenic | -3.81 | Deleterious | 0.994 | Probably Damaging | 0.994 | Probably Damaging | -1.09 | Pathogenic | 0.23 | Tolerated | 0.2019 | 0.4291 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1569C>A | N523K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N523K is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include FoldX and Foldetta, whereas the majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) predict a pathogenic outcome. Uncertain results are reported only by Rosetta and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as benign. Taken together, the preponderance of evidence supports a pathogenic classification for this variant, and this conclusion does not contradict any existing ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.069024 | Structured | 0.033426 | Uncertain | 0.883 | 0.383 | 0.125 | -12.276 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | -0.17 | Likely Benign | 0.2 | -0.58 | Ambiguous | -0.38 | Likely Benign | 0.73 | Ambiguous | 0.607 | Likely Pathogenic | -5.35 | Deleterious | 0.972 | Probably Damaging | 0.728 | Possibly Damaging | -1.30 | Pathogenic | 0.04 | Affected | 0.1837 | 0.2866 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1569C>G | N523K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N523K is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include FoldX and Foldetta, whereas the majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) predict a pathogenic outcome. Uncertain results are reported only by Rosetta and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as benign. Taken together, the preponderance of evidence supports a pathogenic classification for this variant, and this conclusion does not contradict any existing ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.069024 | Structured | 0.033426 | Uncertain | 0.883 | 0.383 | 0.125 | -12.276 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | -0.17 | Likely Benign | 0.2 | -0.58 | Ambiguous | -0.38 | Likely Benign | 0.73 | Ambiguous | 0.607 | Likely Pathogenic | -5.35 | Deleterious | 0.972 | Probably Damaging | 0.728 | Possibly Damaging | -1.30 | Pathogenic | 0.04 | Affected | 0.1837 | 0.2866 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1573G>A | E525K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E525K is reported in gnomAD (ID 6‑33438816‑G‑A) but has no ClinVar entry. Functional prediction tools show a split assessment: benign predictions come from FoldX, Rosetta, Foldetta, and FATHMM, whereas pathogenic predictions are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). The high‑accuracy AlphaMissense‑Optimized tool classifies the variant as pathogenic, the SGM‑Consensus also indicates likely pathogenic, while the Foldetta stability analysis predicts a benign effect. No prediction or stability result is missing or inconclusive. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.206376 | Structured | 0.023618 | Uncertain | 0.937 | 0.382 | 0.125 | 6-33438816-G-A | 1 | 6.20e-7 | -15.628 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | -0.13 | Likely Benign | 0.5 | 0.34 | Likely Benign | 0.11 | Likely Benign | 0.96 | Ambiguous | 0.629 | Likely Pathogenic | -3.98 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | 2.71 | Benign | 0.00 | Affected | 3.37 | 35 | 0.2349 | 0.4293 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||
| c.1576G>C | V526L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V526L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include premPS and SIFT, whereas the majority of algorithms—SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. FoldX, Rosetta, and Foldetta provide uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments reinforce the pathogenic prediction: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates pathogenicity; Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for V526L, and this conclusion does not contradict the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.139895 | Structured | 0.023118 | Uncertain | 0.943 | 0.403 | 0.000 | -9.289 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 1.31 | Ambiguous | 0.5 | 1.74 | Ambiguous | 1.53 | Ambiguous | 0.38 | Likely Benign | 0.643 | Likely Pathogenic | -2.97 | Deleterious | 0.929 | Possibly Damaging | 0.917 | Probably Damaging | -1.16 | Pathogenic | 0.17 | Tolerated | 0.0754 | 0.3894 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1576G>T | V526L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V526L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include premPS and SIFT, whereas the majority of algorithms—SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. FoldX, Rosetta, and Foldetta provide uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments reinforce the pathogenic prediction: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates pathogenicity; Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for V526L, and this conclusion does not contradict the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.139895 | Structured | 0.023118 | Uncertain | 0.943 | 0.403 | 0.000 | -9.289 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 1.31 | Ambiguous | 0.5 | 1.74 | Ambiguous | 1.53 | Ambiguous | 0.38 | Likely Benign | 0.643 | Likely Pathogenic | -2.97 | Deleterious | 0.929 | Possibly Damaging | 0.917 | Probably Damaging | -1.16 | Pathogenic | 0.17 | Tolerated | 0.0754 | 0.3894 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1612G>A | E538K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E538K has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumVar, SIFT, and FATHMM, whereas pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The high‑accuracy assessment indicates that AlphaMissense‑Optimized is uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) supports a pathogenic outcome, and Foldetta (combining FoldX‑MD and Rosetta) predicts benign stability. Overall, the majority of tools lean toward a benign effect, but the consensus of high‑accuracy predictors and several individual pathogenic scores suggest uncertainty. The variant is most likely benign based on the bulk of evidence, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.122885 | Structured | 0.033501 | Uncertain | 0.938 | 0.359 | 0.000 | -11.345 | Likely Pathogenic | 0.922 | Likely Pathogenic | Ambiguous | -0.03 | Likely Benign | 0.0 | -0.16 | Likely Benign | -0.10 | Likely Benign | -0.22 | Likely Benign | 0.215 | Likely Benign | -2.97 | Deleterious | 0.848 | Possibly Damaging | 0.294 | Benign | 3.46 | Benign | 0.16 | Tolerated | 0.2257 | 0.3818 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1618C>A | Q540K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q540K is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default; premPS remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of predictions (10 pathogenic vs. 5 benign) indicate a likely pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.085092 | Structured | 0.029522 | Uncertain | 0.958 | 0.371 | 0.000 | -11.418 | Likely Pathogenic | 0.638 | Likely Pathogenic | Likely Benign | -0.37 | Likely Benign | 0.0 | -0.03 | Likely Benign | -0.20 | Likely Benign | 0.76 | Ambiguous | 0.808 | Likely Pathogenic | -3.98 | Deleterious | 0.985 | Probably Damaging | 0.965 | Probably Damaging | -1.31 | Pathogenic | 0.06 | Tolerated | 0.1508 | 0.2472 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||
| c.1626C>A | N542K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N542K is not reported in ClinVar and has no entries in gnomAD. Prediction tools largely disagree: benign calls come from FoldX, SIFT, and Foldetta; pathogenic calls come from SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Two tools (Rosetta and premPS) give uncertain results. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic, whereas Foldetta predicts benign. No prediction or stability result is missing or inconclusive. Overall, the majority of evidence (nine pathogenic vs three benign) points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.053060 | Structured | 0.026143 | Uncertain | 0.953 | 0.331 | 0.000 | -11.967 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | -0.36 | Likely Benign | 0.1 | 0.53 | Ambiguous | 0.09 | Likely Benign | 0.75 | Ambiguous | 0.610 | Likely Pathogenic | -5.33 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.23 | Pathogenic | 0.10 | Tolerated | 0.1870 | 0.4349 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1626C>G | N542K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N542K is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, SIFT, and Foldetta. Those that predict a pathogenic effect are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Two tools (Rosetta and premPS) returned uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also pathogenic, while Foldetta predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the majority of evidence (nine pathogenic vs. three benign) indicates that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.053060 | Structured | 0.026143 | Uncertain | 0.953 | 0.331 | 0.000 | -11.967 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | -0.36 | Likely Benign | 0.1 | 0.53 | Ambiguous | 0.09 | Likely Benign | 0.75 | Ambiguous | 0.610 | Likely Pathogenic | -5.33 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.23 | Pathogenic | 0.10 | Tolerated | 0.1870 | 0.4349 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.162C>A | N54K 2D AIThe SynGAP1 missense variant N54K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign) and is treated as unavailable. High‑accuracy methods show AlphaMissense‑Optimized as benign; Foldetta results are not provided, so its stability prediction is unavailable. Overall, the balance of evidence (five pathogenic vs. four benign predictions) suggests the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -8.252 | Likely Pathogenic | 0.779 | Likely Pathogenic | Likely Benign | 0.084 | Likely Benign | -0.82 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.23 | Benign | 0.00 | Affected | 0.2010 | 0.5987 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.162C>G | N54K 2D AIThe SynGAP1 missense variant N54K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign) and is treated as unavailable. High‑accuracy methods show AlphaMissense‑Optimized as benign; Foldetta results are not provided, so its stability prediction is unavailable. Overall, the balance of evidence (five pathogenic vs. four benign predictions) suggests the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -8.252 | Likely Pathogenic | 0.779 | Likely Pathogenic | Likely Benign | 0.084 | Likely Benign | -0.82 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.23 | Benign | 0.00 | Affected | 0.2010 | 0.5987 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.163C>A | Q55K 2D AIThe SynGAP1 missense variant Q55K is listed in ClinVar (ID 520688.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33423572‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is not in conflict with the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.332115 | Structured | 0.470108 | Uncertain | 0.461 | 0.657 | 0.000 | Uncertain | 2 | 6-33423572-C-A | 24 | 1.49e-5 | -5.840 | Likely Benign | 0.612 | Likely Pathogenic | Likely Benign | 0.085 | Likely Benign | -1.21 | Neutral | 0.140 | Benign | 0.184 | Benign | 3.91 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2047 | 0.4129 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||||||
| c.1642G>A | E548K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E548K missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM, while those that predict a pathogenic impact are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicting likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) indicating a benign effect on protein stability. Overall, the balance of evidence leans toward a pathogenic interpretation, with no conflict with ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.054297 | Structured | 0.008632 | Uncertain | 0.965 | 0.288 | 0.000 | -13.734 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | -0.34 | Likely Benign | 0.0 | -0.19 | Likely Benign | -0.27 | Likely Benign | 0.13 | Likely Benign | 0.348 | Likely Benign | -3.85 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.33 | Benign | 0.09 | Tolerated | 0.1990 | 0.4491 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1660G>C | V554L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V554L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome, while FoldX, premPS, and ESM1b are uncertain. High‑accuracy assessments reinforce the benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the variant is most likely benign based on the available computational evidence, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.020522 | Structured | 0.007349 | Uncertain | 0.955 | 0.226 | 0.000 | -7.884 | In-Between | 0.645 | Likely Pathogenic | Likely Benign | -1.14 | Ambiguous | 0.0 | 0.29 | Likely Benign | -0.43 | Likely Benign | 0.56 | Ambiguous | 0.162 | Likely Benign | -1.54 | Neutral | 0.204 | Benign | 0.053 | Benign | 3.49 | Benign | 0.08 | Tolerated | 0.0991 | 0.2998 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1660G>T | V554L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V554L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome, while FoldX, premPS, and ESM1b are uncertain. High‑accuracy assessments reinforce the benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the variant is most likely benign based on the available computational evidence, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.020522 | Structured | 0.007349 | Uncertain | 0.955 | 0.226 | 0.000 | -7.884 | In-Between | 0.645 | Likely Pathogenic | Likely Benign | -1.14 | Ambiguous | 0.0 | 0.29 | Likely Benign | -0.43 | Likely Benign | 0.56 | Ambiguous | 0.162 | Likely Benign | -1.54 | Neutral | 0.204 | Benign | 0.053 | Benign | 3.49 | Benign | 0.08 | Tolerated | 0.0991 | 0.2998 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1663G>C | V555L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V555L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The remaining tools—FoldX, Foldetta, ESM1b, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is unavailable due to a tie between pathogenic and benign calls, and Foldetta is uncertain. Overall, the preponderance of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.013265 | Structured | 0.008218 | Uncertain | 0.943 | 0.225 | 0.000 | -7.816 | In-Between | 0.521 | Ambiguous | Likely Benign | -1.43 | Ambiguous | 0.1 | -0.14 | Likely Benign | -0.79 | Ambiguous | 0.16 | Likely Benign | 0.260 | Likely Benign | -0.97 | Neutral | 0.025 | Benign | 0.015 | Benign | -1.22 | Pathogenic | 0.14 | Tolerated | 0.1036 | 0.2958 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1668C>A | N556K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N556K is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include FoldX, Rosetta, premPS, and SIFT, whereas the remaining tools (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized) predict it to be pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of predictions support a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Thus, the variant is most likely pathogenic based on the available computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.015078 | Structured | 0.008655 | Uncertain | 0.925 | 0.225 | 0.000 | -10.017 | Likely Pathogenic | 0.968 | Likely Pathogenic | Likely Pathogenic | 0.10 | Likely Benign | 0.1 | 0.14 | Likely Benign | 0.12 | Likely Benign | 0.40 | Likely Benign | 0.510 | Likely Pathogenic | -4.48 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | -1.08 | Pathogenic | 0.14 | Tolerated | 0.2024 | 0.2240 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1668C>G | N556K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N556K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from FoldX, Rosetta, premPS, and SIFT; pathogenic predictions from SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support this dichotomy: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts a benign effect. Based on the preponderance of evidence, the variant is most likely pathogenic, and this conclusion is consistent with the absence of ClinVar annotation (i.e., no conflicting benign classification). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.015078 | Structured | 0.008655 | Uncertain | 0.925 | 0.225 | 0.000 | -10.017 | Likely Pathogenic | 0.968 | Likely Pathogenic | Likely Pathogenic | 0.10 | Likely Benign | 0.1 | 0.14 | Likely Benign | 0.12 | Likely Benign | 0.40 | Likely Benign | 0.510 | Likely Pathogenic | -4.48 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | -1.08 | Pathogenic | 0.14 | Tolerated | 0.2024 | 0.2240 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
| c.1678G>C | V560L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V560L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Optimized, and Foldetta; pathogenic predictions come from polyPhen‑2 HumDiv, ESM1b, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized predicts benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, also predicts benign. Overall, the bulk of evidence points to a benign effect, but the SGM Consensus and the presence of pathogenic signals from several high‑confidence tools introduce uncertainty. Thus, the variant is most likely benign based on the prevailing predictions, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | -10.191 | Likely Pathogenic | 0.533 | Ambiguous | Likely Benign | -0.44 | Likely Benign | 0.0 | 0.48 | Likely Benign | 0.02 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -2.45 | Neutral | 0.508 | Possibly Damaging | 0.209 | Benign | -1.24 | Pathogenic | 0.40 | Tolerated | 3.37 | 35 | 0.1430 | 0.4162 | 1 | 2 | -0.4 | 14.03 | ||||||||||||||||||||||||||||
| c.1678G>T | V560L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V560L variant has no ClinVar entry (ClinVar status: None) but is catalogued in gnomAD (ID 6‑33440730‑G‑T). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are polyPhen‑2 HumDiv, ESM1b, and FATHMM (polyPhen‑2 HumVar is benign, AlphaMissense‑Default is uncertain). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning pathogenic (2 pathogenic vs 1 benign), and Foldetta indicating a benign stability change. Overall, the majority of conventional tools favor a benign classification, yet the high‑accuracy consensus and Foldetta suggest a pathogenic signal. Based on the most reliable predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | 6-33440730-G-T | 1 | 6.34e-7 | -10.191 | Likely Pathogenic | 0.533 | Ambiguous | Likely Benign | -0.44 | Likely Benign | 0.0 | 0.48 | Likely Benign | 0.02 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -2.45 | Neutral | 0.508 | Possibly Damaging | 0.209 | Benign | -1.24 | Pathogenic | 0.40 | Tolerated | 3.37 | 35 | 0.1430 | 0.4162 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||
| c.1690G>A | E564K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E564K is not reported in ClinVar and is present in the gnomAD database (variant ID 6‑33440742‑G‑A). Functional prediction tools largely agree on a deleterious effect: pathogenic calls are made by REVEL, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while only SIFT predicts a benign outcome. Uncertain results are reported by FoldX, Foldetta, and premPS. High‑accuracy assessments reinforce the pathogenic prediction: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta remains uncertain. Overall, the majority of evidence supports a pathogenic classification, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.023534 | Structured | 0.038418 | Uncertain | 0.891 | 0.208 | 0.000 | 6-33440742-G-A | -15.834 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.76 | Ambiguous | 0.1 | 2.06 | Destabilizing | 1.41 | Ambiguous | 0.89 | Ambiguous | 0.854 | Likely Pathogenic | -3.95 | Deleterious | 0.997 | Probably Damaging | 0.987 | Probably Damaging | -1.35 | Pathogenic | 0.10 | Tolerated | 3.37 | 35 | 0.1988 | 0.5280 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||||
| c.1699G>A | E567K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E567K is not reported in ClinVar (no entry) and is absent from gnomAD. Prediction tools cluster into two groups: benign calls come from REVEL, FoldX, SIFT, and FATHMM, while pathogenic calls arise from Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (likely pathogenic). High‑accuracy methods reinforce the pathogenic assessment: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, remains uncertain. PremPS is likewise inconclusive. Overall, the majority of evidence points to a pathogenic effect, and this conclusion is not contradicted by ClinVar status or gnomAD presence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.021816 | Structured | 0.051008 | Uncertain | 0.916 | 0.234 | 0.000 | -15.568 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | -0.18 | Likely Benign | 0.0 | 2.61 | Destabilizing | 1.22 | Ambiguous | 0.65 | Ambiguous | 0.380 | Likely Benign | -3.75 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.45 | Benign | 0.16 | Tolerated | 0.2115 | 0.5552 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1702G>C | V568L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V568L variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include FoldX, Foldetta, and polyPhen‑2 HumVar, while those that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (which itself is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Uncertain predictions come from Rosetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools predict a pathogenic impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.024826 | Structured | 0.053503 | Uncertain | 0.937 | 0.257 | 0.000 | -9.503 | Likely Pathogenic | 0.921 | Likely Pathogenic | Ambiguous | -0.30 | Likely Benign | 0.3 | 0.57 | Ambiguous | 0.14 | Likely Benign | 0.56 | Ambiguous | 0.651 | Likely Pathogenic | -2.69 | Deleterious | 0.511 | Possibly Damaging | 0.147 | Benign | -1.23 | Pathogenic | 0.04 | Affected | 3.37 | 35 | 0.0944 | 0.3312 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||||
| c.1702G>T | V568L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V568L is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Among the available in‑silico predictors, eight tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus) predict a pathogenic effect, whereas three tools (FoldX, Foldetta, and polyPhen‑2 HumVar) predict a benign outcome; the remaining three (Rosetta, premPS, AlphaMissense‑Optimized) are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta stability outputs) as benign. Overall, the preponderance of evidence points to a pathogenic impact, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.024826 | Structured | 0.053503 | Uncertain | 0.937 | 0.257 | 0.000 | Uncertain | 1 | -9.503 | Likely Pathogenic | 0.921 | Likely Pathogenic | Ambiguous | -0.30 | Likely Benign | 0.3 | 0.57 | Ambiguous | 0.14 | Likely Benign | 0.56 | Ambiguous | 0.651 | Likely Pathogenic | -2.69 | Deleterious | 0.511 | Possibly Damaging | 0.147 | Benign | -1.23 | Pathogenic | 0.04 | Affected | 3.37 | 35 | 0.0944 | 0.3312 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||
| c.1732G>A | E578K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E578K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, and SIFT, whereas polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default predict a pathogenic outcome. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show that the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Pathogenic, AlphaMissense‑Optimized remains Uncertain, and Foldetta predicts a benign effect. Overall, the majority of tools (seven benign vs. five pathogenic) suggest a benign impact, and this assessment does not contradict the absence of ClinVar evidence. Thus, the variant is most likely benign based on the current predictions, with no conflicting ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -13.391 | Likely Pathogenic | 0.870 | Likely Pathogenic | Ambiguous | 0.07 | Likely Benign | 0.1 | -0.19 | Likely Benign | -0.06 | Likely Benign | -0.27 | Likely Benign | 0.450 | Likely Benign | -1.65 | Neutral | 0.996 | Probably Damaging | 0.987 | Probably Damaging | -1.30 | Pathogenic | 0.49 | Tolerated | 0.1954 | 0.5480 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1738G>A | G580S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G580S is listed in ClinVar with an “Uncertain” status (ClinVar ID 1487029.0) and is present in the gnomAD database (gnomAD ID 6‑33440790‑G‑A). Among the available in‑silico predictors, the majority (REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default) indicate a pathogenic effect, whereas only SIFT predicts a benign outcome. Predictions that are inconclusive or uncertain include Rosetta, Foldetta, premPS, AlphaMissense‑Optimized, and the SGM‑Consensus (which is derived from the pathogenic majority of the four contributing tools). High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as pathogenic (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as uncertain (combining a pathogenic FoldX result with an uncertain Rosetta result). Overall, the preponderance of evidence points to a pathogenic effect, which is in contrast to the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.104810 | Structured | 0.025952 | Uncertain | 0.853 | 0.236 | 0.000 | Uncertain | 1 | 6-33440790-G-A | 1 | 6.20e-7 | -10.788 | Likely Pathogenic | 0.861 | Likely Pathogenic | Ambiguous | 2.84 | Destabilizing | 0.2 | 0.59 | Ambiguous | 1.72 | Ambiguous | 0.87 | Ambiguous | 0.644 | Likely Pathogenic | -5.73 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.23 | Pathogenic | 0.07 | Tolerated | 3.37 | 34 | 0.2509 | 0.3085 | 1 | 0 | -0.4 | 30.03 | 233.9 | -49.3 | 0.8 | 0.0 | 0.6 | 0.1 | X | Potentially Benign | Gly580 is located on the outer surface in a short α-α loop turn connecting two α-helices (res. Arg563-Glu578, res. Glu582-Phe608) in the WT simulations. In the variant simulations, the side chain of Ser580 faces outward, and its hydroxyl group does not make any new or additional interactions compared to Gly580 in the WT simulations that could affect the protein structure. | |||||||||||||
| c.1744G>A | E582K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E582K missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a benign effect on protein stability. Taken together, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is available). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -11.826 | Likely Pathogenic | 0.814 | Likely Pathogenic | Ambiguous | 0.20 | Likely Benign | 0.1 | 0.07 | Likely Benign | 0.14 | Likely Benign | 0.02 | Likely Benign | 0.168 | Likely Benign | -1.83 | Neutral | 0.994 | Probably Damaging | 0.994 | Probably Damaging | 3.31 | Benign | 0.33 | Tolerated | 0.2038 | 0.3762 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||
| c.181G>A | E61K 2D AIThe SynGAP1 E61K missense variant has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two consensus groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the aggregate evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | -4.953 | Likely Benign | 0.425 | Ambiguous | Likely Benign | 0.120 | Likely Benign | -0.22 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.34 | Benign | 0.00 | Affected | 0.2736 | 0.5691 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.1834C>A | Q612K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q612K is not reported in ClinVar (ClinVar status: not listed) and has no entry in gnomAD (gnomAD ID: none). Prediction tools that indicate a benign effect include FoldX, Rosetta, Foldetta, and SIFT, whereas pathogenic predictions come from SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default; premPS and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools predict pathogenicity, and the high‑accuracy consensus also leans pathogenic, while the folding‑stability method suggests benign. Thus, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict the ClinVar status, which currently has no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.203988 | Uncertain | 0.822 | 0.263 | 0.000 | -12.393 | Likely Pathogenic | 0.849 | Likely Pathogenic | Ambiguous | 0.15 | Likely Benign | 0.1 | 0.48 | Likely Benign | 0.32 | Likely Benign | 0.81 | Ambiguous | 0.619 | Likely Pathogenic | -3.88 | Deleterious | 0.931 | Possibly Damaging | 0.931 | Probably Damaging | -1.22 | Pathogenic | 0.19 | Tolerated | 0.1810 | 0.3641 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||
| c.1837G>A | E613K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E613K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from FoldX, Foldetta, and premPS, whereas pathogenic calls are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is also pathogenic. High‑accuracy assessments give AlphaMissense‑Optimized as pathogenic, SGM Consensus as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction is inconclusive. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.193489 | Uncertain | 0.816 | 0.254 | 0.000 | -11.892 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.34 | Likely Benign | 0.6 | -0.54 | Ambiguous | -0.10 | Likely Benign | -0.08 | Likely Benign | 0.567 | Likely Pathogenic | -3.72 | Deleterious | 0.996 | Probably Damaging | 0.987 | Probably Damaging | -1.15 | Pathogenic | 0.04 | Affected | 0.2979 | 0.5942 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1849G>A | E617K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E617K is not reported in ClinVar but is present in gnomAD (6‑33440901‑G‑A). Functional prediction tools cluster into two groups: benign predictions come from FoldX, premPS, and SIFT, while pathogenic predictions arise from SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. A third set of methods (Foldetta, AlphaMissense‑Optimized, Rosetta) yield uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic effect for E617K, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.111485 | Structured | 0.155123 | Uncertain | 0.877 | 0.240 | 0.000 | 6-33440901-G-A | 1 | 6.20e-7 | -10.702 | Likely Pathogenic | 0.910 | Likely Pathogenic | Ambiguous | 0.37 | Likely Benign | 0.1 | 1.19 | Ambiguous | 0.78 | Ambiguous | 0.17 | Likely Benign | 0.534 | Likely Pathogenic | -3.32 | Deleterious | 0.997 | Probably Damaging | 0.987 | Probably Damaging | -1.34 | Pathogenic | 0.48 | Tolerated | 3.37 | 35 | 0.1981 | 0.6282 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||
| c.1852C>A | Q618K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q618K is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33440904‑C‑A). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while ESM1b is uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign result; and Foldetta also predicts benign stability. No predictions or folding stability results are missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | 6-33440904-C-A | 24 | 1.49e-5 | -7.708 | In-Between | 0.229 | Likely Benign | Likely Benign | 0.02 | Likely Benign | 0.0 | 0.16 | Likely Benign | 0.09 | Likely Benign | -0.46 | Likely Benign | 0.281 | Likely Benign | -0.05 | Neutral | 0.338 | Benign | 0.111 | Benign | -1.21 | Pathogenic | 0.29 | Tolerated | 3.37 | 35 | 0.1413 | 0.2750 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||
| c.187G>A | E63K 2D AIThe SynGAP1 E63K missense variant (ClinVar ID 2830630.0) is listed as “Uncertain” and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default all predict a pathogenic outcome. AlphaMissense‑Optimized is inconclusive, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. Overall, the high‑accuracy consensus leans toward a benign effect, and this assessment does not contradict the ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.474807 | Uncertain | 0.494 | 0.739 | 0.125 | Uncertain | 1 | -4.976 | Likely Benign | 0.894 | Likely Pathogenic | Ambiguous | 0.103 | Likely Benign | -0.70 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 3.98 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1995 | 0.7261 | 1 | 0 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||
| c.1885G>C | V629L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V629L has no ClinVar entry and is not reported in gnomAD. Prediction tools that indicate a benign effect include REVEL, FoldX, and FATHMM, whereas the majority of algorithms predict a pathogenic outcome: Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Two tools report uncertainty: Foldetta and premPS. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely pathogenic. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus also indicates likely pathogenic, while Foldetta remains uncertain. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.040537 | Structured | 0.034796 | Uncertain | 0.970 | 0.236 | 0.000 | -12.785 | Likely Pathogenic | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.06 | Likely Benign | 0.2 | 2.26 | Destabilizing | 1.16 | Ambiguous | 0.54 | Ambiguous | 0.391 | Likely Benign | -2.79 | Deleterious | 0.975 | Probably Damaging | 0.958 | Probably Damaging | 3.22 | Benign | 0.02 | Affected | 0.0805 | 0.3336 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1891C>A | Q631K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q631K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from FoldX, Foldetta, and FATHMM, while pathogenic predictions arise from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score. Uncertain or inconclusive results are reported by Rosetta, premPS, and AlphaMissense‑Optimized. High‑accuracy methods give a mixed picture: Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a benign effect; the SGM‑Consensus, a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic outcome; AlphaMissense‑Optimized remains uncertain. Overall, the majority of evidence points toward a pathogenic impact, and this assessment does not conflict with the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.038963 | Uncertain | 0.948 | 0.230 | 0.000 | -15.194 | Likely Pathogenic | 0.953 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.1 | 1.13 | Ambiguous | 0.38 | Likely Benign | 0.88 | Ambiguous | 0.596 | Likely Pathogenic | -3.98 | Deleterious | 0.958 | Probably Damaging | 0.931 | Probably Damaging | 2.79 | Benign | 0.01 | Affected | 0.1288 | 0.2157 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||
| c.1896C>A | N632K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N632K is not reported in ClinVar and is present in gnomAD. Prediction tools that indicate a benign effect include FoldX, Rosetta, and Foldetta, whereas the majority of other in silico predictors (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) classify it as pathogenic; premPS is uncertain. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) predicts benign. Overall, the preponderance of evidence points to a pathogenic effect for N632K, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.042364 | Structured | 0.041437 | Uncertain | 0.938 | 0.254 | 0.000 | 6-33440948-C-A | -13.266 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | -0.06 | Likely Benign | 0.2 | 0.12 | Likely Benign | 0.03 | Likely Benign | 0.95 | Ambiguous | 0.766 | Likely Pathogenic | -5.14 | Deleterious | 0.983 | Probably Damaging | 0.714 | Possibly Damaging | -1.43 | Pathogenic | 0.01 | Affected | 3.37 | 34 | 0.2375 | 0.4897 | 0 | 1 | -0.4 | 14.07 | ||||||||||||||||||||||||||
| c.1896C>G | N632K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N632K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include FoldX, Rosetta, and Foldetta, whereas the majority of other in‑silico predictors (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) predict a pathogenic outcome; premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Based on the overall consensus of the majority of tools, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.042364 | Structured | 0.041437 | Uncertain | 0.938 | 0.254 | 0.000 | -13.266 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | -0.06 | Likely Benign | 0.2 | 0.12 | Likely Benign | 0.03 | Likely Benign | 0.95 | Ambiguous | 0.766 | Likely Pathogenic | -5.14 | Deleterious | 0.983 | Probably Damaging | 0.714 | Possibly Damaging | -1.43 | Pathogenic | 0.01 | Affected | 3.37 | 34 | 0.2375 | 0.4897 | 0 | 1 | -0.4 | 14.07 | |||||||||||||||||||||||||||
| c.1905C>A | N635K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant N635K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas a majority of tools predict pathogenicity: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Predictions from FoldX, Rosetta, Foldetta, and premPS are uncertain and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM Consensus as likely pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for N635K, and this conclusion does not contradict any existing ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.039760 | Structured | 0.060246 | Uncertain | 0.900 | 0.252 | 0.000 | -13.144 | Likely Pathogenic | 0.967 | Likely Pathogenic | Likely Pathogenic | 0.60 | Ambiguous | 0.1 | 0.75 | Ambiguous | 0.68 | Ambiguous | 0.85 | Ambiguous | 0.332 | Likely Benign | -5.64 | Deleterious | 0.949 | Possibly Damaging | 0.550 | Possibly Damaging | 2.92 | Benign | 0.00 | Affected | 0.2125 | 0.2510 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||
Found 8751 rows. Show 800 rows per page. Page 4/11 « Previous | Next »