
Table of SynGAP1 Isoform α2 (UniProt Q96PV0-1) Missense Variants.
| c.dna | Variant | SGM Consensus | Domain and Structure information: based on WT protein | Annotated databases | Deep learning-based pathogenicity predictions | Folding stability-based pathogenicity predictions | Sequence/structure-based pathogenicity predictions | Phase Separation | Evolutionary/physical properties | Molecular Dynamics-based analysis | DOI | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Domain | IUPred2 | ANCHOR2 | AlphaFold | MobiDB | PhosphoSitePlus | ClinVar | gnomAD | ESM1b | AlphaMissense | FoldX | Rosetta | Foldetta | PremPS | REVEL | PROVEAN | PolyPhen-2 HumDiv | PolyPhen-2 HumVar | FATHMM | SIFT | PSMutPred | PAM | Physical | SASA | Normalized B-factor backbone | Normalized B-factor sidechain | SynGAP Structural Annotation | |||||||||||||||||||||||||||||||||||||||||||||
| Score | Prediction | Score | Prediction | pLDDT | disorder | disorder | LTP | HTP | KL | PTM | Clinical Status | Review | Subm. | ID | Allele count | Allele freq. | LLR score | Prediction | Pathogenicity | Class | Optimized | Average ΔΔG | Prediction | StdDev | ΔΔG | Prediction | ΔΔG | Prediction | ΔΔG | Prediction | Score | Prediction | Score | Prediction | pph2_prob | Prediction | pph2_prob | Prediction | Nervous System Score | Prediction | Prediction | Status | Conservation | Sequences | IP RF | SP RF | Prediction | PAM250 | PAM120 | Hydropathy Δ | MW Δ | Average | Δ | Δ | StdDev | Δ | StdDev | Secondary | Tertiary bonds | Inside out | GAP-Ras interface | At membrane | No effect | MD Alert | Verdict | Description | |||||
| c.1001A>G | K334R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K334R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Foldetta. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.377384 | Structured | 0.325972 | Uncertain | 0.544 | 0.414 | 0.500 | -5.384 | Likely Benign | 0.162 | Likely Benign | Likely Benign | -0.37 | Likely Benign | 0.1 | -0.53 | Ambiguous | -0.45 | Likely Benign | 0.39 | Likely Benign | 0.247 | Likely Benign | -2.62 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.76 | Pathogenic | 0.07 | Tolerated | 0.4647 | 0.0976 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.1022G>A | G341D 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant G341D is not reported in ClinVar (ClinVar ID: None) but is present in gnomAD (ID: 6-33437927‑G‑A). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, FATHMM, and AlphaMissense‑Default. The remaining tools—Foldetta, AlphaMissense‑Optimized, ESM1b, and Rosetta—return uncertain or inconclusive results and are treated as unavailable for pathogenicity inference. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Considering the majority of standard tools lean benign but the high‑accuracy consensus indicates pathogenicity, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.356642 | Structured | 0.431164 | Uncertain | 0.745 | 0.479 | 0.250 | 6-33437927-G-A | 6 | 3.72e-6 | -7.402 | In-Between | 0.871 | Likely Pathogenic | Ambiguous | 0.28 | Likely Benign | 0.1 | -1.32 | Ambiguous | -0.52 | Ambiguous | -0.04 | Likely Benign | 0.295 | Likely Benign | -0.11 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 0.34 | Pathogenic | 0.25 | Tolerated | 3.42 | 13 | 0.1727 | 0.2241 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||
| c.1025A>T | Y342F 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y342F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive and therefore treated as unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.366687 | Structured | 0.408200 | Uncertain | 0.866 | 0.487 | 0.250 | -6.987 | Likely Benign | 0.145 | Likely Benign | Likely Benign | -0.13 | Likely Benign | 0.1 | 0.42 | Likely Benign | 0.15 | Likely Benign | 0.05 | Likely Benign | 0.160 | Likely Benign | -2.67 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 2.03 | Pathogenic | 0.26 | Tolerated | 0.2626 | 0.3615 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||
| c.1028T>G | V343G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V343G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated algorithms—FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—classify the variant as pathogenic. AlphaMissense‑Optimized is uncertain, providing no definitive direction. High‑accuracy assessments further support pathogenicity: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Thus, the preponderance of evidence indicates that V343G is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.291804 | Structured | 0.383911 | Uncertain | 0.882 | 0.497 | 0.250 | -11.332 | Likely Pathogenic | 0.926 | Likely Pathogenic | Ambiguous | 2.49 | Destabilizing | 0.1 | 4.40 | Destabilizing | 3.45 | Destabilizing | 1.68 | Destabilizing | 0.421 | Likely Benign | -5.84 | Deleterious | 0.898 | Possibly Damaging | 0.996 | Probably Damaging | 1.60 | Pathogenic | 0.00 | Affected | 0.1982 | 0.3341 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1034T>A | L345Q 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant L345Q is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into benign (REVEL, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) and pathogenic (premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM). FoldX, Rosetta, and Foldetta give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is unavailable. Overall, the majority of predictions lean toward pathogenicity, while the single high‑accuracy tool that is definitive is benign. Thus, the variant is most likely pathogenic based on the collective evidence, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.260850 | Structured | 0.354989 | Uncertain | 0.936 | 0.478 | 0.125 | -6.034 | Likely Benign | 0.213 | Likely Benign | Likely Benign | 1.32 | Ambiguous | 0.1 | 0.86 | Ambiguous | 1.09 | Ambiguous | 1.26 | Destabilizing | 0.218 | Likely Benign | -3.67 | Deleterious | 0.993 | Probably Damaging | 0.844 | Possibly Damaging | 1.82 | Pathogenic | 0.03 | Affected | 0.0937 | 0.0842 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||
| c.1037T>G | V346G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V346G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). All available in‑silico predictors classify it as pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a pathogenic verdict; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports pathogenic. With all evidence converging on a deleterious impact and no ClinVar annotation to contradict, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.260850 | Structured | 0.350921 | Uncertain | 0.949 | 0.461 | 0.000 | -12.779 | Likely Pathogenic | 0.969 | Likely Pathogenic | Likely Pathogenic | 4.24 | Destabilizing | 0.2 | 4.62 | Destabilizing | 4.43 | Destabilizing | 2.15 | Destabilizing | 0.667 | Likely Pathogenic | -6.03 | Deleterious | 0.991 | Probably Damaging | 0.999 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.2378 | 0.2522 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1039A>C | T347P 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 T347P variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and FATHMM. Three tools (FoldX, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.268042 | Structured | 0.349915 | Uncertain | 0.951 | 0.434 | 0.000 | -9.323 | Likely Pathogenic | 0.417 | Ambiguous | Likely Benign | 0.55 | Ambiguous | 0.0 | 0.28 | Likely Benign | 0.42 | Likely Benign | 0.65 | Ambiguous | 0.236 | Likely Benign | -2.41 | Neutral | 0.627 | Possibly Damaging | 0.139 | Benign | 1.63 | Pathogenic | 0.12 | Tolerated | 0.2000 | 0.5350 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.1042G>A | V348M 2D  3DClick to see structure in 3D Viewer AISynGAP1 variant V348M is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that report a clear outcome fall into two groups: benign calls come from REVEL, Foldetta, PROVEAN, and AlphaMissense‑Optimized; pathogenic calls come from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Default, ESM1b) give uncertain results, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is unavailable due to no majority. High‑accuracy methods specifically show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus is not available. With four benign and four pathogenic predictions, the evidence is evenly split, providing no definitive direction. Therefore, the variant is not clearly benign or pathogenic based on current predictions, and this lack of consensus does not contradict its ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.170161 | Structured | 0.346556 | Uncertain | 0.951 | 0.414 | 0.000 | Uncertain | 1 | -7.076 | In-Between | 0.546 | Ambiguous | Likely Benign | -1.19 | Ambiguous | 0.1 | 0.72 | Ambiguous | -0.24 | Likely Benign | 0.76 | Ambiguous | 0.191 | Likely Benign | -1.62 | Neutral | 0.966 | Probably Damaging | 0.564 | Possibly Damaging | 1.58 | Pathogenic | 0.03 | Affected | 3.37 | 25 | 0.0903 | 0.4748 | 2 | 1 | -2.3 | 32.06 | 253.8 | -47.4 | -0.3 | 0.1 | 0.2 | 0.1 | X | Potentially Benign | The iso-propyl side chain of Val348, located in an anti-parallel β sheet strand (res. Gly341-Pro349), packs against multiple hydrophobic C2 domain residues (e.g., Leu353, Leu323, Leu402). In the variant simulations, the thioether side chain of Met348 can form similar interactions as valine due to its comparable hydrophobic profile. In fact, the thioether group of methionine can even stack favorably with the phenol ring of Tyr363 in the anti-parallel β sheet strand (res. Ala399-Ile411). Overall, the residue swap does not appear to cause negative effects on the protein structure based on the simulations. | |||||||||||||||||
| c.1043T>G | V348G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V348G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are limited to REVEL, which scores the variant as benign. All other evaluated algorithms—FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. High‑accuracy methods reinforce this assessment: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) indicates a pathogenic effect. No prediction or stability result is missing or inconclusive. Based on the overwhelming agreement among pathogenic predictions and the lack of a benign consensus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.170161 | Structured | 0.346556 | Uncertain | 0.951 | 0.414 | 0.000 | -12.793 | Likely Pathogenic | 0.964 | Likely Pathogenic | Likely Pathogenic | 3.97 | Destabilizing | 0.2 | 5.70 | Destabilizing | 4.84 | Destabilizing | 2.23 | Destabilizing | 0.419 | Likely Benign | -6.18 | Deleterious | 0.637 | Possibly Damaging | 0.989 | Probably Damaging | 1.56 | Pathogenic | 0.00 | Affected | 0.2229 | 0.2082 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1045C>T | P349S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 P349S missense variant is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Predictions that are inconclusive or uncertain are FoldX, ESM1b, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of tools, including the high‑accuracy methods, predict a pathogenic effect. Thus, the variant is most likely pathogenic, which does not contradict its current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.167087 | Structured | 0.348607 | Uncertain | 0.947 | 0.396 | 0.000 | Uncertain | 1 | -7.654 | In-Between | 0.217 | Likely Benign | Likely Benign | 1.92 | Ambiguous | 0.1 | 2.28 | Destabilizing | 2.10 | Destabilizing | 0.87 | Ambiguous | 0.277 | Likely Benign | -6.13 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 1.66 | Pathogenic | 0.06 | Tolerated | 3.37 | 25 | 0.3771 | 0.5756 | 1 | -1 | 0.8 | -10.04 | 194.9 | -18.1 | -0.1 | 0.0 | 0.2 | 0.1 | X | X | Potentially Pathogenic | The cyclic pyrrolidine side chain of Pro349, located at the end of an anti-parallel β sheet strand (res. Gly341-Pro349), allows the strand to end and make a tight turn before a short α helical section within a loop connecting to another β strand (res. Thr359-Pro364). In the variant simulations, the hydroxyl group of Ser349 forms a hydrogen bond with the backbone amide group of Ala351 in the short helical section. Conversely, the backbone amide group of Ser349 (absent in proline) does not form any intra-protein hydrogen bonds. However, the β strand end connects to the α helical section in a more stable and consistent manner compared to the WT. Although the residue swap does not cause major adverse effects on the protein structure in the simulations, it is possible that the tight turn at the β strand end could not be created during folding without the presence of proline. | ||||||||||||||||
| c.1048G>A | V350M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V350M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. The remaining tools (FoldX, premPS, ESM1b) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as benign. Taken together, the majority of evidence supports a benign classification. There is no ClinVar entry to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.179055 | Structured | 0.353227 | Uncertain | 0.931 | 0.371 | 0.000 | -7.493 | In-Between | 0.273 | Likely Benign | Likely Benign | -0.65 | Ambiguous | 0.1 | 0.40 | Likely Benign | -0.13 | Likely Benign | 0.69 | Ambiguous | 0.075 | Likely Benign | -1.95 | Neutral | 0.868 | Possibly Damaging | 0.383 | Benign | 1.63 | Pathogenic | 0.06 | Tolerated | 0.1005 | 0.4359 | 2 | 1 | -2.3 | 32.06 | ||||||||||||||||||||||||||||||
| c.1049T>G | V350G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V350G lies in the C2 domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that classify it as benign include REVEL, polyPhen‑2 HumDiv, and AlphaMissense‑Optimized. The remaining tools—FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—predict it to be pathogenic. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. All predictions are available and consistent. Based on the preponderance of pathogenic calls, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.179055 | Structured | 0.353227 | Uncertain | 0.931 | 0.371 | 0.000 | -12.267 | Likely Pathogenic | 0.597 | Likely Pathogenic | Likely Benign | 2.57 | Destabilizing | 0.0 | 4.25 | Destabilizing | 3.41 | Destabilizing | 1.93 | Destabilizing | 0.330 | Likely Benign | -5.46 | Deleterious | 0.435 | Benign | 0.920 | Probably Damaging | 1.61 | Pathogenic | 0.02 | Affected | 0.2316 | 0.2522 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.104T>G | V35G 2D AIThe SynGAP1 missense variant V35G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is benign. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; the SGM Consensus also predicts benign; Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status because the variant is not currently catalogued in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.613573 | Disordered | 0.434838 | Uncertain | 0.360 | 0.851 | 0.375 | -3.614 | Likely Benign | 0.109 | Likely Benign | Likely Benign | 0.143 | Likely Benign | 0.66 | Neutral | 0.693 | Possibly Damaging | 0.824 | Possibly Damaging | 4.41 | Benign | 0.00 | Affected | 0.1514 | 0.2618 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.1051G>C | A351P 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A351P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are Foldetta, PROVEAN, SIFT, FATHMM, and Rosetta. FoldX and premPS give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of predictions lean toward a benign impact, and this is consistent with the lack of ClinVar reporting; thus the variant is most likely benign rather than pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.216401 | Structured | 0.362025 | Uncertain | 0.925 | 0.342 | 0.000 | -6.559 | Likely Benign | 0.147 | Likely Benign | Likely Benign | -0.89 | Ambiguous | 0.2 | 6.19 | Destabilizing | 2.65 | Destabilizing | 0.62 | Ambiguous | 0.051 | Likely Benign | -3.08 | Deleterious | 0.016 | Benign | 0.007 | Benign | 1.67 | Pathogenic | 0.03 | Affected | 0.1862 | 0.5472 | 1 | -1 | -3.4 | 26.04 | ||||||||||||||||||||||||||||||
| c.1052C>A | A351D 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A351D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, AlphaMissense‑Optimized, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and FATHMM; AlphaMissense‑Default and ESM1b are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward pathogenic due to two pathogenic and two uncertain calls. Overall, the majority of tools predict a benign outcome, and this does not contradict the lack of ClinVar annotation. Thus, based on the available predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.216401 | Structured | 0.362025 | Uncertain | 0.925 | 0.342 | 0.000 | -7.917 | In-Between | 0.543 | Ambiguous | Likely Benign | -0.02 | Likely Benign | 0.0 | -0.12 | Likely Benign | -0.07 | Likely Benign | 0.49 | Likely Benign | 0.114 | Likely Benign | -3.61 | Deleterious | 0.842 | Possibly Damaging | 0.321 | Benign | 1.77 | Pathogenic | 0.11 | Tolerated | 0.1535 | 0.1386 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1052C>G | A351G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A351G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, FoldX, polyPhen‑2 HumVar, AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. Predictions that are uncertain or inconclusive are Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs two pathogenic), and Foldetta is uncertain. Taken together, the majority of available predictions lean toward a benign impact, and this assessment does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.216401 | Structured | 0.362025 | Uncertain | 0.925 | 0.342 | 0.000 | -4.983 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.39 | Likely Benign | 0.0 | 1.04 | Ambiguous | 0.72 | Ambiguous | 0.55 | Ambiguous | 0.041 | Likely Benign | -2.53 | Deleterious | 0.688 | Possibly Damaging | 0.138 | Benign | 1.77 | Pathogenic | 0.04 | Affected | 0.2298 | 0.4737 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||
| c.1061C>A | A354D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A354D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. Those that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. Four tools (Foldetta, premPS, ESM1b, and Rosetta) return uncertain results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors a pathogenic interpretation. Foldetta’s stability prediction is uncertain. Overall, the majority of consensus tools lean toward a benign effect, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.381329 | Uncertain | 0.882 | 0.335 | 0.125 | -7.914 | In-Between | 0.725 | Likely Pathogenic | Likely Benign | 0.22 | Likely Benign | 0.1 | 1.10 | Ambiguous | 0.66 | Ambiguous | 0.57 | Ambiguous | 0.087 | Likely Benign | -1.49 | Neutral | 0.255 | Benign | 0.053 | Benign | 1.75 | Pathogenic | 0.07 | Tolerated | 0.1759 | 0.1345 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1064G>C | G355A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 G355A missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. FoldX and Foldetta are uncertain and therefore not counted as evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta is uncertain. Consequently, the overall prediction leans toward a benign effect, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.155435 | Structured | 0.388832 | Uncertain | 0.810 | 0.354 | 0.125 | -5.431 | Likely Benign | 0.245 | Likely Benign | Likely Benign | 0.88 | Ambiguous | 0.6 | 0.46 | Likely Benign | 0.67 | Ambiguous | 0.49 | Likely Benign | 0.286 | Likely Benign | -4.80 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.80 | Pathogenic | 0.03 | Affected | 0.4111 | 0.5045 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||
| c.1069C>G | H357D 2D 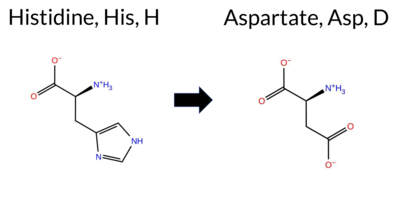 3DClick to see structure in 3D Viewer AIThe SynGAP1 H357D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta also predicts a benign outcome. No prediction or folding‑stability result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -12.013 | Likely Pathogenic | 0.559 | Ambiguous | Likely Benign | -0.23 | Likely Benign | 0.4 | -0.27 | Likely Benign | -0.25 | Likely Benign | 0.48 | Likely Benign | 0.208 | Likely Benign | -1.90 | Neutral | 0.495 | Possibly Damaging | 0.169 | Benign | 4.22 | Benign | 0.08 | Tolerated | 0.2782 | 0.1976 | 1 | -1 | -0.3 | -22.05 | ||||||||||||||||||||||||||||||
| c.1070A>C | H357P 2D 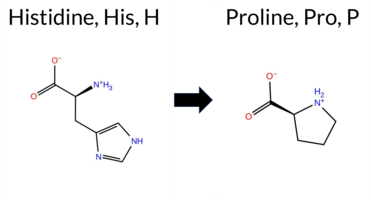 3DClick to see structure in 3D Viewer AIThe SynGAP1 H357P missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are Rosetta, PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Two tools give uncertain results: Foldetta (protein‑folding stability) and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign effect. Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -7.953 | In-Between | 0.267 | Likely Benign | Likely Benign | 0.17 | Likely Benign | 0.9 | 3.52 | Destabilizing | 1.85 | Ambiguous | -0.06 | Likely Benign | 0.197 | Likely Benign | -2.70 | Deleterious | 0.936 | Possibly Damaging | 0.469 | Possibly Damaging | 4.18 | Benign | 0.17 | Tolerated | 0.2433 | 0.4468 | 0 | -2 | 1.6 | -40.02 | ||||||||||||||||||||||||||||||
| c.1070A>G | H357R 2D 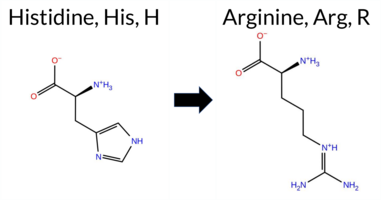 3DClick to see structure in 3D Viewer AIThe SynGAP1 H357R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. Two tools remain uncertain: Rosetta and AlphaMissense‑Default. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority; and Foldetta also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -9.212 | Likely Pathogenic | 0.348 | Ambiguous | Likely Benign | -0.10 | Likely Benign | 0.3 | 1.07 | Ambiguous | 0.49 | Likely Benign | 0.32 | Likely Benign | 0.107 | Likely Benign | -1.03 | Neutral | 0.495 | Possibly Damaging | 0.095 | Benign | 4.22 | Benign | 0.59 | Tolerated | 0.2147 | 0.2818 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||
| c.1070A>T | H357L 2D 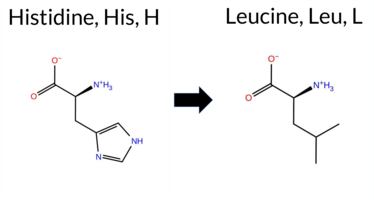 3DClick to see structure in 3D Viewer AIThe SynGAP1 H357L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and polyPhen‑2 HumDiv, while polyPhen‑2 HumVar and ESM1b are benign or uncertain, respectively. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign (3 benign vs 1 pathogenic), and Foldetta also predicts benign. No predictions are missing or inconclusive. Overall, the variant is most likely benign based on the majority of computational evidence, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -7.281 | In-Between | 0.140 | Likely Benign | Likely Benign | -0.18 | Likely Benign | 0.1 | 0.14 | Likely Benign | -0.02 | Likely Benign | 0.10 | Likely Benign | 0.203 | Likely Benign | -3.39 | Deleterious | 0.704 | Possibly Damaging | 0.169 | Benign | 4.20 | Benign | 0.25 | Tolerated | 0.1084 | 0.5971 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||
| c.1072T>C | F358L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 F358L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain or inconclusive results are reported for Rosetta, Foldetta, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic, while Foldetta’s stability prediction is unavailable due to inconclusiveness. Overall, the majority of available evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.222385 | Structured | 0.407113 | Uncertain | 0.912 | 0.441 | 0.250 | -7.865 | In-Between | 0.964 | Likely Pathogenic | Likely Pathogenic | 0.18 | Likely Benign | 0.1 | 1.62 | Ambiguous | 0.90 | Ambiguous | 0.97 | Ambiguous | 0.290 | Likely Benign | -4.21 | Deleterious | 0.982 | Probably Damaging | 0.952 | Probably Damaging | 4.12 | Benign | 0.22 | Tolerated | 0.2555 | 0.3602 | 2 | 0 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||
| c.1073T>G | F358C 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F358C is not reported in ClinVar and is absent from gnomAD. Consensus from standard in‑silico predictors shows a split: benign calls come from REVEL, SIFT, and FATHMM, whereas pathogenic calls arise from Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default. High‑accuracy assessments are less definitive: AlphaMissense‑Optimized is inconclusive, Foldetta is inconclusive, and the SGM Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward pathogenic. Because the majority of available predictions favor a damaging effect, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.222385 | Structured | 0.407113 | Uncertain | 0.912 | 0.441 | 0.250 | -7.966 | In-Between | 0.927 | Likely Pathogenic | Ambiguous | 1.68 | Ambiguous | 0.1 | 2.19 | Destabilizing | 1.94 | Ambiguous | 1.18 | Destabilizing | 0.460 | Likely Benign | -6.36 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 4.02 | Benign | 0.06 | Tolerated | 0.2364 | 0.1800 | -4 | -2 | -0.3 | -44.04 | ||||||||||||||||||||||||||||||
| c.1074C>A | F358L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 F358L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain or inconclusive results are reported for Rosetta, Foldetta, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic, while Foldetta’s stability prediction is unavailable due to inconclusiveness. Overall, the majority of available evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.222385 | Structured | 0.407113 | Uncertain | 0.912 | 0.441 | 0.250 | -7.865 | In-Between | 0.964 | Likely Pathogenic | Likely Pathogenic | 0.18 | Likely Benign | 0.1 | 1.62 | Ambiguous | 0.90 | Ambiguous | 0.97 | Ambiguous | 0.215 | Likely Benign | -4.21 | Deleterious | 0.982 | Probably Damaging | 0.952 | Probably Damaging | 4.12 | Benign | 0.22 | Tolerated | 0.2555 | 0.3602 | 2 | 0 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||
| c.1074C>G | F358L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 F358L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain or inconclusive results come from Rosetta, Foldetta, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic, while Foldetta’s stability prediction is unavailable. Overall, the majority of reliable tools predict a pathogenic impact for F358L. This conclusion does not contradict ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.222385 | Structured | 0.407113 | Uncertain | 0.912 | 0.441 | 0.250 | -7.865 | In-Between | 0.964 | Likely Pathogenic | Likely Pathogenic | 0.18 | Likely Benign | 0.1 | 1.62 | Ambiguous | 0.90 | Ambiguous | 0.97 | Ambiguous | 0.215 | Likely Benign | -4.21 | Deleterious | 0.982 | Probably Damaging | 0.952 | Probably Damaging | 4.12 | Benign | 0.22 | Tolerated | 0.2555 | 0.3602 | 2 | 0 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||
| c.1076C>A | T359K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 T359K missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. The remaining tools—FoldX, Rosetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions (seven benign vs. two pathogenic) support a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.281712 | Structured | 0.414952 | Uncertain | 0.939 | 0.480 | 0.250 | -10.822 | Likely Pathogenic | 0.517 | Ambiguous | Likely Benign | -0.65 | Ambiguous | 0.1 | 0.66 | Ambiguous | 0.01 | Likely Benign | 0.75 | Ambiguous | 0.163 | Likely Benign | -2.23 | Neutral | 0.255 | Benign | 0.045 | Benign | 1.79 | Pathogenic | 0.24 | Tolerated | 0.1618 | 0.3483 | 0 | -1 | -3.2 | 27.07 | ||||||||||||||||||||||||||||||
| c.1081C>G | Q361E 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q361E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Uncertain predictions come from Rosetta, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, while Foldetta remains uncertain. Taken together, the majority of evidence supports a benign impact for Q361E, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.305330 | Structured | 0.427593 | Uncertain | 0.945 | 0.534 | 0.250 | -7.544 | In-Between | 0.289 | Likely Benign | Likely Benign | 0.29 | Likely Benign | 0.0 | 1.02 | Ambiguous | 0.66 | Ambiguous | 0.21 | Likely Benign | 0.153 | Likely Benign | -0.89 | Neutral | 0.969 | Probably Damaging | 0.930 | Probably Damaging | 1.70 | Pathogenic | 0.26 | Tolerated | 0.1217 | 0.3018 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1083G>C | Q361H 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q361H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The remaining tools—FoldX, Rosetta, ESM1b, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also unavailable. Overall, the majority of conventional predictors lean toward pathogenicity, while the single high‑accuracy tool suggests benign. No ClinVar annotation exists to contradict these findings. Thus, based on the collective evidence, the variant is most likely pathogenic, with no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.305330 | Structured | 0.427593 | Uncertain | 0.945 | 0.534 | 0.250 | -7.269 | In-Between | 0.410 | Ambiguous | Likely Benign | 0.68 | Ambiguous | 0.1 | 0.52 | Ambiguous | 0.60 | Ambiguous | 0.32 | Likely Benign | 0.281 | Likely Benign | -2.73 | Deleterious | 0.996 | Probably Damaging | 0.986 | Probably Damaging | 1.79 | Pathogenic | 0.04 | Affected | 0.1352 | 0.4151 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1083G>T | Q361H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q361H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The remaining tools—FoldX, Rosetta, ESM1b, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also unavailable. Overall, the majority of conventional predictors (five pathogenic vs. three benign) lean toward a pathogenic interpretation, while the single high‑accuracy tool suggests benign. No ClinVar entry exists, so there is no contradiction with clinical database status. Based on the collective evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.305330 | Structured | 0.427593 | Uncertain | 0.945 | 0.534 | 0.250 | -7.269 | In-Between | 0.410 | Ambiguous | Likely Benign | 0.68 | Ambiguous | 0.1 | 0.52 | Ambiguous | 0.60 | Ambiguous | 0.32 | Likely Benign | 0.281 | Likely Benign | -2.73 | Deleterious | 0.996 | Probably Damaging | 0.986 | Probably Damaging | 1.79 | Pathogenic | 0.04 | Affected | 0.1352 | 0.4151 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1088A>T | Y363F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y363F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, seven tools predict benign while four predict pathogenic, with no evidence of pathogenicity from the most accurate methods. Therefore, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.321458 | Structured | 0.435392 | Uncertain | 0.954 | 0.586 | 0.125 | -6.621 | Likely Benign | 0.114 | Likely Benign | Likely Benign | -0.23 | Likely Benign | 0.0 | 0.42 | Likely Benign | 0.10 | Likely Benign | 0.58 | Ambiguous | 0.293 | Likely Benign | -3.42 | Deleterious | 0.997 | Probably Damaging | 0.970 | Probably Damaging | 1.74 | Pathogenic | 0.18 | Tolerated | 0.2781 | 0.3583 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||
| c.1090C>A | P364T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P364T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Four tools (FoldX, Foldetta, premPS, ESM1b) return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evaluated tools (five benign versus four pathogenic) lean toward a benign classification, and this assessment does not contradict any ClinVar annotation because no ClinVar claim exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.390993 | Structured | 0.439474 | Uncertain | 0.942 | 0.590 | 0.250 | -7.951 | In-Between | 0.230 | Likely Benign | Likely Benign | 1.18 | Ambiguous | 0.6 | 0.47 | Likely Benign | 0.83 | Ambiguous | 0.53 | Ambiguous | 0.342 | Likely Benign | -5.60 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 1.60 | Pathogenic | 0.09 | Tolerated | 0.1660 | 0.5726 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1094T>G | V365G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V365G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that assess pathogenicity unanimously classify the variant as pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. No tool predicts a benign effect. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. All available evidence points to a pathogenic effect. Therefore, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.414856 | Structured | 0.441505 | Uncertain | 0.923 | 0.608 | 0.250 | -13.020 | Likely Pathogenic | 0.944 | Likely Pathogenic | Ambiguous | 4.18 | Destabilizing | 0.2 | 4.83 | Destabilizing | 4.51 | Destabilizing | 2.18 | Destabilizing | 0.576 | Likely Pathogenic | -5.88 | Deleterious | 0.688 | Possibly Damaging | 0.989 | Probably Damaging | 1.66 | Pathogenic | 0.00 | Affected | 0.1835 | 0.2717 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1100T>G | L367R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 L367R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Those that predict a pathogenic effect are SIFT, FATHMM, and Rosetta. Tools with uncertain or inconclusive results are FoldX, premPS, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of evidence points to a benign impact, with a minority of pathogenic predictions. The variant’s status is not contradicted by ClinVar, as it is not yet classified there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.370445 | Structured | 0.441805 | Uncertain | 0.790 | 0.657 | 0.250 | -6.527 | Likely Benign | 0.515 | Ambiguous | Likely Benign | 0.68 | Ambiguous | 0.4 | 6.63 | Destabilizing | 3.66 | Destabilizing | 0.70 | Ambiguous | 0.196 | Likely Benign | -0.39 | Neutral | 0.146 | Benign | 0.057 | Benign | 1.66 | Pathogenic | 0.02 | Affected | 0.1608 | 0.1173 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||
| c.1102C>A | P368T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P368T missense variant is not listed in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. FoldX, Rosetta, and Foldetta report uncertain or inconclusive stability changes and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie (2 benign, 2 pathogenic) and thus inconclusive, and Foldetta remains uncertain. Overall, the predictions are evenly split between benign and pathogenic, providing no definitive classification. The variant’s status does not contradict ClinVar, which has no entry for it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -5.308 | Likely Benign | 0.284 | Likely Benign | Likely Benign | 1.95 | Ambiguous | 0.6 | 1.61 | Ambiguous | 1.78 | Ambiguous | 0.45 | Likely Benign | 0.188 | Likely Benign | -5.43 | Deleterious | 0.941 | Possibly Damaging | 0.527 | Possibly Damaging | 1.72 | Pathogenic | 0.01 | Affected | 0.1983 | 0.6155 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1102C>G | P368A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 P368A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, AlphaMissense‑Default, AlphaMissense‑Optimized, ESM1b, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta (combining FoldX‑MD and Rosetta outputs) is also inconclusive. Overall, the balance of evidence leans toward a benign impact for P368A. This conclusion does not contradict any ClinVar annotation, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -4.608 | Likely Benign | 0.174 | Likely Benign | Likely Benign | 1.49 | Ambiguous | 0.3 | 1.47 | Ambiguous | 1.48 | Ambiguous | 0.47 | Likely Benign | 0.144 | Likely Benign | -5.42 | Deleterious | 0.767 | Possibly Damaging | 0.344 | Benign | 1.74 | Pathogenic | 0.02 | Affected | 0.3861 | 0.5635 | 1 | -1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||
| c.1102C>T | P368S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P368S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and FATHMM. The remaining methods (FoldX, Rosetta, Foldetta, premPS) yield uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is uncertain, so these do not alter the overall interpretation. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -4.790 | Likely Benign | 0.247 | Likely Benign | Likely Benign | 1.68 | Ambiguous | 0.4 | 1.60 | Ambiguous | 1.64 | Ambiguous | 0.52 | Ambiguous | 0.090 | Likely Benign | -5.12 | Deleterious | 0.384 | Benign | 0.113 | Benign | 1.80 | Pathogenic | 0.10 | Tolerated | 0.3700 | 0.5635 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||
| c.1103C>A | P368Q 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 P368Q missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Remaining tools (AlphaMissense‑Default, FoldX, Rosetta, Foldetta, premPS) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of definitive predictions (five pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -6.019 | Likely Benign | 0.413 | Ambiguous | Likely Benign | 1.56 | Ambiguous | 0.8 | 1.71 | Ambiguous | 1.64 | Ambiguous | 0.71 | Ambiguous | 0.205 | Likely Benign | -5.10 | Deleterious | 0.991 | Probably Damaging | 0.881 | Possibly Damaging | 1.71 | Pathogenic | 0.04 | Affected | 0.1694 | 0.5283 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||
| c.1103C>T | P368L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P368L is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438008‑C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Predictions that are uncertain or inconclusive are FoldX, Rosetta, premPS, AlphaMissense‑Default, and Foldetta. High‑accuracy assessments give AlphaMissense‑Optimized a benign score, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. Based on the overall distribution of predictions, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | 6-33438008-C-T | 1 | 6.33e-7 | -6.520 | Likely Benign | 0.444 | Ambiguous | Likely Benign | 1.52 | Ambiguous | 0.7 | 1.15 | Ambiguous | 1.34 | Ambiguous | 0.52 | Ambiguous | 0.248 | Likely Benign | -6.61 | Deleterious | 0.991 | Probably Damaging | 0.831 | Possibly Damaging | 1.77 | Pathogenic | 0.00 | Affected | 3.42 | 19 | 0.2336 | 0.7125 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||
| c.1106C>A | T369K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T369K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. Foldetta and Rosetta give uncertain results, which are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs two pathogenic). Foldetta’s stability prediction is uncertain. Overall, the majority of evidence (nine benign vs two pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | -5.884 | Likely Benign | 0.616 | Likely Pathogenic | Likely Benign | -0.10 | Likely Benign | 0.1 | 1.13 | Ambiguous | 0.52 | Ambiguous | 0.27 | Likely Benign | 0.102 | Likely Benign | -1.87 | Neutral | 0.118 | Benign | 0.054 | Benign | 1.84 | Pathogenic | 0.34 | Tolerated | 0.1387 | 0.3855 | 0 | -1 | -3.2 | 27.07 | ||||||||||||||||||||||||||||||
| c.1106C>G | T369R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T369R is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438011‑C‑G). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. Rosetta and Foldetta are uncertain, so their results are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is also uncertain. Overall, the majority of evidence (nine benign vs two pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | 6-33438011-C-G | 3 | 1.93e-6 | -6.772 | Likely Benign | 0.571 | Likely Pathogenic | Likely Benign | -0.27 | Likely Benign | 0.1 | 1.48 | Ambiguous | 0.61 | Ambiguous | 0.29 | Likely Benign | 0.148 | Likely Benign | -2.15 | Neutral | 0.244 | Benign | 0.107 | Benign | 1.72 | Pathogenic | 0.32 | Tolerated | 3.42 | 19 | 0.1217 | 0.3737 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||
| c.1108G>T | G370C 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G370C has no ClinVar entry and is not reported in gnomAD. Functional prediction tools fall into two groups: benign predictions come from premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from REVEL, FoldX, Rosetta, Foldetta, and FATHMM. ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. No prediction or stability result is missing. Overall, the majority of tools predict a benign effect, and the high‑accuracy consensus also leans benign, while only one high‑accuracy method (Foldetta) suggests pathogenicity. Thus, the variant is most likely benign based on the available predictions, and this assessment does not contradict any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.461924 | Structured | 0.434325 | Uncertain | 0.359 | 0.720 | 0.500 | -7.071 | In-Between | 0.119 | Likely Benign | Likely Benign | 3.01 | Destabilizing | 2.1 | 2.03 | Destabilizing | 2.52 | Destabilizing | 0.29 | Likely Benign | 0.511 | Likely Pathogenic | -1.00 | Neutral | 0.353 | Benign | 0.010 | Benign | 1.32 | Pathogenic | 0.06 | Tolerated | 0.1245 | 0.4412 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||
| c.1109G>A | G370D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G370D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX, FATHMM, and AlphaMissense‑Default. The high‑accuracy methods give mixed results: AlphaMissense‑Optimized predicts benign, Foldetta (a folding‑stability method that integrates FoldX‑MD and Rosetta outputs) predicts pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive because it yields an equal split of benign and pathogenic calls. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.461924 | Structured | 0.434325 | Uncertain | 0.359 | 0.720 | 0.500 | -5.332 | Likely Benign | 0.597 | Likely Pathogenic | Likely Benign | 3.64 | Destabilizing | 3.8 | 0.83 | Ambiguous | 2.24 | Destabilizing | 0.30 | Likely Benign | 0.372 | Likely Benign | -0.44 | Neutral | 0.007 | Benign | 0.001 | Benign | 1.32 | Pathogenic | 0.64 | Tolerated | 0.1632 | 0.1494 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||
| c.1114G>A | G372R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G372R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, Rosetta, Foldetta, SIFT, FATHMM, and AlphaMissense‑Default. Two tools, FoldX and ESM1b, returned uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta predicting pathogenic. Overall, the majority of predictions (seven pathogenic vs. five benign) and the consensus of high‑accuracy methods indicate a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.433034 | Structured | 0.430335 | Uncertain | 0.322 | 0.774 | 0.375 | -7.344 | In-Between | 0.617 | Likely Pathogenic | Likely Benign | 1.49 | Ambiguous | 0.3 | 2.87 | Destabilizing | 2.18 | Destabilizing | 0.20 | Likely Benign | 0.572 | Likely Pathogenic | -0.61 | Neutral | 0.001 | Benign | 0.001 | Benign | -0.74 | Pathogenic | 0.02 | Affected | 0.1323 | 0.4313 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1114G>C | G372R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G372R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, Rosetta, Foldetta, SIFT, FATHMM, and AlphaMissense‑Default. Two tools (FoldX and ESM1b) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. Overall, the majority of predictions (seven pathogenic vs five benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.433034 | Structured | 0.430335 | Uncertain | 0.322 | 0.774 | 0.375 | -7.344 | In-Between | 0.617 | Likely Pathogenic | Likely Benign | 1.49 | Ambiguous | 0.3 | 2.87 | Destabilizing | 2.18 | Destabilizing | 0.20 | Likely Benign | 0.572 | Likely Pathogenic | -0.61 | Neutral | 0.001 | Benign | 0.001 | Benign | -0.74 | Pathogenic | 0.02 | Affected | 0.1323 | 0.4313 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1114G>T | G372W 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G372W has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, FoldX, polyPhen‑2 HumDiv, SIFT, ESM1b, and FATHMM. The remaining tools (Rosetta, Foldetta, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.433034 | Structured | 0.430335 | Uncertain | 0.322 | 0.774 | 0.375 | -8.262 | Likely Pathogenic | 0.478 | Ambiguous | Likely Benign | 2.28 | Destabilizing | 0.5 | 1.14 | Ambiguous | 1.71 | Ambiguous | 0.21 | Likely Benign | 0.649 | Likely Pathogenic | -1.25 | Neutral | 0.657 | Possibly Damaging | 0.075 | Benign | -0.74 | Pathogenic | 0.00 | Affected | 0.1057 | 0.4368 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||
| c.1115G>A | G372E 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G372E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are REVEL, Rosetta, Foldetta, FATHMM, and AlphaMissense‑Default; FoldX is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while Foldetta predicts pathogenic. The SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split and is treated as unavailable. Overall, the majority of evidence (seven benign vs. five pathogenic) supports a benign classification. This conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.433034 | Structured | 0.430335 | Uncertain | 0.322 | 0.774 | 0.375 | -6.682 | Likely Benign | 0.604 | Likely Pathogenic | Likely Benign | 1.58 | Ambiguous | 0.4 | 2.91 | Destabilizing | 2.25 | Destabilizing | 0.34 | Likely Benign | 0.566 | Likely Pathogenic | -0.81 | Neutral | 0.001 | Benign | 0.001 | Benign | -0.74 | Pathogenic | 0.08 | Tolerated | 0.1606 | 0.4103 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1117G>A | G373R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G373R is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438022‑G‑A). Prediction tools that classify the variant as benign include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized, and premPS. Those that predict pathogenicity are REVEL, FoldX, Foldetta, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as pathogenic. No prediction or folding result is missing or inconclusive. Overall, the majority of tools (six versus five) and the consensus of high‑accuracy methods lean toward a benign effect. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.529623 | Disordered | 0.429267 | Uncertain | 0.295 | 0.799 | 0.625 | 6-33438022-G-A | 1 | 6.28e-7 | -7.878 | In-Between | 0.653 | Likely Pathogenic | Likely Benign | 4.28 | Destabilizing | 3.5 | 0.14 | Likely Benign | 2.21 | Destabilizing | 0.21 | Likely Benign | 0.510 | Likely Pathogenic | -0.64 | Neutral | 0.001 | Benign | 0.000 | Benign | 3.90 | Benign | 0.01 | Affected | 3.53 | 16 | 0.1089 | 0.4524 | -2 | -3 | -4.1 | 99.14 | |||||||||||||||||||||||||
| c.1117G>C | G373R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G373R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Rosetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Foldetta, SIFT, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as pathogenic. Overall, the majority of tools (seven benign vs five pathogenic) lean toward a benign interpretation, and this does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.529623 | Disordered | 0.429267 | Uncertain | 0.295 | 0.799 | 0.625 | -7.878 | In-Between | 0.653 | Likely Pathogenic | Likely Benign | 4.28 | Destabilizing | 3.5 | 0.14 | Likely Benign | 2.21 | Destabilizing | 0.21 | Likely Benign | 0.522 | Likely Pathogenic | -0.64 | Neutral | 0.001 | Benign | 0.000 | Benign | 3.90 | Benign | 0.01 | Affected | 3.53 | 16 | 0.1089 | 0.4524 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||
| c.1118G>A | G373E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G373E is listed in ClinVar with an Uncertain significance and is not present in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, Foldetta, SIFT, and AlphaMissense‑Default. Predictions from Rosetta and ESM1b are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as pathogenic. Overall, the majority of evidence points to a benign impact, which does not contradict the ClinVar status of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.529623 | Disordered | 0.429267 | Uncertain | 0.295 | 0.799 | 0.625 | Uncertain | 1 | -7.281 | In-Between | 0.569 | Likely Pathogenic | Likely Benign | 4.13 | Destabilizing | 3.2 | 0.52 | Ambiguous | 2.33 | Destabilizing | -0.02 | Likely Benign | 0.420 | Likely Benign | -0.69 | Neutral | 0.001 | Benign | 0.000 | Benign | 3.90 | Benign | 0.01 | Affected | 0.1572 | 0.4309 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||
| c.1121C>A | S374Y 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant S374Y is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, whereas polyPhen‑2 HumDiv and SIFT predict a pathogenic impact. Uncertain calls come from FoldX, Rosetta, Foldetta, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome; the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive, and Foldetta likewise yields an uncertain stability change. Overall, the majority of available predictions favor a benign effect, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.642678 | Disordered | 0.428948 | Uncertain | 0.333 | 0.812 | 0.625 | Uncertain | 1 | -7.774 | In-Between | 0.344 | Ambiguous | Likely Benign | 0.71 | Ambiguous | 1.2 | 0.66 | Ambiguous | 0.69 | Ambiguous | -0.02 | Likely Benign | 0.310 | Likely Benign | -1.18 | Neutral | 0.875 | Possibly Damaging | 0.271 | Benign | 5.41 | Benign | 0.01 | Affected | 4.32 | 13 | 0.1175 | 0.6705 | -3 | -2 | -0.5 | 76.10 | 237.3 | -76.9 | 0.5 | 0.4 | 0.5 | 0.3 | Uncertain | Ser374 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because the Ω loop is assumed to directly interact with the membrane, it moves arbitrarily throughout the WT solvent simulations. The Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play major roles in protein functions that require flexibility, and thus, large and relatively hydrophobic residues like tyrosine are rarely tolerated. Additionally, the hydroxyl group of Tyr374 frequently forms various hydrogen bonds with other loop residues in the variant simulations. Although no negative structural effects are observed in the variant simulations, Tyr374 may exert drastic effects on the SynGAP-membrane complex dynamics and stability. However, since the effect on Gly-rich Ω loop dynamics can only be studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||||||
| c.1123G>T | G375W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G375W is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438028‑G‑T). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of evidence, including the high‑accuracy tools, points to a pathogenic effect for G375W. This conclusion is consistent with the absence of a ClinVar classification, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.604312 | Disordered | 0.428340 | Uncertain | 0.301 | 0.836 | 0.625 | 6-33438028-G-T | -9.654 | Likely Pathogenic | 0.464 | Ambiguous | Likely Benign | 4.33 | Destabilizing | 2.0 | 7.01 | Destabilizing | 5.67 | Destabilizing | 0.22 | Likely Benign | 0.450 | Likely Benign | -1.26 | Neutral | 0.992 | Probably Damaging | 0.869 | Possibly Damaging | 1.31 | Pathogenic | 0.01 | Affected | 4.32 | 12 | 0.0992 | 0.4368 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||||||
| c.1124G>A | G375E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G375E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, FATHMM, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) favors pathogenic, and Foldetta also predicts pathogenic. Overall, the majority of tools, including the high‑accuracy ones, indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.604312 | Disordered | 0.428340 | Uncertain | 0.301 | 0.836 | 0.625 | -7.780 | In-Between | 0.600 | Likely Pathogenic | Likely Benign | 2.89 | Destabilizing | 1.4 | 9.47 | Destabilizing | 6.18 | Destabilizing | 0.45 | Likely Benign | 0.545 | Likely Pathogenic | -1.07 | Neutral | 0.845 | Possibly Damaging | 0.244 | Benign | 1.32 | Pathogenic | 0.09 | Tolerated | 0.1619 | 0.4299 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1126G>T | G376C 2D AISynGAP1 missense variant G376C is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from Rosetta, premPS, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic calls come from REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Two tools report uncertainty: Foldetta and ESM1b. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also yields a benign verdict; Foldetta remains uncertain. Overall, the majority of conventional predictors lean toward pathogenicity, whereas the most accurate methods favor a benign effect. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.680603 | Disordered | 0.428979 | Uncertain | 0.326 | 0.869 | 0.625 | Uncertain | 1 | -7.686 | In-Between | 0.125 | Likely Benign | Likely Benign | 2.56 | Destabilizing | 0.5 | 0.22 | Likely Benign | 1.39 | Ambiguous | 0.16 | Likely Benign | 0.560 | Likely Pathogenic | -1.15 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.32 | Pathogenic | 0.01 | Affected | 0.1476 | 0.3929 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||
| c.1127G>A | G376D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G376D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. The remaining tools—Rosetta, Foldetta, premPS, and ESM1b—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) yielding an uncertain stability change. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.680603 | Disordered | 0.428979 | Uncertain | 0.326 | 0.869 | 0.625 | -7.125 | In-Between | 0.569 | Likely Pathogenic | Likely Benign | 3.10 | Destabilizing | 1.1 | -1.08 | Ambiguous | 1.01 | Ambiguous | 0.52 | Ambiguous | 0.572 | Likely Pathogenic | -1.05 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.32 | Pathogenic | 0.09 | Tolerated | 0.1938 | 0.1235 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||
| c.1132G>T | G378C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G378C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of tools (8/12) predict pathogenicity, and the high‑accuracy consensus leans toward benign only for AlphaMissense‑Optimized and SGM Consensus, while Foldetta indicates instability. Thus, the variant is most likely pathogenic based on the preponderance of evidence, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.767246 | Disordered | 0.432858 | Uncertain | 0.341 | 0.915 | 0.625 | -7.981 | In-Between | 0.203 | Likely Benign | Likely Benign | 7.63 | Destabilizing | 2.7 | 12.14 | Destabilizing | 9.89 | Destabilizing | 0.26 | Likely Benign | 0.635 | Likely Pathogenic | -1.18 | Neutral | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 1.32 | Pathogenic | 0.01 | Affected | 0.1595 | 0.4240 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||
| c.1133G>A | G378D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G378D is not reported in ClinVar (ClinVar ID: None) but is present in gnomAD (ID 6‑33438038‑G‑A). Prediction tools that classify the variant as benign include premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Tools that predict pathogenicity are REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. ESM1b is uncertain, and no other high‑accuracy predictions are available. Overall, the majority of evidence supports a pathogenic effect, and this conclusion does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.767246 | Disordered | 0.432858 | Uncertain | 0.341 | 0.915 | 0.625 | 6-33438038-G-A | 1 | 6.97e-7 | -7.767 | In-Between | 0.576 | Likely Pathogenic | Likely Benign | 11.41 | Destabilizing | 5.0 | 11.84 | Destabilizing | 11.63 | Destabilizing | 0.50 | Likely Benign | 0.619 | Likely Pathogenic | -0.63 | Neutral | 1.000 | Probably Damaging | 0.992 | Probably Damaging | 1.32 | Pathogenic | 0.08 | Tolerated | 4.32 | 12 | 0.2130 | 0.2035 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||
| c.1136C>G | S379W 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S379W is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438041‑C‑G). Prediction tools that indicate a benign effect include premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic impact comprise REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus as benign. Because the majority of conventional tools favor pathogenicity while the high‑accuracy subset is split, the overall evidence leans toward a pathogenic effect. This conclusion does not contradict the ClinVar uncertain status, which remains unresolved. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.433206 | Uncertain | 0.327 | 0.931 | 0.625 | Uncertain | 1 | 6-33438041-C-G | -8.898 | Likely Pathogenic | 0.388 | Ambiguous | Likely Benign | 4.32 | Destabilizing | 3.4 | 3.56 | Destabilizing | 3.94 | Destabilizing | 0.16 | Likely Benign | 0.520 | Likely Pathogenic | -1.02 | Neutral | 0.998 | Probably Damaging | 0.844 | Possibly Damaging | 3.82 | Benign | 0.01 | Affected | 4.32 | 11 | 0.1196 | 0.6070 | -2 | -3 | -0.1 | 99.14 | 271.3 | -75.7 | 1.4 | 1.0 | 0.6 | 0.5 | Uncertain | Ser379 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because the Ω loop is assumed to directly interact with the membrane, it moves arbitrarily throughout the WT solvent simulations. The Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play major roles in protein functions that require flexibility, and thus hydrophobic residues like tryptophan are rarely tolerated. Although no major negative structural effects are observed in the variant simulations, Trp379 may exert drastic effects on the SynGAP-membrane complex dynamics and stability. However, since the effect on Gly-rich Ω loop dynamics can only be studied through the SynGAP-membrane complex, no definite conclusions can be drawn | |||||||||||||||||
| c.1138G>A | G380R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 G380R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized, while those that agree on a pathogenic effect are REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, Foldetta predicts a pathogenic outcome, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence (10 pathogenic vs. 4 benign) points to a pathogenic impact. The variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar assertion exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.432982 | Uncertain | 0.316 | 0.939 | 0.750 | -9.205 | Likely Pathogenic | 0.635 | Likely Pathogenic | Likely Benign | 5.94 | Destabilizing | 2.6 | 0.90 | Ambiguous | 3.42 | Destabilizing | 0.11 | Likely Benign | 0.640 | Likely Pathogenic | -0.86 | Neutral | 0.940 | Possibly Damaging | 0.459 | Possibly Damaging | 2.53 | Benign | 0.01 | Affected | 0.1319 | 0.3956 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1138G>C | G380R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 G380R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized, while those that agree on a pathogenic effect are REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, Foldetta predicts pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of tools (10 pathogenic vs 4 benign) and the Foldetta result support a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.432982 | Uncertain | 0.316 | 0.939 | 0.750 | -9.205 | Likely Pathogenic | 0.635 | Likely Pathogenic | Likely Benign | 5.94 | Destabilizing | 2.6 | 0.90 | Ambiguous | 3.42 | Destabilizing | 0.11 | Likely Benign | 0.619 | Likely Pathogenic | -0.86 | Neutral | 0.940 | Possibly Damaging | 0.459 | Possibly Damaging | 2.53 | Benign | 0.01 | Affected | 0.1319 | 0.3956 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1139G>A | G380E 2D AIThe SynGAP1 missense variant G380E has no ClinVar record and is not reported in gnomAD. Prediction tools that agree on a benign effect include Rosetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Foldetta, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give mixed results: AlphaMissense‑Optimized classifies the variant as benign, Foldetta predicts a pathogenic impact on protein stability, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of tools (7 benign vs. 6 pathogenic) lean toward a benign interpretation, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Thus, based on current computational predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.432982 | Uncertain | 0.316 | 0.939 | 0.750 | -9.334 | Likely Pathogenic | 0.617 | Likely Pathogenic | Likely Benign | 6.25 | Destabilizing | 5.1 | -0.28 | Likely Benign | 2.99 | Destabilizing | 0.24 | Likely Benign | 0.582 | Likely Pathogenic | -0.86 | Neutral | 0.056 | Benign | 0.010 | Benign | 2.53 | Benign | 0.03 | Affected | 0.1585 | 0.3741 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1141G>T | G381W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G381W is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438046‑G‑T). Prediction tools that agree on a benign effect include premPS, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments give AlphaMissense‑Optimized a benign call, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) a pathogenic call, and Foldetta (combining FoldX‑MD and Rosetta outputs) a pathogenic call. No prediction or folding result is missing or inconclusive. Overall, the majority of tools and the high‑accuracy methods indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.431692 | Uncertain | 0.301 | 0.951 | 0.750 | 6-33438046-G-T | 1 | 6.24e-7 | -10.173 | Likely Pathogenic | 0.438 | Ambiguous | Likely Benign | 8.81 | Destabilizing | 2.0 | 3.17 | Destabilizing | 5.99 | Destabilizing | 0.02 | Likely Benign | 0.576 | Likely Pathogenic | -1.04 | Neutral | 0.996 | Probably Damaging | 0.920 | Probably Damaging | 1.31 | Pathogenic | 0.01 | Affected | 4.32 | 9 | 0.0993 | 0.4000 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||||
| c.1142G>A | G381E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G381E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Foldetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Two tools give uncertain results: Rosetta and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. With seven pathogenic versus four benign predictions and two high‑accuracy tools supporting pathogenicity, the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.431692 | Uncertain | 0.301 | 0.951 | 0.750 | -9.360 | Likely Pathogenic | 0.540 | Ambiguous | Likely Benign | 5.52 | Destabilizing | 1.1 | 0.53 | Ambiguous | 3.03 | Destabilizing | 0.24 | Likely Benign | 0.588 | Likely Pathogenic | -0.71 | Neutral | 0.985 | Probably Damaging | 0.720 | Possibly Damaging | 1.32 | Pathogenic | 0.11 | Tolerated | 0.1554 | 0.3735 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1145G>A | G382E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G382E is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438050‑G‑A). Prediction tools that agree on a benign effect include premPS, PROVEAN, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) predicts pathogenic. Overall, the majority of evidence points to a pathogenic impact for G382E, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429690 | Uncertain | 0.315 | 0.951 | 0.750 | 6-33438050-G-A | -9.570 | Likely Pathogenic | 0.564 | Ambiguous | Likely Benign | 4.86 | Destabilizing | 1.7 | 6.64 | Destabilizing | 5.75 | Destabilizing | 0.48 | Likely Benign | 0.594 | Likely Pathogenic | -0.96 | Neutral | 0.994 | Probably Damaging | 0.986 | Probably Damaging | 1.32 | Pathogenic | 0.03 | Affected | 4.32 | 9 | 0.1775 | 0.3932 | -2 | 0 | -3.1 | 72.06 | |||||||||||||||||||||||||||
| c.1147G>A | G383R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G383R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that agree on a pathogenic effect are FoldX, Rosetta, Foldetta, polyPhen2_HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, Foldetta predicts a pathogenic outcome, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs. two benign). Overall, the majority of predictions lean toward pathogenicity, and this does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | -9.067 | Likely Pathogenic | 0.660 | Likely Pathogenic | Likely Benign | 4.03 | Destabilizing | 2.3 | 3.36 | Destabilizing | 3.70 | Destabilizing | 0.29 | Likely Benign | 0.449 | Likely Benign | -0.84 | Neutral | 0.498 | Possibly Damaging | 0.119 | Benign | 4.10 | Benign | 0.00 | Affected | 0.1295 | 0.3741 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1147G>C | G383R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G383R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that agree on a pathogenic effect are FoldX, Rosetta, Foldetta, polyPhen2_HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, Foldetta predicts a pathogenic outcome, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs. two benign). Overall, the majority of predictions lean toward pathogenicity, and this does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | -9.067 | Likely Pathogenic | 0.660 | Likely Pathogenic | Likely Benign | 4.03 | Destabilizing | 2.3 | 3.36 | Destabilizing | 3.70 | Destabilizing | 0.29 | Likely Benign | 0.440 | Likely Benign | -0.84 | Neutral | 0.498 | Possibly Damaging | 0.119 | Benign | 4.10 | Benign | 0.00 | Affected | 0.1295 | 0.3741 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1147G>T | G383W 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G383W is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438052‑G‑T). Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as benign, and Foldetta as pathogenic. Because the majority of conventional predictors favor pathogenicity while the high‑accuracy subset is split, the overall evidence leans toward a pathogenic interpretation. This conclusion does not conflict with the ClinVar uncertain status, which reflects the current lack of definitive clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | Uncertain | 1 | 6-33438052-G-T | 1 | 6.22e-7 | -10.161 | Likely Pathogenic | 0.439 | Ambiguous | Likely Benign | 5.81 | Destabilizing | 3.6 | 4.44 | Destabilizing | 5.13 | Destabilizing | 0.08 | Likely Benign | 0.469 | Likely Benign | -1.01 | Neutral | 0.959 | Probably Damaging | 0.704 | Possibly Damaging | 4.09 | Benign | 0.00 | Affected | 4.32 | 7 | 0.0972 | 0.3785 | -2 | -7 | -0.5 | 129.16 | |||||||||||||||||||||||
| c.1148G>A | G383E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G383E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Tools that predict a pathogenic effect are FoldX, Rosetta, SIFT, ESM1b, AlphaMissense‑Default, and the protein‑folding stability method Foldetta. High‑accuracy assessments give a benign result from AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (pathogenic), FATHMM (benign), and PROVEAN (benign), is inconclusive (tie). Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a pathogenic effect. Overall, the majority of predictions (seven pathogenic vs. six benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | -9.535 | Likely Pathogenic | 0.570 | Likely Pathogenic | Likely Benign | 4.09 | Destabilizing | 2.4 | 2.62 | Destabilizing | 3.36 | Destabilizing | 0.39 | Likely Benign | 0.308 | Likely Benign | -0.78 | Neutral | 0.000 | Benign | 0.002 | Benign | 4.10 | Benign | 0.01 | Affected | 0.1582 | 0.3741 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1150G>T | G384C 2D AIThe SynGAP1 missense variant G384C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts pathogenic; and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split and is treated as unavailable. Overall, the majority of predictions (seven pathogenic vs. five benign) and the pathogenic Foldetta result indicate that the variant is most likely pathogenic, with no contradiction to ClinVar status because the variant is not yet classified there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.427831 | Uncertain | 0.323 | 0.934 | 0.750 | -8.363 | Likely Pathogenic | 0.135 | Likely Benign | Likely Benign | 2.41 | Destabilizing | 2.0 | 5.50 | Destabilizing | 3.96 | Destabilizing | 0.17 | Likely Benign | 0.456 | Likely Benign | -1.07 | Neutral | 0.998 | Probably Damaging | 0.993 | Probably Damaging | 1.32 | Pathogenic | 0.01 | Affected | 0.1530 | 0.4240 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||
| c.1154C>G | S385W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S385W is listed in ClinVar as Benign (ClinVar ID 218691.0) and is present in gnomAD (ID 6‑33438059‑C‑G). Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. Predictions that are inconclusive or unavailable are FoldX, Rosetta, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM Consensus as Benign, and Foldetta as Uncertain. Taken together, the majority of evidence points to a benign impact, which aligns with the ClinVar classification and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.733139 | Disordered | 0.425480 | Uncertain | 0.341 | 0.925 | 0.750 | Benign | 1 | 6-33438059-C-G | -9.353 | Likely Pathogenic | 0.362 | Ambiguous | Likely Benign | 0.53 | Ambiguous | 0.2 | 0.69 | Ambiguous | 0.61 | Ambiguous | 0.00 | Likely Benign | 0.373 | Likely Benign | -0.84 | Neutral | 0.986 | Probably Damaging | 0.968 | Probably Damaging | 4.63 | Benign | 0.00 | Affected | 4.32 | 3 | 0.1272 | 0.6670 | -2 | -3 | -0.1 | 99.14 | 260.4 | -71.2 | 0.5 | 1.3 | 0.7 | 0.4 | Uncertain | Ser385 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because the Ω loop is assumed to directly interact with the membrane, it moves arbitrarily throughout the WT solvent simulations. The Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play major roles in protein functions that require flexibility, and thus hydrophobic residues like tryptophan are rarely tolerated. Although no major negative structural effects are observed in the variant simulations, Trp385 may exert drastic effects on the SynGAP-membrane complex dynamics and stability. However, since the effects on Gly-rich Ω loop dynamics can only be studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | 10.1016/j.ajhg.2020.11.011 | ||||||||||||||||
| c.1156G>A | G386R 2D 3DClick to see structure in 3D Viewer AIClinVar reports no entry for this SynGAP1 G386R variant, and it is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are FoldX, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default, while Rosetta is uncertain. High‑accuracy methods give a benign call from AlphaMissense‑Optimized, a pathogenic result from Foldetta, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Overall, the evidence is mixed; the variant is most likely benign, and this assessment does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.733139 | Disordered | 0.424156 | Uncertain | 0.334 | 0.898 | 0.750 | -9.024 | Likely Pathogenic | 0.709 | Likely Pathogenic | Likely Benign | 3.62 | Destabilizing | 2.9 | 1.07 | Ambiguous | 2.35 | Destabilizing | 0.29 | Likely Benign | 0.453 | Likely Benign | -0.82 | Neutral | 0.753 | Possibly Damaging | 0.220 | Benign | 4.03 | Benign | 0.01 | Affected | 0.1329 | 0.4032 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1156G>C | G386R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G386R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Computational predictors that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise FoldX, polyPhen‑2 (HumDiv), SIFT, ESM1b, AlphaMissense‑Default, and Foldetta; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the balance of evidence favors a pathogenic classification. This conclusion is not contradicted by ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.733139 | Disordered | 0.424156 | Uncertain | 0.334 | 0.898 | 0.750 | -9.024 | Likely Pathogenic | 0.709 | Likely Pathogenic | Likely Benign | 3.62 | Destabilizing | 2.9 | 1.07 | Ambiguous | 2.35 | Destabilizing | 0.29 | Likely Benign | 0.453 | Likely Benign | -0.82 | Neutral | 0.753 | Possibly Damaging | 0.220 | Benign | 4.03 | Benign | 0.01 | Affected | 0.1329 | 0.4032 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1156G>T | G386W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G386W is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Computational predictors that classify the change as benign include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. High‑accuracy assessments give a benign verdict from AlphaMissense‑Optimized, a benign consensus from the SGM method (majority of the four contributing tools are benign), and a pathogenic result from Foldetta. Uncertain calls from AlphaMissense‑Default and Rosetta are treated as unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not conflict with the lack of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.733139 | Disordered | 0.424156 | Uncertain | 0.334 | 0.898 | 0.750 | -10.389 | Likely Pathogenic | 0.519 | Ambiguous | Likely Benign | 6.07 | Destabilizing | 5.5 | 1.28 | Ambiguous | 3.68 | Destabilizing | -0.23 | Likely Benign | 0.471 | Likely Benign | -0.85 | Neutral | 0.996 | Probably Damaging | 0.920 | Probably Damaging | 3.90 | Benign | 0.00 | Affected | 0.1058 | 0.4676 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||
| c.1157G>A | G386E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G386E is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438062‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are FoldX, Foldetta, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. Uncertain predictions come from Rosetta and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Overall, the majority of tools predict a pathogenic impact, suggesting the variant is most likely pathogenic, which does not contradict the ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.733139 | Disordered | 0.424156 | Uncertain | 0.334 | 0.898 | 0.750 | Uncertain | 1 | 6-33438062-G-A | -9.286 | Likely Pathogenic | 0.686 | Likely Pathogenic | Likely Benign | 3.69 | Destabilizing | 2.9 | 0.79 | Ambiguous | 2.24 | Destabilizing | 0.54 | Ambiguous | 0.447 | Likely Benign | -0.83 | Neutral | 0.860 | Possibly Damaging | 0.354 | Benign | 3.93 | Benign | 0.01 | Affected | 4.32 | 3 | 0.1543 | 0.3354 | -2 | 0 | -3.1 | 72.06 | |||||||||||||||||||||||||
| c.1159G>T | G387C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G387C is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438064‑G‑T). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, SIFT, and FATHMM; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of high‑confidence tools lean toward a benign interpretation, and this does not contradict the ClinVar status, which has no pathogenic classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.642678 | Disordered | 0.422910 | Uncertain | 0.293 | 0.861 | 0.750 | 6-33438064-G-T | 1 | 6.21e-7 | -7.609 | In-Between | 0.146 | Likely Benign | Likely Benign | 2.88 | Destabilizing | 0.6 | 2.34 | Destabilizing | 2.61 | Destabilizing | -0.03 | Likely Benign | 0.430 | Likely Benign | -0.58 | Neutral | 0.859 | Possibly Damaging | 0.346 | Benign | 1.32 | Pathogenic | 0.01 | Affected | 4.32 | 3 | 0.1589 | 0.4240 | -3 | -3 | 2.9 | 46.09 | |||||||||||||||||||||||||
| c.1162G>T | G388C 2D 3DClick to see structure in 3D Viewer AISynGAP1 G388C is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from AlphaMissense‑Default, AlphaMissense‑Optimized, premPS, and PROVEAN, while pathogenic predictions arise from REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. High‑accuracy assessments give a mixed picture: AlphaMissense‑Optimized predicts benign, Foldetta predicts pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Because the majority of evidence points to pathogenicity and there is no ClinVar entry to contradict this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.736850 | Disordered | 0.420316 | Uncertain | 0.319 | 0.827 | 0.750 | -8.088 | Likely Pathogenic | 0.121 | Likely Benign | Likely Benign | 4.01 | Destabilizing | 3.7 | 4.95 | Destabilizing | 4.48 | Destabilizing | 0.03 | Likely Benign | 0.603 | Likely Pathogenic | -0.98 | Neutral | 0.998 | Probably Damaging | 0.993 | Probably Damaging | 1.32 | Pathogenic | 0.01 | Affected | 0.1497 | 0.4112 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||
| c.1168G>T | G390W 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G390W is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include AlphaMissense‑Optimized, premPS, and PROVEAN, whereas the remaining tools—REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts benign, but the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic verdict, and Foldetta also indicates pathogenic. With the majority of evidence pointing to a deleterious effect, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | -9.405 | Likely Pathogenic | 0.487 | Ambiguous | Likely Benign | 3.69 | Destabilizing | 1.9 | 5.31 | Destabilizing | 4.50 | Destabilizing | 0.14 | Likely Benign | 0.639 | Likely Pathogenic | -1.11 | Neutral | 0.987 | Probably Damaging | 0.744 | Possibly Damaging | 1.31 | Pathogenic | 0.00 | Affected | 0.0917 | 0.4368 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||
| c.1169G>A | G390E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G390E is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that classify the variant as benign include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that predict pathogenicity are REVEL, FoldX, Rosetta, Foldetta, SIFT, FATHMM, and AlphaMissense‑Default. A high‑accuracy assessment shows AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of predictions support a pathogenic effect, and this aligns with the ClinVar designation of uncertain significance rather than contradicting it. Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | Uncertain | 1 | -7.913 | In-Between | 0.646 | Likely Pathogenic | Likely Benign | 2.61 | Destabilizing | 0.9 | 4.28 | Destabilizing | 3.45 | Destabilizing | 0.47 | Likely Benign | 0.575 | Likely Pathogenic | -0.87 | Neutral | 0.276 | Benign | 0.045 | Benign | 1.32 | Pathogenic | 0.05 | Affected | 4.32 | 8 | 0.1595 | 0.4309 | 0 | -2 | -3.1 | 72.06 | 241.5 | -108.4 | 0.6 | 0.5 | -0.1 | 0.1 | Uncertain | Gly390 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364 and res. Ala399-Ile411). The Ω loop is assumed to directly interact with the membrane, and it is observed to move arbitrarily throughout the WT solvent simulations. This loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play significant roles in protein functions that require flexibility, and so they are rich in glycine residues, prolines, and to a lesser extent, small hydrophilic residues to ensure maximum flexibility. Thus, the variant’s Glu390 may not be as well tolerated in the Ω loop. Additionally, the carboxylate group of Glu390 occasionally forms H-bonds with other loop residues in the variant simulations. The interaction between the acidic carboxylate side chain and the acidic membrane lipids may further influence the SynGAP-membrane complex. However, since the effects on the Gly-rich Ω loop dynamics can only be well studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||||||
| c.1169G>T | G390V 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G390V has no ClinVar entry and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from REVEL, FoldX, Rosetta, Foldetta, SIFT, and FATHMM, while ESM1b remains uncertain. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default (benign), ESM1b (uncertain), FATHMM (pathogenic), and PROVEAN (benign), also yields a benign verdict; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, indicates pathogenic. With two high‑accuracy tools supporting benign and one supporting pathogenic, the overall evidence leans toward a benign effect. This conclusion does not contradict ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | -7.137 | In-Between | 0.146 | Likely Benign | Likely Benign | 3.74 | Destabilizing | 0.6 | 4.88 | Destabilizing | 4.31 | Destabilizing | -0.16 | Likely Benign | 0.509 | Likely Pathogenic | -0.88 | Neutral | 0.002 | Benign | 0.002 | Benign | 1.32 | Pathogenic | 0.01 | Affected | 0.1537 | 0.4004 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||
| c.1171G>T | G391C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G391C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include AlphaMissense‑Default, AlphaMissense‑Optimized, premPS, and PROVEAN, whereas the remaining tools—REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM—predict it to be pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence points to a pathogenic effect. This prediction does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.637480 | Disordered | 0.409509 | Uncertain | 0.279 | 0.741 | 0.750 | -8.596 | Likely Pathogenic | 0.123 | Likely Benign | Likely Benign | 2.65 | Destabilizing | 0.7 | 5.03 | Destabilizing | 3.84 | Destabilizing | 0.11 | Likely Benign | 0.640 | Likely Pathogenic | -1.29 | Neutral | 1.000 | Probably Damaging | 0.970 | Probably Damaging | 1.32 | Pathogenic | 0.03 | Affected | 0.1483 | 0.4225 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||
| c.1172G>A | G391D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G391D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. Two tools, Rosetta and Foldetta, return uncertain results. High‑accuracy methods give a benign call from AlphaMissense‑Optimized; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is also inconclusive. Overall, six tools favor pathogenicity while five favor benignity, with two uncertain. Thus, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.637480 | Disordered | 0.409509 | Uncertain | 0.279 | 0.741 | 0.750 | -4.651 | Likely Benign | 0.674 | Likely Pathogenic | Likely Benign | 2.59 | Destabilizing | 1.1 | 1.26 | Ambiguous | 1.93 | Ambiguous | 0.22 | Likely Benign | 0.562 | Likely Pathogenic | -0.95 | Neutral | 0.999 | Probably Damaging | 0.960 | Probably Damaging | 1.32 | Pathogenic | 0.29 | Tolerated | 0.1900 | 0.1305 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||
| c.1175A>C | K392T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, PROVEAN, and SIFT. Two tools—FoldX and AlphaMissense‑Default—return uncertain results and are treated as unavailable. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign; and Foldetta, a protein‑folding stability method, also predicts benign. No ClinVar entry exists to contradict these predictions. Based on the collective evidence, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -3.224 | Likely Benign | 0.481 | Ambiguous | Likely Benign | 0.52 | Ambiguous | 0.1 | -0.29 | Likely Benign | 0.12 | Likely Benign | 0.05 | Likely Benign | 0.512 | Likely Pathogenic | -3.14 | Deleterious | 0.276 | Benign | 0.045 | Benign | 4.60 | Benign | 0.02 | Affected | 0.2807 | 0.4053 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||
| c.1175A>T | K392M 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 K392M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, ESM1b, and FATHMM, whereas those that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The remaining tools—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—return uncertain or inconclusive results. High‑accuracy methods give no definitive signal: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable, and Foldetta is uncertain. Overall, the majority of available predictions (six pathogenic vs. three benign) lean toward a pathogenic impact. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -3.856 | Likely Benign | 0.788 | Likely Pathogenic | Ambiguous | 0.52 | Ambiguous | 0.1 | 0.67 | Ambiguous | 0.60 | Ambiguous | -0.09 | Likely Benign | 0.665 | Likely Pathogenic | -3.24 | Deleterious | 0.952 | Possibly Damaging | 0.496 | Possibly Damaging | 4.59 | Benign | 0.00 | Affected | 0.1799 | 0.4941 | 0 | -1 | 5.8 | 3.02 | ||||||||||||||||||||||||||||||
| c.1176G>C | K392N 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 K392N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts benign. Overall, the majority of evidence (9 benign vs 3 pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.136 | Likely Benign | 0.780 | Likely Pathogenic | Likely Benign | 0.24 | Likely Benign | 0.1 | -0.01 | Likely Benign | 0.12 | Likely Benign | 0.20 | Likely Benign | 0.229 | Likely Benign | -2.61 | Deleterious | 0.276 | Benign | 0.083 | Benign | 4.60 | Benign | 0.02 | Affected | 0.4586 | 0.2454 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.1176G>T | K392N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split and is therefore treated as unavailable. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the majority of evidence (nine benign vs. three pathogenic predictions) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.136 | Likely Benign | 0.780 | Likely Pathogenic | Likely Benign | 0.24 | Likely Benign | 0.1 | -0.01 | Likely Benign | 0.12 | Likely Benign | 0.20 | Likely Benign | 0.229 | Likely Benign | -2.61 | Deleterious | 0.276 | Benign | 0.083 | Benign | 4.60 | Benign | 0.02 | Affected | 0.4586 | 0.2454 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.1178G>T | G393V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 G393V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are REVEL, FoldX, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Rosetta is uncertain and is treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while Foldetta predicts pathogenic. The SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑vs‑2 split. Overall, the majority of evidence (8 pathogenic vs. 4 benign) points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.538167 | Disordered | 0.402365 | Uncertain | 0.333 | 0.670 | 0.625 | -6.358 | Likely Benign | 0.154 | Likely Benign | Likely Benign | 5.56 | Destabilizing | 2.3 | -0.72 | Ambiguous | 2.42 | Destabilizing | -0.01 | Likely Benign | 0.639 | Likely Pathogenic | -2.69 | Deleterious | 0.982 | Probably Damaging | 0.648 | Possibly Damaging | 1.32 | Pathogenic | 0.01 | Affected | 0.1712 | 0.4144 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||
| c.1180A>C | K394Q 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 K394Q missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic outcome are SIFT and Rosetta. The remaining tools—Foldetta, premPS, ESM1b, and AlphaMissense‑Default—return uncertain results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also leans toward benign, with two benign votes and two uncertain votes. Foldetta’s stability prediction is uncertain and thus not considered. Overall, the majority of reliable predictions indicate a benign effect, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | -7.261 | In-Between | 0.468 | Ambiguous | Likely Benign | 0.15 | Likely Benign | 0.0 | 2.00 | Destabilizing | 1.08 | Ambiguous | 0.64 | Ambiguous | 0.330 | Likely Benign | -2.46 | Neutral | 0.001 | Benign | 0.009 | Benign | 4.61 | Benign | 0.01 | Affected | 0.5365 | 0.2106 | Weaken | 1 | 1 | 0.4 | -0.04 | |||||||||||||||||||||||||||||
| c.1180A>G | K394E 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K394E is not listed in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438085‑A‑G). Prediction tools that agree on a benign effect include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Those that predict a pathogenic effect are premPS, PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. No prediction or folding‑stability result is available that decisively supports either outcome. Overall, the majority of tools (six benign vs four pathogenic) lean toward a benign interpretation, and this assessment does not contradict the absence of a ClinVar classification. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | 6-33438085-A-G | 1 | 6.20e-7 | -6.903 | Likely Benign | 0.896 | Likely Pathogenic | Ambiguous | 0.07 | Likely Benign | 0.1 | 3.71 | Destabilizing | 1.89 | Ambiguous | 1.20 | Destabilizing | 0.446 | Likely Benign | -2.54 | Deleterious | 0.063 | Benign | 0.038 | Benign | 4.61 | Benign | 0.04 | Affected | 3.44 | 14 | 0.4556 | 0.1916 | 1 | 0 | 0.4 | 0.94 | |||||||||||||||||||||||||
| c.1181A>C | K394T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K394T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. Predictions that are uncertain or inconclusive are FoldX, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, the balance of evidence (five benign versus three pathogenic predictions) suggests the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | -6.487 | Likely Benign | 0.599 | Likely Pathogenic | Likely Benign | 0.50 | Ambiguous | 0.1 | 2.46 | Destabilizing | 1.48 | Ambiguous | 0.57 | Ambiguous | 0.482 | Likely Benign | -3.35 | Deleterious | 0.247 | Benign | 0.166 | Benign | 4.61 | Benign | 0.01 | Affected | 0.2727 | 0.4453 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||
| c.1182A>C | K394N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K394N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, FATHMM, and polyPhen‑2 HumVar. Those that agree on a pathogenic effect are Rosetta, PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. Predictions that are uncertain or inconclusive are Foldetta, premPS, ESM1b, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, Foldetta as uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of available predictions lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | -7.408 | In-Between | 0.861 | Likely Pathogenic | Ambiguous | 0.08 | Likely Benign | 0.1 | 2.02 | Destabilizing | 1.05 | Ambiguous | 0.66 | Ambiguous | 0.299 | Likely Benign | -3.17 | Deleterious | 0.535 | Possibly Damaging | 0.188 | Benign | 4.60 | Benign | 0.01 | Affected | 0.4353 | 0.2654 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.1182A>T | K394N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K394N missense variant has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a mixed signal: benign calls come from REVEL, FoldX, FATHMM, and polyPhen‑2 HumVar, while pathogenic calls come from Rosetta, PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. Four tools (Foldetta, premPS, ESM1b, AlphaMissense‑Optimized) return uncertain results. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves as pathogenic; and Foldetta is uncertain. Taken together, the majority of evidence—including the high‑accuracy consensus—points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | -7.408 | In-Between | 0.861 | Likely Pathogenic | Ambiguous | 0.08 | Likely Benign | 0.1 | 2.02 | Destabilizing | 1.05 | Ambiguous | 0.66 | Ambiguous | 0.299 | Likely Benign | -3.17 | Deleterious | 0.535 | Possibly Damaging | 0.188 | Benign | 4.60 | Benign | 0.01 | Affected | 0.4353 | 0.2654 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.1183G>A | G395R 2D AIThe SynGAP1 missense variant G395R has no ClinVar record and is not reported in gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from SIFT, ESM1b, and AlphaMissense‑Default. Two tools, FoldX and Foldetta, yield uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta remains uncertain. Overall, the balance of evidence favors a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.513880 | Disordered | 0.396199 | Uncertain | 0.474 | 0.601 | 0.500 | -9.851 | Likely Pathogenic | 0.768 | Likely Pathogenic | Likely Benign | 1.26 | Ambiguous | 2.1 | -0.25 | Likely Benign | 0.51 | Ambiguous | 0.50 | Likely Benign | 0.430 | Likely Benign | -2.01 | Neutral | 0.037 | Benign | 0.027 | Benign | 4.23 | Benign | 0.02 | Affected | 0.0953 | 0.4313 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1183G>C | G395R 2D AIThe SynGAP1 missense variant G395R has no ClinVar record and is not reported in gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from SIFT, ESM1b, and AlphaMissense‑Default. Two tools, FoldX and Foldetta, yield uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta remains uncertain. Overall, the balance of evidence favors a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.513880 | Disordered | 0.396199 | Uncertain | 0.474 | 0.601 | 0.500 | -9.851 | Likely Pathogenic | 0.768 | Likely Pathogenic | Likely Benign | 1.26 | Ambiguous | 2.1 | -0.25 | Likely Benign | 0.51 | Ambiguous | 0.50 | Likely Benign | 0.430 | Likely Benign | -2.01 | Neutral | 0.037 | Benign | 0.027 | Benign | 4.23 | Benign | 0.02 | Affected | 0.0953 | 0.4313 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1189T>C | C397R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, and AlphaMissense‑Default. The remaining tools (FoldX, premPS, ESM1b) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign (2 benign vs. 1 pathogenic, 1 uncertain), and Foldetta predicts a benign impact on protein folding stability. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -7.094 | In-Between | 0.708 | Likely Pathogenic | Likely Benign | 0.55 | Ambiguous | 0.8 | 0.40 | Likely Benign | 0.48 | Likely Benign | 0.96 | Ambiguous | 0.514 | Likely Pathogenic | -1.46 | Neutral | 0.480 | Possibly Damaging | 0.226 | Benign | 4.65 | Benign | 0.11 | Tolerated | 0.1780 | 0.1853 | -4 | -3 | -7.0 | 53.05 | ||||||||||||||||||||||||||||||
| c.1190G>A | C397Y 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, Rosetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and the combined Foldetta method. Uncertain results come from AlphaMissense‑Default and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as pathogenic. Overall, the majority of tools and the high‑accuracy consensus lean toward a benign interpretation, and this does not contradict any ClinVar classification (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -7.213 | In-Between | 0.397 | Ambiguous | Likely Benign | 2.01 | Destabilizing | 2.3 | 8.64 | Destabilizing | 5.33 | Destabilizing | 0.12 | Likely Benign | 0.455 | Likely Benign | -1.82 | Neutral | 0.952 | Possibly Damaging | 0.497 | Possibly Damaging | 4.64 | Benign | 0.07 | Tolerated | 0.1299 | 0.5084 | 0 | -2 | -3.8 | 60.04 | ||||||||||||||||||||||||||||||
| c.1192C>A | P398T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant P398T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, and SIFT. The remaining tools (Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy methods give a benign consensus: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta is uncertain. Overall, the majority of high‑confidence predictions lean toward a benign impact, although several other predictors indicate pathogenicity. There is no conflict with ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | -6.670 | Likely Benign | 0.536 | Ambiguous | Likely Benign | 2.11 | Destabilizing | 0.4 | 1.57 | Ambiguous | 1.84 | Ambiguous | 0.78 | Ambiguous | 0.608 | Likely Pathogenic | -5.70 | Deleterious | 0.816 | Possibly Damaging | 0.307 | Benign | 5.51 | Benign | 0.01 | Affected | 0.1671 | 0.6607 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1192C>T | P398S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P398S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include polyPhen‑2 (HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv), and SIFT. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of definitive predictions lean toward a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | -6.757 | Likely Benign | 0.490 | Ambiguous | Likely Benign | 1.86 | Ambiguous | 0.3 | 1.78 | Ambiguous | 1.82 | Ambiguous | 0.85 | Ambiguous | 0.544 | Likely Pathogenic | -5.58 | Deleterious | 0.478 | Possibly Damaging | 0.130 | Benign | 5.68 | Benign | 0.03 | Affected | 0.3619 | 0.5737 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||
| c.1193C>A | P398Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P398Q has no ClinVar record (ClinVar ID None) and is not reported in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM, while the majority of tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default) predict a pathogenic impact. Two tools report uncertainty: ESM1b and AlphaMissense‑Optimized. High‑accuracy assessments further support pathogenicity: the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict, and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. No prediction or stability result is missing or inconclusive. Consequently, the variant is most likely pathogenic based on the collective evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | -7.402 | In-Between | 0.792 | Likely Pathogenic | Ambiguous | 2.13 | Destabilizing | 0.1 | 2.01 | Destabilizing | 2.07 | Destabilizing | 1.01 | Destabilizing | 0.719 | Likely Pathogenic | -5.57 | Deleterious | 0.996 | Probably Damaging | 0.724 | Possibly Damaging | 5.48 | Benign | 0.00 | Affected | 0.1522 | 0.5176 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||
| c.1193C>T | P398L 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant P398L (ClinVar ID 2415189.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33438098‑C‑T). Functional prediction tools that agree on a benign effect include Foldetta, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 HumDiv, and SIFT. Predictions that are uncertain or inconclusive are FoldX, Rosetta, premPS, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Based on the available predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | Uncertain | 1 | 6-33438098-C-T | 8 | 4.96e-6 | -7.518 | In-Between | 0.547 | Ambiguous | Likely Benign | 1.48 | Ambiguous | 0.2 | -0.54 | Ambiguous | 0.47 | Likely Benign | 0.62 | Ambiguous | 0.599 | Likely Pathogenic | -7.10 | Deleterious | 0.961 | Probably Damaging | 0.256 | Benign | 5.72 | Benign | 0.01 | Affected | 3.40 | 16 | 0.2248 | 0.7157 | -3 | -3 | 5.4 | 16.04 | 245.8 | -68.6 | -0.1 | 0.0 | -0.3 | 0.2 | X | Potentially Pathogenic | Pro398 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364 and res. Ala399-Ile411). The Ω loop is assumed to directly interact with the membrane, and it is observed to move arbitrarily throughout the WT solvent simulations. Although the residue swap does not influence the nearby secondary structure elements, proline is often found at the ends of β sheets due to its disfavored status during folding.Additionally, the Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone. Ω loops are known to play significant roles in protein functions that require flexibility, and thus hydrophobic residues like leucine are rarely tolerated. Although no negative structural effects are visualized in the variant’s simulations, Leu398 may exert drastic effects on the SynGAP-membrane complex dynamics and stability. Since the effects on the Gly-rich Ω loop dynamics can only be well studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||
| c.1195G>C | A399P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A399P missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, SIFT, FATHMM, and polyPhen‑2 HumVar; pathogenic predictions come from FoldX, Rosetta, polyPhen‑2 HumDiv, ESM1b, AlphaMissense‑Default, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a pathogenic effect. Overall, the majority of evidence points toward a deleterious impact. The variant is most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.394753 | Structured | 0.407674 | Uncertain | 0.939 | 0.490 | 0.125 | -8.809 | Likely Pathogenic | 0.942 | Likely Pathogenic | Ambiguous | 2.29 | Destabilizing | 0.1 | 4.00 | Destabilizing | 3.15 | Destabilizing | 0.74 | Ambiguous | 0.498 | Likely Benign | -1.82 | Neutral | 0.596 | Possibly Damaging | 0.188 | Benign | 5.56 | Benign | 0.10 | Tolerated | 0.1847 | 0.5472 | 1 | -1 | -3.4 | 26.04 | ||||||||||||||||||||||||||||||
| c.1196C>A | A399D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A399D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that agree on a pathogenic effect are REVEL, FoldX, SIFT, ESM1b, and AlphaMissense‑Default. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized—return uncertain or inconclusive results. High‑accuracy methods give no definitive verdict: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable, and Foldetta is uncertain. Overall, the majority of available predictions (five pathogenic vs. four benign) lean toward pathogenicity. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.394753 | Structured | 0.407674 | Uncertain | 0.939 | 0.490 | 0.125 | -10.441 | Likely Pathogenic | 0.943 | Likely Pathogenic | Ambiguous | 2.45 | Destabilizing | 0.2 | 1.17 | Ambiguous | 1.81 | Ambiguous | 0.91 | Ambiguous | 0.525 | Likely Pathogenic | -1.84 | Neutral | 0.421 | Benign | 0.054 | Benign | 5.38 | Benign | 0.05 | Affected | 0.1533 | 0.1760 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1199T>C | V400A 2D  3DClick to see structure in 3D Viewer AISynGAP1 V400A is not reported in ClinVar and is absent from gnomAD. Consensus from standard predictors shows a split: benign calls from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls come from FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT and AlphaMissense‑Default. Two high‑accuracy tools give a clear signal: AlphaMissense‑Optimized is uncertain, but the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta also predicts pathogenic. With most evidence pointing to deleterious effects, the variant is most likely pathogenic, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.398279 | Structured | 0.415488 | Uncertain | 0.951 | 0.451 | 0.000 | -7.564 | In-Between | 0.871 | Likely Pathogenic | Ambiguous | 3.14 | Destabilizing | 0.1 | 3.12 | Destabilizing | 3.13 | Destabilizing | 2.29 | Destabilizing | 0.479 | Likely Benign | -3.12 | Deleterious | 0.435 | Benign | 0.049 | Benign | 5.32 | Benign | 0.01 | Affected | 0.3291 | 0.2767 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||||||||
| c.1199T>G | V400G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V400G missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect are polyPhen‑2 HumDiv and FATHMM; all other evaluated algorithms (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, is pathogenic. No prediction or stability result is missing or inconclusive. Based on the overwhelming agreement among both general and high‑accuracy tools, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.398279 | Structured | 0.415488 | Uncertain | 0.951 | 0.451 | 0.000 | -11.508 | Likely Pathogenic | 0.969 | Likely Pathogenic | Likely Pathogenic | 4.84 | Destabilizing | 0.1 | 5.40 | Destabilizing | 5.12 | Destabilizing | 2.40 | Destabilizing | 0.819 | Likely Pathogenic | -5.70 | Deleterious | 0.335 | Benign | 0.920 | Probably Damaging | 5.27 | Benign | 0.00 | Affected | 0.2296 | 0.2522 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1208A>G | K403R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K403R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. The remaining tools—FoldX, ESM1b, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a tie, and Foldetta also predicts benign. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.275179 | Structured | 0.424920 | Uncertain | 0.960 | 0.372 | 0.000 | -7.362 | In-Between | 0.355 | Ambiguous | Likely Benign | -0.78 | Ambiguous | 0.1 | 0.23 | Likely Benign | -0.28 | Likely Benign | 0.41 | Likely Benign | 0.335 | Likely Benign | -2.56 | Deleterious | 0.983 | Probably Damaging | 0.926 | Probably Damaging | 3.91 | Benign | 0.15 | Tolerated | 0.4834 | 0.1861 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.1211C>T | A404V 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A404V missense variant is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438116‑C‑T). Prediction tools that agree on a benign effect include REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and ESM1b. Four tools (FoldX, Rosetta, AlphaMissense‑Default, Foldetta) give uncertain or inconclusive results. High‑accuracy methods give mixed evidence: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta remains uncertain. Overall, the majority of predictions lean toward a benign impact, though a key consensus method suggests pathogenicity, leaving the variant’s effect unresolved. **The variant is most likely benign based on the prevailing predictions, and this assessment does not contradict ClinVar status, as no ClinVar classification exists.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.232838 | Structured | 0.415505 | Uncertain | 0.965 | 0.355 | 0.000 | 6-33438116-C-T | 1 | 6.20e-7 | -8.219 | Likely Pathogenic | 0.343 | Ambiguous | Likely Benign | 0.56 | Ambiguous | 0.3 | -1.55 | Ambiguous | -0.50 | Ambiguous | -0.05 | Likely Benign | 0.118 | Likely Benign | -2.96 | Deleterious | 0.345 | Benign | 0.018 | Benign | 4.20 | Benign | 0.47 | Tolerated | 3.38 | 28 | 0.1288 | 0.6842 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||
| c.1219C>G | Q407E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q407E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b all predict a pathogenic outcome. AlphaMissense‑Default, FoldX, Rosetta, and Foldetta are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of tools and the SGM Consensus support a pathogenic classification, while a minority predict benign. No ClinVar entry exists to contradict these predictions. Thus, the variant is most likely pathogenic based on the available computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.109221 | Structured | 0.382522 | Uncertain | 0.916 | 0.271 | 0.000 | -12.631 | Likely Pathogenic | 0.466 | Ambiguous | Likely Benign | 0.50 | Ambiguous | 0.1 | 1.68 | Ambiguous | 1.09 | Ambiguous | 1.30 | Destabilizing | 0.243 | Likely Benign | -2.66 | Deleterious | 0.989 | Probably Damaging | 0.930 | Probably Damaging | 3.96 | Benign | 0.03 | Affected | 0.1199 | 0.2000 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1220A>T | Q407L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q407L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Four tools are uncertain (AlphaMissense‑Default, FoldX, Rosetta, Foldetta). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, more tools predict pathogenicity than benign, and the high‑accuracy consensus leans pathogenic. Therefore, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.109221 | Structured | 0.382522 | Uncertain | 0.916 | 0.271 | 0.000 | -12.730 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | -0.65 | Ambiguous | 0.2 | -0.69 | Ambiguous | -0.67 | Ambiguous | 0.35 | Likely Benign | 0.359 | Likely Benign | -6.32 | Deleterious | 0.939 | Possibly Damaging | 0.838 | Possibly Damaging | 3.91 | Benign | 0.02 | Affected | 0.0585 | 0.4977 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||
| c.1222A>C | T408P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T408P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. FoldX, premPS, and Foldetta are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable; Foldetta remains uncertain. Overall, the majority of available predictions (six pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.161087 | Structured | 0.370935 | Uncertain | 0.907 | 0.239 | 0.000 | -10.384 | Likely Pathogenic | 0.230 | Likely Benign | Likely Benign | 1.08 | Ambiguous | 0.3 | 2.27 | Destabilizing | 1.68 | Ambiguous | 0.73 | Ambiguous | 0.323 | Likely Benign | -4.19 | Deleterious | 0.998 | Probably Damaging | 0.963 | Probably Damaging | 4.07 | Benign | 0.05 | Affected | 0.1985 | 0.5779 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.1222A>G | T408A 2D  AISynGAP1 missense variant T408A is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 (HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv), and ESM1b. The high‑accuracy AlphaMissense‑Optimized score is benign, while the SGM consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Foldetta, a protein‑folding stability method, also predicts benign. Overall, the balance of evidence favors a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.161087 | Structured | 0.370935 | Uncertain | 0.907 | 0.239 | 0.000 | Uncertain | 1 | -8.304 | Likely Pathogenic | 0.114 | Likely Benign | Likely Benign | 0.37 | Likely Benign | 0.6 | -0.06 | Likely Benign | 0.16 | Likely Benign | 0.72 | Ambiguous | 0.118 | Likely Benign | -3.07 | Deleterious | 0.540 | Possibly Damaging | 0.131 | Benign | 4.16 | Benign | 0.14 | Tolerated | 0.3970 | 0.4674 | 1 | 0 | 2.5 | -30.03 | ||||||||||||||||||||||||||||
| c.1223C>T | T408I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 T408I missense variant is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438128‑C‑T). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. Tools with uncertain or mixed outputs are AlphaMissense‑Default and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of evidence points toward a benign effect, and this conclusion does not contradict ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.161087 | Structured | 0.370935 | Uncertain | 0.907 | 0.239 | 0.000 | 6-33438128-C-T | 1 | 6.19e-7 | -10.023 | Likely Pathogenic | 0.542 | Ambiguous | Likely Benign | -0.16 | Likely Benign | 0.1 | -0.67 | Ambiguous | -0.42 | Likely Benign | 0.38 | Likely Benign | 0.131 | Likely Benign | -4.53 | Deleterious | 0.976 | Probably Damaging | 0.607 | Possibly Damaging | 4.08 | Benign | 0.05 | Affected | 3.38 | 28 | 0.0905 | 0.6700 | -1 | 0 | 5.2 | 12.05 | |||||||||||||||||||||||||
| c.1228A>C | S410R 2D  3DClick to see structure in 3D Viewer AIClinVar contains no record for the SynGAP1 S410R variant, and it is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, Foldetta predicts a benign impact, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Taken together, the majority of evidence points to a benign outcome. This conclusion does not contradict ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -8.203 | Likely Pathogenic | 0.941 | Likely Pathogenic | Ambiguous | -0.67 | Ambiguous | 0.2 | 0.56 | Ambiguous | -0.06 | Likely Benign | 0.62 | Ambiguous | 0.242 | Likely Benign | -2.47 | Neutral | 0.871 | Possibly Damaging | 0.298 | Benign | 4.20 | Benign | 0.40 | Tolerated | 0.0955 | 0.3927 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||
| c.1228A>G | S410G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S410G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and polyPhen‑2 HumDiv. The remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -7.147 | In-Between | 0.137 | Likely Benign | Likely Benign | 0.58 | Ambiguous | 0.1 | 1.33 | Ambiguous | 0.96 | Ambiguous | 0.85 | Ambiguous | 0.117 | Likely Benign | -2.54 | Deleterious | 0.952 | Possibly Damaging | 0.145 | Benign | 4.13 | Benign | 0.19 | Tolerated | 0.2651 | 0.5143 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||
| c.1228A>T | S410C 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S410C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict it as pathogenic are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or folding stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -7.552 | In-Between | 0.144 | Likely Benign | Likely Benign | -0.24 | Likely Benign | 0.1 | 0.31 | Likely Benign | 0.04 | Likely Benign | 0.22 | Likely Benign | 0.230 | Likely Benign | -3.10 | Deleterious | 0.993 | Probably Damaging | 0.536 | Possibly Damaging | 4.12 | Benign | 0.11 | Tolerated | 0.1041 | 0.6543 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||
| c.1229G>T | S410I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S410I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Predictions that are uncertain or inconclusive are FoldX, Foldetta, and AlphaMissense‑Default. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized classifies the variant as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans pathogenic, and Foldetta remains uncertain. Overall, the majority of tools suggest a benign impact, but the high‑accuracy consensus indicates potential pathogenicity, leaving the variant’s effect ambiguous. Based on the aggregate predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -9.383 | Likely Pathogenic | 0.432 | Ambiguous | Likely Benign | -1.08 | Ambiguous | 0.2 | -0.36 | Likely Benign | -0.72 | Ambiguous | -0.41 | Likely Benign | 0.114 | Likely Benign | -3.21 | Deleterious | 0.993 | Probably Damaging | 0.589 | Possibly Damaging | 4.15 | Benign | 0.21 | Tolerated | 0.0965 | 0.6579 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||
| c.1230C>A | S410R 2D 3DClick to see structure in 3D Viewer AIClinVar contains no record for the SynGAP1 S410R variant, and it is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, Foldetta predicts a benign impact, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Taken together, the majority of evidence points to a benign outcome. This conclusion does not contradict ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -8.203 | Likely Pathogenic | 0.941 | Likely Pathogenic | Ambiguous | -0.67 | Ambiguous | 0.2 | 0.56 | Ambiguous | -0.06 | Likely Benign | 0.62 | Ambiguous | 0.148 | Likely Benign | -2.47 | Neutral | 0.871 | Possibly Damaging | 0.298 | Benign | 4.20 | Benign | 0.40 | Tolerated | 0.0955 | 0.3927 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||
| c.1230C>G | S410R 2D 3DClick to see structure in 3D Viewer AIClinVar contains no record for the SynGAP1 S410R variant, and it is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, Foldetta predicts a benign impact, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Taken together, the majority of evidence points to a benign outcome. This conclusion does not contradict ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -8.203 | Likely Pathogenic | 0.941 | Likely Pathogenic | Ambiguous | -0.67 | Ambiguous | 0.2 | 0.56 | Ambiguous | -0.06 | Likely Benign | 0.62 | Ambiguous | 0.146 | Likely Benign | -2.47 | Neutral | 0.871 | Possibly Damaging | 0.298 | Benign | 4.20 | Benign | 0.40 | Tolerated | 0.0955 | 0.3927 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||
| c.1231A>C | I411L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I411L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas a majority (premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. Tools with uncertain or mixed outputs are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, yielding no definitive call; and Foldetta also reports an uncertain stability change. Overall, the majority of standard predictors lean toward pathogenicity, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current computational evidence, though high‑accuracy tools remain inconclusive. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.116183 | Structured | 0.339366 | Uncertain | 0.927 | 0.198 | 0.000 | -10.723 | Likely Pathogenic | 0.819 | Likely Pathogenic | Ambiguous | 1.02 | Ambiguous | 0.3 | 1.89 | Ambiguous | 1.46 | Ambiguous | 1.17 | Destabilizing | 0.285 | Likely Benign | -1.84 | Neutral | 0.908 | Possibly Damaging | 0.943 | Probably Damaging | 3.36 | Benign | 0.01 | Affected | 0.0876 | 0.3539 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1234T>A | L412M 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L412M has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and FATHMM, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Five tools give uncertain or inconclusive results (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized). High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑vs‑2 tie; and Foldetta is uncertain. Consequently, the overall evidence leans toward a pathogenic effect, with no ClinVar record to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.122885 | Structured | 0.331108 | Uncertain | 0.937 | 0.196 | 0.000 | -9.342 | Likely Pathogenic | 0.944 | Likely Pathogenic | Ambiguous | 0.75 | Ambiguous | 0.0 | 1.99 | Ambiguous | 1.37 | Ambiguous | 0.86 | Ambiguous | 0.246 | Likely Benign | -1.84 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.25 | Benign | 0.05 | Affected | 0.0746 | 0.3482 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.1240A>C | M414L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M414L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, and FATHMM, while polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default predict a pathogenic outcome. Uncertain predictions come from premPS and ESM1b. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign verdict; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. Overall, the preponderance of evidence indicates that M414L is most likely benign, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | -7.980 | In-Between | 0.779 | Likely Pathogenic | Likely Benign | 0.26 | Likely Benign | 0.0 | 0.33 | Likely Benign | 0.30 | Likely Benign | 0.64 | Ambiguous | 0.336 | Likely Benign | -2.40 | Neutral | 0.559 | Possibly Damaging | 0.495 | Possibly Damaging | 3.63 | Benign | 0.90 | Tolerated | 0.1323 | 0.4236 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.1240A>G | M414V 2D  AISynGAP1 M414V is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized; pathogenic calls come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and ESM1b; the remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) are inconclusive. The SGM consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a pathogenic majority. High‑accuracy assessments give AlphaMissense‑Optimized benign, SGM consensus pathogenic, and Foldetta uncertain. Because the high‑accuracy predictions are divided and the overall tool set is evenly split, there is no definitive evidence for pathogenicity or benignity. Thus, the variant is most likely inconclusive, and this lack of consensus does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | Uncertain | 1 | -8.003 | Likely Pathogenic | 0.541 | Ambiguous | Likely Benign | 1.81 | Ambiguous | 0.4 | 1.73 | Ambiguous | 1.77 | Ambiguous | 0.95 | Ambiguous | 0.261 | Likely Benign | -2.95 | Deleterious | 0.999 | Probably Damaging | 0.987 | Probably Damaging | 3.43 | Benign | 0.24 | Tolerated | 0.2585 | 0.3482 | 2 | 1 | 2.3 | -32.06 | ||||||||||||||||||||||||||||
| c.1240A>T | M414L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M414L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, and FATHMM, while polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default predict a pathogenic outcome. Uncertain predictions come from premPS and ESM1b. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign verdict; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. Overall, the preponderance of evidence indicates that M414L is most likely benign, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | -7.980 | In-Between | 0.779 | Likely Pathogenic | Likely Benign | 0.26 | Likely Benign | 0.0 | 0.33 | Likely Benign | 0.30 | Likely Benign | 0.64 | Ambiguous | 0.336 | Likely Benign | -2.40 | Neutral | 0.559 | Possibly Damaging | 0.495 | Possibly Damaging | 3.63 | Benign | 0.90 | Tolerated | 0.1323 | 0.4236 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.1242G>A | M414I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 M414I missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain or inconclusive results come from FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, more tools (five) predict pathogenicity than benign (four), and no ClinVar evidence contradicts this assessment. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | -6.203 | Likely Benign | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.83 | Ambiguous | 0.0 | 0.86 | Ambiguous | 0.85 | Ambiguous | 0.84 | Ambiguous | 0.265 | Likely Benign | -3.00 | Deleterious | 0.870 | Possibly Damaging | 0.801 | Possibly Damaging | 3.44 | Benign | 0.36 | Tolerated | 0.1168 | 0.3161 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1242G>C | M414I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M414I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta is uncertain, so these results are treated as unavailable. Overall, the majority of evaluated tools (5 pathogenic vs 4 benign) and the single high‑accuracy pathogenic prediction support a likely pathogenic classification. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | -6.203 | Likely Benign | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.83 | Ambiguous | 0.0 | 0.86 | Ambiguous | 0.85 | Ambiguous | 0.84 | Ambiguous | 0.265 | Likely Benign | -3.00 | Deleterious | 0.870 | Possibly Damaging | 0.801 | Possibly Damaging | 3.44 | Benign | 0.36 | Tolerated | 0.1168 | 0.3161 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1242G>T | M414I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M414I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta is uncertain, so these results are treated as unavailable. Overall, the majority of evaluated tools (5 pathogenic vs 4 benign) and the single high‑accuracy pathogenic prediction support a likely pathogenic classification. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | -6.203 | Likely Benign | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.83 | Ambiguous | 0.0 | 0.86 | Ambiguous | 0.85 | Ambiguous | 0.84 | Ambiguous | 0.265 | Likely Benign | -3.00 | Deleterious | 0.870 | Possibly Damaging | 0.801 | Possibly Damaging | 3.44 | Benign | 0.36 | Tolerated | 0.1168 | 0.3161 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1245G>C | E415D 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 E415D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. Predictions that are inconclusive or uncertain are FoldX, ESM1b, and AlphaMissense‑Optimized. High‑accuracy assessments show that the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenicity, while Foldetta predicts a benign effect. Overall, the balance of evidence leans toward a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.100716 | Structured | 0.330366 | Uncertain | 0.915 | 0.236 | 0.000 | -7.766 | In-Between | 0.952 | Likely Pathogenic | Ambiguous | 0.75 | Ambiguous | 0.1 | 0.21 | Likely Benign | 0.48 | Likely Benign | 0.49 | Likely Benign | 0.220 | Likely Benign | -2.60 | Deleterious | 0.995 | Probably Damaging | 0.960 | Probably Damaging | 3.22 | Benign | 0.04 | Affected | 0.1777 | 0.2075 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1245G>T | E415D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E415D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. Predictions that are uncertain or inconclusive are FoldX, ESM1b, and AlphaMissense‑Optimized. High‑accuracy assessments show that AlphaMissense‑Optimized is uncertain, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta indicates a benign effect. Overall, the majority of standard tools lean toward pathogenicity, and the high‑accuracy consensus also supports a pathogenic classification. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.100716 | Structured | 0.330366 | Uncertain | 0.915 | 0.236 | 0.000 | -7.766 | In-Between | 0.952 | Likely Pathogenic | Ambiguous | 0.75 | Ambiguous | 0.1 | 0.21 | Likely Benign | 0.48 | Likely Benign | 0.49 | Likely Benign | 0.220 | Likely Benign | -2.60 | Deleterious | 0.995 | Probably Damaging | 0.960 | Probably Damaging | 3.22 | Benign | 0.04 | Affected | 0.1777 | 0.2075 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1246C>A | L416I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 L416I missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, and FATHMM. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Predictions that remain inconclusive are FoldX, Rosetta, AlphaMissense‑Default, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign prediction. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is uncertain. Overall, the preponderance of evidence points to a benign effect. This conclusion is not contradicted by any ClinVar annotation, as the variant has no existing ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.104810 | Structured | 0.336105 | Uncertain | 0.935 | 0.227 | 0.000 | -8.613 | Likely Pathogenic | 0.396 | Ambiguous | Likely Benign | 0.57 | Ambiguous | 0.0 | 0.69 | Ambiguous | 0.63 | Ambiguous | 0.50 | Likely Benign | 0.127 | Likely Benign | -1.28 | Neutral | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 3.38 | Benign | 0.37 | Tolerated | 0.1073 | 0.3480 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1247T>G | L416R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L416R missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also leans toward benign, while Foldetta remains uncertain. Overall, the majority of reliable predictors classify the variant as benign, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.104810 | Structured | 0.336105 | Uncertain | 0.935 | 0.227 | 0.000 | -8.600 | Likely Pathogenic | 0.511 | Ambiguous | Likely Benign | 0.71 | Ambiguous | 0.0 | 0.97 | Ambiguous | 0.84 | Ambiguous | 0.84 | Ambiguous | 0.238 | Likely Benign | -2.05 | Neutral | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 3.35 | Benign | 0.43 | Tolerated | 0.1445 | 0.0702 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||
| c.1257G>C | E419D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E419D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and FATHMM. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Uncertain or inconclusive predictions come from Foldetta, AlphaMissense‑Optimized, ESM1b, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain (treated as unavailable), the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta remains uncertain (unavailable). Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.371949 | Uncertain | 0.961 | 0.261 | 0.000 | -7.036 | In-Between | 0.869 | Likely Pathogenic | Ambiguous | 0.17 | Likely Benign | 0.1 | 0.87 | Ambiguous | 0.52 | Ambiguous | 0.48 | Likely Benign | 0.170 | Likely Benign | -2.40 | Neutral | 0.995 | Probably Damaging | 0.960 | Probably Damaging | 3.33 | Benign | 0.10 | Tolerated | 0.1966 | 0.4906 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1257G>T | E419D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E419D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and FATHMM. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Uncertain or inconclusive predictions come from Foldetta, AlphaMissense‑Optimized, ESM1b, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain (treated as unavailable), the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta remains uncertain (unavailable). Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.371949 | Uncertain | 0.961 | 0.261 | 0.000 | -7.036 | In-Between | 0.869 | Likely Pathogenic | Ambiguous | 0.17 | Likely Benign | 0.1 | 0.87 | Ambiguous | 0.52 | Ambiguous | 0.48 | Likely Benign | 0.170 | Likely Benign | -2.40 | Neutral | 0.995 | Probably Damaging | 0.960 | Probably Damaging | 3.33 | Benign | 0.10 | Tolerated | 0.1966 | 0.4906 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1264G>C | E422Q 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 E422Q missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, and FATHMM, while those that predict a pathogenic effect are SIFT, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool rates the variant as uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.067594 | Structured | 0.426709 | Uncertain | 0.965 | 0.255 | 0.000 | -9.460 | Likely Pathogenic | 0.926 | Likely Pathogenic | Ambiguous | 0.32 | Likely Benign | 0.0 | 0.21 | Likely Benign | 0.27 | Likely Benign | -0.15 | Likely Benign | 0.208 | Likely Benign | -2.26 | Neutral | 0.997 | Probably Damaging | 0.973 | Probably Damaging | 3.38 | Benign | 0.03 | Affected | 0.1045 | 0.4913 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.1266G>C | E422D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E422D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM; pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors pathogenic; whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts benign. Overall, the majority of tools (six benign vs five pathogenic) lean toward a benign effect, but the two most accurate predictors and the consensus vote indicate a pathogenic tendency. Thus, the variant is most likely benign based on the broader tool set, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.067594 | Structured | 0.426709 | Uncertain | 0.965 | 0.255 | 0.000 | -7.092 | In-Between | 0.969 | Likely Pathogenic | Likely Pathogenic | 0.39 | Likely Benign | 0.0 | -0.03 | Likely Benign | 0.18 | Likely Benign | 0.21 | Likely Benign | 0.199 | Likely Benign | -2.76 | Deleterious | 0.995 | Probably Damaging | 0.960 | Probably Damaging | 3.38 | Benign | 0.07 | Tolerated | 0.1686 | 0.2864 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1266G>T | E422D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E422D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM; pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors pathogenic; whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts benign. Overall, the majority of tools (six benign vs five pathogenic) lean toward a benign effect, but the two most accurate predictors and the consensus vote indicate a pathogenic tendency. Thus, the variant is most likely benign based on the broader tool set, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.067594 | Structured | 0.426709 | Uncertain | 0.965 | 0.255 | 0.000 | -7.092 | In-Between | 0.969 | Likely Pathogenic | Likely Pathogenic | 0.39 | Likely Benign | 0.0 | -0.03 | Likely Benign | 0.18 | Likely Benign | 0.21 | Likely Benign | 0.199 | Likely Benign | -2.76 | Deleterious | 0.995 | Probably Damaging | 0.960 | Probably Damaging | 3.38 | Benign | 0.07 | Tolerated | 0.1686 | 0.2864 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1267T>C | Y423H 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 Y423H missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect comprise FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and AlphaMissense‑Default. AlphaMissense‑Optimized is reported as uncertain. High‑accuracy assessments show that AlphaMissense‑Optimized is inconclusive, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and therefore inconclusive, while Foldetta predicts a pathogenic impact. Overall, the majority of evaluated tools (10 pathogenic vs. 4 benign) support a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.088832 | Structured | 0.421885 | Uncertain | 0.975 | 0.242 | 0.000 | -6.638 | Likely Benign | 0.873 | Likely Pathogenic | Ambiguous | 3.09 | Destabilizing | 0.1 | 2.44 | Destabilizing | 2.77 | Destabilizing | 1.66 | Destabilizing | 0.257 | Likely Benign | -3.88 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.39 | Benign | 0.61 | Tolerated | 0.1603 | 0.0352 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||
| c.1271T>G | V424G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V424G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM. All other evaluated algorithms—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic consensus (3 pathogenic vs. 1 benign); and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, also classifies the variant as pathogenic. No predictions are missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.050641 | Structured | 0.411431 | Uncertain | 0.973 | 0.248 | 0.000 | -13.697 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 3.90 | Destabilizing | 0.1 | 4.84 | Destabilizing | 4.37 | Destabilizing | 2.44 | Destabilizing | 0.622 | Likely Pathogenic | -6.26 | Deleterious | 0.994 | Probably Damaging | 1.000 | Probably Damaging | 3.34 | Benign | 0.00 | Affected | 0.1396 | 0.2029 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1273A>T | T425S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T425S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Four tools (Foldetta, premPS, ESM1b, Rosetta) return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of tools (five benign vs. four pathogenic) lean toward a benign interpretation, and this assessment does not contradict ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.041405 | Structured | 0.401218 | Uncertain | 0.964 | 0.280 | 0.000 | -7.420 | In-Between | 0.676 | Likely Pathogenic | Likely Benign | 0.22 | Likely Benign | 0.1 | 0.77 | Ambiguous | 0.50 | Ambiguous | 0.63 | Ambiguous | 0.275 | Likely Benign | -3.05 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | 3.42 | Benign | 0.07 | Tolerated | 0.2511 | 0.2324 | 1 | 1 | -0.1 | -14.03 | ||||||||||||||||||||||||||||||
| c.1274C>G | T425S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T425S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Four tools (Foldetta, premPS, ESM1b, Rosetta) return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of tools (five benign vs. four pathogenic) lean toward a benign interpretation, and this assessment does not contradict ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.041405 | Structured | 0.401218 | Uncertain | 0.964 | 0.280 | 0.000 | -7.420 | In-Between | 0.676 | Likely Pathogenic | Likely Benign | 0.22 | Likely Benign | 0.1 | 0.77 | Ambiguous | 0.50 | Ambiguous | 0.63 | Ambiguous | 0.184 | Likely Benign | -3.05 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | 3.42 | Benign | 0.07 | Tolerated | 0.2511 | 0.2324 | 1 | 1 | -0.1 | -14.03 | ||||||||||||||||||||||||||||||
| c.1276A>C | N426H 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N426H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Predictions that are uncertain or inconclusive are FoldX and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign; and Foldetta also predicts a benign outcome. No prediction tool is missing or inconclusive in this set. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.394941 | Uncertain | 0.959 | 0.287 | 0.000 | -7.004 | In-Between | 0.248 | Likely Benign | Likely Benign | 0.64 | Ambiguous | 0.0 | -0.14 | Likely Benign | 0.25 | Likely Benign | 0.24 | Likely Benign | 0.237 | Likely Benign | -3.57 | Deleterious | 0.998 | Probably Damaging | 0.985 | Probably Damaging | 3.29 | Benign | 0.14 | Tolerated | 0.1267 | 0.3124 | 2 | 1 | 0.3 | 23.04 | ||||||||||||||||||||||||||||||
| c.1276A>G | N426D 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N426D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Three tools (FoldX, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions (six benign vs. four pathogenic) and the two high‑accuracy benign calls suggest the variant is most likely benign, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.394941 | Uncertain | 0.959 | 0.287 | 0.000 | -11.159 | Likely Pathogenic | 0.554 | Ambiguous | Likely Benign | 0.80 | Ambiguous | 0.0 | 0.18 | Likely Benign | 0.49 | Likely Benign | 0.64 | Ambiguous | 0.173 | Likely Benign | -3.09 | Deleterious | 0.998 | Probably Damaging | 0.980 | Probably Damaging | 3.34 | Benign | 0.09 | Tolerated | 0.1730 | 0.1746 | 2 | 1 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1276A>T | N426Y 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N426Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two main groups: benign predictions come from REVEL, FoldX, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and ESM1b. Two tools remain uncertain: Rosetta and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on the current predictive landscape. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.394941 | Uncertain | 0.959 | 0.287 | 0.000 | -8.510 | Likely Pathogenic | 0.541 | Ambiguous | Likely Benign | 0.47 | Likely Benign | 0.0 | -0.50 | Ambiguous | -0.02 | Likely Benign | 0.23 | Likely Benign | 0.341 | Likely Benign | -5.48 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 3.30 | Benign | 0.25 | Tolerated | 0.0578 | 0.3338 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||
| c.1277A>C | N426T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense change N426T lies in the GAP domain. It is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Predictions that are uncertain or inconclusive are FoldX, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, while Foldetta remains uncertain and is not taken as evidence. Overall, the majority of tools, including the high‑accuracy methods, predict a benign effect. This consensus does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.394941 | Uncertain | 0.959 | 0.287 | 0.000 | -7.389 | In-Between | 0.194 | Likely Benign | Likely Benign | 0.84 | Ambiguous | 0.0 | 0.38 | Likely Benign | 0.61 | Ambiguous | 0.12 | Likely Benign | 0.126 | Likely Benign | -3.14 | Deleterious | 0.983 | Probably Damaging | 0.926 | Probably Damaging | 3.47 | Benign | 0.10 | Tolerated | 0.1090 | 0.3002 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||
| c.1280A>G | H427R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H427R is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438185‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT, while premPS and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign stability change. No predictions or folding results are missing or inconclusive. Overall, the majority of evidence indicates the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.394261 | Uncertain | 0.962 | 0.287 | 0.000 | 6-33438185-A-G | -3.259 | Likely Benign | 0.439 | Ambiguous | Likely Benign | -0.04 | Likely Benign | 0.1 | 0.48 | Likely Benign | 0.22 | Likely Benign | 0.72 | Ambiguous | 0.168 | Likely Benign | -2.61 | Deleterious | 0.174 | Benign | 0.018 | Benign | 3.51 | Benign | 0.03 | Affected | 3.38 | 25 | 0.2108 | 0.2121 | 0 | 2 | -1.3 | 19.05 | |||||||||||||||||||||||||||
| c.1283A>T | Y428F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y428F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b) predict a pathogenic impact. Uncertain or inconclusive results are reported for FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as unavailable. Overall, the balance of evidence—five pathogenic versus three benign predictions, with a pathogenic SGM Consensus and a benign AlphaMissense‑Optimized—suggests that the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.118441 | Structured | 0.389652 | Uncertain | 0.965 | 0.292 | 0.000 | -10.464 | Likely Pathogenic | 0.530 | Ambiguous | Likely Benign | 0.82 | Ambiguous | 0.1 | 0.52 | Ambiguous | 0.67 | Ambiguous | 0.89 | Ambiguous | 0.366 | Likely Benign | -3.82 | Deleterious | 0.999 | Probably Damaging | 0.985 | Probably Damaging | 3.41 | Benign | 0.04 | Affected | 0.2719 | 0.3146 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||
| c.1285C>G | R429G 2D  3DClick to see structure in 3D Viewer AISynGAP1 R429G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta’s stability analysis is uncertain. No evidence from FoldX or Rosetta is available. Overall, the predictions are evenly split, with no clear consensus. The variant is most likely of uncertain significance; it is not contradicted by ClinVar status, which has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.390504 | Uncertain | 0.959 | 0.290 | 0.000 | -9.157 | Likely Pathogenic | 0.333 | Likely Benign | Likely Benign | 1.58 | Ambiguous | 0.0 | 1.68 | Ambiguous | 1.63 | Ambiguous | 1.21 | Destabilizing | 0.257 | Likely Benign | -3.37 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.43 | Benign | 0.34 | Tolerated | 0.2888 | 0.2717 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||
| c.1285C>T | R429W 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R429W is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33438190‑C‑T). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; premPS and AlphaMissense‑Default are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions lean toward a benign impact, and this does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.390504 | Uncertain | 0.959 | 0.290 | 0.000 | Conflicting | 5 | 6-33438190-C-T | 65 | 4.03e-5 | -10.666 | Likely Pathogenic | 0.500 | Ambiguous | Likely Benign | 0.31 | Likely Benign | 0.1 | -0.13 | Likely Benign | 0.09 | Likely Benign | 0.52 | Ambiguous | 0.282 | Likely Benign | -3.19 | Deleterious | 1.000 | Probably Damaging | 0.990 | Probably Damaging | 3.41 | Benign | 0.03 | Affected | 3.38 | 25 | 0.1232 | 0.3422 | 2 | -3 | 3.6 | 30.03 | 252.3 | 45.5 | 0.0 | 0.0 | 0.2 | 0.1 | X | Potentially Pathogenic | The guanidinium group of Arg429, located in an α helix (res. Met414-Glu436), either forms a salt bridge with the carboxylate group of an acidic residue (Asp474, Asp467) or a H-bond with the hydroxyl group of Ser471 in an opposing α helix (res. Ala461-Phe476). In the variant simulations, the indole ring of the Trp429 side chain cannot form ionic interactions with the acidic residues. Although it forms a H-bond with Ser471, the bonding is not as strong as that of arginine. The residue swap could affect the tertiary structure assembly during folding; however, no large-scale negative effects were seen during the simulations. | ||||||||||||||
| c.1286G>C | R429P 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R429P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and ESM1b; AlphaMissense‑Default and FoldX are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of tools and the high‑accuracy methods favor a pathogenic interpretation. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar assertion exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.390504 | Uncertain | 0.959 | 0.290 | 0.000 | -9.771 | Likely Pathogenic | 0.556 | Ambiguous | Likely Benign | 1.53 | Ambiguous | 0.2 | 5.13 | Destabilizing | 3.33 | Destabilizing | 1.01 | Destabilizing | 0.265 | Likely Benign | -2.89 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.43 | Benign | 0.25 | Tolerated | 0.1881 | 0.3790 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||
| c.1289T>A | M430K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M430K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and ESM1b. The remaining tools—FoldX, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized classifies the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors a pathogenic outcome; Foldetta remains uncertain. Overall, the majority of individual predictors lean toward a benign interpretation, whereas the SGM Consensus suggests pathogenicity. Given the lack of ClinVar evidence, there is no contradiction with existing clinical annotations. Based on the aggregate predictions, the variant is most likely benign, and this assessment does not conflict with the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.071867 | Structured | 0.385298 | Uncertain | 0.952 | 0.306 | 0.000 | -10.816 | Likely Pathogenic | 0.494 | Ambiguous | Likely Benign | 0.78 | Ambiguous | 0.1 | 0.44 | Likely Benign | 0.61 | Ambiguous | 0.86 | Ambiguous | 0.149 | Likely Benign | -2.66 | Deleterious | 0.134 | Benign | 0.033 | Benign | 3.47 | Benign | 0.32 | Tolerated | 0.1660 | 0.1271 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||
| c.1289T>G | M430R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M430R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and ESM1b. The remaining tools—FoldX, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of consensus tools lean toward a benign classification, while a subset of high‑accuracy methods suggest pathogenicity, leaving the variant’s impact ambiguous. Based on the aggregate predictions, the variant is most likely benign, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.071867 | Structured | 0.385298 | Uncertain | 0.952 | 0.306 | 0.000 | -10.636 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | 0.98 | Ambiguous | 0.1 | 0.47 | Likely Benign | 0.73 | Ambiguous | 0.83 | Ambiguous | 0.153 | Likely Benign | -2.87 | Deleterious | 0.620 | Possibly Damaging | 0.046 | Benign | 3.48 | Benign | 0.38 | Tolerated | 0.1775 | 0.1652 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||
| c.1291C>A | L431M 2D AIThe SynGAP1 missense variant L431M (ClinVar ID 4327026) is not found in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, FATHMM, AlphaMissense‑Optimized, and Foldetta, whereas premPS, polyPhen‑2 (HumDiv and HumVar), and SIFT all predict a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts benign. No predictions or stability results are missing or inconclusive. Overall, the majority of computational evidence indicates that the variant is most likely benign, which contradicts the ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.094817 | Structured | 0.374755 | Uncertain | 0.959 | 0.300 | 0.000 | 1 | -7.471 | In-Between | 0.375 | Ambiguous | Likely Benign | 0.18 | Likely Benign | 0.3 | 0.50 | Ambiguous | 0.34 | Likely Benign | 1.07 | Destabilizing | 0.102 | Likely Benign | -1.50 | Neutral | 0.995 | Probably Damaging | 0.849 | Possibly Damaging | 2.94 | Benign | 0.02 | Affected | 0.0748 | 0.3121 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||
| c.1291C>G | L431V 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 L431V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, AlphaMissense‑Optimized, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are FoldX, premPS, PROVEAN, and polyPhen‑2 HumDiv. Four tools (Rosetta, Foldetta, ESM1b, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 1‑to‑1 split between benign and pathogenic signals, and Foldetta also yields an uncertain outcome. Overall, the balance of evidence—including the higher number of benign predictions and the benign call from the most accurate tool—suggests that the variant is most likely benign. This conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.094817 | Structured | 0.374755 | Uncertain | 0.959 | 0.300 | 0.000 | -7.949 | In-Between | 0.505 | Ambiguous | Likely Benign | 2.17 | Destabilizing | 0.0 | 1.50 | Ambiguous | 1.84 | Ambiguous | 1.32 | Destabilizing | 0.093 | Likely Benign | -2.58 | Deleterious | 0.861 | Possibly Damaging | 0.332 | Benign | 3.04 | Benign | 0.20 | Tolerated | 0.1377 | 0.3198 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||
| c.1298C>A | A433E 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A433E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools (premPS and AlphaMissense‑Default) give uncertain results. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign prediction; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports benign. No prediction is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.352258 | Uncertain | 0.938 | 0.302 | 0.000 | -8.210 | Likely Pathogenic | 0.506 | Ambiguous | Likely Benign | -0.20 | Likely Benign | 0.0 | 0.15 | Likely Benign | -0.03 | Likely Benign | 0.58 | Ambiguous | 0.148 | Likely Benign | -1.65 | Neutral | 0.998 | Probably Damaging | 0.824 | Possibly Damaging | 3.42 | Benign | 0.18 | Tolerated | 0.0783 | 0.1695 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||
| c.1298C>T | A433V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A433V missense variant is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438203‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive and therefore unavailable as evidence. Overall, the majority of predictions support a benign impact, and this is consistent with the lack of ClinVar pathogenic classification. Thus, the variant is most likely benign based on current computational evidence, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.352258 | Uncertain | 0.938 | 0.302 | 0.000 | 6-33438203-C-T | 240 | 1.49e-4 | -8.200 | Likely Pathogenic | 0.129 | Likely Benign | Likely Benign | 0.48 | Likely Benign | 0.1 | -0.07 | Likely Benign | 0.21 | Likely Benign | 0.48 | Likely Benign | 0.096 | Likely Benign | -2.91 | Deleterious | 0.994 | Probably Damaging | 0.527 | Possibly Damaging | 3.43 | Benign | 0.04 | Affected | 3.37 | 29 | 0.0753 | 0.5104 | 0 | 0 | 2.4 | 28.05 | 214.5 | -45.8 | 0.0 | 0.0 | 0.0 | 0.2 | X | Potentially Benign | The methyl group of Ala433, located on the outer surface of an α helix (res. Met414-Glu436), does not interact with any nearby residues in the WT simulations. In the variant simulations, the iso-propyl side chain of Val433, which has a similarly hydrophobic profile as alanine, also does not form any lasting interactions at the helix surface. Accordingly, the residue swap does not negatively affect the protein structure based on the simulations. | ||||||||||||||||
| c.1300G>C | V434L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V434L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools, Rosetta and AlphaMissense‑Default, give uncertain results. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta, a protein‑folding stability method, also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the preponderance of evidence indicates that V434L is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.158265 | Structured | 0.342846 | Uncertain | 0.954 | 0.306 | 0.000 | -8.697 | Likely Pathogenic | 0.477 | Ambiguous | Likely Benign | -0.05 | Likely Benign | 0.0 | 1.02 | Ambiguous | 0.49 | Likely Benign | 0.20 | Likely Benign | 0.225 | Likely Benign | -2.30 | Neutral | 0.947 | Possibly Damaging | 0.851 | Possibly Damaging | 3.64 | Benign | 0.27 | Tolerated | 0.0895 | 0.3813 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1301T>C | V434A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V434A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and ESM1b; FoldX and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. Overall, the majority of evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.158265 | Structured | 0.342846 | Uncertain | 0.954 | 0.306 | 0.000 | -8.531 | Likely Pathogenic | 0.441 | Ambiguous | Likely Benign | 1.72 | Ambiguous | 0.0 | 2.51 | Destabilizing | 2.12 | Destabilizing | 1.00 | Destabilizing | 0.238 | Likely Benign | -2.51 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.58 | Benign | 0.75 | Tolerated | 0.2338 | 0.2292 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||||||||
| c.1301T>G | V434G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V434G has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect are SIFT and FATHMM, while the remaining tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default) all predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as pathogenic. Taken together, the overwhelming majority of evidence indicates a deleterious effect. Therefore, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.158265 | Structured | 0.342846 | Uncertain | 0.954 | 0.306 | 0.000 | -12.519 | Likely Pathogenic | 0.809 | Likely Pathogenic | Ambiguous | 3.08 | Destabilizing | 0.0 | 4.37 | Destabilizing | 3.73 | Destabilizing | 1.91 | Destabilizing | 0.564 | Likely Pathogenic | -5.16 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.39 | Benign | 0.07 | Tolerated | 0.1758 | 0.2408 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1303T>G | L435V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L435V has no ClinVar assertion and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and AlphaMissense‑Default. ESM1b is uncertain. High‑accuracy assessments further support a pathogenic bias: AlphaMissense‑Optimized predicts benign, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (benign), and PROVEAN (pathogenic)—favors pathogenicity. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenic. With the preponderance of pathogenic calls from both general and high‑accuracy tools, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.229226 | Structured | 0.333584 | Uncertain | 0.954 | 0.292 | 0.000 | -7.134 | In-Between | 0.571 | Likely Pathogenic | Likely Benign | 2.97 | Destabilizing | 0.1 | 2.33 | Destabilizing | 2.65 | Destabilizing | 1.36 | Destabilizing | 0.217 | Likely Benign | -2.80 | Deleterious | 0.995 | Probably Damaging | 0.970 | Probably Damaging | 3.23 | Benign | 0.06 | Tolerated | 0.1216 | 0.2628 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||
| c.1309C>A | P437T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P437T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The remaining tools—FoldX, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta yielding an uncertain stability change. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.306196 | Uncertain | 0.921 | 0.298 | 0.000 | -13.011 | Likely Pathogenic | 0.484 | Ambiguous | Likely Benign | 1.35 | Ambiguous | 0.0 | -3.46 | Stabilizing | -1.06 | Ambiguous | 0.54 | Ambiguous | 0.305 | Likely Benign | -6.67 | Deleterious | 0.999 | Probably Damaging | 0.985 | Probably Damaging | 3.45 | Benign | 0.01 | Affected | 0.1659 | 0.5782 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1309C>G | P437A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P437A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, Rosetta, FATHMM, AlphaMissense‑Optimized, and premPS. Those that predict pathogenicity are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Tools with uncertain or inconclusive results are AlphaMissense‑Default, FoldX, and Foldetta. High‑accuracy assessments give AlphaMissense‑Optimized a benign prediction. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default (uncertain), ESM1b (pathogenic), FATHMM (benign), and PROVEAN (pathogenic), yields a pathogenic consensus. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is uncertain. Overall, the balance of evidence leans toward a pathogenic effect for P437A, and this conclusion does not contradict the current ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.306196 | Uncertain | 0.921 | 0.298 | 0.000 | -12.059 | Likely Pathogenic | 0.392 | Ambiguous | Likely Benign | 1.23 | Ambiguous | 0.0 | -3.14 | Stabilizing | -0.96 | Ambiguous | 0.50 | Likely Benign | 0.266 | Likely Benign | -6.53 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 3.51 | Benign | 0.03 | Affected | 0.3489 | 0.4775 | 1 | -1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||
| c.1309C>T | P437S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P437S is not reported in ClinVar (ClinVar ID: None) but is present in gnomAD (ID 6‑33438214‑C‑T). Prediction tools that agree on a benign effect include REVEL, Rosetta, FATHMM, and AlphaMissense‑Optimized, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b all predict a pathogenic outcome; the remaining methods (FoldX, Foldetta, premPS, AlphaMissense‑Default) are uncertain and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the balance of evidence favors a pathogenic interpretation. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.306196 | Uncertain | 0.921 | 0.298 | 0.000 | 6-33438214-C-T | 2 | 1.24e-6 | -11.964 | Likely Pathogenic | 0.518 | Ambiguous | Likely Benign | 1.14 | Ambiguous | 0.0 | -3.31 | Stabilizing | -1.09 | Ambiguous | 0.60 | Ambiguous | 0.226 | Likely Benign | -6.60 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.49 | Benign | 0.01 | Affected | 3.38 | 26 | 0.3548 | 0.4843 | -1 | 1 | 0.8 | -10.04 | |||||||||||||||||||||||||
| c.130T>A | W44R 2D  AIThe SynGAP1 W44R missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of tools (seven pathogenic vs. three benign) indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this prediction does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.301917 | Structured | 0.431379 | Uncertain | 0.377 | 0.748 | 0.375 | -4.850 | Likely Benign | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.291 | Likely Benign | -5.06 | Deleterious | 0.943 | Possibly Damaging | 0.888 | Possibly Damaging | 3.16 | Benign | 0.00 | Affected | 0.4268 | 0.0749 | 2 | -3 | -3.6 | -30.03 | ||||||||||||||||||||||||||||||||||||||||
| c.130T>C | W44R 2D AISynGAP1 W44R is not reported in ClinVar and is absent from gnomAD. Consensus from standard predictors shows a split: benign calls come from REVEL, ESM1b, and FATHMM, while pathogenic calls come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessment focuses on AlphaMissense‑Optimized, which predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive, and Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic effect, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.301917 | Structured | 0.431379 | Uncertain | 0.377 | 0.748 | 0.375 | -4.850 | Likely Benign | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.291 | Likely Benign | -5.06 | Deleterious | 0.943 | Possibly Damaging | 0.888 | Possibly Damaging | 3.16 | Benign | 0.00 | Affected | 0.4268 | 0.0749 | 2 | -3 | -3.6 | -30.03 | ||||||||||||||||||||||||||||||||||||||||
| c.130T>G | W44G 2D  AIThe SynGAP1 missense variant W44G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, more tools predict pathogenicity (5) than benign (3), and no ClinVar evidence contradicts this assessment. Thus, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.301917 | Structured | 0.431379 | Uncertain | 0.377 | 0.748 | 0.375 | -4.658 | Likely Benign | 0.850 | Likely Pathogenic | Ambiguous | 0.323 | Likely Benign | -4.80 | Deleterious | 0.659 | Possibly Damaging | 0.693 | Possibly Damaging | 3.16 | Benign | 0.00 | Affected | 0.4198 | 0.2164 | -7 | -2 | 0.5 | -129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.1312G>C | A438P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A438P missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas those that predict a pathogenic impact are FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv), SIFT, and AlphaMissense‑Default; premPS and ESM1b are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as pathogenic. Overall, six tools predict pathogenicity versus four predicting benign, and the high‑accuracy consensus is split, but the majority of evidence points toward a deleterious effect. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.147574 | Structured | 0.290154 | Uncertain | 0.929 | 0.293 | 0.000 | -7.955 | In-Between | 0.721 | Likely Pathogenic | Likely Benign | 2.21 | Destabilizing | 0.1 | 6.36 | Destabilizing | 4.29 | Destabilizing | 0.83 | Ambiguous | 0.158 | Likely Benign | -2.46 | Neutral | 0.815 | Possibly Damaging | 0.137 | Benign | 4.09 | Benign | 0.05 | Affected | 0.1624 | 0.4150 | 1 | -1 | -3.4 | 26.04 | ||||||||||||||||||||||||||||||
| c.1313C>A | A438D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A438D missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, polyPhen‑2 HumVar, and FATHMM; pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default; the remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments indicate that AlphaMissense‑Optimized is inconclusive, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta is also inconclusive. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Thus, based on the available predictions, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.147574 | Structured | 0.290154 | Uncertain | 0.929 | 0.293 | 0.000 | -7.410 | In-Between | 0.808 | Likely Pathogenic | Ambiguous | 1.60 | Ambiguous | 0.1 | 0.70 | Ambiguous | 1.15 | Ambiguous | 0.92 | Ambiguous | 0.175 | Likely Benign | -3.07 | Deleterious | 0.859 | Possibly Damaging | 0.124 | Benign | 4.10 | Benign | 0.04 | Affected | 0.1490 | 0.1941 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1318A>G | N440D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N440D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while FoldX, Foldetta, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -9.335 | Likely Pathogenic | 0.407 | Ambiguous | Likely Benign | -0.62 | Ambiguous | 0.0 | -0.41 | Likely Benign | -0.52 | Ambiguous | 0.47 | Likely Benign | 0.074 | Likely Benign | -1.71 | Neutral | 0.229 | Benign | 0.045 | Benign | 3.43 | Benign | 0.43 | Tolerated | 0.1544 | 0.2025 | 2 | 1 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.131G>C | W44S 2D  AIThe SynGAP1 missense variant W44S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.301917 | Structured | 0.431379 | Uncertain | 0.377 | 0.748 | 0.375 | -5.113 | Likely Benign | 0.921 | Likely Pathogenic | Ambiguous | 0.275 | Likely Benign | -4.68 | Deleterious | 0.824 | Possibly Damaging | 0.775 | Possibly Damaging | 3.16 | Benign | 0.00 | Affected | 0.4139 | 0.2371 | -2 | -3 | 0.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.131G>T | W44L 2D  AIThe SynGAP1 missense variant W44L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Consensus from standard predictors shows a split: benign calls come from REVEL, ESM1b, and FATHMM, while pathogenic calls come from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized returns an uncertain result. High‑accuracy assessment is inconclusive: the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑vs‑2 tie, and Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a pathogenic interpretation, with no conflict with ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.301917 | Structured | 0.431379 | Uncertain | 0.377 | 0.748 | 0.375 | -5.743 | Likely Benign | 0.869 | Likely Pathogenic | Ambiguous | 0.211 | Likely Benign | -4.37 | Deleterious | 0.659 | Possibly Damaging | 0.693 | Possibly Damaging | 3.20 | Benign | 0.00 | Affected | 0.2584 | 0.3413 | -2 | -2 | 4.7 | -73.05 | ||||||||||||||||||||||||||||||||||||||||
| c.1320T>A | N440K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N440K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. The remaining methods—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Based on the overall distribution of predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -10.114 | Likely Pathogenic | 0.895 | Likely Pathogenic | Ambiguous | 0.92 | Ambiguous | 0.1 | 1.04 | Ambiguous | 0.98 | Ambiguous | 0.40 | Likely Benign | 0.058 | Likely Benign | -1.97 | Neutral | 0.206 | Benign | 0.021 | Benign | 3.50 | Benign | 0.19 | Tolerated | 0.1660 | 0.2550 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||
| c.1320T>G | N440K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N440K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. The remaining methods—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Based on the overall distribution of predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -10.114 | Likely Pathogenic | 0.895 | Likely Pathogenic | Ambiguous | 0.92 | Ambiguous | 0.1 | 1.04 | Ambiguous | 0.98 | Ambiguous | 0.40 | Likely Benign | 0.057 | Likely Benign | -1.97 | Neutral | 0.206 | Benign | 0.021 | Benign | 3.50 | Benign | 0.19 | Tolerated | 0.1660 | 0.2550 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||
| c.1321G>C | V441L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V441L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while Rosetta and AlphaMissense‑Default are uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts benign. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the preponderance of evidence, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | -9.546 | Likely Pathogenic | 0.395 | Ambiguous | Likely Benign | -0.43 | Likely Benign | 0.0 | 0.59 | Ambiguous | 0.08 | Likely Benign | 0.45 | Likely Benign | 0.135 | Likely Benign | -2.27 | Neutral | 0.165 | Benign | 0.028 | Benign | 3.43 | Benign | 0.11 | Tolerated | 0.0961 | 0.3881 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1322T>C | V441A 2D 3DClick to see structure in 3D Viewer AISynGAP1 variant V441A is listed in ClinVar as uncertain and is present in gnomAD (ID 6‑33438227‑T‑C). Consensus from most in silico predictors favors a benign effect: REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized all report benign. Pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, and ESM1b, while premPS and AlphaMissense‑Default remain uncertain. High‑accuracy assessments give a mixed picture: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports benign. Overall, the preponderance of evidence points to a benign impact, aligning with the ClinVar uncertain designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | Conflicting | 2 | 6-33438227-T-C | 3 | 1.86e-6 | -9.439 | Likely Pathogenic | 0.359 | Ambiguous | Likely Benign | -0.14 | Likely Benign | 0.0 | 0.33 | Likely Benign | 0.10 | Likely Benign | 0.95 | Ambiguous | 0.053 | Likely Benign | -2.92 | Deleterious | 0.513 | Possibly Damaging | 0.214 | Benign | 3.44 | Benign | 0.93 | Tolerated | 3.37 | 29 | 0.2390 | 0.1800 | 0 | 0 | -2.4 | -28.05 | 195.0 | 44.6 | 0.0 | 0.1 | 0.5 | 0.0 | X | X | Uncertain | The iso-propyl side chain of Val441, located on the outer surface of an α helix (res. Asn440-Thr458), does not interact with other residues in the WT simulations. In the variant simulations, the methyl side chain of Ala441 is similarly hydrophobic and does not form any interactions on the outer helix surface. Although the residue swap does not negatively affect the protein structure based on the simulations, it is noteworthy that the residue faces the RasGTPase interface. Thus, the effect of the residue swap on the SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | |||||||||||||
| c.1322T>G | V441G 2D 3DClick to see structure in 3D Viewer AISynGAP1 variant V441G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into three groups: benign predictions come from REVEL, FoldX, SIFT, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b; the remaining tools (Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized remains benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) favors pathogenic, and Foldetta is inconclusive. Taken together, the majority of evidence points to a benign effect, but the SGM Consensus and several individual pathogenic predictors introduce uncertainty. Therefore, the variant is most likely benign based on the overall prediction landscape, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | -12.380 | Likely Pathogenic | 0.478 | Ambiguous | Likely Benign | 0.18 | Likely Benign | 0.0 | 1.25 | Ambiguous | 0.72 | Ambiguous | 0.95 | Ambiguous | 0.273 | Likely Benign | -5.88 | Deleterious | 0.841 | Possibly Damaging | 0.997 | Probably Damaging | 3.41 | Benign | 0.24 | Tolerated | 0.1664 | 0.1715 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1324A>C | K442Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K442Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.170161 | Structured | 0.255766 | Uncertain | 0.912 | 0.225 | 0.000 | -10.410 | Likely Pathogenic | 0.562 | Ambiguous | Likely Benign | 0.05 | Likely Benign | 0.1 | 0.03 | Likely Benign | 0.04 | Likely Benign | 0.25 | Likely Benign | 0.268 | Likely Benign | -3.10 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 3.39 | Benign | 0.18 | Tolerated | 0.3353 | 0.1014 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||
| c.132G>C | W44C 2D  AIThe SynGAP1 missense variant W44C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of evaluated tools (seven pathogenic vs. three benign) indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this prediction does not contradict any ClinVar status because the variant has not yet been classified in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.301917 | Structured | 0.431379 | Uncertain | 0.377 | 0.748 | 0.375 | -6.216 | Likely Benign | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.321 | Likely Benign | -4.70 | Deleterious | 0.943 | Possibly Damaging | 0.941 | Probably Damaging | 3.14 | Benign | 0.00 | Affected | 0.3542 | 0.2370 | -8 | -2 | 3.4 | -83.07 | ||||||||||||||||||||||||||||||||||||||||
| c.132G>T | W44C 2D AIThe SynGAP1 missense variant W44C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated tools (seven pathogenic vs. three benign) indicate a pathogenic effect. This prediction is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.301917 | Structured | 0.431379 | Uncertain | 0.377 | 0.748 | 0.375 | -6.216 | Likely Benign | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.321 | Likely Benign | -4.70 | Deleterious | 0.943 | Possibly Damaging | 0.941 | Probably Damaging | 3.14 | Benign | 0.00 | Affected | 0.3542 | 0.2370 | -8 | -2 | 3.4 | -83.07 | ||||||||||||||||||||||||||||||||||||||||
| c.1340T>G | V447G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V447G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and FATHMM, while the remaining tools—FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. No prediction or stability result is missing or inconclusive. Overall, the preponderance of evidence indicates that V447G is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.250310 | Structured | 0.283801 | Uncertain | 0.970 | 0.243 | 0.000 | -13.648 | Likely Pathogenic | 0.861 | Likely Pathogenic | Ambiguous | 3.81 | Destabilizing | 0.1 | 4.62 | Destabilizing | 4.22 | Destabilizing | 2.28 | Destabilizing | 0.499 | Likely Benign | -5.43 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.31 | Benign | 0.01 | Affected | 0.1863 | 0.2240 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1346G>T | S449I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S449I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen2_HumVar. Those that predict a pathogenic effect are PROVEAN, polyPhen2_HumDiv, and ESM1b. Rosetta and Foldetta give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic). Foldetta also remains uncertain. Overall, the majority of evidence (7 benign vs. 3 pathogenic) points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.254060 | Structured | 0.301437 | Uncertain | 0.958 | 0.251 | 0.000 | -9.549 | Likely Pathogenic | 0.310 | Likely Benign | Likely Benign | 0.04 | Likely Benign | 0.1 | -1.39 | Ambiguous | -0.68 | Ambiguous | 0.13 | Likely Benign | 0.105 | Likely Benign | -3.23 | Deleterious | 0.559 | Possibly Damaging | 0.044 | Benign | 3.40 | Benign | 0.14 | Tolerated | 0.0756 | 0.5006 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||
| c.1348G>A | A450T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A450T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Because the majority of conventional tools lean toward benign and no ClinVar evidence contradicts this, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.321458 | Structured | 0.306281 | Uncertain | 0.963 | 0.234 | 0.000 | -9.149 | Likely Pathogenic | 0.380 | Ambiguous | Likely Benign | 0.50 | Ambiguous | 0.2 | 0.98 | Ambiguous | 0.74 | Ambiguous | 0.81 | Ambiguous | 0.233 | Likely Benign | -3.35 | Deleterious | 0.996 | Probably Damaging | 0.973 | Probably Damaging | 3.40 | Benign | 0.06 | Tolerated | 0.0943 | 0.5902 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||
| c.1348G>T | A450S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A450S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The remaining methods—FoldX, Rosetta, Foldetta, and premPS—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and thus unavailable; Foldetta is uncertain. Overall, the balance of evidence (five benign versus four pathogenic predictions, with three uncertain) suggests the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.321458 | Structured | 0.306281 | Uncertain | 0.963 | 0.234 | 0.000 | -9.257 | Likely Pathogenic | 0.274 | Likely Benign | Likely Benign | 0.81 | Ambiguous | 0.0 | 1.35 | Ambiguous | 1.08 | Ambiguous | 0.69 | Ambiguous | 0.268 | Likely Benign | -2.70 | Deleterious | 0.965 | Probably Damaging | 0.972 | Probably Damaging | 3.47 | Benign | 0.10 | Tolerated | 0.2003 | 0.4322 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.1351C>A | L451I 2D AIThe SynGAP1 L451I missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Uncertain results come from FoldX, Rosetta, premPS, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, the majority of available predictions (five pathogenic versus four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict the absence of a ClinVar record. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.281712 | Structured | 0.314017 | Uncertain | 0.978 | 0.232 | 0.000 | -11.046 | Likely Pathogenic | 0.658 | Likely Pathogenic | Likely Benign | 1.43 | Ambiguous | 0.8 | 0.70 | Ambiguous | 1.07 | Ambiguous | 0.94 | Ambiguous | 0.284 | Likely Benign | -1.94 | Neutral | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 2.75 | Benign | 0.02 | Affected | 0.0681 | 0.2958 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1354G>A | V452I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 V452I variant is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Rosetta and AlphaMissense‑Default return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.335645 | Structured | 0.315167 | Uncertain | 0.970 | 0.229 | 0.000 | Uncertain | 2 | -8.985 | Likely Pathogenic | 0.361 | Ambiguous | Likely Benign | -0.08 | Likely Benign | 0.1 | 0.51 | Ambiguous | 0.22 | Likely Benign | 0.25 | Likely Benign | 0.218 | Likely Benign | -0.99 | Neutral | 0.947 | Possibly Damaging | 0.851 | Possibly Damaging | 3.26 | Benign | 0.05 | Affected | 0.0630 | 0.3476 | 4 | 3 | 0.3 | 14.03 | ||||||||||||||||||||||||||||
| c.1355T>G | V452G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V452G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining tools—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—consistently predict a pathogenic impact. AlphaMissense‑Optimized is uncertain, providing no definitive direction. High‑accuracy assessments further support pathogenicity: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenicity. AlphaMissense‑Optimized remains uncertain. Overall, the preponderance of evidence indicates that V452G is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.335645 | Structured | 0.315167 | Uncertain | 0.970 | 0.229 | 0.000 | -13.410 | Likely Pathogenic | 0.946 | Likely Pathogenic | Ambiguous | 3.70 | Destabilizing | 0.1 | 3.37 | Destabilizing | 3.54 | Destabilizing | 2.46 | Destabilizing | 0.570 | Likely Pathogenic | -6.92 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.17 | Benign | 0.00 | Affected | 0.1948 | 0.2567 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1360A>C | I454L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I454L is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, while polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default predict a pathogenic impact. The remaining tools—FoldX, Rosetta, Foldetta, premPS, ESM1b, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward benign, and Foldetta also uncertain. Overall, the majority of reliable predictors and the SGM Consensus favor a benign classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.254060 | Structured | 0.312811 | Uncertain | 0.965 | 0.182 | 0.000 | -7.852 | In-Between | 0.786 | Likely Pathogenic | Ambiguous | 0.57 | Ambiguous | 0.1 | 1.36 | Ambiguous | 0.97 | Ambiguous | 0.80 | Ambiguous | 0.246 | Likely Benign | -1.98 | Neutral | 0.908 | Possibly Damaging | 0.943 | Probably Damaging | 3.51 | Benign | 0.26 | Tolerated | 0.0653 | 0.2857 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1363C>A | L455M 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L455M is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie, and Foldetta is uncertain. No folding‑stability method provides a definitive result. Consequently, the computational evidence does not favor either benign or pathogenic classification. The variant’s status is therefore inconclusive, and this lack of consensus does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.188120 | Structured | 0.310377 | Uncertain | 0.963 | 0.168 | 0.000 | -10.086 | Likely Pathogenic | 0.802 | Likely Pathogenic | Ambiguous | 0.88 | Ambiguous | 0.0 | 1.27 | Ambiguous | 1.08 | Ambiguous | 0.59 | Ambiguous | 0.147 | Likely Benign | -1.64 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.25 | Benign | 0.11 | Tolerated | 0.0606 | 0.3362 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.1366C>G | Q456E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q456E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) yield uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) remaining uncertain. Overall, the predictions are split, but the high‑accuracy consensus leans toward pathogenicity. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.170161 | Structured | 0.302348 | Uncertain | 0.939 | 0.164 | 0.000 | -12.982 | Likely Pathogenic | 0.503 | Ambiguous | Likely Benign | 0.71 | Ambiguous | 0.1 | 0.87 | Ambiguous | 0.79 | Ambiguous | 0.65 | Ambiguous | 0.233 | Likely Benign | -2.76 | Deleterious | 0.998 | Probably Damaging | 0.964 | Probably Damaging | 3.41 | Benign | 0.15 | Tolerated | 0.1191 | 0.1276 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1367A>T | Q456L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q456L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Tools with uncertain or inconclusive results are FoldX, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of predictions (six benign vs. four pathogenic) lean toward a benign interpretation, with no evidence of contradiction with the ClinVar status. Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.170161 | Structured | 0.302348 | Uncertain | 0.939 | 0.164 | 0.000 | -10.272 | Likely Pathogenic | 0.492 | Ambiguous | Likely Benign | -0.88 | Ambiguous | 0.1 | -0.30 | Likely Benign | -0.59 | Ambiguous | 0.28 | Likely Benign | 0.347 | Likely Benign | -6.65 | Deleterious | 0.987 | Probably Damaging | 0.914 | Probably Damaging | 3.49 | Benign | 0.36 | Tolerated | 0.0554 | 0.4317 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||
| c.1368G>C | Q456H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q456H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Tools with uncertain or inconclusive results are FoldX, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of predictions lean toward a benign impact, and this does not contradict the lack of ClinVar annotation. Thus, based on the available computational evidence, the variant is most likely benign, though a high‑accuracy consensus suggests a possible pathogenic effect that warrants further investigation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.170161 | Structured | 0.302348 | Uncertain | 0.939 | 0.164 | 0.000 | -7.993 | In-Between | 0.598 | Likely Pathogenic | Likely Benign | 0.74 | Ambiguous | 0.1 | 0.39 | Likely Benign | 0.57 | Ambiguous | 0.32 | Likely Benign | 0.214 | Likely Benign | -4.07 | Deleterious | 0.996 | Probably Damaging | 0.974 | Probably Damaging | 3.37 | Benign | 0.19 | Tolerated | 0.0987 | 0.2358 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1368G>T | Q456H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q456H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Tools with uncertain or inconclusive results are FoldX, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of predictions lean toward a benign impact, and this does not contradict the lack of ClinVar annotation. Thus, based on the available computational evidence, the variant is most likely benign, though a high‑accuracy consensus suggests a possible pathogenic effect that warrants further investigation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.170161 | Structured | 0.302348 | Uncertain | 0.939 | 0.164 | 0.000 | -7.993 | In-Between | 0.598 | Likely Pathogenic | Likely Benign | 0.74 | Ambiguous | 0.1 | 0.39 | Likely Benign | 0.57 | Ambiguous | 0.32 | Likely Benign | 0.214 | Likely Benign | -4.07 | Deleterious | 0.996 | Probably Damaging | 0.974 | Probably Damaging | 3.37 | Benign | 0.19 | Tolerated | 0.0987 | 0.2358 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1381G>A | A461T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A461T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and ESM1b. Four tools (FoldX, Rosetta, Foldetta, premPS) returned uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta is also inconclusive. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.292531 | Uncertain | 0.936 | 0.151 | 0.125 | -10.885 | Likely Pathogenic | 0.323 | Likely Benign | Likely Benign | 1.21 | Ambiguous | 0.2 | 0.51 | Ambiguous | 0.86 | Ambiguous | 0.65 | Ambiguous | 0.214 | Likely Benign | -3.27 | Deleterious | 0.508 | Possibly Damaging | 0.042 | Benign | 3.40 | Benign | 0.13 | Tolerated | 0.1083 | 0.5930 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||
| c.1381G>T | A461S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A461S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 (HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv), SIFT, and ESM1b. Four tools (FoldX, Rosetta, Foldetta, premPS) give uncertain results and are not considered evidence for either side. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta’s stability analysis is also unavailable. Overall, the balance of evidence leans toward a benign effect, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.292531 | Uncertain | 0.936 | 0.151 | 0.125 | -10.663 | Likely Pathogenic | 0.309 | Likely Benign | Likely Benign | 0.87 | Ambiguous | 0.0 | 1.18 | Ambiguous | 1.03 | Ambiguous | 0.63 | Ambiguous | 0.236 | Likely Benign | -2.74 | Deleterious | 0.600 | Possibly Damaging | 0.289 | Benign | 3.36 | Benign | 0.02 | Affected | 0.2401 | 0.4551 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.1382C>G | A461G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A461G has no ClinVar entry and is not reported in gnomAD. Prediction tools cluster into three groups: benign predictions come from REVEL, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions arise from Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; the remaining tools (FoldX, premPS, AlphaMissense‑Default, Foldetta) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points toward a pathogenic effect, with no ClinVar record to contradict this assessment. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.292531 | Uncertain | 0.936 | 0.151 | 0.125 | -11.360 | Likely Pathogenic | 0.556 | Ambiguous | Likely Benign | 1.46 | Ambiguous | 0.1 | 2.06 | Destabilizing | 1.76 | Ambiguous | 0.83 | Ambiguous | 0.276 | Likely Benign | -3.79 | Deleterious | 0.991 | Probably Damaging | 0.628 | Possibly Damaging | 3.32 | Benign | 0.00 | Affected | 0.1988 | 0.3317 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||
| c.1382C>T | A461V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A461V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that predict a pathogenic effect are PROVEAN, polyPhen2_HumDiv, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Because the majority of standard predictors (nine benign vs. three pathogenic) favor a benign outcome, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.292531 | Uncertain | 0.936 | 0.151 | 0.125 | -9.968 | Likely Pathogenic | 0.436 | Ambiguous | Likely Benign | -0.08 | Likely Benign | 0.3 | 0.34 | Likely Benign | 0.13 | Likely Benign | 0.02 | Likely Benign | 0.141 | Likely Benign | -3.05 | Deleterious | 0.983 | Probably Damaging | 0.273 | Benign | 3.43 | Benign | 0.66 | Tolerated | 0.0819 | 0.5078 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||
| c.1385A>G | K462R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K462R is catalogued in gnomAD (ID 6‑33438290‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Three tools (Rosetta, Foldetta, premPS) yield uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta is uncertain, so neither provides decisive evidence. Overall, the majority of reliable predictors lean toward a benign effect. This consensus does not contradict ClinVar status, which is currently absent. Thus, based on available predictions, K462R is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.264545 | Structured | 0.297737 | Uncertain | 0.921 | 0.159 | 0.125 | 6-33438290-A-G | 1 | 6.20e-7 | -9.980 | Likely Pathogenic | 0.184 | Likely Benign | Likely Benign | 0.13 | Likely Benign | 0.1 | 0.90 | Ambiguous | 0.52 | Ambiguous | 0.84 | Ambiguous | 0.246 | Likely Benign | -2.75 | Deleterious | 0.983 | Probably Damaging | 0.962 | Probably Damaging | 3.43 | Benign | 0.09 | Tolerated | 3.37 | 34 | 0.4995 | 0.1201 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||
| c.1389C>A | D463E 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 D463E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and premPS. Only PROVEAN predicts a pathogenic outcome, while Rosetta and AlphaMissense‑Default are uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.260850 | Structured | 0.305622 | Uncertain | 0.940 | 0.176 | 0.000 | -5.170 | Likely Benign | 0.527 | Ambiguous | Likely Benign | -0.42 | Likely Benign | 0.1 | 0.50 | Ambiguous | 0.04 | Likely Benign | 0.47 | Likely Benign | 0.162 | Likely Benign | -2.58 | Deleterious | 0.012 | Benign | 0.003 | Benign | 3.47 | Benign | 0.29 | Tolerated | 0.1330 | 0.5177 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||
| c.1389C>G | D463E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D463E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and premPS. Only PROVEAN predicts a pathogenic outcome, while Rosetta and AlphaMissense‑Default are uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) resolves to benign, and Foldetta also indicates benign stability. No prediction or stability result is missing or inconclusive. Overall, the variant is most likely benign based on the collective evidence, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.260850 | Structured | 0.305622 | Uncertain | 0.940 | 0.176 | 0.000 | -5.170 | Likely Benign | 0.527 | Ambiguous | Likely Benign | -0.42 | Likely Benign | 0.1 | 0.50 | Ambiguous | 0.04 | Likely Benign | 0.47 | Likely Benign | 0.162 | Likely Benign | -2.58 | Deleterious | 0.012 | Benign | 0.003 | Benign | 3.47 | Benign | 0.29 | Tolerated | 0.1330 | 0.5177 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||
| c.1393C>A | L465I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L465I missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score classifies the variant as benign, whereas the Foldetta stability assessment is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive due to a 2‑to‑2 split. Overall, the evidence is mixed; the balance of predictions leans toward a benign interpretation, and this does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.346032 | Structured | 0.319240 | Uncertain | 0.956 | 0.202 | 0.000 | -9.672 | Likely Pathogenic | 0.770 | Likely Pathogenic | Likely Benign | 1.21 | Ambiguous | 0.1 | 1.27 | Ambiguous | 1.24 | Ambiguous | 0.78 | Ambiguous | 0.258 | Likely Benign | -1.99 | Neutral | 0.998 | Probably Damaging | 0.997 | Probably Damaging | 2.54 | Benign | 0.08 | Tolerated | 0.0967 | 0.3539 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.139C>T | R47W 2D AIThe SynGAP1 missense variant R47W is listed in ClinVar as a variant of uncertain significance and is present in the gnomAD database (ID 6‑33423548‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign votes), and Foldetta results are unavailable. Overall, the majority of conventional tools suggest a pathogenic impact, but the single high‑accuracy benign prediction and the inconclusive consensus temper this view. Consequently, the variant is most likely pathogenic based on the preponderance of evidence, and this assessment does not contradict the ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.318242 | Structured | 0.436559 | Uncertain | 0.520 | 0.719 | 0.125 | Uncertain | 1 | 6-33423548-C-T | 1 | 6.20e-7 | -9.201 | Likely Pathogenic | 0.752 | Likely Pathogenic | Likely Benign | 0.201 | Likely Benign | -2.17 | Neutral | 0.994 | Probably Damaging | 0.919 | Probably Damaging | 4.00 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1101 | 0.4158 | -3 | 2 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||
| c.1402A>G | M468V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M468V is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from PROVEAN, SIFT, and AlphaMissense‑Optimized, while pathogenic predictions are made by REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools, premPS and AlphaMissense‑Default, return uncertain results. High‑accuracy assessments further clarify the variant’s impact: AlphaMissense‑Optimized predicts a benign effect; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates pathogenicity; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also classifies the variant as pathogenic. Overall, the preponderance of evidence points to a pathogenic effect, which does not contradict the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.284882 | Structured | 0.339253 | Uncertain | 0.932 | 0.257 | 0.000 | Uncertain | 1 | -9.461 | Likely Pathogenic | 0.361 | Ambiguous | Likely Benign | 2.69 | Destabilizing | 0.1 | 2.20 | Destabilizing | 2.45 | Destabilizing | 0.89 | Ambiguous | 0.570 | Likely Pathogenic | -1.66 | Neutral | 0.998 | Probably Damaging | 0.993 | Probably Damaging | -1.21 | Pathogenic | 0.08 | Tolerated | 3.37 | 31 | 0.3309 | 0.3845 | 1 | 2 | 2.3 | -32.06 | ||||||||||||||||||||||||||
| c.1406C>G | A469G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A469G is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD. Prediction tools that agree on a benign effect include PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise REVEL, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default; FoldX is uncertain. High‑accuracy methods give mixed results: AlphaMissense‑Optimized indicates benign, Foldetta indicates pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of tools (12 vs. 4) predict pathogenicity, and there is no ClinVar entry to contradict this assessment. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.278302 | Structured | 0.343926 | Uncertain | 0.910 | 0.276 | 0.000 | -4.438 | Likely Benign | 0.733 | Likely Pathogenic | Likely Benign | 1.99 | Ambiguous | 0.1 | 2.22 | Destabilizing | 2.11 | Destabilizing | 1.01 | Destabilizing | 0.569 | Likely Pathogenic | -2.42 | Neutral | 0.997 | Probably Damaging | 0.990 | Probably Damaging | -1.32 | Pathogenic | 0.37 | Tolerated | 0.1797 | 0.3078 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||
| c.1406C>T | A469V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A469V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect comprise FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and Foldetta; premPS and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta predicts pathogenic. With seven tools supporting pathogenicity versus four supporting benignity, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.278302 | Structured | 0.343926 | Uncertain | 0.910 | 0.276 | 0.000 | -8.858 | Likely Pathogenic | 0.459 | Ambiguous | Likely Benign | 2.30 | Destabilizing | 0.4 | 2.66 | Destabilizing | 2.48 | Destabilizing | -0.77 | Ambiguous | 0.426 | Likely Benign | 0.26 | Neutral | 0.997 | Probably Damaging | 0.976 | Probably Damaging | -1.20 | Pathogenic | 0.60 | Tolerated | 0.0827 | 0.5421 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||
| c.140G>C | R47P 2D AIThe SynGAP1 missense variant R47P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic, two benign) and therefore unavailable as evidence. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant and is likewise unavailable. High‑accuracy tools specifically indicate a benign prediction from AlphaMissense‑Optimized, while the other high‑accuracy methods (SGM Consensus, Foldetta) provide no definitive assessment. Overall, the balance of evidence (five pathogenic vs. four benign predictions) suggests the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.318242 | Structured | 0.436559 | Uncertain | 0.520 | 0.719 | 0.125 | -10.316 | Likely Pathogenic | 0.746 | Likely Pathogenic | Likely Benign | 0.195 | Likely Benign | -1.63 | Neutral | 0.841 | Possibly Damaging | 0.809 | Possibly Damaging | 4.02 | Benign | 0.00 | Affected | 0.2050 | 0.4956 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||||||||||
| c.1412C>G | S471C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S471C is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33438444‑C‑G). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of predictions (10 benign vs. 3 pathogenic) indicate that the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no reported pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.305330 | Structured | 0.355411 | Uncertain | 0.888 | 0.261 | 0.000 | 6-33438444-C-G | 1 | 6.20e-7 | -3.454 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.36 | Likely Benign | 0.0 | -0.05 | Likely Benign | 0.16 | Likely Benign | 0.07 | Likely Benign | 0.273 | Likely Benign | -2.90 | Deleterious | 0.000 | Benign | 0.001 | Benign | -1.32 | Pathogenic | 0.01 | Affected | 3.37 | 34 | 0.1048 | 0.4913 | -1 | 0 | 3.3 | 16.06 | |||||||||||||||||||||||||
| c.1417G>A | V473I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V473I is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438449‑G‑A). Functional prediction tools that agree on benign impact include REVEL, FoldX, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Pathogenic predictions are provided by both polyPhen‑2 HumDiv and HumVar. Predictions that are inconclusive are AlphaMissense‑Default, ESM1b, Foldetta, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is unavailable due to no majority, and Foldetta is uncertain. Overall, the balance of evidence favors a benign effect for V473I, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.362529 | Uncertain | 0.884 | 0.239 | 0.000 | Uncertain | 1 | 6-33438449-G-A | 1 | 6.20e-7 | -7.481 | In-Between | 0.418 | Ambiguous | Likely Benign | -0.12 | Likely Benign | 0.0 | 1.20 | Ambiguous | 0.54 | Ambiguous | -0.06 | Likely Benign | 0.203 | Likely Benign | -0.91 | Neutral | 0.929 | Possibly Damaging | 0.917 | Probably Damaging | 3.74 | Benign | 0.18 | Tolerated | 3.37 | 34 | 0.0568 | 0.3335 | 3 | 4 | 0.3 | 14.03 | |||||||||||||||||||||||
| c.1418T>G | V473G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V473G has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include only FATHMM, while the remaining eleven tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) uniformly predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is uncertain, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) votes pathogenic, and Foldetta also predicts pathogenic. No predictions are missing or inconclusive. Consequently, the variant is most likely pathogenic based on the collective evidence, and this conclusion does not contradict any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.362529 | Uncertain | 0.884 | 0.239 | 0.000 | -14.782 | Likely Pathogenic | 0.936 | Likely Pathogenic | Ambiguous | 3.45 | Destabilizing | 0.0 | 3.40 | Destabilizing | 3.43 | Destabilizing | 2.69 | Destabilizing | 0.683 | Likely Pathogenic | -6.91 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.13 | Benign | 0.00 | Affected | 0.1885 | 0.2567 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1426T>A | F476I 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant F476I has no ClinVar entry and is not reported in gnomAD. Functional prediction tools fall into two groups: benign predictions come from REVEL, premPS, PROVEAN, SIFT, and FATHMM; pathogenic predictions come from FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—remains inconclusive and is treated as unavailable. With seven tools indicating pathogenicity versus five indicating benign, and two high‑accuracy tools supporting pathogenicity, the evidence points to a likely pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no record for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | -12.617 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 3.90 | Destabilizing | 0.1 | 3.09 | Destabilizing | 3.50 | Destabilizing | 0.39 | Likely Benign | 0.239 | Likely Benign | -1.23 | Neutral | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 3.50 | Benign | 0.37 | Tolerated | 0.1383 | 0.2202 | 1 | 0 | 1.7 | -34.02 | ||||||||||||||||||||||||||||||
| c.1426T>C | F476L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F476L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also inconclusive. No other tools provide definitive evidence. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | -10.109 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 1.00 | Ambiguous | 0.1 | 1.04 | Ambiguous | 1.02 | Ambiguous | 0.75 | Ambiguous | 0.196 | Likely Benign | -1.10 | Neutral | 0.997 | Probably Damaging | 0.978 | Probably Damaging | 3.53 | Benign | 0.60 | Tolerated | 3.40 | 22 | 0.1653 | 0.2916 | 2 | 0 | 1.0 | -34.02 | 235.9 | 16.1 | 0.0 | 0.1 | -0.2 | 0.0 | X | Potentially Benign | In the WT simulations, the phenyl ring of Phe476, located at the end of an α-helix (res. Ala461-Phe476), packs with the hydrophobic side chains of Leu482 and Ile483. Additionally, Phe476 stacks with the Arg475 side chain on the preceding α-α loop connecting the two α-helices (res. Ala461-Phe476 and res. Leu489-Glu519) near the GAP-Ras interface.In the variant simulations, Leu476 can maintain hydrophobic packing with neighboring residues, although not as efficiently as the phenylalanine in the WT system. The absence of Phe476/Arg475 stacking weakens the integrity of the α-helix end in the variant simulations. Nonetheless, no large-scale adverse effects are observed in the simulations. Lastly, the potential effect of the residue swap on SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | |||||||||||||||||||
| c.1426T>G | F476V 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 F476V variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM. Those that agree on a pathogenic effect are FoldX, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and Foldetta. Tools with uncertain or mixed outputs are Rosetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a pathogenic impact. Overall, the majority of evidence points toward a pathogenic effect. The variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | -12.329 | Likely Pathogenic | 0.918 | Likely Pathogenic | Ambiguous | 3.01 | Destabilizing | 0.2 | 1.90 | Ambiguous | 2.46 | Destabilizing | 0.64 | Ambiguous | 0.353 | Likely Benign | -1.63 | Neutral | 0.996 | Probably Damaging | 0.993 | Probably Damaging | 3.49 | Benign | 0.53 | Tolerated | 0.1478 | 0.2251 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||
| c.1427T>A | F476Y 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F476Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default; FoldX is uncertain and therefore not counted. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta predicts a benign outcome. Overall, the balance of evidence leans toward a benign impact for F476Y, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | -9.707 | Likely Pathogenic | 0.576 | Likely Pathogenic | Likely Benign | 0.50 | Ambiguous | 0.2 | 0.30 | Likely Benign | 0.40 | Likely Benign | 1.10 | Destabilizing | 0.169 | Likely Benign | -1.10 | Neutral | 0.965 | Probably Damaging | 0.919 | Probably Damaging | 3.46 | Benign | 0.90 | Tolerated | 0.1109 | 0.1568 | 7 | 3 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||
| c.1427T>C | F476S 2D  AIThe SynGAP1 missense variant F476S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, whereas a majority of tools predict a pathogenic impact: FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields an equal split of pathogenic and benign signals. Overall, the balance of evidence favors a pathogenic effect for F476S, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | -12.675 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 3.02 | Destabilizing | 0.9 | 4.07 | Destabilizing | 3.55 | Destabilizing | 1.23 | Destabilizing | 0.278 | Likely Benign | -2.33 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.56 | Benign | 0.44 | Tolerated | 0.3387 | 0.0558 | -3 | -2 | -3.6 | -60.10 | ||||||||||||||||||||||||||||||
| c.1427T>G | F476C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F476C is catalogued in gnomAD (ID 6‑33438459‑T‑G) but has no ClinVar entry. Functional prediction tools split in two groups: benign predictions come from REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from FoldX, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, seven tools predict pathogenicity versus six predicting benign, with no ClinVar evidence to contradict this assessment. Thus, the variant is most likely pathogenic based on the current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | 6-33438459-T-G | -9.270 | Likely Pathogenic | 0.745 | Likely Pathogenic | Likely Benign | 2.05 | Destabilizing | 0.1 | 2.62 | Destabilizing | 2.34 | Destabilizing | -0.30 | Likely Benign | 0.280 | Likely Benign | 2.69 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.46 | Benign | 0.83 | Tolerated | 3.40 | 22 | 0.2051 | 0.1251 | -2 | -4 | -0.3 | -44.04 | |||||||||||||||||||||||||||
| c.1428C>A | F476L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F476L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also inconclusive. No other tools provide definitive evidence. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | -10.109 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 1.00 | Ambiguous | 0.1 | 1.04 | Ambiguous | 1.02 | Ambiguous | 0.75 | Ambiguous | 0.180 | Likely Benign | -1.10 | Neutral | 0.997 | Probably Damaging | 0.978 | Probably Damaging | 3.53 | Benign | 0.60 | Tolerated | 3.40 | 22 | 0.1653 | 0.2916 | 2 | 0 | 1.0 | -34.02 | 235.9 | 16.1 | 0.0 | 0.1 | -0.2 | 0.0 | X | Potentially Benign | In the WT simulations, the phenyl ring of Phe476, located at the end of an α-helix (res. Ala461-Phe476), packs with the hydrophobic side chains of Leu482 and Ile483. Additionally, Phe476 stacks with the Arg475 side chain on the preceding α-α loop connecting the two α-helices (res. Ala461-Phe476 and res. Leu489-Glu519) near the GAP-Ras interface.In the variant simulations, Leu476 can maintain hydrophobic packing with neighboring residues, although not as efficiently as the phenylalanine in the WT system. The absence of Phe476/Arg475 stacking weakens the integrity of the α-helix end in the variant simulations. Nonetheless, no large-scale adverse effects are observed in the simulations. Lastly, the potential effect of the residue swap on SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | |||||||||||||||||||
| c.1428C>G | F476L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F476L is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438460‑C‑G). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, SIFT, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that returned uncertain results—FoldX, Rosetta, Foldetta, and premPS—do not contribute to the assessment. High‑accuracy methods give the following: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, with two pathogenic and two benign calls; Foldetta also reports an uncertain stability change. Overall, the balance of evidence favors a pathogenic effect for F476L, which contrasts with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.397815 | Uncertain | 0.821 | 0.250 | 0.000 | Uncertain | 2 | 6-33438460-C-G | 4 | 2.48e-6 | -10.109 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 1.00 | Ambiguous | 0.1 | 1.04 | Ambiguous | 1.02 | Ambiguous | 0.75 | Ambiguous | 0.180 | Likely Benign | -1.10 | Neutral | 0.997 | Probably Damaging | 0.978 | Probably Damaging | 3.53 | Benign | 0.60 | Tolerated | 3.40 | 22 | 0.1653 | 0.2916 | 2 | 0 | 1.0 | -34.02 | 235.9 | 16.1 | 0.0 | 0.1 | -0.2 | 0.0 | X | Potentially Benign | In the WT simulations, the phenyl ring of Phe476, located at the end of an α-helix (res. Ala461-Phe476), packs with the hydrophobic side chains of Leu482 and Ile483. Additionally, Phe476 stacks with the Arg475 side chain on the preceding α-α loop connecting the two α-helices (res. Ala461-Phe476 and res. Leu489-Glu519) near the GAP-Ras interface.In the variant simulations, Leu476 can maintain hydrophobic packing with neighboring residues, although not as efficiently as the phenylalanine in the WT system. The absence of Phe476/Arg475 stacking weakens the integrity of the α-helix end in the variant simulations. Nonetheless, no large-scale adverse effects are observed in the simulations. Lastly, the potential effect of the residue swap on SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | ||||||||||||||
| c.142T>A | F48I 2D AIThe SynGAP1 missense variant F48I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM, while SIFT and AlphaMissense‑Default predict a pathogenic impact. Two tools—ESM1b and AlphaMissense‑Optimized—return uncertain results. High‑accuracy assessments further clarify the variant’s likely benign nature: the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, favors benign; AlphaMissense‑Optimized remains uncertain, and Foldetta (which would evaluate protein‑folding stability) is unavailable. Overall, the balance of evidence points to a benign classification, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.298791 | Structured | 0.440452 | Uncertain | 0.558 | 0.707 | 0.125 | -7.994 | In-Between | 0.808 | Likely Pathogenic | Ambiguous | 0.108 | Likely Benign | -1.67 | Neutral | 0.092 | Benign | 0.050 | Benign | 3.99 | Benign | 0.00 | Affected | 0.2561 | 0.2301 | 1 | 0 | 1.7 | -34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.142T>G | F48V 2D AIThe SynGAP1 missense variant F48V is not reported in ClinVar and is absent from gnomAD. Consensus from routine in silico predictors shows six tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized) classifying the change as benign, while two (SIFT, AlphaMissense‑Default) predict pathogenicity; ESM1b remains uncertain. High‑accuracy assessment further supports a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority vote, and Foldetta data are unavailable. Consequently, the overall evidence points to a benign effect for F48V, and this conclusion does not conflict with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.298791 | Structured | 0.440452 | Uncertain | 0.558 | 0.707 | 0.125 | -7.591 | In-Between | 0.756 | Likely Pathogenic | Likely Benign | 0.144 | Likely Benign | -1.87 | Neutral | 0.092 | Benign | 0.037 | Benign | 4.00 | Benign | 0.00 | Affected | 0.2533 | 0.2300 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.1430T>A | M477K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS and FATHMM. Predictions that are uncertain or inconclusive are FoldX, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is unavailable due to mixed or uncertain inputs. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the current ClinVar status, which contains no report of pathogenicity. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -7.519 | In-Between | 0.553 | Ambiguous | Likely Benign | 0.54 | Ambiguous | 0.1 | 0.37 | Likely Benign | 0.46 | Likely Benign | 1.12 | Destabilizing | 0.371 | Likely Benign | -1.32 | Neutral | 0.254 | Benign | 0.122 | Benign | -1.15 | Pathogenic | 0.14 | Tolerated | 0.1817 | 0.0847 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||
| c.1430T>C | M477T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477T has no ClinVar entry and is present in gnomAD (ID 6‑33438462‑T‑C). Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. The remaining tools (FoldX, Foldetta, premPS, AlphaMissense‑Default) return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign (2 benign vs. 1 pathogenic votes); Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | 6-33438462-T-C | 2 | 1.24e-6 | -2.509 | Likely Benign | 0.373 | Ambiguous | Likely Benign | 1.62 | Ambiguous | 0.2 | 0.16 | Likely Benign | 0.89 | Ambiguous | 0.51 | Ambiguous | 0.273 | Likely Benign | -1.33 | Neutral | 0.765 | Possibly Damaging | 0.363 | Benign | -1.10 | Pathogenic | 0.40 | Tolerated | 3.37 | 34 | 0.2177 | 0.1950 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||
| c.1430T>G | M477R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are premPS, polyPhen‑2 HumDiv, and FATHMM. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, while Foldetta remains uncertain and is not taken as evidence. Overall, the majority of reliable predictors indicate a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -6.786 | Likely Benign | 0.552 | Ambiguous | Likely Benign | 0.48 | Likely Benign | 0.2 | 0.77 | Ambiguous | 0.63 | Ambiguous | 1.24 | Destabilizing | 0.442 | Likely Benign | -1.12 | Neutral | 0.720 | Possibly Damaging | 0.242 | Benign | -1.22 | Pathogenic | 0.11 | Tolerated | 0.1901 | 0.0828 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||
| c.1431G>A | M477I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while FoldX, Foldetta, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, indicating no contradiction with existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -2.662 | Likely Benign | 0.467 | Ambiguous | Likely Benign | 0.72 | Ambiguous | 0.1 | 0.36 | Likely Benign | 0.54 | Ambiguous | 0.45 | Likely Benign | 0.291 | Likely Benign | -1.57 | Neutral | 0.000 | Benign | 0.001 | Benign | -1.21 | Pathogenic | 0.09 | Tolerated | 0.1368 | 0.3176 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1431G>C | M477I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized all predict benign. Only FATHMM predicts pathogenic, while FoldX, Foldetta, and AlphaMissense‑Default are uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta remains uncertain. Overall, the majority of evidence supports a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -2.662 | Likely Benign | 0.467 | Ambiguous | Likely Benign | 0.72 | Ambiguous | 0.1 | 0.36 | Likely Benign | 0.54 | Ambiguous | 0.45 | Likely Benign | 0.290 | Likely Benign | -1.57 | Neutral | 0.000 | Benign | 0.001 | Benign | -1.21 | Pathogenic | 0.09 | Tolerated | 0.1368 | 0.3176 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1431G>T | M477I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized all predict benign. Only FATHMM predicts pathogenic, while FoldX, Foldetta, and AlphaMissense‑Default are uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta remains uncertain. Overall, the majority of evidence supports a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -2.662 | Likely Benign | 0.467 | Ambiguous | Likely Benign | 0.72 | Ambiguous | 0.1 | 0.36 | Likely Benign | 0.54 | Ambiguous | 0.45 | Likely Benign | 0.291 | Likely Benign | -1.57 | Neutral | 0.000 | Benign | 0.001 | Benign | -1.21 | Pathogenic | 0.09 | Tolerated | 0.1368 | 0.3176 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1432G>C | E478Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E478Q is listed in gnomAD (ID 6‑33438464‑G‑C) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑vs‑2 split. Overall, the majority of evidence (nine benign vs three pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | 6-33438464-G-C | 1 | 6.20e-7 | -9.881 | Likely Pathogenic | 0.603 | Likely Pathogenic | Likely Benign | -0.04 | Likely Benign | 0.0 | 0.31 | Likely Benign | 0.14 | Likely Benign | 0.07 | Likely Benign | 0.222 | Likely Benign | -2.49 | Neutral | 0.623 | Possibly Damaging | 0.199 | Benign | 3.40 | Benign | 0.14 | Tolerated | 3.37 | 34 | 0.1027 | 0.5867 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||
| c.1433A>G | E478G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 E478G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and polyPhen‑2 HumDiv. Uncertain predictions come from Rosetta, ESM1b, AlphaMissense‑Default, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to equal benign and pathogenic signals and two uncertain calls; Foldetta likewise yields an uncertain result. Overall, the majority of evaluated tools (seven benign vs. two pathogenic) support a benign classification. This consensus does not contradict any ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | -7.897 | In-Between | 0.418 | Ambiguous | Likely Benign | 0.40 | Likely Benign | 0.0 | 0.73 | Ambiguous | 0.57 | Ambiguous | 0.02 | Likely Benign | 0.306 | Likely Benign | -4.91 | Deleterious | 0.923 | Possibly Damaging | 0.427 | Benign | 3.41 | Benign | 0.15 | Tolerated | 0.2758 | 0.5572 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||
| c.1440G>C | E480D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E480D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and ESM1b, whereas polyPhen‑2 (HumDiv and HumVar) and FATHMM predict a pathogenic impact. The remaining tools—FoldX, Rosetta, premPS, AlphaMissense‑Default, and Foldetta—return uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority, and Foldetta remains uncertain. Overall, the balance of evidence favors a benign classification for E480D, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.216401 | Structured | 0.426867 | Uncertain | 0.798 | 0.250 | 0.000 | -3.001 | Likely Benign | 0.475 | Ambiguous | Likely Benign | 0.62 | Ambiguous | 0.2 | 1.39 | Ambiguous | 1.01 | Ambiguous | 0.61 | Ambiguous | 0.405 | Likely Benign | -0.77 | Neutral | 0.989 | Probably Damaging | 0.979 | Probably Damaging | -1.27 | Pathogenic | 0.28 | Tolerated | 0.1526 | 0.4130 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1440G>T | E480D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E480D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and ESM1b, while polyPhen‑2 (HumDiv and HumVar) and FATHMM predict a pathogenic outcome. The remaining tools—FoldX, Rosetta, premPS, AlphaMissense‑Default, and Foldetta—return uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward benign, and Foldetta providing no definitive stability change. Overall, the balance of evidence favors a benign interpretation, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.216401 | Structured | 0.426867 | Uncertain | 0.798 | 0.250 | 0.000 | -3.001 | Likely Benign | 0.475 | Ambiguous | Likely Benign | 0.62 | Ambiguous | 0.2 | 1.39 | Ambiguous | 1.01 | Ambiguous | 0.61 | Ambiguous | 0.405 | Likely Benign | -0.77 | Neutral | 0.989 | Probably Damaging | 0.979 | Probably Damaging | -1.27 | Pathogenic | 0.28 | Tolerated | 0.1526 | 0.4130 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1441C>A | H481N 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 H481N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools—Rosetta and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current predictive data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.257454 | Structured | 0.430977 | Uncertain | 0.764 | 0.247 | 0.000 | -8.229 | Likely Pathogenic | 0.381 | Ambiguous | Likely Benign | 0.16 | Likely Benign | 0.1 | 0.54 | Ambiguous | 0.35 | Likely Benign | 0.09 | Likely Benign | 0.205 | Likely Benign | -4.11 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.68 | Benign | 0.57 | Tolerated | 0.1205 | 0.1631 | 2 | 1 | -0.3 | -23.04 | ||||||||||||||||||||||||||||||
| c.1447A>G | I483V 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I483V is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, Rosetta, PROVEAN, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions are reported by premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Predictions marked as uncertain include FoldX, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, whereas Foldetta remains uncertain. Overall, the balance of evidence from both general and high‑accuracy tools leans toward a benign effect, which does not contradict the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.206376 | Structured | 0.415850 | Uncertain | 0.798 | 0.254 | 0.000 | Conflicting | 2 | -10.121 | Likely Pathogenic | 0.523 | Ambiguous | Likely Benign | 1.00 | Ambiguous | 0.0 | 0.27 | Likely Benign | 0.64 | Ambiguous | 1.02 | Destabilizing | 0.228 | Likely Benign | -0.86 | Neutral | 0.914 | Possibly Damaging | 0.921 | Probably Damaging | 3.23 | Benign | 0.03 | Affected | 3.37 | 32 | 0.0876 | 0.2691 | 3 | 4 | -0.3 | -14.03 | ||||||||||||||||||||||||||
| c.145T>A | C49S 2D  AIThe SynGAP1 missense variant C49S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of standard predictors lean toward pathogenicity, but the high‑accuracy tools do not provide definitive support. Thus, the variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.575 | Likely Benign | 0.704 | Likely Pathogenic | Likely Benign | 0.260 | Likely Benign | -3.07 | Deleterious | 0.462 | Possibly Damaging | 0.478 | Possibly Damaging | 3.91 | Benign | 0.00 | Affected | 0.3855 | 0.1686 | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.145T>G | C49G 2D  AIThe SynGAP1 missense variant C49G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic versus four benign) lean toward a pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.464 | Likely Benign | 0.674 | Likely Pathogenic | Likely Benign | 0.351 | Likely Benign | -3.64 | Deleterious | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.87 | Benign | 0.00 | Affected | 0.2419 | 0.2528 | -3 | -3 | -2.9 | -46.09 | ||||||||||||||||||||||||||||||||||||||||
| c.1462A>T | T488S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T488S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Four tools (Foldetta, premPS, ESM1b, and Rosetta) return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of standard tools lean toward a benign interpretation, but the high‑accuracy consensus is split, leaving the variant’s impact ambiguous. No ClinVar annotation contradicts these predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.206376 | Structured | 0.332663 | Uncertain | 0.928 | 0.233 | 0.125 | -7.662 | In-Between | 0.745 | Likely Pathogenic | Likely Benign | 0.35 | Likely Benign | 0.1 | 0.90 | Ambiguous | 0.63 | Ambiguous | 0.77 | Ambiguous | 0.257 | Likely Benign | -3.51 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 3.73 | Benign | 0.31 | Tolerated | 0.2256 | 0.2813 | 1 | 1 | -0.1 | -14.03 | ||||||||||||||||||||||||||||||
| c.1465C>A | L489I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L489I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into three groups: benign predictions come from REVEL, PROVEAN, SIFT, and AlphaMissense‑Optimized; pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; the remaining methods (FoldX, Rosetta, Foldetta, premPS, ESM1b, AlphaMissense‑Default) yield uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is indeterminate due to a tie between pathogenic and benign signals, and Foldetta reports an uncertain stability change. Overall, the preponderance of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.326126 | Uncertain | 0.949 | 0.234 | 0.125 | -7.333 | In-Between | 0.342 | Ambiguous | Likely Benign | 1.01 | Ambiguous | 0.0 | 0.52 | Ambiguous | 0.77 | Ambiguous | 0.90 | Ambiguous | 0.490 | Likely Benign | -1.55 | Neutral | 0.999 | Probably Damaging | 0.997 | Probably Damaging | -1.42 | Pathogenic | 0.21 | Tolerated | 0.1069 | 0.4080 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.146G>C | C49S 2D AIThe SynGAP1 missense variant C49S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the majority of conventional tools (five pathogenic vs four benign) lean toward a pathogenic interpretation, but the single high‑accuracy benign prediction and the inconclusive SGM Consensus leave the assessment uncertain. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.575 | Likely Benign | 0.704 | Likely Pathogenic | Likely Benign | 0.224 | Likely Benign | -3.07 | Deleterious | 0.462 | Possibly Damaging | 0.478 | Possibly Damaging | 3.91 | Benign | 0.00 | Affected | 0.3855 | 0.1686 | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.146G>T | C49F 2D  AIThe SynGAP1 missense variant C49F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and FATHMM, while a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default) predict a pathogenic impact. The remaining tools, ESM1b and AlphaMissense‑Optimized, return uncertain results. High‑accuracy assessments further support a deleterious interpretation: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (benign), and PROVEAN (pathogenic)—classifies the variant as pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -7.194 | In-Between | 0.893 | Likely Pathogenic | Ambiguous | 0.301 | Likely Benign | -3.21 | Deleterious | 0.676 | Possibly Damaging | 0.695 | Possibly Damaging | 3.86 | Benign | 0.00 | Affected | 0.1199 | 0.3690 | -4 | -2 | 0.3 | 44.04 | ||||||||||||||||||||||||||||||||||||||||
| c.1480A>C | I494L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I494L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the balance of evidence (five pathogenic versus four benign predictions, with several uncertain calls) leans toward a pathogenic interpretation. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | -10.175 | Likely Pathogenic | 0.478 | Ambiguous | Likely Benign | 0.28 | Likely Benign | 0.1 | 1.11 | Ambiguous | 0.70 | Ambiguous | 0.91 | Ambiguous | 0.513 | Likely Pathogenic | -1.69 | Neutral | 0.645 | Possibly Damaging | 0.718 | Possibly Damaging | -0.88 | Pathogenic | 0.11 | Tolerated | 0.0816 | 0.2851 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1480A>G | I494V 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant I494V is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438512‑A‑G). Functional prediction tools that agree on benign impact include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Pathogenic predictions come from premPS and FATHMM. Predictions that are inconclusive are FoldX, Rosetta, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign; Foldetta remains uncertain. Overall, the majority of evidence supports a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | Conflicting | 2 | 6-33438512-A-G | 36 | 2.23e-5 | -7.102 | In-Between | 0.112 | Likely Benign | Likely Benign | 1.16 | Ambiguous | 0.0 | 0.71 | Ambiguous | 0.94 | Ambiguous | 1.02 | Destabilizing | 0.439 | Likely Benign | -0.83 | Neutral | 0.278 | Benign | 0.179 | Benign | -1.30 | Pathogenic | 0.07 | Tolerated | 3.37 | 35 | 0.0965 | 0.2491 | 4 | 3 | -0.3 | -14.03 | 248.6 | 29.3 | 0.0 | 0.0 | -1.1 | 0.5 | X | Potentially Benign | The sec-butyl side chain of Ile494, located in an α-helix (res. Leu489-Glu519), packs against hydrophobic residues (e.g., Phe484, Leu465, Trp572, Ala493, Met468) in an inter-helix space (res. Leu489-Glu519 and res. Ala461-Phe476). In the variant simulations, the hydrophobic iso-propyl side chain of Val494, which is of a similar size and has similar physicochemical properties to Ile494 in the WT, resides similarly in the inter-helix hydrophobic space. Thus, no negative effects on the protein structure are observed. | ||||||||||||||
| c.1480A>T | I494L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I494L variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta indicating uncertain stability change. Overall, the balance of evidence leans toward pathogenicity, with a majority of tools and the SGM Consensus supporting a deleterious effect. This conclusion does not contradict ClinVar status, as the variant is currently unreported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | -10.175 | Likely Pathogenic | 0.478 | Ambiguous | Likely Benign | 0.28 | Likely Benign | 0.1 | 1.11 | Ambiguous | 0.70 | Ambiguous | 0.91 | Ambiguous | 0.513 | Likely Pathogenic | -1.69 | Neutral | 0.645 | Possibly Damaging | 0.718 | Possibly Damaging | -0.88 | Pathogenic | 0.11 | Tolerated | 0.0816 | 0.2851 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1482A>G | I494M 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 I494M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the balance of evidence leans toward pathogenicity, with a majority of tools predicting a deleterious effect. This conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | -8.223 | Likely Pathogenic | 0.356 | Ambiguous | Likely Benign | 0.35 | Likely Benign | 0.2 | 1.98 | Ambiguous | 1.17 | Ambiguous | 0.98 | Ambiguous | 0.542 | Likely Pathogenic | -2.25 | Neutral | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.15 | Pathogenic | 0.23 | Tolerated | 0.0676 | 0.1789 | 2 | 1 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||
| c.1494G>A | M498I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M498I has no ClinVar record (ClinVar ID None) and is not reported in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, SIFT, FATHMM, and AlphaMissense‑Default. Two tools, Rosetta and premPS, give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of predictions lean toward a benign impact, and this is not contradicted by ClinVar status, which lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.092881 | Structured | 0.399612 | Uncertain | 0.932 | 0.158 | 0.000 | -4.796 | Likely Benign | 0.760 | Likely Pathogenic | Likely Benign | 2.56 | Destabilizing | 0.3 | 1.65 | Ambiguous | 2.11 | Destabilizing | 0.88 | Ambiguous | 0.418 | Likely Benign | -1.09 | Neutral | 0.018 | Benign | 0.012 | Benign | -1.28 | Pathogenic | 0.04 | Affected | 0.1184 | 0.2537 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1494G>C | M498I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M498I has no ClinVar record (ClinVar ID None) and is not reported in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, SIFT, FATHMM, and AlphaMissense‑Default. Two tools, Rosetta and premPS, give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of predictions lean toward a benign impact, and this is not contradicted by ClinVar status, which lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.092881 | Structured | 0.399612 | Uncertain | 0.932 | 0.158 | 0.000 | -4.796 | Likely Benign | 0.760 | Likely Pathogenic | Likely Benign | 2.56 | Destabilizing | 0.3 | 1.65 | Ambiguous | 2.11 | Destabilizing | 0.88 | Ambiguous | 0.418 | Likely Benign | -1.09 | Neutral | 0.018 | Benign | 0.012 | Benign | -1.28 | Pathogenic | 0.04 | Affected | 0.1184 | 0.2537 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1494G>T | M498I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M498I has no ClinVar record (ClinVar ID None) and is not reported in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, SIFT, FATHMM, and AlphaMissense‑Default. Two tools, Rosetta and premPS, give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of predictions lean toward a benign impact, and this is not contradicted by ClinVar status, which lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.092881 | Structured | 0.399612 | Uncertain | 0.932 | 0.158 | 0.000 | -4.796 | Likely Benign | 0.760 | Likely Pathogenic | Likely Benign | 2.56 | Destabilizing | 0.3 | 1.65 | Ambiguous | 2.11 | Destabilizing | 0.88 | Ambiguous | 0.414 | Likely Benign | -1.09 | Neutral | 0.018 | Benign | 0.012 | Benign | -1.28 | Pathogenic | 0.04 | Affected | 0.1184 | 0.2537 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||
| c.1498C>A | L500M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L500M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The premPS score is uncertain. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; Foldetta predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive because it yields a 2‑to‑2 split. No other high‑accuracy predictions are available. Overall, the majority of high‑accuracy tools lean toward a benign interpretation, and this assessment does not contradict the lack of ClinVar reporting. Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.382942 | Uncertain | 0.893 | 0.150 | 0.000 | -9.945 | Likely Pathogenic | 0.301 | Likely Benign | Likely Benign | 0.15 | Likely Benign | 0.0 | -0.09 | Likely Benign | 0.03 | Likely Benign | 0.92 | Ambiguous | 0.590 | Likely Pathogenic | -1.99 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.36 | Pathogenic | 0.01 | Affected | 0.0710 | 0.2202 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.149T>A | I50N 2D  AIThe SynGAP1 missense variant I50N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, whereas the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves as benign. Foldetta results are unavailable. Overall, the predictions are mixed, with an equal split between benign and pathogenic calls, but the most reliable single‑tool prediction (AlphaMissense‑Optimized) and the majority of individual tools lean toward pathogenicity. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.295083 | Structured | 0.449965 | Uncertain | 0.545 | 0.708 | 0.000 | -7.091 | In-Between | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.135 | Likely Benign | -2.37 | Neutral | 0.842 | Possibly Damaging | 0.272 | Benign | 3.73 | Benign | 0.00 | Affected | 0.0824 | 0.0412 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.1507C>G | Q503E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q503E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and Rosetta. Uncertain or inconclusive results come from FoldX, Foldetta, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, while Foldetta remains uncertain. Overall, the majority of available predictions favor a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.322935 | Uncertain | 0.848 | 0.168 | 0.000 | -7.909 | In-Between | 0.146 | Likely Benign | Likely Benign | 0.51 | Ambiguous | 0.1 | 2.11 | Destabilizing | 1.31 | Ambiguous | 0.85 | Ambiguous | 0.410 | Likely Benign | -2.21 | Neutral | 0.931 | Possibly Damaging | 0.500 | Possibly Damaging | -1.43 | Pathogenic | 0.17 | Tolerated | 0.1168 | 0.1508 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1509G>C | Q503H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q503H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict pathogenicity are PROVEAN, SIFT, and FATHMM. FoldX, Rosetta, and Foldetta report uncertain or inconclusive stability changes and are therefore considered unavailable for pathogenicity inference. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and thus inconclusive, and Foldetta remains uncertain. Overall, the majority of evaluated predictors (7 benign vs. 3 pathogenic) lean toward a benign effect. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.322935 | Uncertain | 0.848 | 0.168 | 0.000 | -5.335 | Likely Benign | 0.227 | Likely Benign | Likely Benign | 0.94 | Ambiguous | 0.2 | 0.88 | Ambiguous | 0.91 | Ambiguous | 0.48 | Likely Benign | 0.331 | Likely Benign | -3.40 | Deleterious | 0.002 | Benign | 0.004 | Benign | -1.52 | Pathogenic | 0.01 | Affected | 0.1091 | 0.2125 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1509G>T | Q503H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q503H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict pathogenicity are PROVEAN, SIFT, and FATHMM. FoldX, Rosetta, and Foldetta report uncertain or inconclusive stability changes and are therefore considered unavailable for pathogenicity inference. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and thus inconclusive, and Foldetta remains uncertain. Overall, the majority of evaluated predictors (7 benign vs. 3 pathogenic) lean toward a benign effect. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.322935 | Uncertain | 0.848 | 0.168 | 0.000 | -5.335 | Likely Benign | 0.227 | Likely Benign | Likely Benign | 0.94 | Ambiguous | 0.2 | 0.88 | Ambiguous | 0.91 | Ambiguous | 0.48 | Likely Benign | 0.331 | Likely Benign | -3.40 | Deleterious | 0.002 | Benign | 0.004 | Benign | -1.52 | Pathogenic | 0.01 | Affected | 0.1091 | 0.2125 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1510A>C | K504Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K504Q missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta predicts a benign stability change. Overall, seven tools support a benign outcome while four support pathogenicity, with no ClinVar evidence to contradict this assessment. Thus, the variant is most likely benign based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.028107 | Structured | 0.304984 | Uncertain | 0.850 | 0.189 | 0.000 | -6.685 | Likely Benign | 0.238 | Likely Benign | Likely Benign | 0.13 | Likely Benign | 0.2 | -0.01 | Likely Benign | 0.06 | Likely Benign | 0.91 | Ambiguous | 0.269 | Likely Benign | -3.07 | Deleterious | 0.945 | Possibly Damaging | 0.918 | Probably Damaging | -1.37 | Pathogenic | 0.46 | Tolerated | 0.3104 | 0.0780 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||
| c.1514A>T | Y505F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y505F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Predictions that are uncertain or inconclusive are Rosetta, Foldetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of tools (five benign versus four pathogenic) lean toward a benign interpretation, and this is consistent with the lack of ClinVar evidence; thus the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.047319 | Structured | 0.292227 | Uncertain | 0.909 | 0.188 | 0.000 | -8.855 | Likely Pathogenic | 0.437 | Ambiguous | Likely Benign | 0.40 | Likely Benign | 0.0 | 0.76 | Ambiguous | 0.58 | Ambiguous | 0.69 | Ambiguous | 0.434 | Likely Benign | -3.98 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.93 | Benign | 0.07 | Tolerated | 0.2409 | 0.2598 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||
| c.1516C>A | L506I 2D AIThe SynGAP1 missense variant L506I is not reported in ClinVar and is absent from gnomAD. Consensus from standard in‑silico predictors shows a split: benign calls come from REVEL, PROVEAN, and AlphaMissense‑Optimized, whereas pathogenic calls arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Uncertain results are reported by FoldX, premPS, and AlphaMissense‑Default. High‑accuracy assessments give a pathogenic verdict from Foldetta (a combined FoldX‑MD/Rosetta stability analysis) and from the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). AlphaMissense‑Optimized, however, predicts benign. Overall, the majority of reliable tools and the high‑accuracy methods favor a pathogenic effect, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.034884 | Structured | 0.279180 | Uncertain | 0.924 | 0.196 | 0.000 | -10.386 | Likely Pathogenic | 0.501 | Ambiguous | Likely Benign | 1.73 | Ambiguous | 1.0 | 2.85 | Destabilizing | 2.29 | Destabilizing | 0.98 | Ambiguous | 0.365 | Likely Benign | -1.99 | Neutral | 0.998 | Probably Damaging | 0.997 | Probably Damaging | 1.65 | Pathogenic | 0.01 | Affected | 0.0745 | 0.2033 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1519A>C | K507Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K507Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Uncertain results come from Rosetta, premPS, and ESM1b. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. Taken together, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.019401 | Structured | 0.262601 | Uncertain | 0.885 | 0.222 | 0.000 | -7.698 | In-Between | 0.180 | Likely Benign | Likely Benign | 0.28 | Likely Benign | 0.0 | -0.70 | Ambiguous | -0.21 | Likely Benign | -0.52 | Ambiguous | 0.443 | Likely Benign | 0.22 | Neutral | 0.999 | Probably Damaging | 0.999 | Probably Damaging | -1.52 | Pathogenic | 0.86 | Tolerated | 0.2952 | 0.0713 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||
| c.1519A>G | K507E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K507E missense change is not listed in ClinVar and has no gnomAD allele, indicating it is not a common polymorphism. Functional prediction tools cluster into two groups: benign predictions come from Rosetta, PROVEAN, SIFT, and AlphaMissense‑Optimized, while pathogenic predictions arise from REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Four tools give uncertain results: FoldX, Foldetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments provide a mixed picture: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans pathogenic, and Foldetta remains uncertain. Overall, the majority of standard predictors favor a pathogenic effect, and the high‑accuracy consensus also tilts toward pathogenicity, though not decisively. Therefore, the variant is most likely pathogenic, and this assessment does not contradict ClinVar status because the variant is not yet reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.019401 | Structured | 0.262601 | Uncertain | 0.885 | 0.222 | 0.000 | -11.507 | Likely Pathogenic | 0.448 | Ambiguous | Likely Benign | 0.70 | Ambiguous | 0.0 | 0.31 | Likely Benign | 0.51 | Ambiguous | 0.67 | Ambiguous | 0.536 | Likely Pathogenic | -0.84 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | -1.45 | Pathogenic | 0.21 | Tolerated | 0.2537 | 0.0488 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||
| c.1520A>G | K507R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K507R is not listed in ClinVar (ClinVar status: not reported) but is present in gnomAD (gnomAD ID 6-33438552-A-G). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No prediction or folding stability result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no pathogenic annotation for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.019401 | Structured | 0.262601 | Uncertain | 0.885 | 0.222 | 0.000 | 6-33438552-A-G | 1 | 6.20e-7 | -7.363 | In-Between | 0.085 | Likely Benign | Likely Benign | -0.08 | Likely Benign | 0.1 | 0.38 | Likely Benign | 0.15 | Likely Benign | 0.36 | Likely Benign | 0.503 | Likely Pathogenic | -0.68 | Neutral | 0.992 | Probably Damaging | 0.980 | Probably Damaging | -1.48 | Pathogenic | 0.27 | Tolerated | 3.37 | 35 | 0.3155 | 0.0579 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||
| c.1522G>A | D508N 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 D508N missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions (REVEL, FoldX, premPS, FATHMM, AlphaMissense‑Optimized) and pathogenic predictions (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b). Three tools remain uncertain (Rosetta, Foldetta, AlphaMissense‑Default). High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized classifies the change as benign; the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—returns a pathogenic verdict; Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, yields an inconclusive result. Because the consensus of the high‑accuracy methods is split, the overall prediction is ambiguous, but the balance of evidence leans toward pathogenicity. This assessment does not contradict ClinVar, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.019401 | Structured | 0.255890 | Uncertain | 0.890 | 0.228 | 0.000 | -9.909 | Likely Pathogenic | 0.411 | Ambiguous | Likely Benign | 0.11 | Likely Benign | 0.1 | 1.24 | Ambiguous | 0.68 | Ambiguous | -0.12 | Likely Benign | 0.265 | Likely Benign | -4.62 | Deleterious | 0.870 | Possibly Damaging | 0.615 | Possibly Damaging | 3.32 | Benign | 0.04 | Affected | 0.1577 | 0.4599 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.1525G>A | A509T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A509T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain and therefore unavailable for interpretation. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.025762 | Structured | 0.250110 | Uncertain | 0.923 | 0.256 | 0.000 | -8.961 | Likely Pathogenic | 0.171 | Likely Benign | Likely Benign | 0.95 | Ambiguous | 0.4 | 0.61 | Ambiguous | 0.78 | Ambiguous | 0.04 | Likely Benign | 0.360 | Likely Benign | -2.15 | Neutral | 0.031 | Benign | 0.058 | Benign | -1.25 | Pathogenic | 0.32 | Tolerated | 0.1344 | 0.5593 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||
| c.1525G>T | A509S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A509S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, polyPhen‑2 HumDiv, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM. Four tools give uncertain or inconclusive results (FoldX, Rosetta, Foldetta, premPS). High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and thus inconclusive, and Foldetta is also inconclusive. Overall, the majority of standard predictors lean toward pathogenicity, while the most reliable single‑tool prediction (AlphaMissense‑Optimized) suggests benign. Given the balance of evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.025762 | Structured | 0.250110 | Uncertain | 0.923 | 0.256 | 0.000 | -9.997 | Likely Pathogenic | 0.171 | Likely Benign | Likely Benign | 0.83 | Ambiguous | 0.1 | 1.63 | Ambiguous | 1.23 | Ambiguous | 0.59 | Ambiguous | 0.621 | Likely Pathogenic | -2.18 | Neutral | 0.119 | Benign | 0.468 | Possibly Damaging | -1.35 | Pathogenic | 0.01 | Affected | 0.2410 | 0.4384 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.152T>A | I51N 2D AIThe SynGAP1 missense variant I51N is not reported in ClinVar (ClinVar status: not reported) and is absent from gnomAD (gnomAD: not present). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method, has no available output for this variant. Consequently, the evidence is split evenly between benign and pathogenic predictions, with no decisive support from the most accurate methods. The variant is therefore inconclusive; it is not contradicted by any ClinVar record. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.291804 | Structured | 0.454181 | Uncertain | 0.606 | 0.710 | 0.000 | -9.287 | Likely Pathogenic | 0.909 | Likely Pathogenic | Ambiguous | 0.155 | Likely Benign | -1.77 | Neutral | 0.704 | Possibly Damaging | 0.272 | Benign | 4.13 | Benign | 0.00 | Affected | 0.1005 | 0.0769 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.152T>G | I51S 2D  AIThe SynGAP1 missense variant I51S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The remaining tools, ESM1b and AlphaMissense‑Optimized, are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign (2 benign vs. 1 pathogenic vote). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.291804 | Structured | 0.454181 | Uncertain | 0.606 | 0.710 | 0.000 | -7.603 | In-Between | 0.879 | Likely Pathogenic | Ambiguous | 0.220 | Likely Benign | -1.39 | Neutral | 0.182 | Benign | 0.099 | Benign | 4.15 | Benign | 0.00 | Affected | 0.3142 | 0.0957 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.1530T>G | I510M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I510M missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign impact include PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b) returned uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (benign), ESM1b (uncertain), FATHMM (pathogenic), and PROVEAN (benign)—also favors benign. Foldetta, a protein‑folding stability method, yielded an uncertain outcome. Taken together, the consensus of the most reliable predictors indicates a benign effect. This conclusion does not contradict the ClinVar status, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.025762 | Structured | 0.250630 | Uncertain | 0.945 | 0.273 | 0.000 | -7.988 | In-Between | 0.235 | Likely Benign | Likely Benign | 0.56 | Ambiguous | 0.3 | 1.61 | Ambiguous | 1.09 | Ambiguous | 0.55 | Ambiguous | 0.532 | Likely Pathogenic | -0.97 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | -1.42 | Pathogenic | 0.02 | Affected | 0.0674 | 0.1789 | 2 | 1 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||
| c.1540A>C | I514L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I514L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools (premPS and AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta also predicts a benign outcome. No prediction or folding stability result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation—there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.049374 | Structured | 0.221408 | Uncertain | 0.948 | 0.266 | 0.000 | -10.239 | Likely Pathogenic | 0.490 | Ambiguous | Likely Benign | -0.05 | Likely Benign | 0.2 | 0.34 | Likely Benign | 0.15 | Likely Benign | 0.81 | Ambiguous | 0.310 | Likely Benign | -1.99 | Neutral | 0.879 | Possibly Damaging | 0.985 | Probably Damaging | 3.31 | Benign | 0.12 | Tolerated | 0.0767 | 0.2678 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1546G>T | A516S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A516S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Two tools remain uncertain: premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.067594 | Structured | 0.167423 | Uncertain | 0.938 | 0.284 | 0.000 | -9.639 | Likely Pathogenic | 0.562 | Ambiguous | Likely Benign | 0.22 | Likely Benign | 0.2 | 0.24 | Likely Benign | 0.23 | Likely Benign | 0.52 | Ambiguous | 0.448 | Likely Benign | -2.45 | Neutral | 0.973 | Probably Damaging | 0.993 | Probably Damaging | -1.24 | Pathogenic | 0.17 | Tolerated | 0.2730 | 0.4730 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.154T>C | S52P 2D  AIThe SynGAP1 missense variant S52P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default all predict a pathogenic outcome. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign votes); and Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact for S52P. This conclusion does not contradict ClinVar, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.291804 | Structured | 0.457753 | Uncertain | 0.499 | 0.677 | 0.000 | -10.007 | Likely Pathogenic | 0.797 | Likely Pathogenic | Ambiguous | 0.144 | Likely Benign | -1.28 | Neutral | 0.676 | Possibly Damaging | 0.693 | Possibly Damaging | 4.07 | Benign | 0.00 | Affected | 0.2276 | 0.5909 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||||
| c.1553A>T | Y518F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 Y518F missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, AlphaMissense‑Optimized, and Foldetta. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools are uncertain: Rosetta and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of individual predictors (seven benign vs. four pathogenic) support a benign classification, and this does not contradict the ClinVar status, which has no entry for this variant. Thus, based on the available predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.139895 | Structured | 0.126970 | Uncertain | 0.897 | 0.321 | 0.000 | -10.617 | Likely Pathogenic | 0.466 | Ambiguous | Likely Benign | 0.34 | Likely Benign | 0.1 | -0.61 | Ambiguous | -0.14 | Likely Benign | 0.41 | Likely Benign | 0.318 | Likely Benign | -3.68 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 3.50 | Benign | 0.22 | Tolerated | 0.2107 | 0.2230 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||
| c.1555G>C | E519Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E519Q missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a benign effect on protein stability. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.078022 | Structured | 0.104514 | Uncertain | 0.899 | 0.328 | 0.000 | -8.693 | Likely Pathogenic | 0.792 | Likely Pathogenic | Ambiguous | -0.35 | Likely Benign | 0.1 | -0.14 | Likely Benign | -0.25 | Likely Benign | -0.21 | Likely Benign | 0.195 | Likely Benign | -2.48 | Neutral | 0.994 | Probably Damaging | 0.986 | Probably Damaging | 3.28 | Benign | 0.14 | Tolerated | 0.1394 | 0.3418 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.155C>G | S52W 2D AIThe SynGAP1 missense variant S52W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). AlphaMissense‑Optimized is uncertain, and Foldetta results are unavailable. Overall, more tools (five) predict pathogenicity than benign (three), and no high‑accuracy consensus or folding‑stability evidence contradicts this trend. Thus, the variant is most likely pathogenic based on the available predictions, and this assessment does not conflict with the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.291804 | Structured | 0.457753 | Uncertain | 0.499 | 0.677 | 0.000 | -8.649 | Likely Pathogenic | 0.909 | Likely Pathogenic | Ambiguous | 0.155 | Likely Benign | -1.85 | Neutral | 0.986 | Probably Damaging | 0.968 | Probably Damaging | 4.05 | Benign | 0.00 | Affected | 0.0580 | 0.6254 | -2 | -3 | -0.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.155C>T | S52L 2D  AISynGAP1 missense variant S52L is listed in ClinVar with an uncertain significance and is present in the gnomAD database (ID 6‑33423564‑C‑T). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors benign. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the collective evidence points to a likely benign effect, which does not contradict the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.291804 | Structured | 0.457753 | Uncertain | 0.499 | 0.677 | 0.000 | Uncertain | 1 | 6-33423564-C-T | 1 | 6.20e-7 | -7.199 | In-Between | 0.688 | Likely Pathogenic | Likely Benign | 0.087 | Likely Benign | -1.41 | Neutral | 0.829 | Possibly Damaging | 0.706 | Possibly Damaging | 4.10 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0991 | 0.6100 | -3 | -2 | 4.6 | 26.08 | |||||||||||||||||||||||||||||||||
| c.1561G>C | E521Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E521Q missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta predicts a benign stability change. Overall, the majority of evidence (9 benign vs. 4 pathogenic predictions) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.086953 | Structured | 0.062387 | Uncertain | 0.865 | 0.349 | 0.000 | -8.310 | Likely Pathogenic | 0.777 | Likely Pathogenic | Likely Benign | -0.28 | Likely Benign | 0.2 | 0.19 | Likely Benign | -0.05 | Likely Benign | -0.06 | Likely Benign | 0.262 | Likely Benign | -2.15 | Neutral | 0.992 | Probably Damaging | 0.993 | Probably Damaging | 3.30 | Benign | 0.11 | Tolerated | 0.1580 | 0.6552 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.1562A>G | E521G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E521G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The high‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it is a 2‑vs‑2 split and is treated as unavailable. Overall, the majority of evidence points to a benign impact. The variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.086953 | Structured | 0.062387 | Uncertain | 0.865 | 0.349 | 0.000 | -6.636 | Likely Benign | 0.767 | Likely Pathogenic | Likely Benign | 0.18 | Likely Benign | 0.1 | 0.58 | Ambiguous | 0.38 | Likely Benign | 0.14 | Likely Benign | 0.289 | Likely Benign | -3.64 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.31 | Benign | 0.07 | Tolerated | 0.2932 | 0.5455 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||
| c.1568A>G | N523S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N523S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and FATHMM. Four tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Default) returned uncertain or inconclusive results. For high‑accuracy assessment, AlphaMissense‑Optimized classifies the variant as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—predicts pathogenicity. Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, reports an uncertain folding‑stability change. Taken together, the majority of evidence (five benign versus three pathogenic predictions) points to a benign effect, and this conclusion does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.033426 | Uncertain | 0.883 | 0.383 | 0.125 | -6.188 | Likely Benign | 0.552 | Ambiguous | Likely Benign | 0.64 | Ambiguous | 0.2 | 0.66 | Ambiguous | 0.65 | Ambiguous | 0.17 | Likely Benign | 0.492 | Likely Benign | -4.31 | Deleterious | 0.976 | Probably Damaging | 0.410 | Benign | -1.27 | Pathogenic | 0.27 | Tolerated | 0.2213 | 0.4178 | 1 | 1 | 2.7 | -27.03 | ||||||||||||||||||||||||||||||
| c.1576G>A | V526I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V526I variant is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (gnomAD ID: 6‑33438819‑G‑A). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized, while those that predict a pathogenic outcome are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of tools (seven benign vs. five pathogenic) lean toward a benign interpretation. Thus, the variant is most likely benign based on current predictions, and this assessment does not contradict the ClinVar status, which has no entry for V526I. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.139895 | Structured | 0.023118 | Uncertain | 0.943 | 0.403 | 0.000 | 6-33438819-G-A | -9.962 | Likely Pathogenic | 0.526 | Ambiguous | Likely Benign | 0.07 | Likely Benign | 0.3 | 0.41 | Likely Benign | 0.24 | Likely Benign | 0.02 | Likely Benign | 0.540 | Likely Pathogenic | -0.99 | Neutral | 0.929 | Possibly Damaging | 0.917 | Probably Damaging | -1.34 | Pathogenic | 0.15 | Tolerated | 3.37 | 35 | 0.0630 | 0.3307 | 3 | 4 | 0.3 | 14.03 | |||||||||||||||||||||||||||
| c.1577T>G | V526G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V526G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on pathogenicity include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default; no tool predicts it benign. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. All available evidence points to a deleterious effect. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.139895 | Structured | 0.023118 | Uncertain | 0.943 | 0.403 | 0.000 | -15.811 | Likely Pathogenic | 0.926 | Likely Pathogenic | Ambiguous | 3.21 | Destabilizing | 0.3 | 4.79 | Destabilizing | 4.00 | Destabilizing | 2.19 | Destabilizing | 0.935 | Likely Pathogenic | -6.92 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.49 | Pathogenic | 0.00 | Affected | 0.1636 | 0.1837 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.157G>A | G53R 2D AIThe SynGAP1 missense variant G53R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic + 2 benign), and Foldetta (combining FoldX‑MD and Rosetta outputs) is unavailable. Overall, the majority of available predictions (5 pathogenic vs. 3 benign) lean toward a pathogenic impact. Because there is no ClinVar entry to contradict this assessment, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.268042 | Structured | 0.460894 | Uncertain | 0.386 | 0.666 | 0.000 | -8.400 | Likely Pathogenic | 0.901 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -1.20 | Neutral | 0.994 | Probably Damaging | 0.990 | Probably Damaging | 4.13 | Benign | 0.00 | Affected | 0.0953 | 0.4347 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.157G>C | G53R 2D AIThe SynGAP1 missense variant G53R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic + 2 benign), and Foldetta (combining FoldX‑MD and Rosetta outputs) is unavailable. Overall, the majority of tools (5 pathogenic vs. 3 benign) predict a pathogenic impact. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.268042 | Structured | 0.460894 | Uncertain | 0.386 | 0.666 | 0.000 | -8.400 | Likely Pathogenic | 0.901 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -1.20 | Neutral | 0.994 | Probably Damaging | 0.990 | Probably Damaging | 4.13 | Benign | 0.00 | Affected | 0.0953 | 0.4347 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.157G>T | G53W 2D AIThe SynGAP1 missense variant G53W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of available predictions (six pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.268042 | Structured | 0.460894 | Uncertain | 0.386 | 0.666 | 0.000 | -11.012 | Likely Pathogenic | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.243 | Likely Benign | -1.69 | Neutral | 0.998 | Probably Damaging | 0.993 | Probably Damaging | 4.08 | Benign | 0.00 | Affected | 0.0678 | 0.5071 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.158G>T | G53V 2D AIThe SynGAP1 missense variant G53V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of predictions (six pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.268042 | Structured | 0.460894 | Uncertain | 0.386 | 0.666 | 0.000 | -8.308 | Likely Pathogenic | 0.959 | Likely Pathogenic | Likely Pathogenic | 0.238 | Likely Benign | -1.71 | Neutral | 0.994 | Probably Damaging | 0.990 | Probably Damaging | 4.11 | Benign | 0.00 | Affected | 0.1193 | 0.4833 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.1598C>A | A533E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A533E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.026324 | Uncertain | 0.843 | 0.393 | 0.000 | -10.810 | Likely Pathogenic | 0.553 | Ambiguous | Likely Benign | -0.13 | Likely Benign | 0.0 | -0.26 | Likely Benign | -0.20 | Likely Benign | 0.43 | Likely Benign | 0.314 | Likely Benign | -1.89 | Neutral | 0.259 | Benign | 0.107 | Benign | -1.18 | Pathogenic | 0.18 | Tolerated | 0.1147 | 0.1498 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||
| c.1601C>G | S534C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S534C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include REVEL, FoldX, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is uncertain, so both are treated as unavailable. No other high‑accuracy predictions are available. Overall, the evidence is evenly split between benign and pathogenic predictions, leaving the variant’s clinical significance inconclusive. There is no ClinVar status to contradict this balanced prediction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.167087 | Structured | 0.032173 | Uncertain | 0.860 | 0.362 | 0.000 | -8.077 | Likely Pathogenic | 0.247 | Likely Benign | Likely Benign | 0.18 | Likely Benign | 0.1 | 0.90 | Ambiguous | 0.54 | Ambiguous | 0.56 | Ambiguous | 0.308 | Likely Benign | -4.05 | Deleterious | 0.998 | Probably Damaging | 0.997 | Probably Damaging | 3.25 | Benign | 0.00 | Affected | 0.0984 | 0.4861 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.1601C>T | S534F 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S534F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FoldX, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. Tools with uncertain results are AlphaMissense‑Default and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic. Overall, the majority of predictions lean toward a benign effect, and this is consistent with the lack of ClinVar evidence; thus the variant is most likely benign and does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.167087 | Structured | 0.032173 | Uncertain | 0.860 | 0.362 | 0.000 | -10.948 | Likely Pathogenic | 0.492 | Ambiguous | Likely Benign | -0.03 | Likely Benign | 0.1 | 0.88 | Ambiguous | 0.43 | Likely Benign | 0.43 | Likely Benign | 0.313 | Likely Benign | -5.09 | Deleterious | 0.998 | Probably Damaging | 0.998 | Probably Damaging | 3.26 | Benign | 0.00 | Affected | 0.0621 | 0.5379 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||
| c.1603A>T | S535C 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant S535C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include Rosetta, Foldetta, premPS, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. Two tools, FoldX and ESM1b, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of predictions lean toward a benign effect, and this conclusion is not contradicted by ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.236433 | Structured | 0.041365 | Uncertain | 0.918 | 0.343 | 0.000 | -7.526 | In-Between | 0.165 | Likely Benign | Likely Benign | 0.57 | Ambiguous | 0.1 | 0.25 | Likely Benign | 0.41 | Likely Benign | 0.47 | Likely Benign | 0.500 | Likely Pathogenic | -2.96 | Deleterious | 0.933 | Possibly Damaging | 0.419 | Benign | -1.33 | Pathogenic | 0.02 | Affected | 0.1139 | 0.5633 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.1604G>T | S535I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S535I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a benign outcome. Overall, the preponderance of evidence points to a benign impact for S535I, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.236433 | Structured | 0.041365 | Uncertain | 0.918 | 0.343 | 0.000 | -8.073 | Likely Pathogenic | 0.330 | Likely Benign | Likely Benign | 0.50 | Ambiguous | 0.0 | -0.42 | Likely Benign | 0.04 | Likely Benign | 0.14 | Likely Benign | 0.246 | Likely Benign | -2.49 | Neutral | 0.004 | Benign | 0.004 | Benign | -1.25 | Pathogenic | 0.07 | Tolerated | 0.1123 | 0.5571 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||
| c.1606T>A | L536M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L536M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.137348 | Structured | 0.042188 | Uncertain | 0.931 | 0.341 | 0.000 | -10.124 | Likely Pathogenic | 0.462 | Ambiguous | Likely Benign | 0.37 | Likely Benign | 0.1 | 1.18 | Ambiguous | 0.78 | Ambiguous | 0.93 | Ambiguous | 0.571 | Likely Pathogenic | -1.94 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.32 | Pathogenic | 0.01 | Affected | 0.0937 | 0.3299 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.160A>T | N54Y 2D AIThe SynGAP1 missense variant N54Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2). High‑accuracy methods show AlphaMissense‑Optimized as benign; the SGM Consensus is unavailable, and Foldetta results are not provided, so its stability prediction is also unavailable. Overall, more tools (five) predict pathogenicity than benign (four), and the high‑accuracy benign prediction is outweighed by the majority of pathogenic calls. Thus, the variant is most likely pathogenic based on current computational predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -9.313 | Likely Pathogenic | 0.640 | Likely Pathogenic | Likely Benign | 0.183 | Likely Benign | -1.57 | Neutral | 0.943 | Possibly Damaging | 0.924 | Probably Damaging | 4.16 | Benign | 0.00 | Affected | 0.0512 | 0.6687 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||||||||||||
| c.1610C>A | A537E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A537E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or stability result is missing or inconclusive. Based on the overall consensus of the available tools, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.116183 | Structured | 0.037313 | Uncertain | 0.928 | 0.362 | 0.000 | -5.120 | Likely Benign | 0.450 | Ambiguous | Likely Benign | -0.14 | Likely Benign | 0.1 | 0.00 | Likely Benign | -0.07 | Likely Benign | 0.23 | Likely Benign | 0.378 | Likely Benign | -1.78 | Neutral | 0.987 | Probably Damaging | 0.838 | Possibly Damaging | -1.04 | Pathogenic | 1.00 | Tolerated | 0.1347 | 0.1566 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||
| c.1612G>C | E538Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E538Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence supports a benign impact. This conclusion does not contradict ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.122885 | Structured | 0.033501 | Uncertain | 0.938 | 0.359 | 0.000 | -10.380 | Likely Pathogenic | 0.733 | Likely Pathogenic | Likely Benign | 0.07 | Likely Benign | 0.0 | 0.07 | Likely Benign | 0.07 | Likely Benign | -0.08 | Likely Benign | 0.188 | Likely Benign | -2.14 | Neutral | 0.890 | Possibly Damaging | 0.436 | Benign | 3.37 | Benign | 0.11 | Tolerated | 0.1127 | 0.3952 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.161A>T | N54I 2D  AIThe SynGAP1 missense variant N54I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -9.919 | Likely Pathogenic | 0.890 | Likely Pathogenic | Ambiguous | 0.201 | Likely Benign | -1.70 | Neutral | 0.943 | Possibly Damaging | 0.924 | Probably Damaging | 4.15 | Benign | 0.00 | Affected | 0.0637 | 0.6793 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.1621G>A | A541T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A541T missense variant is not listed in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that report a benign effect include Rosetta, Foldetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Two tools (FoldX and AlphaMissense‑Default) returned uncertain results. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized predicts benign; Foldetta predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans pathogenic. Overall, the majority of tools (six benign vs. five pathogenic) suggest a benign impact, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.050641 | Structured | 0.029947 | Uncertain | 0.955 | 0.365 | 0.000 | -8.555 | Likely Pathogenic | 0.541 | Ambiguous | Likely Benign | 0.52 | Ambiguous | 0.0 | -0.07 | Likely Benign | 0.23 | Likely Benign | 0.45 | Likely Benign | 0.500 | Likely Pathogenic | -2.02 | Neutral | 0.998 | Probably Damaging | 0.993 | Probably Damaging | -1.28 | Pathogenic | 0.15 | Tolerated | 0.1088 | 0.4164 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||
| c.1622C>G | A541G 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A541G is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438865‑C‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default, and ESM1b) return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is also inconclusive. Overall, the balance of evidence leans toward a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.050641 | Structured | 0.029947 | Uncertain | 0.955 | 0.365 | 0.000 | Uncertain | 1 | 6-33438865-C-G | 2 | 1.24e-6 | -7.233 | In-Between | 0.341 | Ambiguous | Likely Benign | 0.67 | Ambiguous | 0.0 | 0.94 | Ambiguous | 0.81 | Ambiguous | 0.76 | Ambiguous | 0.421 | Likely Benign | -1.48 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | -1.31 | Pathogenic | 0.57 | Tolerated | 3.37 | 35 | 0.1787 | 0.2428 | 1 | 0 | -2.2 | -14.03 | 170.1 | 23.6 | 0.0 | 0.0 | 0.0 | 0.0 | X | Potentially Pathogenic | Ala541 is located on the outer surface of an α-helix (res. Ala533-Val560). The methyl group of Ala541 is on the surface and does not form any interactions. Glycine, known as an “α-helix breaker,” weakens the integrity of the helix. Indeed, in the variant simulations, the hydrogen bond formation between Gly541 and the backbone carbonyl of Ala537 is disrupted. | ||||||||||||||
| c.162C>A | N54K 2D AIThe SynGAP1 missense variant N54K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign) and is treated as unavailable. High‑accuracy methods show AlphaMissense‑Optimized as benign; Foldetta results are not provided, so its stability prediction is unavailable. Overall, the balance of evidence (five pathogenic vs. four benign predictions) suggests the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -8.252 | Likely Pathogenic | 0.779 | Likely Pathogenic | Likely Benign | 0.084 | Likely Benign | -0.82 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.23 | Benign | 0.00 | Affected | 0.2010 | 0.5987 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.162C>G | N54K 2D AIThe SynGAP1 missense variant N54K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign) and is treated as unavailable. High‑accuracy methods show AlphaMissense‑Optimized as benign; Foldetta results are not provided, so its stability prediction is unavailable. Overall, the balance of evidence (five pathogenic vs. four benign predictions) suggests the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -8.252 | Likely Pathogenic | 0.779 | Likely Pathogenic | Likely Benign | 0.084 | Likely Benign | -0.82 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.23 | Benign | 0.00 | Affected | 0.2010 | 0.5987 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.1644G>C | E548D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E548D variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, premPS, SIFT, and FATHMM, whereas a separate group predicts pathogenicity: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Predictions that are uncertain or unavailable are FoldX, Rosetta, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a pathogenic verdict (2 pathogenic, 1 benign, 1 uncertain). Foldetta’s stability prediction is unavailable. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status, which simply lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.054297 | Structured | 0.008632 | Uncertain | 0.965 | 0.288 | 0.000 | -7.359 | In-Between | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.74 | Ambiguous | 0.1 | 1.29 | Ambiguous | 1.02 | Ambiguous | 0.32 | Likely Benign | 0.254 | Likely Benign | -2.85 | Deleterious | 0.998 | Probably Damaging | 0.989 | Probably Damaging | 3.51 | Benign | 0.09 | Tolerated | 0.1374 | 0.2524 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1644G>T | E548D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E548D variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, premPS, SIFT, and FATHMM, whereas a separate group predicts a pathogenic effect: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Predictions that are uncertain or unavailable are FoldX, Rosetta, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a pathogenic verdict (2 pathogenic, 1 benign, 1 uncertain). Foldetta’s stability prediction is unavailable. Overall, the majority of evidence points to a pathogenic impact for E548D, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.054297 | Structured | 0.008632 | Uncertain | 0.965 | 0.288 | 0.000 | -7.359 | In-Between | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.74 | Ambiguous | 0.1 | 1.29 | Ambiguous | 1.02 | Ambiguous | 0.32 | Likely Benign | 0.254 | Likely Benign | -2.85 | Deleterious | 0.998 | Probably Damaging | 0.989 | Probably Damaging | 3.51 | Benign | 0.09 | Tolerated | 0.1374 | 0.2524 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.164A>C | Q55P 2D  AIThe SynGAP1 missense variant Q55P is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33423573‑A‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs two benign), and Foldetta results are unavailable. Overall, more tools predict pathogenicity than benignity, and no ClinVar entry contradicts this assessment. Therefore, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.332115 | Structured | 0.470108 | Uncertain | 0.461 | 0.657 | 0.000 | 6-33423573-A-C | 1 | 6.20e-7 | -13.163 | Likely Pathogenic | 0.897 | Likely Pathogenic | Ambiguous | 0.260 | Likely Benign | -2.06 | Neutral | 0.462 | Possibly Damaging | 0.480 | Possibly Damaging | 3.83 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2557 | 0.5508 | -1 | 0 | 1.9 | -31.01 | |||||||||||||||||||||||||||||||||||
| c.1651C>A | L551M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L551M is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33438894‑C‑A). Prediction tools that classify the variant as benign include FoldX, Rosetta, Foldetta, PROVEAN, SIFT, and AlphaMissense‑Optimized, while those that predict pathogenicity are REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Two tools report an uncertain outcome: premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of predictions lean toward a benign effect, and this does not contradict the ClinVar “Uncertain” classification. Thus, the variant is most likely benign based on the current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.009977 | Structured | 0.006653 | Uncertain | 0.960 | 0.254 | 0.000 | Uncertain | 1 | 6-33438894-C-A | 7 | 4.34e-6 | -9.937 | Likely Pathogenic | 0.480 | Ambiguous | Likely Benign | -0.07 | Likely Benign | 0.1 | 0.13 | Likely Benign | 0.03 | Likely Benign | 0.71 | Ambiguous | 0.544 | Likely Pathogenic | -0.56 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.48 | Pathogenic | 0.06 | Tolerated | 3.37 | 35 | 0.0838 | 0.2701 | 4 | 2 | -1.9 | 18.03 | 246.5 | -18.6 | 0.0 | 0.0 | 0.3 | 0.0 | X | Potentially Benign | L551 is located on an α-helix (res. Ala533-Val560). The iso-butyl side chain of Leu551 hydrophobically packs with nearby hydrophobic residues such as Cys547, Phe652, Leu633, and Ile630 in the inter-helix space. In the variant simulations, the thioether side chain of Met551 can maintain similar hydrophobic interactions as Leu551 in the WT, thus causing no negative effect on the protein structure during the simulations. | ||||||||||||||
| c.1651C>G | L551V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L551V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Two tools give uncertain results: AlphaMissense‑Default and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic. Foldetta’s stability prediction is uncertain. Overall, the majority of evidence points to a pathogenic impact for L551V. This conclusion does not contradict ClinVar status, as the variant is currently unreported in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.009977 | Structured | 0.006653 | Uncertain | 0.960 | 0.254 | 0.000 | -10.154 | Likely Pathogenic | 0.556 | Ambiguous | Likely Benign | 2.04 | Destabilizing | 0.0 | 1.41 | Ambiguous | 1.73 | Ambiguous | 1.03 | Destabilizing | 0.575 | Likely Pathogenic | -1.39 | Neutral | 0.998 | Probably Damaging | 0.992 | Probably Damaging | -1.48 | Pathogenic | 0.22 | Tolerated | 0.1376 | 0.2778 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||
| c.1660G>A | V554M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V554M is not reported in ClinVar but is present in gnomAD (ID 6‑33438903‑G‑A). Functional prediction tools show a split opinion: benign calls come from REVEL, Rosetta, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default; FoldX, Foldetta, and premPS are inconclusive. High‑accuracy assessment focuses on AlphaMissense‑Optimized, the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta. AlphaMissense‑Optimized predicts benign, the SGM Consensus is a tie and thus unavailable, and Foldetta is uncertain, so it is treated as unavailable. Overall, the available evidence leans toward a benign effect, and this does not contradict ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.020522 | Structured | 0.007349 | Uncertain | 0.955 | 0.226 | 0.000 | 6-33438903-G-A | 1 | 6.20e-7 | -8.118 | Likely Pathogenic | 0.671 | Likely Pathogenic | Likely Benign | -1.11 | Ambiguous | 0.0 | -0.20 | Likely Benign | -0.66 | Ambiguous | 0.73 | Ambiguous | 0.217 | Likely Benign | -2.26 | Neutral | 0.994 | Probably Damaging | 0.867 | Possibly Damaging | 3.22 | Benign | 0.00 | Affected | 3.37 | 35 | 0.0743 | 0.2810 | 1 | 2 | -2.3 | 32.06 | |||||||||||||||||||||||||
| c.1660G>C | V554L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V554L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome, while FoldX, premPS, and ESM1b are uncertain. High‑accuracy assessments reinforce the benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the variant is most likely benign based on the available computational evidence, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.020522 | Structured | 0.007349 | Uncertain | 0.955 | 0.226 | 0.000 | -7.884 | In-Between | 0.645 | Likely Pathogenic | Likely Benign | -1.14 | Ambiguous | 0.0 | 0.29 | Likely Benign | -0.43 | Likely Benign | 0.56 | Ambiguous | 0.162 | Likely Benign | -1.54 | Neutral | 0.204 | Benign | 0.053 | Benign | 3.49 | Benign | 0.08 | Tolerated | 0.0991 | 0.2998 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1660G>T | V554L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V554L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome, while FoldX, premPS, and ESM1b are uncertain. High‑accuracy assessments reinforce the benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the variant is most likely benign based on the available computational evidence, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.020522 | Structured | 0.007349 | Uncertain | 0.955 | 0.226 | 0.000 | -7.884 | In-Between | 0.645 | Likely Pathogenic | Likely Benign | -1.14 | Ambiguous | 0.0 | 0.29 | Likely Benign | -0.43 | Likely Benign | 0.56 | Ambiguous | 0.162 | Likely Benign | -1.54 | Neutral | 0.204 | Benign | 0.053 | Benign | 3.49 | Benign | 0.08 | Tolerated | 0.0991 | 0.2998 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1661T>G | V554G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V554G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM, while the remaining eleven tools—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as pathogenic. No predictions are missing or inconclusive. Overall, the variant is most likely pathogenic based on the collective evidence, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.020522 | Structured | 0.007349 | Uncertain | 0.955 | 0.226 | 0.000 | -13.879 | Likely Pathogenic | 0.942 | Likely Pathogenic | Ambiguous | 3.31 | Destabilizing | 0.1 | 4.13 | Destabilizing | 3.72 | Destabilizing | 2.40 | Destabilizing | 0.594 | Likely Pathogenic | -6.96 | Deleterious | 0.997 | Probably Damaging | 1.000 | Probably Damaging | 3.21 | Benign | 0.00 | Affected | 0.1619 | 0.1548 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1663G>C | V555L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V555L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The remaining tools—FoldX, Foldetta, ESM1b, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is unavailable due to a tie between pathogenic and benign calls, and Foldetta is uncertain. Overall, the preponderance of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.013265 | Structured | 0.008218 | Uncertain | 0.943 | 0.225 | 0.000 | -7.816 | In-Between | 0.521 | Ambiguous | Likely Benign | -1.43 | Ambiguous | 0.1 | -0.14 | Likely Benign | -0.79 | Ambiguous | 0.16 | Likely Benign | 0.260 | Likely Benign | -0.97 | Neutral | 0.025 | Benign | 0.015 | Benign | -1.22 | Pathogenic | 0.14 | Tolerated | 0.1036 | 0.2958 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.1664T>G | V555G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V555G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on pathogenicity include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default; no tool predicts it benign. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. No predictions are inconclusive or missing. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.013265 | Structured | 0.008218 | Uncertain | 0.943 | 0.225 | 0.000 | -13.327 | Likely Pathogenic | 0.899 | Likely Pathogenic | Ambiguous | 2.07 | Destabilizing | 0.1 | 2.22 | Destabilizing | 2.15 | Destabilizing | 1.07 | Destabilizing | 0.798 | Likely Pathogenic | -6.35 | Deleterious | 0.984 | Probably Damaging | 1.000 | Probably Damaging | -1.39 | Pathogenic | 0.01 | Affected | 0.1994 | 0.1677 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1667A>G | N556S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant N556S (ClinVar ID 941099.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33438910‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta predicts a benign effect. No other high‑accuracy or folding‑stability methods provide additional evidence. Overall, the majority of predictions support a benign impact, which does not contradict the ClinVar Uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.015078 | Structured | 0.008655 | Uncertain | 0.925 | 0.225 | 0.000 | Uncertain | 1 | 6-33438910-A-G | 3 | 1.86e-6 | -6.576 | Likely Benign | 0.197 | Likely Benign | Likely Benign | 0.52 | Ambiguous | 0.1 | 0.14 | Likely Benign | 0.33 | Likely Benign | 0.16 | Likely Benign | 0.449 | Likely Benign | -3.60 | Deleterious | 1.000 | Probably Damaging | 0.989 | Probably Damaging | -1.22 | Pathogenic | 0.14 | Tolerated | 3.37 | 35 | 0.2641 | 0.3556 | 1 | 1 | 2.7 | -27.03 | 198.8 | 31.0 | 0.0 | 0.0 | -0.5 | 0.2 | X | Potentially Benign | Asn556 is located on the outer surface of an α-helix (res. Ala533-Val560). The carboxamide group of Asn556 forms hydrogen bonds with nearby residues such as Lys553 and Cys552. It also forms a hydrogen bond with the backbone carbonyl group of Cys552, which weakens the α-helix integrity. In the variant simulations, the hydroxyl group of Ser556 forms a more stable hydrogen bond with the backbone carbonyl oxygen of the same helix residue, Cys552, compared to Asn556 in the WT. Serine has a slightly lower propensity to reside in an α-helix than asparagine, which may exacerbate the negative effect on the α-helix integrity. However, the residue swap does not cause negative structural effects during the simulations. | ||||||||||||||
| c.166C>A | L56M 2D AIThe SynGAP1 missense variant L56M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools, ESM1b and AlphaMissense‑Default, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it contains two benign and two uncertain calls, and Foldetta data are unavailable. Overall, the balance of evidence (four benign versus three pathogenic predictions, with two uncertain) suggests the variant is most likely benign. This conclusion does not contradict ClinVar status, as the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.342579 | Structured | 0.476218 | Uncertain | 0.495 | 0.657 | 0.000 | -7.470 | In-Between | 0.345 | Ambiguous | Likely Benign | 0.109 | Likely Benign | -0.51 | Neutral | 0.824 | Possibly Damaging | 0.910 | Probably Damaging | 3.86 | Benign | 0.00 | Affected | 0.0797 | 0.3574 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.166C>G | L56V 2D AIThe SynGAP1 missense variant L56V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also leans benign (2 benign vs 1 pathogenic, 1 uncertain). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for the L56V variant, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.342579 | Structured | 0.476218 | Uncertain | 0.495 | 0.657 | 0.000 | -8.104 | Likely Pathogenic | 0.471 | Ambiguous | Likely Benign | 0.123 | Likely Benign | -0.99 | Neutral | 0.458 | Possibly Damaging | 0.745 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.1635 | 0.3655 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.1672C>T | H558Y 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 H558Y missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while FoldX, premPS, and ESM1b are uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign result; and Foldetta also predicts benign. No prediction or folding stability result is missing or inconclusive. Overall, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.033407 | Structured | 0.011039 | Uncertain | 0.897 | 0.200 | 0.000 | -7.208 | In-Between | 0.187 | Likely Benign | Likely Benign | -0.58 | Ambiguous | 0.0 | -0.36 | Likely Benign | -0.47 | Likely Benign | -0.56 | Ambiguous | 0.359 | Likely Benign | 0.55 | Neutral | 0.019 | Benign | 0.013 | Benign | -1.24 | Pathogenic | 1.00 | Tolerated | 0.0938 | 0.2363 | 0 | 2 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||
| c.1678G>A | V560M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V560M missense variant is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6-33440730-G-A). Functional prediction tools that agree on a benign effect include FoldX, Foldetta, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Uncertain predictions come from Rosetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of high‑confidence tools predict a benign impact, with only one consensus pathogenic prediction. Therefore, the variant is most likely benign based on current computational evidence, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | Uncertain | 2 | 6-33440730-G-A | 15 | 9.50e-6 | -9.598 | Likely Pathogenic | 0.517 | Ambiguous | Likely Benign | -0.33 | Likely Benign | 0.1 | 0.88 | Ambiguous | 0.28 | Likely Benign | 0.72 | Ambiguous | 0.520 | Likely Pathogenic | -2.42 | Neutral | 0.999 | Probably Damaging | 0.863 | Possibly Damaging | -1.25 | Pathogenic | 0.14 | Tolerated | 3.37 | 35 | 0.1161 | 0.3980 | 2 | 1 | -2.3 | 32.06 | 234.9 | -52.6 | 0.0 | 0.0 | -0.1 | 0.1 | X | Potentially Benign | Val560 is located on the surface at the end of an α-helix (res. Ala533-Val560). The iso-propyl group of Val560 favorably packs against Asp508 of the opposing α-helix (res. Gln503-Glu519). However, in the variant simulations, the bulkier thioether side chain of Met560 does not form equally favorable inter-helix interactions. Regardless, no negative structural effects are observed during the simulations. | ||||||||||||||
| c.1678G>C | V560L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V560L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Optimized, and Foldetta; pathogenic predictions come from polyPhen‑2 HumDiv, ESM1b, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized predicts benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, also predicts benign. Overall, the bulk of evidence points to a benign effect, but the SGM Consensus and the presence of pathogenic signals from several high‑confidence tools introduce uncertainty. Thus, the variant is most likely benign based on the prevailing predictions, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | -10.191 | Likely Pathogenic | 0.533 | Ambiguous | Likely Benign | -0.44 | Likely Benign | 0.0 | 0.48 | Likely Benign | 0.02 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -2.45 | Neutral | 0.508 | Possibly Damaging | 0.209 | Benign | -1.24 | Pathogenic | 0.40 | Tolerated | 3.37 | 35 | 0.1430 | 0.4162 | 1 | 2 | -0.4 | 14.03 | ||||||||||||||||||||||||||||
| c.1678G>T | V560L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V560L variant has no ClinVar entry (ClinVar status: None) but is catalogued in gnomAD (ID 6‑33440730‑G‑T). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are polyPhen‑2 HumDiv, ESM1b, and FATHMM (polyPhen‑2 HumVar is benign, AlphaMissense‑Default is uncertain). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning pathogenic (2 pathogenic vs 1 benign), and Foldetta indicating a benign stability change. Overall, the majority of conventional tools favor a benign classification, yet the high‑accuracy consensus and Foldetta suggest a pathogenic signal. Based on the most reliable predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | 6-33440730-G-T | 1 | 6.34e-7 | -10.191 | Likely Pathogenic | 0.533 | Ambiguous | Likely Benign | -0.44 | Likely Benign | 0.0 | 0.48 | Likely Benign | 0.02 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -2.45 | Neutral | 0.508 | Possibly Damaging | 0.209 | Benign | -1.24 | Pathogenic | 0.40 | Tolerated | 3.37 | 35 | 0.1430 | 0.4162 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||
| c.1679T>G | V560G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V560G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only SIFT, whereas the majority of tools (REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic impact. Uncertain results come from FoldX, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show that the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, AlphaMissense‑Optimized is uncertain, and Foldetta is uncertain. Overall, the preponderance of evidence indicates that V560G is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | -12.485 | Likely Pathogenic | 0.799 | Likely Pathogenic | Ambiguous | 0.66 | Ambiguous | 0.1 | 2.12 | Destabilizing | 1.39 | Ambiguous | 1.80 | Destabilizing | 0.753 | Likely Pathogenic | -5.87 | Deleterious | 0.981 | Probably Damaging | 1.000 | Probably Damaging | -1.25 | Pathogenic | 0.19 | Tolerated | 0.1738 | 0.2036 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.167T>A | L56Q 2D AIThe SynGAP1 missense variant L56Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs 2 benign), and Foldetta results are unavailable. Overall, the majority of available predictions (5 pathogenic vs 3 benign) indicate a pathogenic impact. There is no ClinVar entry to contradict this assessment, so the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.342579 | Structured | 0.476218 | Uncertain | 0.495 | 0.657 | 0.000 | -11.064 | Likely Pathogenic | 0.926 | Likely Pathogenic | Ambiguous | 0.293 | Likely Benign | -2.03 | Neutral | 0.943 | Possibly Damaging | 0.944 | Probably Damaging | 3.79 | Benign | 0.00 | Affected | 0.1083 | 0.0842 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||||
| c.167T>G | L56R 2D AIThe SynGAP1 missense variant L56R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of available predictions (5 pathogenic vs 3 benign) indicate a likely pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.342579 | Structured | 0.476218 | Uncertain | 0.495 | 0.657 | 0.000 | -10.194 | Likely Pathogenic | 0.918 | Likely Pathogenic | Ambiguous | 0.264 | Likely Benign | -1.92 | Neutral | 0.943 | Possibly Damaging | 0.944 | Probably Damaging | 3.79 | Benign | 0.00 | Affected | 0.1191 | 0.0685 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.1688G>C | R563T 2D  3DClick to see structure in 3D Viewer AISynGAP1 R563T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, and FATHMM, while those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The remaining tools—FoldX, Rosetta, ESM1b, AlphaMissense‑Optimized, and Foldetta—give uncertain or inconclusive results. High‑accuracy methods provide the following: AlphaMissense‑Optimized is unavailable; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic; Foldetta is unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.031987 | Uncertain | 0.876 | 0.209 | 0.000 | -7.088 | In-Between | 0.913 | Likely Pathogenic | Ambiguous | 1.13 | Ambiguous | 0.1 | 0.77 | Ambiguous | 0.95 | Ambiguous | 0.18 | Likely Benign | 0.307 | Likely Benign | -4.77 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.49 | Benign | 0.23 | Tolerated | 0.2115 | 0.3148 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||
| c.1689G>C | R563S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 R563S missense change is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Five tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized) return uncertain or inconclusive results. High‑accuracy assessments are likewise ambiguous: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑to‑2 tie and therefore unavailable; Foldetta also reports an uncertain stability change. Consequently, the evidence does not strongly support either benign or pathogenic status. The variant’s classification is therefore inconclusive and does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.031987 | Uncertain | 0.876 | 0.209 | 0.000 | -6.503 | Likely Benign | 0.949 | Likely Pathogenic | Ambiguous | 1.16 | Ambiguous | 0.1 | 1.46 | Ambiguous | 1.31 | Ambiguous | 0.69 | Ambiguous | 0.251 | Likely Benign | -4.49 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.48 | Benign | 0.23 | Tolerated | 0.3031 | 0.2534 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||
| c.1689G>T | R563S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant R563S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑to‑2 tie, and Foldetta is uncertain. Consequently, the evidence does not strongly favor either outcome. Based on the current predictions, the variant is most likely benign, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.031987 | Uncertain | 0.876 | 0.209 | 0.000 | -6.503 | Likely Benign | 0.949 | Likely Pathogenic | Ambiguous | 1.16 | Ambiguous | 0.1 | 1.46 | Ambiguous | 1.31 | Ambiguous | 0.69 | Ambiguous | 0.251 | Likely Benign | -4.49 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.48 | Benign | 0.23 | Tolerated | 0.3031 | 0.2534 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||
| c.1693C>A | L565M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L565M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Two tools, Rosetta and premPS, give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it is a 2‑vs‑2 split. No prediction or folding stability result is missing; the only unavailable result is the SGM Consensus. Overall, the balance of evidence (five benign vs four pathogenic predictions, with two high‑accuracy tools supporting benign) indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.034884 | Structured | 0.045819 | Uncertain | 0.922 | 0.205 | 0.000 | -10.346 | Likely Pathogenic | 0.666 | Likely Pathogenic | Likely Benign | 0.24 | Likely Benign | 0.1 | 0.51 | Ambiguous | 0.38 | Likely Benign | 0.69 | Ambiguous | 0.255 | Likely Benign | -1.99 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.86 | Benign | 0.08 | Tolerated | 0.0934 | 0.2613 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.1697A>G | K566R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K566R missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include premPS, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. FoldX, Rosetta, and Foldetta give uncertain or inconclusive results. High‑accuracy methods show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is unavailable. Overall, predictions are split evenly between benign and pathogenic, with no clear consensus. The variant is therefore most likely of uncertain significance; this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.027463 | Structured | 0.047887 | Uncertain | 0.924 | 0.219 | 0.000 | -8.493 | Likely Pathogenic | 0.279 | Likely Benign | Likely Benign | 1.26 | Ambiguous | 0.5 | 1.04 | Ambiguous | 1.15 | Ambiguous | 0.29 | Likely Benign | 0.555 | Likely Pathogenic | -2.39 | Neutral | 0.996 | Probably Damaging | 0.990 | Probably Damaging | -0.79 | Pathogenic | 0.43 | Tolerated | 0.4093 | 0.1099 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.1708G>T | A570S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A570S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The remaining tools (FoldX, Rosetta, Foldetta, premPS, and ESM1b) yield uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors benign, while Foldetta remains uncertain. Overall, the majority of reliable predictions indicate a benign effect. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.046336 | Structured | 0.054494 | Uncertain | 0.932 | 0.263 | 0.000 | -7.893 | In-Between | 0.194 | Likely Benign | Likely Benign | 0.77 | Ambiguous | 0.1 | 1.68 | Ambiguous | 1.23 | Ambiguous | 0.51 | Ambiguous | 0.399 | Likely Benign | -2.26 | Neutral | 0.983 | Probably Damaging | 0.993 | Probably Damaging | -1.19 | Pathogenic | 0.17 | Tolerated | 0.2091 | 0.3256 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.1709C>G | A570G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A570G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include SIFT and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. The remaining tools (FoldX, premPS, ESM1b, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and Foldetta are unavailable due to mixed or uncertain outputs. Overall, the majority of evaluated tools (seven pathogenic vs. two benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.046336 | Structured | 0.054494 | Uncertain | 0.932 | 0.263 | 0.000 | -7.509 | In-Between | 0.562 | Ambiguous | Likely Benign | 1.34 | Ambiguous | 0.1 | 2.12 | Destabilizing | 1.73 | Ambiguous | 0.99 | Ambiguous | 0.607 | Likely Pathogenic | -3.62 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | -1.30 | Pathogenic | 0.09 | Tolerated | 0.1700 | 0.2499 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||
| c.170T>C | L57P 2D  AIThe SynGAP1 missense variant L57P is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (six pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.254060 | Structured | 0.481044 | Uncertain | 0.554 | 0.642 | 0.000 | -10.724 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.242 | Likely Benign | -1.77 | Neutral | 0.943 | Possibly Damaging | 0.944 | Probably Damaging | 3.90 | Benign | 0.00 | Affected | 0.3651 | 0.1703 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.1711T>G | S571A 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant S571A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include FoldX, Rosetta, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑2 split. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta (combining FoldX‑MD and Rosetta outputs) as benign, and the SGM Consensus remains unavailable. Overall, the preponderance of evidence points to a benign impact for S571A, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.045569 | Uncertain | 0.928 | 0.270 | 0.000 | -6.344 | Likely Benign | 0.233 | Likely Benign | Likely Benign | -0.44 | Likely Benign | 0.1 | -0.19 | Likely Benign | -0.32 | Likely Benign | 0.51 | Ambiguous | 0.563 | Likely Pathogenic | -2.69 | Deleterious | 0.980 | Probably Damaging | 0.994 | Probably Damaging | -1.22 | Pathogenic | 0.09 | Tolerated | 0.4739 | 0.2671 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||
| c.1720C>A | L574M 2D AIThe SynGAP1 missense variant L574M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as benign. No predictions or stability results are missing or inconclusive. Based on the overall consensus, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.083462 | Structured | 0.026427 | Uncertain | 0.927 | 0.246 | 0.000 | -7.195 | In-Between | 0.098 | Likely Benign | Likely Benign | 0.14 | Likely Benign | 0.2 | 0.34 | Likely Benign | 0.24 | Likely Benign | -0.09 | Likely Benign | 0.113 | Likely Benign | 0.85 | Neutral | 0.691 | Possibly Damaging | 0.278 | Benign | -1.29 | Pathogenic | 0.11 | Tolerated | 0.0894 | 0.3087 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.1721T>G | L574R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L574R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and FATHMM. Two tools remain inconclusive: Rosetta (Uncertain) and AlphaMissense‑Default (Uncertain). High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of individual predictors (seven benign vs. three pathogenic) support a benign classification, and this conclusion does not contradict the ClinVar status, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.083462 | Structured | 0.026427 | Uncertain | 0.927 | 0.246 | 0.000 | -8.702 | Likely Pathogenic | 0.563 | Ambiguous | Likely Benign | 0.04 | Likely Benign | 0.2 | 0.88 | Ambiguous | 0.46 | Likely Benign | -0.12 | Likely Benign | 0.322 | Likely Benign | 0.30 | Neutral | 0.907 | Possibly Damaging | 0.292 | Benign | -1.22 | Pathogenic | 0.52 | Tolerated | 0.1419 | 0.0530 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||
| c.1724G>A | R575H 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant R575H (ClinVar ID 1029088.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33440776‑G‑A). Prediction tools that indicate a benign effect include Rosetta, Foldetta, PROVEAN, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, Foldetta as Benign, and the SGM Consensus as Pathogenic. Overall, the majority of evidence points to a pathogenic impact, which contrasts with the ClinVar designation of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.061840 | Structured | 0.021362 | Uncertain | 0.916 | 0.259 | 0.000 | Conflicting | 4 | 6-33440776-G-A | 204 | 1.27e-4 | -11.142 | Likely Pathogenic | 0.496 | Ambiguous | Likely Benign | 0.81 | Ambiguous | 0.2 | -0.22 | Likely Benign | 0.30 | Likely Benign | 1.31 | Destabilizing | 0.707 | Likely Pathogenic | -2.34 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.33 | Pathogenic | 0.05 | Affected | 3.37 | 35 | 0.2361 | 0.1292 | 2 | 0 | 1.3 | -19.05 | 244.7 | 80.6 | 0.0 | 0.0 | 0.3 | 0.0 | X | Potentially Pathogenic | The guanidinium group of Arg575, located in an α-helix (res. Arg563-Glu578), forms salt bridges with the carboxylate groups of Asp463 and Asp467, and it also hydrogen bonds with the hydroxyl group of Ser466 on an opposing α-helix (res. Ala461-Phe476) in the WT simulations. In the variant simulations, the imidazole ring of His575 (in its neutral epsilon protonated form) cannot form the same salt bridges as the guanidinium group of the non-mutated Arg575. Instead, His575 only forms weak hydrogen bonds with the hydroxyl groups of Ser466 and Ser571. Overall, the residue swap has the potential to substantially affect the tertiary structure assembly during the protein folding process. | ||||||||||||||
| c.1730C>T | A577V 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A577V is catalogued in gnomAD (ID 6‑33440782‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores benign; the SGM consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also reports benign. No prediction or stability result is missing or inconclusive in these key analyses. Consequently, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.113710 | Structured | 0.019074 | Uncertain | 0.913 | 0.239 | 0.000 | 6-33440782-C-T | 2 | 1.24e-6 | -4.265 | Likely Benign | 0.432 | Ambiguous | Likely Benign | 0.69 | Ambiguous | 0.1 | 0.00 | Likely Benign | 0.35 | Likely Benign | 0.16 | Likely Benign | 0.417 | Likely Benign | -2.11 | Neutral | 0.997 | Probably Damaging | 0.976 | Probably Damaging | -1.32 | Pathogenic | 0.26 | Tolerated | 3.37 | 34 | 0.1278 | 0.4790 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||
| c.1732G>C | E578Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E578Q missense variant is not reported in ClinVar and is absent from gnomAD. Consensus from most in silico predictors indicates a benign effect: REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, AlphaMissense‑Optimized, and Foldetta all predict benign. Pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessment shows AlphaMissense‑Optimized as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic. Foldetta, a protein‑folding stability method, also predicts benign. Overall, the majority of tools, including the high‑accuracy AlphaMissense‑Optimized and Foldetta, support a benign classification, and this is consistent with the lack of ClinVar evidence. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -9.771 | Likely Pathogenic | 0.491 | Ambiguous | Likely Benign | 0.01 | Likely Benign | 0.1 | -0.12 | Likely Benign | -0.06 | Likely Benign | -0.16 | Likely Benign | 0.353 | Likely Benign | -1.21 | Neutral | 0.994 | Probably Damaging | 0.986 | Probably Damaging | -1.40 | Pathogenic | 0.36 | Tolerated | 0.1000 | 0.5319 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.1733A>C | E578A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 E578A missense change is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized; pathogenic predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. ESM1b is uncertain. High‑accuracy assessment shows AlphaMissense‑Optimized predicts benign, while the SGM consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) favors pathogenic, and Foldetta also predicts benign. Overall, the majority of conventional tools lean benign, but the high‑accuracy consensus and AlphaMissense‑Optimized suggest a pathogenic effect. Thus, the variant is most likely pathogenic according to the most reliable predictors, and this assessment does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -7.369 | In-Between | 0.604 | Likely Pathogenic | Likely Benign | 0.34 | Likely Benign | 0.1 | -0.48 | Likely Benign | -0.07 | Likely Benign | -0.02 | Likely Benign | 0.525 | Likely Pathogenic | -2.30 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | -1.40 | Pathogenic | 0.44 | Tolerated | 0.3326 | 0.5118 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||
| c.1733A>G | E578G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E578G missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. Four tools give uncertain or inconclusive results (FoldX, Rosetta, Foldetta, ESM1b). High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—predicts pathogenic. Foldetta, a protein‑folding stability method, yields an uncertain result. Overall, the majority of high‑confidence predictions lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -7.680 | In-Between | 0.601 | Likely Pathogenic | Likely Benign | 0.98 | Ambiguous | 0.1 | 0.63 | Ambiguous | 0.81 | Ambiguous | 0.08 | Likely Benign | 0.589 | Likely Pathogenic | -2.40 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.49 | Pathogenic | 0.19 | Tolerated | 0.2608 | 0.4843 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||
| c.1734G>C | E578D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E578D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) leans toward benign (2 benign vs 1 pathogenic votes), and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -4.366 | Likely Benign | 0.447 | Ambiguous | Likely Benign | 0.46 | Likely Benign | 0.1 | 0.15 | Likely Benign | 0.31 | Likely Benign | -0.05 | Likely Benign | 0.318 | Likely Benign | -0.53 | Neutral | 0.989 | Probably Damaging | 0.979 | Probably Damaging | -1.43 | Pathogenic | 0.33 | Tolerated | 0.1561 | 0.3554 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1734G>T | E578D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E578D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) resolves to benign (2 benign vs 1 pathogenic votes), and Foldetta predicts a benign stability change. No prediction or stability result is missing or inconclusive. Overall, the majority of evidence indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -4.366 | Likely Benign | 0.447 | Ambiguous | Likely Benign | 0.46 | Likely Benign | 0.1 | 0.15 | Likely Benign | 0.31 | Likely Benign | -0.05 | Likely Benign | 0.318 | Likely Benign | -0.53 | Neutral | 0.989 | Probably Damaging | 0.979 | Probably Damaging | -1.43 | Pathogenic | 0.33 | Tolerated | 0.1561 | 0.3554 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1744G>A | E582K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 E582K missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a benign effect on protein stability. Taken together, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is available). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -11.826 | Likely Pathogenic | 0.814 | Likely Pathogenic | Ambiguous | 0.20 | Likely Benign | 0.1 | 0.07 | Likely Benign | 0.14 | Likely Benign | 0.02 | Likely Benign | 0.168 | Likely Benign | -1.83 | Neutral | 0.994 | Probably Damaging | 0.994 | Probably Damaging | 3.31 | Benign | 0.33 | Tolerated | 0.2038 | 0.3762 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||
| c.1745A>C | E582A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E582A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Two tools (FoldX and ESM1b) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -7.432 | In-Between | 0.661 | Likely Pathogenic | Likely Benign | 0.78 | Ambiguous | 0.2 | 0.15 | Likely Benign | 0.47 | Likely Benign | 0.27 | Likely Benign | 0.263 | Likely Benign | -2.99 | Deleterious | 0.998 | Probably Damaging | 0.999 | Probably Damaging | 3.19 | Benign | 0.26 | Tolerated | 0.3236 | 0.4000 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||
| c.1746G>C | E582D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E582D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two consensus groups: a benign group comprising REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized, and a pathogenic group containing polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Remaining tools (FoldX, Rosetta, ESM1b, AlphaMissense‑Default) yield uncertain results and are treated as unavailable. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans toward benign, and Foldetta indicates no significant destabilization. Consequently, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -7.974 | In-Between | 0.520 | Ambiguous | Likely Benign | 0.57 | Ambiguous | 0.2 | 0.95 | Ambiguous | 0.76 | Ambiguous | 0.40 | Likely Benign | 0.093 | Likely Benign | -1.63 | Neutral | 0.986 | Probably Damaging | 0.989 | Probably Damaging | 3.22 | Benign | 0.27 | Tolerated | 0.1481 | 0.2236 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1746G>T | E582D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E582D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two consensus groups: a benign group comprising REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized, and a pathogenic group consisting of polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Remaining tools (FoldX, Rosetta, ESM1b, AlphaMissense‑Default) yield uncertain results. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because two of the four inputs are uncertain; Foldetta, which integrates FoldX‑MD and Rosetta outputs, also reports an uncertain stability change. Consequently, the preponderance of evidence points to a benign effect. This conclusion aligns with the absence of a ClinVar assertion, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -7.974 | In-Between | 0.520 | Ambiguous | Likely Benign | 0.57 | Ambiguous | 0.2 | 0.95 | Ambiguous | 0.76 | Ambiguous | 0.40 | Likely Benign | 0.093 | Likely Benign | -1.63 | Neutral | 0.986 | Probably Damaging | 0.989 | Probably Damaging | 3.22 | Benign | 0.27 | Tolerated | 0.1481 | 0.2236 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1750A>C | I584L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I584L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM, while premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the preponderance of evidence points to a benign impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.046673 | Uncertain | 0.846 | 0.244 | 0.000 | -8.266 | Likely Pathogenic | 0.285 | Likely Benign | Likely Benign | -0.18 | Likely Benign | 0.1 | -0.30 | Likely Benign | -0.24 | Likely Benign | 0.84 | Ambiguous | 0.420 | Likely Benign | -1.74 | Neutral | 0.008 | Benign | 0.046 | Benign | -1.23 | Pathogenic | 0.18 | Tolerated | 0.0927 | 0.2817 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1750A>G | I584V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I584V is catalogued in gnomAD (ID 6‑33440802‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are premPS, polyPhen‑2 HumDiv, and FATHMM. Two tools (FoldX and ESM1b) returned uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign (2 benign vs. 1 pathogenic vote), and Foldetta predicts benign stability. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar classification is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.046673 | Uncertain | 0.846 | 0.244 | 0.000 | 6-33440802-A-G | 1 | 6.20e-7 | -7.562 | In-Between | 0.234 | Likely Benign | Likely Benign | 0.67 | Ambiguous | 0.1 | 0.29 | Likely Benign | 0.48 | Likely Benign | 1.16 | Destabilizing | 0.405 | Likely Benign | -0.95 | Neutral | 0.642 | Possibly Damaging | 0.349 | Benign | -1.18 | Pathogenic | 0.18 | Tolerated | 3.37 | 34 | 0.1007 | 0.2659 | 3 | 4 | -0.3 | -14.03 | |||||||||||||||||||||||||
| c.1754C>G | A585G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A585G is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta reports an uncertain stability change. No evidence from these high‑confidence tools supports pathogenicity. Overall, the balance of evidence favors a benign effect for A585G, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.060549 | Structured | 0.055884 | Uncertain | 0.880 | 0.244 | 0.000 | -3.879 | Likely Benign | 0.629 | Likely Pathogenic | Likely Benign | 1.62 | Ambiguous | 0.0 | 1.94 | Ambiguous | 1.78 | Ambiguous | 0.59 | Ambiguous | 0.384 | Likely Benign | -1.16 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | -1.33 | Pathogenic | 0.24 | Tolerated | 0.1724 | 0.2299 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||
| c.1758C>A | D586E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D586E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Optimized, and Foldetta. Those that predict a pathogenic outcome are polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence points to a benign effect. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.060549 | Structured | 0.066018 | Uncertain | 0.866 | 0.241 | 0.000 | -3.233 | Likely Benign | 0.683 | Likely Pathogenic | Likely Benign | -0.42 | Likely Benign | 0.1 | 0.88 | Ambiguous | 0.23 | Likely Benign | 0.38 | Likely Benign | 0.367 | Likely Benign | -0.12 | Neutral | 0.929 | Possibly Damaging | 0.969 | Probably Damaging | -1.20 | Pathogenic | 1.00 | Tolerated | 0.1304 | 0.5126 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||
| c.1758C>G | D586E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D586E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.060549 | Structured | 0.066018 | Uncertain | 0.866 | 0.241 | 0.000 | -3.233 | Likely Benign | 0.683 | Likely Pathogenic | Likely Benign | -0.42 | Likely Benign | 0.1 | 0.88 | Ambiguous | 0.23 | Likely Benign | 0.38 | Likely Benign | 0.367 | Likely Benign | -0.12 | Neutral | 0.929 | Possibly Damaging | 0.969 | Probably Damaging | -1.20 | Pathogenic | 1.00 | Tolerated | 0.1304 | 0.5126 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||
| c.1765A>G | I589V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I589V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), and FATHMM. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta is uncertain. Taken together, the majority of evidence, including the high‑accuracy predictions, points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.018415 | Structured | 0.084536 | Uncertain | 0.927 | 0.214 | 0.000 | -6.966 | Likely Benign | 0.464 | Ambiguous | Likely Benign | 0.98 | Ambiguous | 0.0 | 0.78 | Ambiguous | 0.88 | Ambiguous | 0.96 | Ambiguous | 0.535 | Likely Pathogenic | -1.00 | Neutral | 0.969 | Probably Damaging | 0.960 | Probably Damaging | -1.50 | Pathogenic | 0.31 | Tolerated | 0.1356 | 0.3568 | 4 | 3 | -0.3 | -14.03 | ||||||||||||||||||||||||||||||
| c.1769G>C | S590T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S590T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and ESM1b. Two tools (premPS and AlphaMissense‑Default) return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.022667 | Structured | 0.088943 | Uncertain | 0.918 | 0.199 | 0.000 | -12.472 | Likely Pathogenic | 0.458 | Ambiguous | Likely Benign | 0.23 | Likely Benign | 0.1 | 0.42 | Likely Benign | 0.33 | Likely Benign | 0.64 | Ambiguous | 0.459 | Likely Benign | -2.89 | Deleterious | 0.183 | Benign | 0.050 | Benign | 3.15 | Benign | 0.08 | Tolerated | 0.1420 | 0.5723 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||
| c.176T>C | L59P 2D AIThe SynGAP1 missense variant L59P is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.212910 | Structured | 0.484882 | Uncertain | 0.510 | 0.668 | 0.000 | -7.076 | In-Between | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.362 | Likely Benign | -2.74 | Deleterious | 0.943 | Possibly Damaging | 0.944 | Probably Damaging | 3.25 | Benign | 0.00 | Affected | 0.3787 | 0.1382 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.1777C>A | L593I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L593I missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are FoldX, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Predictions that are uncertain are Rosetta, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign effect. The variant’s predicted benign status does not contradict its ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.009728 | Structured | 0.110534 | Uncertain | 0.941 | 0.151 | 0.000 | -8.617 | Likely Pathogenic | 0.448 | Ambiguous | Likely Benign | 2.16 | Destabilizing | 0.3 | 0.76 | Ambiguous | 1.46 | Ambiguous | 0.39 | Likely Benign | 0.169 | Likely Benign | -1.59 | Neutral | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 3.14 | Benign | 0.17 | Tolerated | 0.1071 | 0.3582 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1777C>G | L593V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L593V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are FoldX, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Uncertain or inconclusive results come from Rosetta, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments: AlphaMissense‑Optimized classifies the variant as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic prediction; Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. Overall, the majority of evidence (six pathogenic vs. four benign) points to a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, so there is no contradiction with existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.009728 | Structured | 0.110534 | Uncertain | 0.941 | 0.151 | 0.000 | -10.327 | Likely Pathogenic | 0.551 | Ambiguous | Likely Benign | 2.81 | Destabilizing | 0.2 | 1.10 | Ambiguous | 1.96 | Ambiguous | 1.39 | Destabilizing | 0.268 | Likely Benign | -2.59 | Deleterious | 0.998 | Probably Damaging | 0.992 | Probably Damaging | 3.04 | Benign | 0.12 | Tolerated | 0.1599 | 0.3577 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||
| c.1783C>A | L595M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L595M is not reported in ClinVar (ClinVar ID: None) and has no entries in gnomAD (gnomAD ID: None). Functional prediction tools show a split consensus: benign predictions come from REVEL, FoldX, Rosetta, PROVEAN, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a benign effect on protein stability. No prediction or folding result is missing; all available outputs are reported. Based on the balanced distribution of benign and pathogenic calls, the variant is most likely benign, but the evidence is conflicting and does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.015344 | Structured | 0.128444 | Uncertain | 0.920 | 0.150 | 0.000 | -11.325 | Likely Pathogenic | 0.840 | Likely Pathogenic | Ambiguous | 0.32 | Likely Benign | 0.0 | 0.41 | Likely Benign | 0.37 | Likely Benign | 0.90 | Ambiguous | 0.387 | Likely Benign | -1.99 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.74 | Benign | 0.02 | Affected | 0.0930 | 0.3140 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.178G>T | D60Y 2D  AIThe SynGAP1 D60Y missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the D60Y variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.284882 | Structured | 0.480942 | Uncertain | 0.521 | 0.676 | 0.000 | -7.748 | In-Between | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.221 | Likely Benign | -2.60 | Deleterious | 0.972 | Probably Damaging | 0.969 | Probably Damaging | 3.90 | Benign | 0.00 | Affected | 0.0517 | 0.7790 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||||
| c.1792C>A | L598I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L598I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar) and ESM1b. The remaining tools (FoldX, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of reliable predictions indicate a benign impact. This conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.007259 | Structured | 0.147872 | Uncertain | 0.953 | 0.154 | 0.000 | -9.396 | Likely Pathogenic | 0.512 | Ambiguous | Likely Benign | 0.80 | Ambiguous | 0.0 | 0.38 | Likely Benign | 0.59 | Ambiguous | 0.78 | Ambiguous | 0.246 | Likely Benign | -1.96 | Neutral | 0.988 | Probably Damaging | 0.910 | Probably Damaging | 3.47 | Benign | 0.10 | Tolerated | 0.0792 | 0.2000 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.179A>T | D60V 2D  AIThe SynGAP1 D60V missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of evaluated predictors (7 of 10) indicate pathogenicity, while only three suggest benignity. Therefore, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.284882 | Structured | 0.480942 | Uncertain | 0.521 | 0.676 | 0.000 | -6.576 | Likely Benign | 0.980 | Likely Pathogenic | Likely Pathogenic | 0.254 | Likely Benign | -2.72 | Deleterious | 0.972 | Probably Damaging | 0.954 | Probably Damaging | 3.91 | Benign | 0.00 | Affected | 0.0765 | 0.7962 | -2 | -3 | 7.7 | -15.96 | ||||||||||||||||||||||||||||||||||||||||
| c.1801G>T | A601S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A601S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The remaining tools—FoldX, Rosetta, Foldetta, and premPS—return uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie and thus inconclusive. Foldetta, which integrates FoldX‑MD and Rosetta outputs, also yields an uncertain result. Overall, more tools (six) predict pathogenicity than benign (three), and no ClinVar evidence contradicts this assessment. Therefore, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.008895 | Structured | 0.174517 | Uncertain | 0.955 | 0.156 | 0.000 | -11.248 | Likely Pathogenic | 0.136 | Likely Benign | Likely Benign | 0.79 | Ambiguous | 0.1 | 1.63 | Ambiguous | 1.21 | Ambiguous | 0.79 | Ambiguous | 0.541 | Likely Pathogenic | -2.99 | Deleterious | 0.983 | Probably Damaging | 0.993 | Probably Damaging | 2.56 | Benign | 0.01 | Affected | 0.2732 | 0.4730 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.1802C>G | A601G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A601G missense variant is listed in gnomAD (ID 6‑33440854‑C‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect are FATHMM and AlphaMissense‑Optimized; those that agree on a pathogenic effect are REVEL, FoldX, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The high‑accuracy consensus methods give a pathogenic verdict: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also pathogenic. AlphaMissense‑Default and premPS are uncertain, and no evidence is available from other folding‑stability tools. Overall, the preponderance of evidence points to a pathogenic impact for A601G, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.008895 | Structured | 0.174517 | Uncertain | 0.955 | 0.156 | 0.000 | 6-33440854-C-G | 1 | 6.19e-7 | -11.772 | Likely Pathogenic | 0.543 | Ambiguous | Likely Benign | 2.03 | Destabilizing | 0.0 | 2.31 | Destabilizing | 2.17 | Destabilizing | 0.94 | Ambiguous | 0.511 | Likely Pathogenic | -3.98 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 2.55 | Benign | 0.01 | Affected | 3.37 | 35 | 0.2337 | 0.4370 | 0 | 1 | -2.2 | -14.03 | |||||||||||||||||||||||||
| c.1804A>C | I602L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I602L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, PROVEAN, and AlphaMissense‑Optimized, whereas a majority of tools predict pathogenicity: REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Three tools (Rosetta, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the balance of evidence leans toward pathogenicity, with no conflict with the ClinVar status because the variant is not yet classified in that database. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.010221 | Structured | 0.186541 | Uncertain | 0.963 | 0.171 | 0.000 | -9.660 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | -0.15 | Likely Benign | 0.1 | 1.12 | Ambiguous | 0.49 | Likely Benign | 0.91 | Ambiguous | 0.631 | Likely Pathogenic | -1.99 | Neutral | 0.645 | Possibly Damaging | 0.718 | Possibly Damaging | -1.54 | Pathogenic | 0.04 | Affected | 0.1044 | 0.3199 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1807A>C | M603L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 M603L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include FoldX, premPS, PROVEAN, SIFT, AlphaMissense‑Optimized, and the protein‑folding stability method Foldetta. Those that predict pathogenicity are REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. Two tools give uncertain results: Rosetta and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, more evidence supports a benign effect (six benign versus five pathogenic predictions, with two uncertain). Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.011342 | Structured | 0.197847 | Uncertain | 0.942 | 0.176 | 0.000 | -7.134 | In-Between | 0.590 | Likely Pathogenic | Likely Benign | 0.04 | Likely Benign | 0.1 | 0.86 | Ambiguous | 0.45 | Likely Benign | -0.18 | Likely Benign | 0.548 | Likely Pathogenic | -2.16 | Neutral | 0.484 | Possibly Damaging | 0.654 | Possibly Damaging | -0.98 | Pathogenic | 0.52 | Tolerated | 0.1345 | 0.3227 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.1807A>T | M603L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 M603L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. Rosetta and ESM1b give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools lean toward a benign interpretation, but the SGM Consensus indicates pathogenicity, leaving the variant’s clinical significance uncertain. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for M603L. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.011342 | Structured | 0.197847 | Uncertain | 0.942 | 0.176 | 0.000 | -7.134 | In-Between | 0.590 | Likely Pathogenic | Likely Benign | 0.04 | Likely Benign | 0.1 | 0.86 | Ambiguous | 0.45 | Likely Benign | -0.18 | Likely Benign | 0.548 | Likely Pathogenic | -2.16 | Neutral | 0.484 | Possibly Damaging | 0.654 | Possibly Damaging | -0.98 | Pathogenic | 0.52 | Tolerated | 0.1345 | 0.3227 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.1817G>C | S606T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S606T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, FATHMM, and AlphaMissense‑Optimized, whereas a pathogenic consensus is reached by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Uncertain results come from Rosetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools predict pathogenicity, and the SGM Consensus supports this view, while the high‑accuracy methods give mixed results. Thus, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.041405 | Structured | 0.191720 | Uncertain | 0.875 | 0.247 | 0.000 | -11.052 | Likely Pathogenic | 0.554 | Ambiguous | Likely Benign | 0.06 | Likely Benign | 0.1 | -0.91 | Ambiguous | -0.43 | Likely Benign | 0.57 | Ambiguous | 0.203 | Likely Benign | -2.99 | Deleterious | 0.826 | Possibly Damaging | 0.933 | Probably Damaging | 3.34 | Benign | 0.03 | Affected | 0.1260 | 0.4513 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||
| c.1826G>A | G609E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G609E has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools (Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.203786 | Uncertain | 0.851 | 0.252 | 0.000 | -10.470 | Likely Pathogenic | 0.421 | Ambiguous | Likely Benign | 2.95 | Destabilizing | 0.4 | -1.65 | Ambiguous | 0.65 | Ambiguous | 0.61 | Ambiguous | 0.531 | Likely Pathogenic | -2.28 | Neutral | 0.916 | Possibly Damaging | 0.588 | Possibly Damaging | -1.41 | Pathogenic | 0.17 | Tolerated | 0.1709 | 0.4491 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1826G>C | G609A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G609A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX, PROVEAN, and FATHMM. One tool, Foldetta, yields an uncertain result. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.203786 | Uncertain | 0.851 | 0.252 | 0.000 | -6.790 | Likely Benign | 0.153 | Likely Benign | Likely Benign | 2.38 | Destabilizing | 0.3 | 0.01 | Likely Benign | 1.20 | Ambiguous | 0.43 | Likely Benign | 0.494 | Likely Benign | -2.65 | Deleterious | 0.282 | Benign | 0.164 | Benign | -1.43 | Pathogenic | 0.10 | Tolerated | 0.3812 | 0.3896 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||
| c.1828C>A | L610I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L610I is listed in gnomAD (ID 6‑33440880‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM. Four tools are uncertain (FoldX, Foldetta, premPS, AlphaMissense‑Default). High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized scores benign, the SGM consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, whereas Foldetta’s stability estimate is unavailable. Overall, the balance of evidence points to a benign effect for L610I, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.271506 | Structured | 0.209504 | Uncertain | 0.888 | 0.253 | 0.000 | 6-33440880-C-A | 1 | 6.19e-7 | -6.362 | Likely Benign | 0.389 | Ambiguous | Likely Benign | 1.50 | Ambiguous | 0.2 | 0.18 | Likely Benign | 0.84 | Ambiguous | 0.76 | Ambiguous | 0.544 | Likely Pathogenic | -1.86 | Neutral | 0.992 | Probably Damaging | 0.997 | Probably Damaging | -1.34 | Pathogenic | 0.15 | Tolerated | 3.37 | 35 | 0.0999 | 0.3219 | 2 | 2 | 0.7 | 0.00 | |||||||||||||||||||||||||
| c.1831A>G | M611V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M611V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—yields an inconclusive result (two benign, two pathogenic). Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports an uncertain effect, so its result is treated as unavailable. Overall, the balance of evidence leans toward a benign impact, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.236433 | Structured | 0.210791 | Uncertain | 0.870 | 0.253 | 0.000 | -8.057 | Likely Pathogenic | 0.176 | Likely Benign | Likely Benign | 1.42 | Ambiguous | 0.4 | 1.56 | Ambiguous | 1.49 | Ambiguous | 0.66 | Ambiguous | 0.315 | Likely Benign | -2.06 | Neutral | 0.960 | Probably Damaging | 0.474 | Possibly Damaging | -1.04 | Pathogenic | 0.49 | Tolerated | 0.2324 | 0.2721 | 2 | 1 | 2.3 | -32.06 | ||||||||||||||||||||||||||||||
| c.1841A>T | Y614F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y614F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Uncertain or unavailable results come from Foldetta, premPS, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is unavailable. Overall, the majority of reliable predictors indicate a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.314870 | Structured | 0.182134 | Uncertain | 0.852 | 0.254 | 0.000 | -5.584 | Likely Benign | 0.630 | Likely Pathogenic | Likely Benign | 0.09 | Likely Benign | 0.2 | 0.98 | Ambiguous | 0.54 | Ambiguous | 0.78 | Ambiguous | 0.364 | Likely Benign | -3.75 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.42 | Benign | 0.07 | Tolerated | 0.1909 | 0.2762 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||
| c.1846G>A | D616N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D616N missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into benign (REVEL, premPS, SIFT, FATHMM, AlphaMissense‑Optimized) and pathogenic (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b). Four tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) favors pathogenic, and Foldetta remains uncertain. Overall, the majority of conventional predictors lean toward a benign effect, whereas the SGM Consensus suggests pathogenicity, leaving the variant’s clinical significance ambiguous. Based on the prevailing evidence, the variant is most likely benign, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.166689 | Uncertain | 0.867 | 0.252 | 0.000 | -8.292 | Likely Pathogenic | 0.349 | Ambiguous | Likely Benign | 0.54 | Ambiguous | 0.2 | 1.05 | Ambiguous | 0.80 | Ambiguous | 0.03 | Likely Benign | 0.149 | Likely Benign | -3.74 | Deleterious | 0.875 | Possibly Damaging | 0.581 | Possibly Damaging | 3.41 | Benign | 0.11 | Tolerated | 0.1053 | 0.3976 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.1847A>G | D616G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 D616G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Tools with uncertain or inconclusive results—FoldX, AlphaMissense‑Default, and Foldetta—are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the predictions are split evenly between benign and pathogenic, with no clear consensus. Thus, the variant is most likely of uncertain significance; it does not contradict any ClinVar status because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.166689 | Uncertain | 0.867 | 0.252 | 0.000 | -10.310 | Likely Pathogenic | 0.547 | Ambiguous | Likely Benign | 1.48 | Ambiguous | 0.1 | 2.13 | Destabilizing | 1.81 | Ambiguous | 0.49 | Likely Benign | 0.144 | Likely Benign | -5.60 | Deleterious | 0.985 | Probably Damaging | 0.800 | Possibly Damaging | 3.37 | Benign | 0.06 | Tolerated | 0.3496 | 0.4386 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||
| c.1848T>A | D616E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D616E missense variant is catalogued in gnomAD (ID 6‑33440900‑T‑A) but has no ClinVar submission. Functional prediction tools show a split assessment: benign calls come from REVEL, both polyPhen‑2 HumDiv and HumVar, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls arise from PROVEAN, SIFT, and AlphaMissense‑Default. The remaining predictors (FoldX, Rosetta, Foldetta, premPS, ESM1b) are inconclusive. A high‑accuracy consensus (SGM) that aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a pathogenic verdict. AlphaMissense‑Optimized remains benign, and Foldetta, which evaluates protein‑folding stability, is uncertain. Overall, the majority of evidence leans toward pathogenicity, and this conclusion does not conflict with ClinVar because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.166689 | Uncertain | 0.867 | 0.252 | 0.000 | 6-33440900-T-A | 1 | 6.20e-7 | -7.250 | In-Between | 0.695 | Likely Pathogenic | Likely Benign | 0.96 | Ambiguous | 0.1 | 1.52 | Ambiguous | 1.24 | Ambiguous | 0.58 | Ambiguous | 0.092 | Likely Benign | -2.85 | Deleterious | 0.421 | Benign | 0.232 | Benign | 3.32 | Benign | 0.03 | Affected | 3.37 | 35 | 0.1225 | 0.4128 | 2 | 3 | 0.0 | 14.03 | |||||||||||||||||||||||||
| c.1848T>G | D616E 2D 3DClick to see structure in 3D Viewer AISynGAP1 D616E is not reported in ClinVar but is present in gnomAD (ID 6‑33440900‑T‑G). Functional prediction tools that agree on benign impact include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, SIFT, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b) are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of conventional predictors lean toward a benign effect, but the high‑accuracy consensus is split, leaving the variant’s clinical significance unresolved. Thus, the variant is most likely benign based on the bulk of predictions, and this does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.166689 | Uncertain | 0.867 | 0.252 | 0.000 | 6-33440900-T-G | 3 | 1.86e-6 | -7.250 | In-Between | 0.695 | Likely Pathogenic | Likely Benign | 0.96 | Ambiguous | 0.1 | 1.52 | Ambiguous | 1.24 | Ambiguous | 0.58 | Ambiguous | 0.092 | Likely Benign | -2.85 | Deleterious | 0.421 | Benign | 0.232 | Benign | 3.32 | Benign | 0.03 | Affected | 3.37 | 35 | 0.1225 | 0.4128 | 2 | 3 | 0.0 | 14.03 | |||||||||||||||||||||||||
| c.1852C>A | Q618K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q618K is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33440904‑C‑A). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while ESM1b is uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign result; and Foldetta also predicts benign stability. No predictions or folding stability results are missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | 6-33440904-C-A | 24 | 1.49e-5 | -7.708 | In-Between | 0.229 | Likely Benign | Likely Benign | 0.02 | Likely Benign | 0.0 | 0.16 | Likely Benign | 0.09 | Likely Benign | -0.46 | Likely Benign | 0.281 | Likely Benign | -0.05 | Neutral | 0.338 | Benign | 0.111 | Benign | -1.21 | Pathogenic | 0.29 | Tolerated | 3.37 | 35 | 0.1413 | 0.2750 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||
| c.1852C>G | Q618E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q618E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the preponderance of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, and there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | -12.535 | Likely Pathogenic | 0.162 | Likely Benign | Likely Benign | 0.46 | Likely Benign | 0.1 | 0.50 | Ambiguous | 0.48 | Likely Benign | 0.33 | Likely Benign | 0.233 | Likely Benign | -1.16 | Neutral | 0.046 | Benign | 0.021 | Benign | -1.17 | Pathogenic | 0.26 | Tolerated | 0.1003 | 0.1469 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1854G>C | Q618H 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q618H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, premPS, PROVEAN, and AlphaMissense‑Optimized; pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Four tools (FoldX, Foldetta, ESM1b, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive and Foldetta is uncertain. Overall, the majority of evidence points toward a benign effect, and this conclusion does not contradict the lack of ClinVar annotation or gnomAD presence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | -7.092 | In-Between | 0.489 | Ambiguous | Likely Benign | 0.59 | Ambiguous | 0.1 | 0.44 | Likely Benign | 0.52 | Ambiguous | 0.34 | Likely Benign | 0.475 | Likely Benign | -1.99 | Neutral | 0.946 | Possibly Damaging | 0.638 | Possibly Damaging | -1.35 | Pathogenic | 0.04 | Affected | 0.1015 | 0.2655 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1854G>T | Q618H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q618H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, premPS, PROVEAN, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Four tools (FoldX, Foldetta, ESM1b, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is also uncertain. Overall, the balance of evidence leans toward a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | -7.092 | In-Between | 0.489 | Ambiguous | Likely Benign | 0.59 | Ambiguous | 0.1 | 0.44 | Likely Benign | 0.52 | Ambiguous | 0.34 | Likely Benign | 0.475 | Likely Benign | -1.99 | Neutral | 0.946 | Possibly Damaging | 0.638 | Possibly Damaging | -1.35 | Pathogenic | 0.04 | Affected | 0.1015 | 0.2655 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.1873C>A | L625I 2D AIThe SynGAP1 missense variant L625I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas a majority of tools (premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. Tools with uncertain or inconclusive results—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—are treated as unavailable. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Consequently, the overall evidence leans toward pathogenicity, with no conflict with ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.229226 | Structured | 0.045896 | Uncertain | 0.966 | 0.215 | 0.000 | -11.713 | Likely Pathogenic | 0.866 | Likely Pathogenic | Ambiguous | 0.75 | Ambiguous | 0.6 | 0.72 | Ambiguous | 0.74 | Ambiguous | 1.09 | Destabilizing | 0.412 | Likely Benign | -1.96 | Neutral | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 3.06 | Benign | 0.01 | Affected | 0.0864 | 0.3588 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1876A>C | I626L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I626L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default predict a pathogenic outcome. Three tools—Foldetta, premPS, and Rosetta—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta also yields an uncertain stability prediction. Overall, the balance of evidence leans toward a benign interpretation, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.109221 | Structured | 0.040732 | Uncertain | 0.970 | 0.223 | 0.000 | -10.696 | Likely Pathogenic | 0.622 | Likely Pathogenic | Likely Benign | 0.32 | Likely Benign | 0.1 | 1.00 | Ambiguous | 0.66 | Ambiguous | 0.83 | Ambiguous | 0.311 | Likely Benign | -1.86 | Neutral | 0.955 | Possibly Damaging | 0.985 | Probably Damaging | 3.52 | Benign | 0.12 | Tolerated | 0.0741 | 0.2461 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1879G>T | A627S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A627S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are Rosetta, PROVEAN, and ESM1b. The remaining tools—FoldX, Foldetta, and premPS—return uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta is uncertain. Overall, the majority of available predictions lean toward a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.100716 | Structured | 0.037862 | Uncertain | 0.970 | 0.210 | 0.000 | -10.782 | Likely Pathogenic | 0.329 | Likely Benign | Likely Benign | 1.11 | Ambiguous | 0.2 | 2.05 | Destabilizing | 1.58 | Ambiguous | 0.71 | Ambiguous | 0.316 | Likely Benign | -2.94 | Deleterious | 0.411 | Benign | 0.387 | Benign | 2.78 | Benign | 0.11 | Tolerated | 0.2266 | 0.4224 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.1885G>A | V629I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V629I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, AlphaMissense‑Optimized, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar) and SIFT, with ESM1b also indicating pathogenicity. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta also predicts a benign outcome. Taken together, the preponderance of evidence supports a benign classification for V629I, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.034796 | Uncertain | 0.970 | 0.236 | 0.000 | -9.876 | Likely Pathogenic | 0.462 | Ambiguous | Likely Benign | -0.22 | Likely Benign | 0.1 | 0.37 | Likely Benign | 0.08 | Likely Benign | 0.21 | Likely Benign | 0.305 | Likely Benign | -1.00 | Neutral | 0.975 | Probably Damaging | 0.958 | Probably Damaging | 3.26 | Benign | 0.04 | Affected | 0.0650 | 0.2919 | 4 | 3 | 0.3 | 14.03 | ||||||||||||||||||||||||||||||
| c.1886T>G | V629G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V629G missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining eleven tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) uniformly predict a pathogenic outcome; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) is pathogenic. Taken together, the overwhelming majority of evidence indicates that V629G is likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.034796 | Uncertain | 0.970 | 0.236 | 0.000 | -13.150 | Likely Pathogenic | 0.871 | Likely Pathogenic | Ambiguous | 3.81 | Destabilizing | 0.0 | 4.52 | Destabilizing | 4.17 | Destabilizing | 1.94 | Destabilizing | 0.678 | Likely Pathogenic | -6.47 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.07 | Benign | 0.00 | Affected | 0.1903 | 0.2240 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1888A>C | I630L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I630L is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33440940‑A‑C). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign; the SGM Consensus remains unavailable. Based on the overall predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.036106 | Uncertain | 0.966 | 0.236 | 0.000 | 6-33440940-A-C | -8.949 | Likely Pathogenic | 0.277 | Likely Benign | Likely Benign | -0.39 | Likely Benign | 0.0 | 0.23 | Likely Benign | -0.08 | Likely Benign | 0.33 | Likely Benign | 0.165 | Likely Benign | -1.30 | Neutral | 0.102 | Benign | 0.108 | Benign | -0.81 | Pathogenic | 0.27 | Tolerated | 3.37 | 34 | 0.0688 | 0.2461 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||
| c.1888A>G | I630V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I630V is listed in ClinVar as Benign and is present in gnomAD (variant ID 6‑33440940‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome; all other tools (FoldX, Rosetta, Foldetta, premPS, ESM1b) return uncertain or inconclusive results. The high‑accuracy consensus (SGM Consensus) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a benign majority (2 benign vs. 1 pathogenic, 1 uncertain). AlphaMissense‑Optimized also predicts benign. Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, is uncertain. Taken together, the overwhelming majority of predictions support a benign effect, and this conclusion aligns with the ClinVar designation. Thus, the variant is most likely benign, with no contradiction to the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.036106 | Uncertain | 0.966 | 0.236 | 0.000 | Benign/Likely benign | 4 | 6-33440940-A-G | 59 | 3.66e-5 | -7.264 | In-Between | 0.145 | Likely Benign | Likely Benign | 1.33 | Ambiguous | 0.0 | 0.94 | Ambiguous | 1.14 | Ambiguous | 0.64 | Ambiguous | 0.143 | Likely Benign | -0.38 | Neutral | 0.018 | Benign | 0.011 | Benign | -1.37 | Pathogenic | 0.35 | Tolerated | 3.37 | 34 | 0.0960 | 0.2891 | 4 | 3 | -0.3 | -14.03 | 235.0 | 26.2 | -0.1 | 0.0 | -0.3 | 0.1 | X | Potentially Benign | The sec-butyl side chain of Ile630, located in an α helix (res. Glu617-Asn635), packs with hydrophobic residues (e.g., Phe594, Leu633, Ile626, Ile602) in the hydrophobic inter-helix space between two α helices (res. Glu617-Asn635 and res. Glu582-Met603).In the variant simulations, the iso-propyl side chain of Val630, which shares a similar size and physicochemical properties with Ile630 in the WT, maintains similar interactions in the inter-helix space. Although no negative structural effects are observed during the simulations, the implications of the residue swap on the complex formation with the GTPase, due to its location, cannot be investigated using solvent-only simulations. | ||||||||||||||
| c.1889T>C | I630T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I630T has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized indicates benign, whereas the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic majority, and Foldetta also predicts pathogenic. No prediction is missing or inconclusive. Overall, the preponderance of evidence points to a pathogenic effect for I630T, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.036106 | Uncertain | 0.966 | 0.236 | 0.000 | -7.780 | In-Between | 0.754 | Likely Pathogenic | Likely Benign | 2.77 | Destabilizing | 0.1 | 2.27 | Destabilizing | 2.52 | Destabilizing | 1.94 | Destabilizing | 0.734 | Likely Pathogenic | -2.15 | Neutral | 0.997 | Probably Damaging | 0.961 | Probably Damaging | -1.46 | Pathogenic | 0.35 | Tolerated | 0.0985 | 0.0640 | 0 | -1 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||
| c.1890C>G | I630M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I630M is listed in gnomAD (ID 6‑33440942‑C‑G) but has no ClinVar entry. Functional prediction tools show a split: benign calls come from Rosetta, Foldetta, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic calls come from REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. FoldX is uncertain, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. High‑accuracy assessments give AlphaMissense‑Optimized benign, Foldetta benign, and an inconclusive SGM Consensus. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.040537 | Structured | 0.036106 | Uncertain | 0.966 | 0.236 | 0.000 | 6-33440942-C-G | 1 | 6.20e-7 | -10.586 | Likely Pathogenic | 0.259 | Likely Benign | Likely Benign | -0.55 | Ambiguous | 0.1 | 0.32 | Likely Benign | -0.12 | Likely Benign | 1.06 | Destabilizing | 0.508 | Likely Pathogenic | -1.90 | Neutral | 0.833 | Possibly Damaging | 0.700 | Possibly Damaging | -1.38 | Pathogenic | 0.02 | Affected | 3.37 | 34 | 0.0618 | 0.1759 | 1 | 2 | -2.6 | 18.03 | |||||||||||||||||||||||||
| c.1895A>C | N632T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N632T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, PROVEAN, and FATHMM. The remaining tools—FoldX, Rosetta, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of predictions lean toward a benign impact, and this does not contradict any ClinVar annotation (none is available). Thus, based on the current computational evidence, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.041437 | Uncertain | 0.938 | 0.254 | 0.000 | -6.797 | Likely Benign | 0.371 | Ambiguous | Likely Benign | 1.14 | Ambiguous | 0.2 | -0.58 | Ambiguous | 0.28 | Likely Benign | 0.39 | Likely Benign | 0.541 | Likely Pathogenic | -4.48 | Deleterious | 0.214 | Benign | 0.062 | Benign | -1.49 | Pathogenic | 0.13 | Tolerated | 0.1295 | 0.6809 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||
| c.1895A>G | N632S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N632S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen2_HumVar. Those that predict a pathogenic effect are PROVEAN, polyPhen2_HumDiv, and FATHMM. The remaining tools—FoldX, Rosetta, Foldetta, and ESM1b—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of available predictions (six benign vs. three pathogenic) support a benign classification, and this does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.041437 | Uncertain | 0.938 | 0.254 | 0.000 | -7.677 | In-Between | 0.291 | Likely Benign | Likely Benign | 0.81 | Ambiguous | 0.1 | 0.78 | Ambiguous | 0.80 | Ambiguous | 0.41 | Likely Benign | 0.469 | Likely Benign | -3.85 | Deleterious | 0.718 | Possibly Damaging | 0.086 | Benign | -1.37 | Pathogenic | 0.12 | Tolerated | 0.3302 | 0.6486 | 1 | 1 | 2.7 | -27.03 | ||||||||||||||||||||||||||||||
| c.1897C>A | L633M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L633M has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Four tools (FoldX, Rosetta, Foldetta, premPS) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, the majority of definitive predictions (5 pathogenic vs. 4 benign) lean toward a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.045352 | Structured | 0.045407 | Uncertain | 0.952 | 0.252 | 0.000 | -10.300 | Likely Pathogenic | 0.718 | Likely Pathogenic | Likely Benign | 0.61 | Ambiguous | 0.1 | 1.47 | Ambiguous | 1.04 | Ambiguous | 0.90 | Ambiguous | 0.355 | Likely Benign | -1.99 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.77 | Benign | 0.01 | Affected | 0.0984 | 0.2683 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.1900G>T | A634S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A634S variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are FATHMM and AlphaMissense‑Optimized; those that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not contradict the ClinVar status, which simply lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.085092 | Structured | 0.052058 | Uncertain | 0.932 | 0.242 | 0.000 | -9.706 | Likely Pathogenic | 0.434 | Ambiguous | Likely Benign | 0.91 | Ambiguous | 0.1 | 1.28 | Ambiguous | 1.10 | Ambiguous | 0.77 | Ambiguous | 0.506 | Likely Pathogenic | -2.99 | Deleterious | 0.953 | Possibly Damaging | 0.985 | Probably Damaging | 2.67 | Benign | 0.05 | Affected | 0.2607 | 0.4231 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.1903A>C | N635H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N635H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Three tools (FoldX, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy methods give the following: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic; and Foldetta predicts benign. No prediction or folding‑stability result is missing or inconclusive. Based on the available evidence, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.060246 | Uncertain | 0.900 | 0.252 | 0.000 | -12.507 | Likely Pathogenic | 0.419 | Ambiguous | Likely Benign | 1.07 | Ambiguous | 0.2 | -0.10 | Likely Benign | 0.49 | Likely Benign | 0.91 | Ambiguous | 0.429 | Likely Benign | -4.78 | Deleterious | 0.993 | Probably Damaging | 0.879 | Possibly Damaging | 2.90 | Benign | 0.00 | Affected | 0.1184 | 0.3720 | 2 | 1 | 0.3 | 23.04 | ||||||||||||||||||||||||||||||
| c.1904A>C | N635T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N635T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and ESM1b; polyPhen‑2 HumVar, however, classifies it as benign. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain, and FoldX and premPS also yield uncertain results. Overall, the majority of evidence (five benign vs. four pathogenic) supports a benign classification. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.060246 | Uncertain | 0.900 | 0.252 | 0.000 | -11.705 | Likely Pathogenic | 0.256 | Likely Benign | Likely Benign | 1.50 | Ambiguous | 0.1 | 0.44 | Likely Benign | 0.97 | Ambiguous | 0.81 | Ambiguous | 0.240 | Likely Benign | -5.58 | Deleterious | 0.536 | Possibly Damaging | 0.184 | Benign | 2.98 | Benign | 0.04 | Affected | 0.1206 | 0.4032 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||
| c.1904A>G | N635S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant N635S is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6-33440956-A-G). Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and ESM1b. Predictions that are inconclusive or unavailable are FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is also inconclusive. Overall, the majority of available predictions lean toward a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.060246 | Uncertain | 0.900 | 0.252 | 0.000 | Conflicting | 4 | 6-33440956-A-G | 10 | 6.20e-6 | -9.002 | Likely Pathogenic | 0.101 | Likely Benign | Likely Benign | 0.80 | Ambiguous | 0.1 | 0.67 | Ambiguous | 0.74 | Ambiguous | 0.95 | Ambiguous | 0.104 | Likely Benign | -4.45 | Deleterious | 0.261 | Benign | 0.044 | Benign | 3.06 | Benign | 0.05 | Affected | 3.37 | 34 | 0.2816 | 0.4279 | 1 | 1 | 2.7 | -27.03 | 196.0 | 30.9 | 0.1 | 0.0 | -0.3 | 0.2 | X | Uncertain | In the WT simulations, the carboxamide side chain of Asn635, located on the outer surface of an α helix (res. Glu617-Asn635), forms hydrogen bonds with Gln631 on the same α helix and with the hydroxyl side chain of Ser590 on an opposing α helix (res. Glu582-Met603).In the variant simulations, the side chain of Ser635 is shorter than asparagine and thus prefers to hydrogen bond with the carbonyl group of Gln631 on the same helix and, to a lesser extent, with Ser590 compared to Asn635 in the WT. Ser635 forms hydrogen bonds with the backbone atoms of the same helix, which may destabilize the helix, although this is not clearly evident in the simulations. The weakening of the hydrogen bond between Ser635 and Ser590 in the variant may also weaken the tertiary structure assembly between the helices.Additionally, Asn635 is at the GTPase interface. However, the implication of the residue swap on the complex formation with the GTPase cannot be investigated using solvent-only simulations. | ||||||||||||||
| c.1910C>G | S637C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S637C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include REVEL, FoldX, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; Foldetta predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence supports a benign impact, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.076542 | Structured | 0.083482 | Uncertain | 0.920 | 0.253 | 0.000 | -10.040 | Likely Pathogenic | 0.138 | Likely Benign | Likely Benign | 0.25 | Likely Benign | 0.0 | 0.54 | Ambiguous | 0.40 | Likely Benign | 0.22 | Likely Benign | 0.169 | Likely Benign | -2.83 | Deleterious | 0.985 | Probably Damaging | 0.533 | Possibly Damaging | 3.34 | Benign | 0.01 | Affected | 0.1327 | 0.4439 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.1912A>C | K638Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K638Q missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.045352 | Structured | 0.098064 | Uncertain | 0.937 | 0.260 | 0.000 | -9.561 | Likely Pathogenic | 0.556 | Ambiguous | Likely Benign | 0.45 | Likely Benign | 0.0 | 0.37 | Likely Benign | 0.41 | Likely Benign | 0.22 | Likely Benign | 0.421 | Likely Benign | -3.60 | Deleterious | 0.997 | Probably Damaging | 0.991 | Probably Damaging | 3.42 | Benign | 0.12 | Tolerated | 0.3623 | 0.0920 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||
| c.1919C>T | T640I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T640I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only PROVEAN predicts a pathogenic outcome. The remaining tools (FoldX, Rosetta, ESM1b, AlphaMissense‑Default, and Foldetta) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is unavailable due to a tie, and Foldetta also yields an uncertain stability change. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.137043 | Uncertain | 0.893 | 0.284 | 0.000 | -7.256 | In-Between | 0.358 | Ambiguous | Likely Benign | 0.70 | Ambiguous | 0.1 | 0.66 | Ambiguous | 0.68 | Ambiguous | 0.28 | Likely Benign | 0.193 | Likely Benign | -2.75 | Deleterious | 0.322 | Benign | 0.022 | Benign | 3.46 | Benign | 0.12 | Tolerated | 0.1030 | 0.4793 | 0 | -1 | 5.2 | 12.05 | ||||||||||||||||||||||||||||||
| c.1921T>C | S641P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S641P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and SIFT. The remaining tools (FoldX, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward benign, and Foldetta as uncertain. Overall, the majority of reliable predictions indicate a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.125101 | Structured | 0.157322 | Uncertain | 0.786 | 0.270 | 0.000 | -6.907 | Likely Benign | 0.378 | Ambiguous | Likely Benign | 1.73 | Ambiguous | 1.3 | 0.07 | Likely Benign | 0.90 | Ambiguous | 0.59 | Ambiguous | 0.205 | Likely Benign | -3.05 | Deleterious | 0.000 | Benign | 0.000 | Benign | 3.34 | Benign | 0.04 | Affected | 0.2289 | 0.5791 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||
| c.1922C>T | S641L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S641L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields an equal split of benign and pathogenic calls. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.125101 | Structured | 0.157322 | Uncertain | 0.786 | 0.270 | 0.000 | -9.501 | Likely Pathogenic | 0.237 | Likely Benign | Likely Benign | 0.33 | Likely Benign | 0.3 | -0.14 | Likely Benign | 0.10 | Likely Benign | 0.32 | Likely Benign | 0.103 | Likely Benign | -4.33 | Deleterious | 0.115 | Benign | 0.014 | Benign | 3.39 | Benign | 0.07 | Tolerated | 0.1248 | 0.5376 | -3 | -2 | 4.6 | 26.08 | ||||||||||||||||||||||||||||||
| c.1925A>G | K642R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K642R is not reported in ClinVar and has no entries in gnomAD. Prediction tools that assess pathogenicity largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign. In contrast, PROVEAN, SIFT, and ESM1b predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, Foldetta is benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the balance of evidence favors a benign classification, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.061840 | Structured | 0.181468 | Uncertain | 0.806 | 0.289 | 0.000 | -10.562 | Likely Pathogenic | 0.192 | Likely Benign | Likely Benign | -0.13 | Likely Benign | 0.1 | 0.39 | Likely Benign | 0.13 | Likely Benign | 0.04 | Likely Benign | 0.319 | Likely Benign | -2.79 | Deleterious | 0.001 | Benign | 0.001 | Benign | 2.91 | Benign | 0.02 | Affected | 0.4962 | 0.1262 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||
| c.1929G>C | E643D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E643D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, AlphaMissense‑Default, AlphaMissense‑Optimized, FATHMM, and polyPhen2_HumVar. Tools that predict a pathogenic effect are premPS, PROVEAN, polyPhen2_HumDiv, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of predictions (8 benign vs. 5 pathogenic) support a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.033407 | Structured | 0.215915 | Uncertain | 0.871 | 0.315 | 0.000 | -8.083 | Likely Pathogenic | 0.223 | Likely Benign | Likely Benign | 0.46 | Likely Benign | 0.2 | -0.34 | Likely Benign | 0.06 | Likely Benign | 1.09 | Destabilizing | 0.292 | Likely Benign | -2.96 | Deleterious | 0.694 | Possibly Damaging | 0.064 | Benign | 2.98 | Benign | 0.01 | Affected | 0.2040 | 0.4276 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1929G>T | E643D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E643D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, AlphaMissense‑Default, AlphaMissense‑Optimized, FATHMM, and polyPhen2_HumVar. Tools that predict a pathogenic effect are premPS, PROVEAN, polyPhen2_HumDiv, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of predictions (8 benign vs. 5 pathogenic) support a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.033407 | Structured | 0.215915 | Uncertain | 0.871 | 0.315 | 0.000 | -8.083 | Likely Pathogenic | 0.223 | Likely Benign | Likely Benign | 0.46 | Likely Benign | 0.2 | -0.34 | Likely Benign | 0.06 | Likely Benign | 1.09 | Destabilizing | 0.292 | Likely Benign | -2.96 | Deleterious | 0.694 | Possibly Damaging | 0.064 | Benign | 2.98 | Benign | 0.01 | Affected | 0.2040 | 0.4276 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.1930G>C | D644H 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 D644H missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that predict a pathogenic effect are PROVEAN, polyPhen2_HumDiv, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. No other high‑accuracy predictions are available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.248888 | Uncertain | 0.883 | 0.320 | 0.000 | -6.786 | Likely Benign | 0.771 | Likely Pathogenic | Likely Benign | 0.34 | Likely Benign | 0.1 | -0.83 | Ambiguous | -0.25 | Likely Benign | 0.09 | Likely Benign | 0.284 | Likely Benign | -2.93 | Deleterious | 0.789 | Possibly Damaging | 0.158 | Benign | 3.43 | Benign | 0.07 | Tolerated | 0.1656 | 0.6306 | 1 | -1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||
| c.1931A>C | D644A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 D644A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, Foldetta, and premPS. Only PROVEAN predicts it as pathogenic, while Rosetta and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign; and Foldetta also predicts benign stability. Based on the aggregate evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.248888 | Uncertain | 0.883 | 0.320 | 0.000 | -3.358 | Likely Benign | 0.502 | Ambiguous | Likely Benign | -0.14 | Likely Benign | 0.1 | -0.83 | Ambiguous | -0.49 | Likely Benign | -0.18 | Likely Benign | 0.274 | Likely Benign | -3.90 | Deleterious | 0.311 | Benign | 0.032 | Benign | 3.51 | Benign | 0.39 | Tolerated | 0.3883 | 0.5481 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||
| c.1931A>G | D644G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D644G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and AlphaMissense‑Default (polyPhen‑2 HumVar is benign). High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.248888 | Uncertain | 0.883 | 0.320 | 0.000 | -4.496 | Likely Benign | 0.602 | Likely Pathogenic | Likely Benign | 0.13 | Likely Benign | 0.0 | -0.18 | Likely Benign | -0.03 | Likely Benign | 0.04 | Likely Benign | 0.271 | Likely Benign | -4.38 | Deleterious | 0.456 | Possibly Damaging | 0.069 | Benign | 3.46 | Benign | 0.14 | Tolerated | 0.4035 | 0.5610 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||
| c.1931A>T | D644V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D644V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.248888 | Uncertain | 0.883 | 0.320 | 0.000 | -6.230 | Likely Benign | 0.636 | Likely Pathogenic | Likely Benign | 0.50 | Ambiguous | 0.0 | -0.35 | Likely Benign | 0.08 | Likely Benign | -0.03 | Likely Benign | 0.347 | Likely Benign | -4.96 | Deleterious | 0.198 | Benign | 0.052 | Benign | 3.49 | Benign | 0.11 | Tolerated | 0.0951 | 0.6267 | -2 | -3 | 7.7 | -15.96 | ||||||||||||||||||||||||||||||
| c.1940G>A | G647D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G647D is reported in ClinVar as having no entry and is present in gnomAD (6‑33441199‑G‑A). Functional prediction tools largely agree on a benign effect: REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized all predict benign. Only AlphaMissense‑Default predicts pathogenic, while FoldX, Foldetta, premPS, and ESM1b are uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields benign; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is uncertain. Overall, the consensus of most tools and the high‑accuracy predictions indicate that G647D is most likely benign, and this is consistent with the absence of a pathogenic ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.048328 | Structured | 0.325524 | Uncertain | 0.936 | 0.356 | 0.000 | 6-33441199-G-A | 1 | 6.20e-7 | -7.643 | In-Between | 0.582 | Likely Pathogenic | Likely Benign | 0.68 | Ambiguous | 0.1 | 0.33 | Likely Benign | 0.51 | Ambiguous | 0.56 | Ambiguous | 0.098 | Likely Benign | -1.63 | Neutral | 0.282 | Benign | 0.128 | Benign | 3.45 | Benign | 0.24 | Tolerated | 3.37 | 30 | 0.1630 | 0.1372 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||
| c.1945A>C | M649L 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant M649L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN and AlphaMissense‑Default, while premPS remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta predicts benign stability. Overall, the preponderance of evidence points to a benign effect. This conclusion is consistent with the absence of a ClinVar assertion, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.051831 | Structured | 0.360413 | Uncertain | 0.962 | 0.345 | 0.000 | -5.210 | Likely Benign | 0.745 | Likely Pathogenic | Likely Benign | 0.19 | Likely Benign | 0.4 | 0.26 | Likely Benign | 0.23 | Likely Benign | 0.64 | Ambiguous | 0.294 | Likely Benign | -2.99 | Deleterious | 0.009 | Benign | 0.007 | Benign | 3.73 | Benign | 0.15 | Tolerated | 0.1361 | 0.4025 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.1945A>T | M649L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M649L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and AlphaMissense‑Default, while premPS remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta (combining FoldX‑MD and Rosetta outputs) as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.051831 | Structured | 0.360413 | Uncertain | 0.962 | 0.345 | 0.000 | -5.210 | Likely Benign | 0.745 | Likely Pathogenic | Likely Benign | 0.19 | Likely Benign | 0.4 | 0.26 | Likely Benign | 0.23 | Likely Benign | 0.64 | Ambiguous | 0.294 | Likely Benign | -2.99 | Deleterious | 0.009 | Benign | 0.007 | Benign | 3.73 | Benign | 0.15 | Tolerated | 0.1361 | 0.4025 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.1952A>G | E651G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E651G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Predictions that are uncertain or unavailable are FoldX, Rosetta, ESM1b, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as unavailable. Overall, the majority of tools (five benign vs. four pathogenic) lean toward a benign interpretation, and this does not contradict the lack of ClinVar annotation. Thus, based on the available predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.088832 | Structured | 0.365409 | Uncertain | 0.955 | 0.340 | 0.000 | -7.596 | In-Between | 0.591 | Likely Pathogenic | Likely Benign | 0.66 | Ambiguous | 0.1 | 1.61 | Ambiguous | 1.14 | Ambiguous | 0.32 | Likely Benign | 0.444 | Likely Benign | -4.78 | Deleterious | 0.999 | Probably Damaging | 0.935 | Probably Damaging | 3.31 | Benign | 0.06 | Tolerated | 0.2908 | 0.4488 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||
| c.1955T>A | F652Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F652Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that agree on a pathogenic effect are premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. Three tools (FoldX, Foldetta, AlphaMissense‑Optimized) give uncertain results. High‑accuracy methods provide no definitive verdict: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie (2 pathogenic, 2 benign) and thus inconclusive; Foldetta is uncertain. Consequently, the overall prediction landscape is balanced, with an equal number of benign and pathogenic calls and several uncertain results. The variant is therefore most likely **inconclusive** in terms of pathogenicity, and this lack of consensus does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.356594 | Uncertain | 0.966 | 0.338 | 0.000 | -5.437 | Likely Benign | 0.890 | Likely Pathogenic | Ambiguous | 1.05 | Ambiguous | 0.2 | 0.27 | Likely Benign | 0.66 | Ambiguous | 1.24 | Destabilizing | 0.363 | Likely Benign | -2.92 | Deleterious | 0.957 | Probably Damaging | 0.390 | Benign | 3.13 | Benign | 0.00 | Affected | 0.1085 | 0.1584 | 7 | 3 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||
| c.1957C>A | L653M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L653M has no ClinVar record and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic outcome are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Tools with uncertain or missing results (FoldX, Rosetta, Foldetta, premPS) are not considered evidence for either side. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs. two benign votes), and Foldetta is also inconclusive. Overall, the majority of standard predictors lean toward pathogenicity, but the most reliable high‑accuracy tool indicates a benign effect, leaving the variant’s impact uncertain. No ClinVar entry exists, so there is no contradiction with clinical annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.049374 | Structured | 0.335213 | Uncertain | 0.963 | 0.332 | 0.000 | -8.838 | Likely Pathogenic | 0.713 | Likely Pathogenic | Likely Benign | 0.99 | Ambiguous | 0.2 | 1.70 | Ambiguous | 1.35 | Ambiguous | 0.94 | Ambiguous | 0.259 | Likely Benign | -1.76 | Neutral | 1.000 | Probably Damaging | 0.991 | Probably Damaging | 3.13 | Benign | 0.01 | Affected | 0.0935 | 0.3227 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.1964T>C | L655P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L655P is catalogued in gnomAD (ID 6‑33441223‑T‑C) but has no ClinVar entry. Functional prediction tools show a split: benign calls come from REVEL, PROVEAN, SIFT, and FATHMM, whereas pathogenic calls are made by Rosetta, Foldetta, both polyPhen‑2 versions, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; FoldX and premPS are uncertain. High‑accuracy assessments give AlphaMissense‑Optimized as pathogenic, Foldetta as pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Overall, the majority of evidence points to a pathogenic effect. This conclusion is consistent with the absence of a ClinVar classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.015344 | Structured | 0.268808 | Uncertain | 0.961 | 0.274 | 0.000 | 6-33441223-T-C | 1 | 6.20e-7 | -9.771 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 1.33 | Ambiguous | 0.5 | 6.52 | Destabilizing | 3.93 | Destabilizing | 0.57 | Ambiguous | 0.126 | Likely Benign | -1.36 | Neutral | 0.981 | Probably Damaging | 0.772 | Possibly Damaging | 3.47 | Benign | 0.32 | Tolerated | 3.39 | 24 | 0.3598 | 0.1274 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||
| c.1966G>C | E656Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E656Q is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33441225‑G‑C). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default; Rosetta reports an uncertain result. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑2 split. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.032017 | Structured | 0.242242 | Uncertain | 0.963 | 0.264 | 0.000 | Uncertain | 1 | 6-33441225-G-C | 1 | 6.20e-7 | -9.145 | Likely Pathogenic | 0.766 | Likely Pathogenic | Likely Benign | -0.14 | Likely Benign | 0.0 | -0.81 | Ambiguous | -0.48 | Likely Benign | 0.25 | Likely Benign | 0.249 | Likely Benign | -2.29 | Neutral | 0.980 | Probably Damaging | 0.528 | Possibly Damaging | 3.46 | Benign | 0.02 | Affected | 3.39 | 24 | 0.1739 | 0.6645 | 2 | 2 | 0.0 | -0.98 | 224.3 | 1.7 | 0.0 | 0.1 | 0.1 | 0.0 | X | Potentially Benign | The carboxylate side chain of Glu656, located on an α helix (res. Ser641-Glu666), frequently forms a hydrogen bond with the nearby residue Ser659 on the same α helix. In the variant simulations, the carboxamide side chain of Gln656 alternatively forms a hydrogen bond with either Ser659 or Glu548 on an opposing helix (res. Ala533-Val560).Although the frequent interaction between Gln656 and Glu548 may strengthen or stabilize the tertiary structure assembly, the effect is likely to be marginal. | ||||||||||||||
| c.1972G>C | G658R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G658R is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, SIFT, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default. Four tools (FoldX, Foldetta, premPS, ESM1b) return uncertain results. High‑accuracy methods give a mixed signal: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta is uncertain. Because the majority of conventional predictors lean benign and no ClinVar evidence contradicts this, the variant is most likely benign, though the conflicting high‑accuracy predictions leave some uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.029376 | Structured | 0.180299 | Uncertain | 0.942 | 0.251 | 0.000 | -7.725 | In-Between | 0.618 | Likely Pathogenic | Likely Benign | -1.17 | Ambiguous | 0.1 | -0.46 | Likely Benign | -0.82 | Ambiguous | 0.64 | Ambiguous | 0.120 | Likely Benign | -3.11 | Deleterious | 0.955 | Possibly Damaging | 0.591 | Possibly Damaging | 3.40 | Benign | 0.19 | Tolerated | 0.1157 | 0.3888 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1972G>T | G658C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G658C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. One tool, ESM1b, yields an uncertain result. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors benign (2 benign vs. 1 pathogenic, 1 uncertain), and Foldetta predicts a benign impact on protein stability. Overall, the majority of evidence points to a benign effect. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.029376 | Structured | 0.180299 | Uncertain | 0.942 | 0.251 | 0.000 | -7.577 | In-Between | 0.170 | Likely Benign | Likely Benign | 0.05 | Likely Benign | 0.0 | 0.04 | Likely Benign | 0.05 | Likely Benign | 0.46 | Likely Benign | 0.146 | Likely Benign | -3.49 | Deleterious | 0.989 | Probably Damaging | 0.544 | Possibly Damaging | 3.37 | Benign | 0.04 | Affected | 0.1509 | 0.3141 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||
| c.1973G>A | G658D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G658D is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6-33441232‑G‑A). Functional prediction tools that agree on a benign effect include REVEL, FoldX, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only PROVEAN predicts a pathogenic outcome, while Rosetta, Foldetta, ESM1b, and AlphaMissense‑Default are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta, which integrates FoldX‑MD and Rosetta, is also inconclusive. Overall, the preponderance of evidence points to a benign effect, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.029376 | Structured | 0.180299 | Uncertain | 0.942 | 0.251 | 0.000 | Uncertain | 1 | 6-33441232-G-A | 3 | 1.86e-6 | -7.786 | In-Between | 0.442 | Ambiguous | Likely Benign | -0.40 | Likely Benign | 0.1 | -0.59 | Ambiguous | -0.50 | Ambiguous | 0.46 | Likely Benign | 0.144 | Likely Benign | -2.64 | Deleterious | 0.008 | Benign | 0.005 | Benign | 3.53 | Benign | 0.38 | Tolerated | 3.39 | 24 | 0.2106 | 0.2333 | 1 | -1 | -3.1 | 58.04 | 219.8 | -84.3 | 0.0 | 0.0 | 0.2 | 0.1 | X | Potentially Pathogenic | Gly658, located on the outer surface of an α helix (res. Ser641-Glu666), weakens the helix integrity at that spot, which is necessary for the kink in the middle of the long helix. In the variant simulations, the carboxylic acid side chain of Asp658 is on the surface of the α helix and is not involved in any interactions. However, aspartate is not as effective a breaker of the secondary structure element as glycine, which may lead to misfolding. | ||||||||||||||
| c.1976C>G | S659C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S659C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, AlphaMissense‑Default, AlphaMissense‑Optimized, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and SIFT. Two tools (Rosetta and ESM1b) return uncertain results and are not counted as evidence for either side. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.067594 | Structured | 0.154597 | Uncertain | 0.954 | 0.283 | 0.000 | -7.133 | In-Between | 0.161 | Likely Benign | Likely Benign | 0.09 | Likely Benign | 0.0 | 0.69 | Ambiguous | 0.39 | Likely Benign | 0.36 | Likely Benign | 0.173 | Likely Benign | -4.12 | Deleterious | 0.981 | Probably Damaging | 0.397 | Benign | 3.36 | Benign | 0.04 | Affected | 0.1130 | 0.5384 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.1981C>A | Q661K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q661K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Only ESM1b predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority (2 benign vs. 1 pathogenic, with one uncertain), and Foldetta also predicts benign stability. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.048328 | Structured | 0.117089 | Uncertain | 0.924 | 0.309 | 0.000 | -10.581 | Likely Pathogenic | 0.400 | Ambiguous | Likely Benign | -0.01 | Likely Benign | 0.0 | -0.18 | Likely Benign | -0.10 | Likely Benign | 0.04 | Likely Benign | 0.108 | Likely Benign | -1.89 | Neutral | 0.098 | Benign | 0.030 | Benign | 3.59 | Benign | 0.42 | Tolerated | 0.1756 | 0.3592 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||||
| c.1981C>G | Q661E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q661E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Four tools (FoldX, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of available predictions lean toward a benign impact. This conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.048328 | Structured | 0.117089 | Uncertain | 0.924 | 0.309 | 0.000 | -12.121 | Likely Pathogenic | 0.370 | Ambiguous | Likely Benign | 0.64 | Ambiguous | 0.1 | 0.35 | Likely Benign | 0.50 | Ambiguous | 0.61 | Ambiguous | 0.141 | Likely Benign | -2.15 | Neutral | 0.988 | Probably Damaging | 0.619 | Possibly Damaging | 3.42 | Benign | 0.03 | Affected | 0.1199 | 0.2439 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1982A>G | Q661R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q661R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that predict a pathogenic effect are polyPhen2_HumDiv and ESM1b. FoldX, Rosetta, Foldetta, and AlphaMissense‑Default are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign prediction, while Foldetta remains uncertain. Overall, the majority of available evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.048328 | Structured | 0.117089 | Uncertain | 0.924 | 0.309 | 0.000 | -10.386 | Likely Pathogenic | 0.479 | Ambiguous | Likely Benign | -1.12 | Ambiguous | 0.0 | -0.59 | Ambiguous | -0.86 | Ambiguous | -0.15 | Likely Benign | 0.281 | Likely Benign | -2.06 | Neutral | 0.812 | Possibly Damaging | 0.251 | Benign | 3.52 | Benign | 0.18 | Tolerated | 0.1502 | 0.2196 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||
| c.1985A>T | Q662L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q662L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, AlphaMissense‑Default, AlphaMissense‑Optimized, polyPhen2_HumVar, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen2_HumDiv, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Overall, the majority of evidence supports a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, so there is no contradiction with existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.046336 | Structured | 0.103446 | Uncertain | 0.932 | 0.323 | 0.000 | -10.291 | Likely Pathogenic | 0.300 | Likely Benign | Likely Benign | 0.04 | Likely Benign | 0.0 | 0.12 | Likely Benign | 0.08 | Likely Benign | 0.17 | Likely Benign | 0.186 | Likely Benign | -4.09 | Deleterious | 0.699 | Possibly Damaging | 0.057 | Benign | 3.42 | Benign | 0.10 | Tolerated | 0.1070 | 0.5255 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||
| c.1990T>A | L664M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L664M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is also uncertain due to a 2‑vs‑2 split; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. Overall, the majority of conventional tools predict a pathogenic impact, whereas the high‑accuracy methods do not provide a definitive verdict. Consequently, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.100716 | Structured | 0.089318 | Uncertain | 0.937 | 0.339 | 0.000 | -11.819 | Likely Pathogenic | 0.807 | Likely Pathogenic | Ambiguous | 0.49 | Likely Benign | 0.1 | 1.35 | Ambiguous | 0.92 | Ambiguous | 0.96 | Ambiguous | 0.429 | Likely Benign | -2.00 | Neutral | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.92 | Benign | 0.01 | Affected | 0.0620 | 0.2541 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.1993T>A | Y665N 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 Y665N variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Four tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Default) give uncertain results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of conventional predictors lean toward a benign impact, and this assessment does not contradict ClinVar status (none). Thus, the variant is most likely benign based on the prevailing predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.086641 | Uncertain | 0.922 | 0.361 | 0.000 | -11.189 | Likely Pathogenic | 0.470 | Ambiguous | Likely Benign | 1.11 | Ambiguous | 0.1 | 1.43 | Ambiguous | 1.27 | Ambiguous | 0.50 | Likely Benign | 0.219 | Likely Benign | -3.03 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.50 | Benign | 0.88 | Tolerated | 0.1929 | 0.0573 | -2 | -2 | -2.2 | -49.07 | ||||||||||||||||||||||||||||||
| c.1993T>C | Y665H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y665H is not reported in ClinVar (ClinVar status: not reported) but is present in gnomAD (gnomAD ID: 6‑33441252‑T‑C). Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Tools with uncertain or inconclusive results are FoldX, Foldetta, and AlphaMissense‑Default. High‑accuracy methods give a benign consensus: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta remains uncertain. Overall, the majority of predictions (7 benign vs. 4 pathogenic) and the high‑accuracy consensus support a benign classification. This conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.086641 | Uncertain | 0.922 | 0.361 | 0.000 | 6-33441252-T-C | 2 | 1.24e-6 | -9.303 | Likely Pathogenic | 0.389 | Ambiguous | Likely Benign | 0.85 | Ambiguous | 0.0 | 0.28 | Likely Benign | 0.57 | Ambiguous | 1.12 | Destabilizing | 0.129 | Likely Benign | -2.00 | Neutral | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.45 | Benign | 0.48 | Tolerated | 3.38 | 28 | 0.2122 | 0.0513 | 2 | 0 | -1.9 | -26.03 | |||||||||||||||||||||||||
| c.1994A>C | Y665S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y665S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans pathogenic, and Foldetta is uncertain (treated as unavailable). Overall, the balance of evidence, particularly the pathogenic signal from the SGM Consensus and the equal split among standard predictors, indicates that the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.086641 | Uncertain | 0.922 | 0.361 | 0.000 | -9.110 | Likely Pathogenic | 0.453 | Ambiguous | Likely Benign | 1.24 | Ambiguous | 0.2 | 1.40 | Ambiguous | 1.32 | Ambiguous | 0.87 | Ambiguous | 0.202 | Likely Benign | -2.50 | Deleterious | 1.000 | Probably Damaging | 0.994 | Probably Damaging | 3.55 | Benign | 0.62 | Tolerated | 0.3995 | 0.1793 | -3 | -2 | 0.5 | -76.10 | ||||||||||||||||||||||||||||||
| c.1994A>G | Y665C 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y665C is listed in ClinVar with no assertion (status: None) and is present in gnomAD (ID 6‑33441253‑A‑G). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. FoldX, Rosetta, and Foldetta give uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is also unavailable. Overall, the evidence is mixed, but the majority of high‑confidence tools lean toward a benign interpretation. Thus, the variant is most likely benign based on current predictions, and this does not contradict the ClinVar status, which remains unclassified. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.086641 | Uncertain | 0.922 | 0.361 | 0.000 | 6-33441253-A-G | 1 | 6.20e-7 | -9.007 | Likely Pathogenic | 0.261 | Likely Benign | Likely Benign | 1.05 | Ambiguous | 0.1 | 1.60 | Ambiguous | 1.33 | Ambiguous | 1.12 | Destabilizing | 0.210 | Likely Benign | -3.22 | Deleterious | 1.000 | Probably Damaging | 0.981 | Probably Damaging | 3.45 | Benign | 0.14 | Tolerated | 3.38 | 28 | 0.2562 | 0.2019 | -2 | 0 | 3.8 | -60.04 | |||||||||||||||||||||||||
| c.1996G>C | E666Q 2D AIThe SynGAP1 E666Q missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default; FoldX is uncertain and therefore not counted. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split and is treated as unavailable. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the majority of reliable predictions indicate a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.086870 | Uncertain | 0.925 | 0.387 | 0.000 | -10.963 | Likely Pathogenic | 0.737 | Likely Pathogenic | Likely Benign | 0.54 | Ambiguous | 0.5 | 0.20 | Likely Benign | 0.37 | Likely Benign | 0.46 | Likely Benign | 0.268 | Likely Benign | -2.29 | Neutral | 0.997 | Probably Damaging | 0.696 | Possibly Damaging | 3.46 | Benign | 0.13 | Tolerated | 0.1437 | 0.4788 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.1999A>C | I667L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I667L is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, indicating no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.142424 | Structured | 0.083597 | Uncertain | 0.927 | 0.379 | 0.000 | -10.452 | Likely Pathogenic | 0.459 | Ambiguous | Likely Benign | 0.93 | Ambiguous | 0.2 | 0.72 | Ambiguous | 0.83 | Ambiguous | 0.75 | Ambiguous | 0.300 | Likely Benign | -1.97 | Neutral | 0.457 | Possibly Damaging | 0.392 | Benign | 3.36 | Benign | 0.13 | Tolerated | 0.0980 | 0.3145 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.1999A>G | I667V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I667V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Only ESM1b predicts a pathogenic outcome; all other tools (FoldX, Rosetta, premPS, AlphaMissense‑Default, Foldetta) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority vote. Foldetta’s stability prediction is uncertain. Overall, the preponderance of evidence points to a benign effect. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.142424 | Structured | 0.083597 | Uncertain | 0.927 | 0.379 | 0.000 | -8.482 | Likely Pathogenic | 0.406 | Ambiguous | Likely Benign | 1.32 | Ambiguous | 0.0 | 1.06 | Ambiguous | 1.19 | Ambiguous | 0.85 | Ambiguous | 0.234 | Likely Benign | -0.86 | Neutral | 0.341 | Benign | 0.052 | Benign | 3.35 | Benign | 0.15 | Tolerated | 0.1178 | 0.3059 | 4 | 3 | -0.3 | -14.03 | ||||||||||||||||||||||||||||||
| c.2002T>G | S668A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 S668A missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, AlphaMissense‑Optimized, and Foldetta. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools—AlphaMissense‑Default and FoldX—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Taking all evidence together, the majority of predictions (seven benign versus four pathogenic) and the two high‑accuracy benign calls suggest that the variant is most likely benign. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.247041 | Structured | 0.084935 | Uncertain | 0.922 | 0.370 | 0.000 | -12.011 | Likely Pathogenic | 0.506 | Ambiguous | Likely Benign | 0.73 | Ambiguous | 0.3 | -0.44 | Likely Benign | 0.15 | Likely Benign | 0.48 | Likely Benign | 0.399 | Likely Benign | -2.99 | Deleterious | 0.887 | Possibly Damaging | 0.738 | Possibly Damaging | 3.28 | Benign | 0.19 | Tolerated | 0.4866 | 0.3967 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||
| c.2005A>C | N669H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N669H has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) are uncertain and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.142424 | Structured | 0.086615 | Uncertain | 0.872 | 0.380 | 0.000 | -10.364 | Likely Pathogenic | 0.421 | Ambiguous | Likely Benign | 1.26 | Ambiguous | 0.2 | 1.69 | Ambiguous | 1.48 | Ambiguous | 0.80 | Ambiguous | 0.432 | Likely Benign | -4.49 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 3.35 | Benign | 0.01 | Affected | 0.1732 | 0.4839 | 2 | 1 | 0.3 | 23.04 | ||||||||||||||||||||||||||||||
| c.2006A>G | N669S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N669S is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33441265‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta (combining FoldX‑MD and Rosetta) is also inconclusive. No folding‑stability metrics (FoldX, Rosetta, Foldetta) provide definitive evidence. Overall, the majority of predictions lean toward a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.142424 | Structured | 0.086615 | Uncertain | 0.872 | 0.380 | 0.000 | 6-33441265-A-G | 3 | 1.86e-6 | -8.369 | Likely Pathogenic | 0.187 | Likely Benign | Likely Benign | 0.55 | Ambiguous | 0.1 | 1.88 | Ambiguous | 1.22 | Ambiguous | 0.35 | Likely Benign | 0.210 | Likely Benign | -4.02 | Deleterious | 0.999 | Probably Damaging | 0.960 | Probably Damaging | 3.52 | Benign | 0.14 | Tolerated | 3.39 | 27 | 0.3521 | 0.4480 | 1 | 1 | 2.7 | -27.03 | |||||||||||||||||||||||||
| c.2009T>G | L670R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L670R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of reliable predictions indicate a benign effect. There is no ClinVar annotation to contradict this assessment, so the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.161087 | Structured | 0.090855 | Uncertain | 0.812 | 0.385 | 0.000 | -9.801 | Likely Pathogenic | 0.552 | Ambiguous | Likely Benign | 0.64 | Ambiguous | 0.1 | 1.42 | Ambiguous | 1.03 | Ambiguous | 0.67 | Ambiguous | 0.193 | Likely Benign | -1.74 | Neutral | 0.993 | Probably Damaging | 0.755 | Possibly Damaging | 3.43 | Benign | 0.41 | Tolerated | 0.1327 | 0.1145 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||
| c.2014A>C | T672P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T672P missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Uncertain results come from premPS and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. No predictions are missing or inconclusive. Overall, the majority of evidence points to a pathogenic impact for T672P. This conclusion is not contradicted by ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.116183 | Structured | 0.102069 | Uncertain | 0.586 | 0.362 | 0.000 | -11.022 | Likely Pathogenic | 0.346 | Ambiguous | Likely Benign | 2.22 | Destabilizing | 0.2 | 5.54 | Destabilizing | 3.88 | Destabilizing | 0.55 | Ambiguous | 0.087 | Likely Benign | -4.20 | Deleterious | 0.968 | Probably Damaging | 0.824 | Possibly Damaging | 3.40 | Benign | 0.01 | Affected | 0.1988 | 0.5532 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.2015C>T | T672M 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant T672M is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33441274‑C‑T). Prediction tools that classify the variant as benign include REVEL, FoldX, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. Rosetta and Foldetta report uncertain results, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while SGM Consensus and Foldetta remain unavailable. Overall, the balance of evidence favors a benign effect, and this conclusion does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.116183 | Structured | 0.102069 | Uncertain | 0.586 | 0.362 | 0.000 | Conflicting | 3 | 6-33441274-C-T | 19 | 1.18e-5 | -9.472 | Likely Pathogenic | 0.174 | Likely Benign | Likely Benign | 0.31 | Likely Benign | 0.4 | 1.52 | Ambiguous | 0.92 | Ambiguous | 0.41 | Likely Benign | 0.127 | Likely Benign | -4.34 | Deleterious | 0.993 | Probably Damaging | 0.520 | Possibly Damaging | 3.39 | Benign | 0.00 | Affected | 3.40 | 25 | 0.1332 | 0.6677 | -1 | -1 | 2.6 | 30.09 | 231.9 | -52.9 | 1.1 | 0.1 | 0.5 | 0.0 | X | X | Potentially Pathogenic | The hydroxyl group of Thr672, located in an entangled α-α loop connecting the two α-helices (res. Ser641-Glu666 and res. Leu685-Val699), is involved in a highly coordinated hydrogen-bonding network between residues from two α-helices (res. Ser641-Glu666 and res. Arg563-Glu578) and from the α-α loop itself, such as Lys566, Glu666, and Asn669. Met672 can only form a hydrogen bond with the amino group of the Lys566 side chain via its backbone carbonyl group. Nevertheless, the Lys566-Glu666 salt bridge forms intermittently. This is possible because Asn669 keeps the carboxylate group of Glu666 in the vicinity through hydrogen bonding, and the hydrophobic side chain of Met stays mostly rotated away from the salt bridge. Consequently, no drastic disruption of the hydrogen-bond network that keeps the loop close to the helices occurs in the variant simulations. | |||||||||||||
| c.2018T>A | L673Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L673Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect include Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. FoldX and Foldetta, as well as AlphaMissense‑Default, are inconclusive and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta remains uncertain. Overall, the majority of available predictions (seven pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar claim exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.060549 | Structured | 0.104692 | Uncertain | 0.545 | 0.369 | 0.000 | -11.963 | Likely Pathogenic | 0.445 | Ambiguous | Likely Benign | 1.56 | Ambiguous | 0.2 | 2.32 | Destabilizing | 1.94 | Ambiguous | 1.02 | Destabilizing | 0.190 | Likely Benign | -3.40 | Deleterious | 1.000 | Probably Damaging | 0.968 | Probably Damaging | 3.32 | Benign | 0.03 | Affected | 0.1045 | 0.1172 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||
| c.2020A>C | T674P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T674P missense change is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. The high‑accuracy AlphaMissense‑Optimized predicts benign, while Foldetta predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. No other high‑accuracy tool provides a definitive call. Consequently, the evidence is evenly split between benign and pathogenic, leaving the variant’s clinical significance uncertain. This uncertainty does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.109297 | Uncertain | 0.521 | 0.349 | 0.000 | -8.661 | Likely Pathogenic | 0.109 | Likely Benign | Likely Benign | 0.91 | Ambiguous | 0.6 | 4.12 | Destabilizing | 2.52 | Destabilizing | 0.12 | Likely Benign | 0.143 | Likely Benign | -2.69 | Deleterious | 0.995 | Probably Damaging | 0.891 | Possibly Damaging | 3.50 | Benign | 0.16 | Tolerated | 0.2039 | 0.6103 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.2021C>T | T674I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T674I is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (ID 6‑33441280‑C‑T). Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, and ESM1b (polyPhen‑2 HumVar is benign). AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.109297 | Uncertain | 0.521 | 0.349 | 0.000 | 6-33441280-C-T | 2 | 1.24e-6 | -8.951 | Likely Pathogenic | 0.522 | Ambiguous | Likely Benign | -0.05 | Likely Benign | 0.1 | 0.28 | Likely Benign | 0.12 | Likely Benign | -0.04 | Likely Benign | 0.289 | Likely Benign | -3.18 | Deleterious | 0.981 | Probably Damaging | 0.444 | Benign | 3.46 | Benign | 0.11 | Tolerated | 3.40 | 24 | 0.0898 | 0.6945 | -1 | 0 | 5.2 | 12.05 | |||||||||||||||||||||||||
| c.2023A>G | N675D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N675D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. High‑accuracy assessments give AlphaMissense‑Optimized a benign call, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) a pathogenic call, and Foldetta an uncertain result. With an equal split of general predictions but a pathogenic majority in the high‑accuracy consensus, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.111024 | Uncertain | 0.513 | 0.333 | 0.000 | -12.829 | Likely Pathogenic | 0.497 | Ambiguous | Likely Benign | 1.41 | Ambiguous | 0.4 | -0.26 | Likely Benign | 0.58 | Ambiguous | 1.05 | Destabilizing | 0.246 | Likely Benign | -3.87 | Deleterious | 0.997 | Probably Damaging | 0.865 | Possibly Damaging | 3.53 | Benign | 0.17 | Tolerated | 0.1914 | 0.4901 | 2 | 1 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.2023A>T | N675Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N675Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of available predictions (six pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.111024 | Uncertain | 0.513 | 0.333 | 0.000 | -9.981 | Likely Pathogenic | 0.553 | Ambiguous | Likely Benign | 3.02 | Destabilizing | 1.3 | -0.81 | Ambiguous | 1.11 | Ambiguous | 0.19 | Likely Benign | 0.439 | Likely Benign | -5.86 | Deleterious | 1.000 | Probably Damaging | 0.955 | Probably Damaging | 3.38 | Benign | 0.01 | Affected | 0.0603 | 0.6417 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||
| c.2024A>C | N675T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N675T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and Foldetta. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. Uncertain results come from Rosetta and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. No evidence contradicts the ClinVar status. Overall, the balance of evidence, especially from the high‑accuracy tools, indicates that the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.111024 | Uncertain | 0.513 | 0.333 | 0.000 | -10.636 | Likely Pathogenic | 0.208 | Likely Benign | Likely Benign | 0.36 | Likely Benign | 0.3 | -0.67 | Ambiguous | -0.16 | Likely Benign | 0.56 | Ambiguous | 0.258 | Likely Benign | -4.63 | Deleterious | 0.991 | Probably Damaging | 0.710 | Possibly Damaging | 3.49 | Benign | 0.05 | Affected | 0.1283 | 0.7910 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||
| c.2024A>G | N675S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N675S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, FATHMM, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Two tools (premPS and ESM1b) return uncertain results. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign consensus; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. Taken together, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.111024 | Uncertain | 0.513 | 0.333 | 0.000 | -7.871 | In-Between | 0.081 | Likely Benign | Likely Benign | 0.37 | Likely Benign | 0.0 | -0.43 | Likely Benign | -0.03 | Likely Benign | 0.61 | Ambiguous | 0.157 | Likely Benign | -3.78 | Deleterious | 0.993 | Probably Damaging | 0.606 | Possibly Damaging | 3.50 | Benign | 0.11 | Tolerated | 0.3462 | 0.7587 | 1 | 1 | 2.7 | -27.03 | ||||||||||||||||||||||||||||||
| c.2026A>C | S676R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S676R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, FATHMM, and polyPhen‑2 HumVar, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or folding stability result is missing; all available data are reported. Overall, the balance of evidence (six benign versus four pathogenic predictions, with the high‑accuracy Foldetta supporting benign) indicates that the variant is most likely benign. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.209395 | Structured | 0.113632 | Uncertain | 0.551 | 0.338 | 0.125 | -10.665 | Likely Pathogenic | 0.936 | Likely Pathogenic | Ambiguous | 0.22 | Likely Benign | 0.3 | 0.74 | Ambiguous | 0.48 | Likely Benign | 0.45 | Likely Benign | 0.136 | Likely Benign | -2.34 | Neutral | 0.891 | Possibly Damaging | 0.278 | Benign | 3.40 | Benign | 0.02 | Affected | 0.0962 | 0.3454 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2026A>G | S676G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S676G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and SIFT. Two tools (ESM1b and premPS) returned uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, and Foldetta predicts a benign stability change. No prediction or folding result is missing or inconclusive. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.209395 | Structured | 0.113632 | Uncertain | 0.551 | 0.338 | 0.125 | -7.249 | In-Between | 0.117 | Likely Benign | Likely Benign | 0.38 | Likely Benign | 0.1 | 0.25 | Likely Benign | 0.32 | Likely Benign | 0.67 | Ambiguous | 0.132 | Likely Benign | -2.74 | Deleterious | 0.855 | Possibly Damaging | 0.100 | Benign | 3.39 | Benign | 0.02 | Affected | 0.2772 | 0.4342 | 1 | 0 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||
| c.2027G>T | S676I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S676I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and ESM1b. FoldX, Rosetta, Foldetta, and AlphaMissense‑Default are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, while Foldetta remains uncertain. Overall, the balance of evidence (five benign versus three pathogenic predictions) indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.209395 | Structured | 0.113632 | Uncertain | 0.551 | 0.338 | 0.125 | -10.165 | Likely Pathogenic | 0.383 | Ambiguous | Likely Benign | 1.25 | Ambiguous | 0.3 | 1.12 | Ambiguous | 1.19 | Ambiguous | 0.14 | Likely Benign | 0.170 | Likely Benign | -2.46 | Neutral | 0.801 | Possibly Damaging | 0.063 | Benign | 3.38 | Benign | 0.02 | Affected | 0.0879 | 0.5720 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||
| c.2028C>A | S676R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S676R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, FATHMM, and polyPhen‑2 HumVar, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or folding stability result is missing; all available data are included. Overall, the balance of evidence (six benign versus four pathogenic predictions, with the high‑accuracy Foldetta supporting benign) indicates that the variant is most likely benign. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.209395 | Structured | 0.113632 | Uncertain | 0.551 | 0.338 | 0.125 | -10.665 | Likely Pathogenic | 0.936 | Likely Pathogenic | Ambiguous | 0.22 | Likely Benign | 0.3 | 0.74 | Ambiguous | 0.48 | Likely Benign | 0.45 | Likely Benign | 0.157 | Likely Benign | -2.34 | Neutral | 0.891 | Possibly Damaging | 0.278 | Benign | 3.40 | Benign | 0.02 | Affected | 0.0962 | 0.3454 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2028C>G | S676R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S676R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, FATHMM, and polyPhen‑2 HumVar, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or folding stability result is missing; all available data are included. Overall, the balance of evidence (six benign versus four pathogenic predictions, with the high‑accuracy Foldetta supporting benign) indicates that the variant is most likely benign. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.209395 | Structured | 0.113632 | Uncertain | 0.551 | 0.338 | 0.125 | -10.665 | Likely Pathogenic | 0.936 | Likely Pathogenic | Ambiguous | 0.22 | Likely Benign | 0.3 | 0.74 | Ambiguous | 0.48 | Likely Benign | 0.45 | Likely Benign | 0.156 | Likely Benign | -2.34 | Neutral | 0.891 | Possibly Damaging | 0.278 | Benign | 3.40 | Benign | 0.02 | Affected | 0.0962 | 0.3454 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2029A>C | S677R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S677R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Those that predict a pathogenic effect are FoldX, ESM1b, and AlphaMissense‑Default. Uncertain results come from Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, the majority of tools and the high‑accuracy benign prediction suggest the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.194234 | Structured | 0.115685 | Uncertain | 0.555 | 0.338 | 0.125 | -9.388 | Likely Pathogenic | 0.714 | Likely Pathogenic | Likely Benign | 2.59 | Destabilizing | 0.1 | 0.73 | Ambiguous | 1.66 | Ambiguous | 0.71 | Ambiguous | 0.104 | Likely Benign | -2.49 | Neutral | 0.385 | Benign | 0.037 | Benign | 3.31 | Benign | 0.12 | Tolerated | 0.1120 | 0.4195 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2030G>T | S677I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S677I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Pathogenicity is suggested only by PROVEAN and SIFT, while Foldetta and ESM1b give uncertain results. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.194234 | Structured | 0.115685 | Uncertain | 0.555 | 0.338 | 0.125 | -7.942 | In-Between | 0.156 | Likely Benign | Likely Benign | -0.09 | Likely Benign | 0.0 | -2.25 | Stabilizing | -1.17 | Ambiguous | -0.01 | Likely Benign | 0.097 | Likely Benign | -2.81 | Deleterious | 0.002 | Benign | 0.002 | Benign | 3.34 | Benign | 0.05 | Affected | 0.1040 | 0.6463 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||
| c.2031T>A | S677R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S677R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are FoldX, ESM1b, and AlphaMissense‑Default. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and premPS. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is uncertain. Overall, the majority of consensus tools (six benign vs. three pathogenic) lean toward a benign interpretation. This prediction does not contradict any ClinVar status, as none is available. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.194234 | Structured | 0.115685 | Uncertain | 0.555 | 0.338 | 0.125 | -9.388 | Likely Pathogenic | 0.714 | Likely Pathogenic | Likely Benign | 2.59 | Destabilizing | 0.1 | 0.73 | Ambiguous | 1.66 | Ambiguous | 0.71 | Ambiguous | 0.150 | Likely Benign | -2.49 | Neutral | 0.385 | Benign | 0.037 | Benign | 3.31 | Benign | 0.12 | Tolerated | 0.1120 | 0.4195 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2031T>G | S677R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S677R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are FoldX, ESM1b, and AlphaMissense‑Default. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and premPS. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is uncertain. Overall, the majority of consensus tools (six benign vs. three pathogenic) lean toward a benign interpretation. This prediction does not contradict any ClinVar status, as none is available. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.194234 | Structured | 0.115685 | Uncertain | 0.555 | 0.338 | 0.125 | -9.388 | Likely Pathogenic | 0.714 | Likely Pathogenic | Likely Benign | 2.59 | Destabilizing | 0.1 | 0.73 | Ambiguous | 1.66 | Ambiguous | 0.71 | Ambiguous | 0.150 | Likely Benign | -2.49 | Neutral | 0.385 | Benign | 0.037 | Benign | 3.31 | Benign | 0.12 | Tolerated | 0.1120 | 0.4195 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2032A>C | S678R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta predicts a benign effect. Overall, the majority of tools (8 benign vs. 4 pathogenic) support a benign classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.708 | Likely Pathogenic | 0.878 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.2 | 0.48 | Likely Benign | 0.06 | Likely Benign | 0.32 | Likely Benign | 0.106 | Likely Benign | -2.07 | Neutral | 0.454 | Possibly Damaging | 0.057 | Benign | 3.44 | Benign | 0.02 | Affected | 0.0950 | 0.3519 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2032A>T | S678C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FoldX, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Tools with uncertain results are Rosetta and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign (2 benign vs. 1 pathogenic, 1 uncertain), and Foldetta also predicts benign. No prediction or folding stability result is missing or inconclusive. Overall, the preponderance of evidence indicates the variant is most likely benign, and this conclusion does not contradict any ClinVar status because the variant is not yet classified in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -7.879 | In-Between | 0.095 | Likely Benign | Likely Benign | 0.21 | Likely Benign | 0.2 | 0.55 | Ambiguous | 0.38 | Likely Benign | 0.35 | Likely Benign | 0.094 | Likely Benign | -3.31 | Deleterious | 0.947 | Possibly Damaging | 0.527 | Possibly Damaging | 3.37 | Benign | 0.01 | Affected | 0.1080 | 0.5875 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.2033G>T | S678I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence (10 benign vs. 3 pathogenic) supports a benign classification. This consensus does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.735 | Likely Pathogenic | 0.253 | Likely Benign | Likely Benign | 0.36 | Likely Benign | 0.3 | -0.01 | Likely Benign | 0.18 | Likely Benign | -0.11 | Likely Benign | 0.119 | Likely Benign | -3.99 | Deleterious | 0.294 | Benign | 0.057 | Benign | 3.45 | Benign | 0.01 | Affected | 0.0941 | 0.5954 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||
| c.2034C>A | S678R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is classified as uncertain. High‑accuracy assessments show AlphaMissense‑Optimized uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta predicts a benign effect. Overall, the majority of evidence (8 benign vs. 4 pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.708 | Likely Pathogenic | 0.878 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.2 | 0.48 | Likely Benign | 0.06 | Likely Benign | 0.32 | Likely Benign | 0.158 | Likely Benign | -2.07 | Neutral | 0.454 | Possibly Damaging | 0.057 | Benign | 3.44 | Benign | 0.02 | Affected | 0.0950 | 0.3519 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2034C>G | S678R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is classified as uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a benign stability change. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.708 | Likely Pathogenic | 0.878 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.2 | 0.48 | Likely Benign | 0.06 | Likely Benign | 0.32 | Likely Benign | 0.157 | Likely Benign | -2.07 | Neutral | 0.454 | Possibly Damaging | 0.057 | Benign | 3.44 | Benign | 0.02 | Affected | 0.0950 | 0.3519 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2036T>A | F679Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F679Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Two tools give uncertain results: premPS and AlphaMissense‑Default. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) resolves to benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. Overall, the majority of evidence indicates that F679Y is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.222385 | Structured | 0.129316 | Uncertain | 0.700 | 0.320 | 0.000 | -5.842 | Likely Benign | 0.462 | Ambiguous | Likely Benign | 0.48 | Likely Benign | 0.2 | 0.13 | Likely Benign | 0.31 | Likely Benign | 0.71 | Ambiguous | 0.315 | Likely Benign | -2.71 | Deleterious | 0.993 | Probably Damaging | 0.952 | Probably Damaging | 3.47 | Benign | 0.14 | Tolerated | 0.1306 | 0.1954 | 7 | 3 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||
| c.2045A>T | Y682F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y682F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. The SGM Consensus, which is a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic votes) and is treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta (combining FoldX‑MD and Rosetta outputs) as benign, and the SGM Consensus remains unavailable. Overall, the balance of evidence—both from general predictors and from the high‑accuracy tools—leans toward a benign classification. This conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.206376 | Structured | 0.141467 | Uncertain | 0.758 | 0.328 | 0.000 | -9.740 | Likely Pathogenic | 0.225 | Likely Benign | Likely Benign | 0.21 | Likely Benign | 0.1 | -0.09 | Likely Benign | 0.06 | Likely Benign | 0.42 | Likely Benign | 0.278 | Likely Benign | -3.72 | Deleterious | 0.997 | Probably Damaging | 0.947 | Probably Damaging | 3.40 | Benign | 0.04 | Affected | 0.2452 | 0.3662 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||
| c.2047A>C | I683L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I683L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b. Remaining methods (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, while Foldetta remains uncertain. Overall, the balance of evidence favors a benign impact for I683L, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.143268 | Uncertain | 0.848 | 0.314 | 0.000 | -9.988 | Likely Pathogenic | 0.488 | Ambiguous | Likely Benign | 0.63 | Ambiguous | 0.1 | 0.76 | Ambiguous | 0.70 | Ambiguous | 0.69 | Ambiguous | 0.286 | Likely Benign | -2.00 | Neutral | 0.011 | Benign | 0.056 | Benign | 3.43 | Benign | 0.04 | Affected | 0.0957 | 0.3185 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2049C>G | I683M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I683M variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of definitive predictions (five pathogenic vs. three benign) point to a pathogenic impact. Thus, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.143268 | Uncertain | 0.848 | 0.314 | 0.000 | -9.010 | Likely Pathogenic | 0.424 | Ambiguous | Likely Benign | 0.69 | Ambiguous | 0.1 | 0.68 | Ambiguous | 0.69 | Ambiguous | 0.74 | Ambiguous | 0.296 | Likely Benign | -2.88 | Deleterious | 0.999 | Probably Damaging | 0.986 | Probably Damaging | 3.30 | Benign | 0.01 | Affected | 0.0933 | 0.2662 | 2 | 1 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||
| c.2053T>A | L685M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L685M is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, and FATHMM, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. Uncertain predictions come from premPS, Foldetta, Rosetta, and AlphaMissense‑Optimized. High‑accuracy methods give inconclusive results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑to‑2 tie and therefore uncertain; Foldetta also reports an uncertain stability change. Consequently, the overall computational evidence is mixed, with a slight tilt toward pathogenicity. Thus, the variant is most likely pathogenic based on predictions, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.162061 | Uncertain | 0.913 | 0.280 | 0.000 | -10.790 | Likely Pathogenic | 0.902 | Likely Pathogenic | Ambiguous | 0.35 | Likely Benign | 0.1 | 1.01 | Ambiguous | 0.68 | Ambiguous | 0.83 | Ambiguous | 0.281 | Likely Benign | -2.00 | Neutral | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.29 | Benign | 0.01 | Affected | 0.0814 | 0.2758 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.2060G>T | R687L 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant R687L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on benign include REVEL, SIFT, ESM1b, and FATHMM, while those that agree on pathogenic are AlphaMissense‑Default, PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. The remaining tools—AlphaMissense‑Optimized, FoldX, Foldetta, and premPS—return uncertain or inconclusive results. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie, and Foldetta is uncertain. Consequently, the evidence does not strongly support either benign or pathogenic classification. The variant is therefore most likely inconclusive, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.054297 | Structured | 0.191060 | Uncertain | 0.914 | 0.259 | 0.000 | -6.925 | Likely Benign | 0.901 | Likely Pathogenic | Ambiguous | 1.43 | Ambiguous | 0.3 | 0.05 | Likely Benign | 0.74 | Ambiguous | 0.83 | Ambiguous | 0.448 | Likely Benign | -5.76 | Deleterious | 1.000 | Probably Damaging | 0.987 | Probably Damaging | 3.90 | Benign | 0.10 | Tolerated | 0.1252 | 0.3376 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||
| c.2065C>A | L689I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L689I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar, while those that predict a pathogenic effect are polyPhen2_HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also unavailable. No folding‑stability evidence supports a deleterious change. Overall, the balance of evidence slightly favors a benign interpretation, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.227227 | Uncertain | 0.963 | 0.248 | 0.000 | -11.196 | Likely Pathogenic | 0.677 | Likely Pathogenic | Likely Benign | 1.71 | Ambiguous | 0.1 | 1.12 | Ambiguous | 1.42 | Ambiguous | 0.85 | Ambiguous | 0.180 | Likely Benign | -1.97 | Neutral | 0.822 | Possibly Damaging | 0.381 | Benign | 3.44 | Benign | 0.00 | Affected | 0.0921 | 0.3479 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2068T>G | S690A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S690A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta reports an uncertain stability change, so these results are treated as unavailable. Overall, the balance of evidence favors a benign interpretation, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.055536 | Structured | 0.247926 | Uncertain | 0.944 | 0.253 | 0.000 | -8.763 | Likely Pathogenic | 0.318 | Likely Benign | Likely Benign | -0.82 | Ambiguous | 0.0 | -2.17 | Stabilizing | -1.50 | Ambiguous | 0.45 | Likely Benign | 0.217 | Likely Benign | -2.55 | Deleterious | 0.887 | Possibly Damaging | 0.738 | Possibly Damaging | 3.45 | Benign | 0.21 | Tolerated | 0.4619 | 0.3131 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||
| c.2074C>A | L692M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L692M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2) and Foldetta (combining FoldX‑MD and Rosetta outputs) is also inconclusive. Thus, the overall evidence slightly favors pathogenicity, with a majority of standard tools predicting a deleterious impact. The variant is most likely pathogenic based on current predictions, and this assessment does not contradict the ClinVar status, which has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.064632 | Structured | 0.295225 | Uncertain | 0.966 | 0.243 | 0.000 | -9.659 | Likely Pathogenic | 0.746 | Likely Pathogenic | Likely Benign | 0.49 | Likely Benign | 0.0 | 1.81 | Ambiguous | 1.15 | Ambiguous | 1.01 | Destabilizing | 0.302 | Likely Benign | -1.99 | Neutral | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.07 | Benign | 0.01 | Affected | 0.0754 | 0.2585 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.2077C>T | H693Y 2D AIThe SynGAP1 H693Y missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, premPS, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain predictions come from Rosetta, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.073402 | Structured | 0.323991 | Uncertain | 0.964 | 0.260 | 0.000 | -7.963 | In-Between | 0.984 | Likely Pathogenic | Likely Pathogenic | -0.16 | Likely Benign | 1.7 | -1.41 | Ambiguous | -0.79 | Ambiguous | 0.10 | Likely Benign | 0.513 | Likely Pathogenic | -5.98 | Deleterious | 0.553 | Possibly Damaging | 0.046 | Benign | 3.13 | Benign | 0.02 | Affected | 0.0819 | 0.3419 | 0 | 2 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||
| c.2081C>A | A694D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A694D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, FATHMM, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are premPS, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Consequently, the variant’s functional impact remains ambiguous. The predictions do not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.127496 | Structured | 0.352199 | Uncertain | 0.938 | 0.269 | 0.000 | -9.542 | Likely Pathogenic | 0.841 | Likely Pathogenic | Ambiguous | 0.35 | Likely Benign | 0.1 | 1.04 | Ambiguous | 0.70 | Ambiguous | 1.01 | Destabilizing | 0.163 | Likely Benign | -2.47 | Neutral | 0.918 | Possibly Damaging | 0.375 | Benign | 3.48 | Benign | 0.05 | Affected | 0.2085 | 0.2216 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.2081C>T | A694V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A694V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, SIFT, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that predict a pathogenic effect are PROVEAN and polyPhen2_HumDiv, while Rosetta and ESM1b give uncertain results. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign; and Foldetta is benign. No prediction or stability result is missing or inconclusive. Overall, the majority of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.127496 | Structured | 0.352199 | Uncertain | 0.938 | 0.269 | 0.000 | -7.099 | In-Between | 0.221 | Likely Benign | Likely Benign | 0.46 | Likely Benign | 0.1 | -0.76 | Ambiguous | -0.15 | Likely Benign | 0.24 | Likely Benign | 0.149 | Likely Benign | -2.66 | Deleterious | 0.970 | Probably Damaging | 0.207 | Benign | 3.42 | Benign | 0.08 | Tolerated | 0.1238 | 0.4771 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||
| c.2083C>A | L695I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L695I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, and FATHMM, while polyPhen‑2 (HumDiv and HumVar) and ESM1b predict a pathogenic outcome. The remaining tools—AlphaMissense‑Default, Foldetta, premPS, and Rosetta—return uncertain results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, whereas Foldetta remains uncertain. Overall, the majority of reliable predictions support a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.118441 | Structured | 0.373419 | Uncertain | 0.942 | 0.258 | 0.000 | -12.096 | Likely Pathogenic | 0.395 | Ambiguous | Likely Benign | 0.47 | Likely Benign | 0.1 | 0.63 | Ambiguous | 0.55 | Ambiguous | 0.93 | Ambiguous | 0.251 | Likely Benign | -2.00 | Neutral | 0.996 | Probably Damaging | 0.905 | Possibly Damaging | 3.24 | Benign | 0.08 | Tolerated | 0.0815 | 0.2759 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2083C>G | L695V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L695V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. FoldX, Rosetta, and Foldetta give uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic), and Foldetta is also unavailable. Overall, the majority of available predictions (six pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict the current ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.118441 | Structured | 0.373419 | Uncertain | 0.942 | 0.258 | 0.000 | -10.605 | Likely Pathogenic | 0.317 | Likely Benign | Likely Benign | 1.61 | Ambiguous | 0.0 | 1.57 | Ambiguous | 1.59 | Ambiguous | 1.19 | Destabilizing | 0.274 | Likely Benign | -2.99 | Deleterious | 0.993 | Probably Damaging | 0.694 | Possibly Damaging | 3.20 | Benign | 0.02 | Affected | 0.1158 | 0.2828 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||
| c.2086C>A | L696I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 L696I missense change is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. FoldX, Rosetta, Foldetta, and premPS give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is unavailable. Overall, the majority of conventional tools lean toward pathogenicity, and the high‑accuracy prediction that is available (AlphaMissense‑Optimized) indicates benign, leaving the evidence mixed. Thus, the variant is most likely pathogenic based on the preponderance of predictions, and this does not contradict the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.390093 | Uncertain | 0.962 | 0.267 | 0.000 | -10.652 | Likely Pathogenic | 0.698 | Likely Pathogenic | Likely Benign | 1.01 | Ambiguous | 0.1 | 0.58 | Ambiguous | 0.80 | Ambiguous | 0.85 | Ambiguous | 0.250 | Likely Benign | -1.86 | Neutral | 0.996 | Probably Damaging | 0.989 | Probably Damaging | 3.15 | Benign | 0.00 | Affected | 0.0921 | 0.2542 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2089T>G | W697G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 W697G missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. In contrast, a majority of tools predict a pathogenic outcome: FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar). AlphaMissense‑Default and ESM1b are uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it receives one benign, one pathogenic, and two uncertain calls. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus remains unavailable. Overall, the preponderance of evidence from multiple independent pathogenicity predictors and the protein‑stability assessment by Foldetta indicates that W697G is most likely pathogenic. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.400169 | Uncertain | 0.945 | 0.297 | 0.000 | -7.429 | In-Between | 0.480 | Ambiguous | Likely Benign | 2.57 | Destabilizing | 0.1 | 2.38 | Destabilizing | 2.48 | Destabilizing | 1.21 | Destabilizing | 0.485 | Likely Benign | -8.93 | Deleterious | 1.000 | Probably Damaging | 0.987 | Probably Damaging | 3.44 | Benign | 0.06 | Tolerated | 0.3965 | 0.1482 | -7 | -2 | 0.5 | -129.16 | ||||||||||||||||||||||||||||||
| c.2090G>C | W697S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant W697S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS, PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Four tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as unavailable due to uncertainty. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.400169 | Uncertain | 0.945 | 0.297 | 0.000 | -4.900 | Likely Benign | 0.449 | Ambiguous | Likely Benign | 1.90 | Ambiguous | 0.1 | 1.58 | Ambiguous | 1.74 | Ambiguous | 1.13 | Destabilizing | 0.322 | Likely Benign | -8.89 | Deleterious | 1.000 | Probably Damaging | 0.992 | Probably Damaging | 3.53 | Benign | 0.13 | Tolerated | 0.3738 | 0.1089 | -2 | -3 | 0.1 | -99.14 | ||||||||||||||||||||||||||||||
| c.2091G>C | W697C 2D 3DClick to see structure in 3D Viewer AISynGAP1 W697C is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized—return uncertain or inconclusive results. High‑accuracy assessments are likewise ambiguous: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta is uncertain. Consequently, the evidence does not favor either outcome; the variant’s impact remains indeterminate. This lack of consensus does not contradict ClinVar, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.400169 | Uncertain | 0.945 | 0.297 | 0.000 | -5.683 | Likely Benign | 0.811 | Likely Pathogenic | Ambiguous | 1.84 | Ambiguous | 0.1 | 1.01 | Ambiguous | 1.43 | Ambiguous | 0.98 | Ambiguous | 0.318 | Likely Benign | -8.65 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.48 | Benign | 0.10 | Tolerated | 0.3575 | 0.1015 | -8 | -2 | 3.4 | -83.07 | ||||||||||||||||||||||||||||||
| c.2091G>T | W697C 2D 3DClick to see structure in 3D Viewer AISynGAP1 W697C is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Five tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized) yield uncertain or missing results. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑vs‑2 tie; and Foldetta is uncertain. Consequently, the evidence does not strongly favor either outcome. The variant is therefore best classified as of uncertain significance; this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.400169 | Uncertain | 0.945 | 0.297 | 0.000 | -5.683 | Likely Benign | 0.811 | Likely Pathogenic | Ambiguous | 1.84 | Ambiguous | 0.1 | 1.01 | Ambiguous | 1.43 | Ambiguous | 0.98 | Ambiguous | 0.318 | Likely Benign | -8.65 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.48 | Benign | 0.10 | Tolerated | 0.3575 | 0.1015 | -8 | -2 | 3.4 | -83.07 | ||||||||||||||||||||||||||||||
| c.2095G>A | V699M 2D 3DClick to see structure in 3D Viewer AISynGAP1 variant V699M is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33441354‑G‑A). Across in silico predictors, benign calls are made by REVEL, Rosetta, Foldetta, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. Predictions that are inconclusive (FoldX, premPS, AlphaMissense‑Default) are noted but not used as evidence. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) reports benign stability. Overall, the preponderance of evidence indicates the variant is most likely benign, which does not contradict the ClinVar uncertain status but provides a stronger leaning toward benignity. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.432975 | Uncertain | 0.935 | 0.315 | 0.000 | Uncertain | 2 | 6-33441354-G-A | 8 | 4.96e-6 | -8.869 | Likely Pathogenic | 0.484 | Ambiguous | Likely Benign | -0.58 | Ambiguous | 0.1 | 0.29 | Likely Benign | -0.15 | Likely Benign | 0.96 | Ambiguous | 0.276 | Likely Benign | -2.18 | Neutral | 0.994 | Probably Damaging | 0.806 | Possibly Damaging | 3.37 | Benign | 0.03 | Affected | 3.47 | 10 | 0.0734 | 0.3071 | 2 | 1 | -2.3 | 32.06 | 257.8 | -47.2 | 0.0 | 0.0 | 0.9 | 0.1 | X | Potentially Benign | The isopropyl side chain of Val699, located on an α-helix (res. Leu685-Gln702), packs against hydrophobic residues (e.g., Leu703, Leu696, Leu435, Leu439) in the inter-helix space. In the variant simulations, the thioether side chain of Met699 has similar physicochemical properties to Val699 in the WT, and thus, it is able to maintain similar interactions. Consequently, the mutation causes no apparent changes in the structure. | ||||||||||||||
| c.2095G>C | V699L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V699L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while FoldX, premPS, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority; and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the preponderance of evidence, and this assessment does not contradict any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.432975 | Uncertain | 0.935 | 0.315 | 0.000 | -8.301 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | -0.58 | Ambiguous | 0.1 | -0.36 | Likely Benign | -0.47 | Likely Benign | 0.69 | Ambiguous | 0.150 | Likely Benign | -2.14 | Neutral | 0.448 | Benign | 0.153 | Benign | 3.48 | Benign | 0.10 | Tolerated | 0.0900 | 0.3289 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.2095G>T | V699L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V699L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while FoldX, premPS, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority; and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the consensus of the available tools, and this benign prediction does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.432975 | Uncertain | 0.935 | 0.315 | 0.000 | -8.301 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | -0.58 | Ambiguous | 0.1 | -0.36 | Likely Benign | -0.47 | Likely Benign | 0.69 | Ambiguous | 0.150 | Likely Benign | -2.14 | Neutral | 0.448 | Benign | 0.153 | Benign | 3.48 | Benign | 0.10 | Tolerated | 0.0900 | 0.3289 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.2096T>G | V699G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V699G has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include FATHMM and AlphaMissense‑Optimized, while the remaining tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b) uniformly predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic verdict; and Foldetta also predicts pathogenic. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.432975 | Uncertain | 0.935 | 0.315 | 0.000 | -11.912 | Likely Pathogenic | 0.481 | Ambiguous | Likely Benign | 2.22 | Destabilizing | 0.0 | 3.25 | Destabilizing | 2.74 | Destabilizing | 1.77 | Destabilizing | 0.514 | Likely Pathogenic | -5.11 | Deleterious | 0.972 | Probably Damaging | 0.999 | Probably Damaging | 3.37 | Benign | 0.01 | Affected | 0.1830 | 0.2240 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.2102C>A | P701H 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 P701H missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. Predictions that are uncertain or inconclusive are FoldX, Foldetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict the ClinVar status, which contains no pathogenic assertion for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.404318 | Uncertain | 0.918 | 0.345 | 0.000 | -8.786 | Likely Pathogenic | 0.553 | Ambiguous | Likely Benign | 1.40 | Ambiguous | 0.0 | -0.41 | Likely Benign | 0.50 | Ambiguous | 0.62 | Ambiguous | 0.141 | Likely Benign | -2.12 | Neutral | 0.936 | Possibly Damaging | 0.539 | Possibly Damaging | 3.38 | Benign | 0.03 | Affected | 0.1495 | 0.3495 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||
| c.2102C>G | P701R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 P701R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. Two tools (FoldX and premPS) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta (combining FoldX‑MD and Rosetta outputs) as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.404318 | Uncertain | 0.918 | 0.345 | 0.000 | -11.060 | Likely Pathogenic | 0.779 | Likely Pathogenic | Likely Benign | 0.54 | Ambiguous | 0.0 | -0.19 | Likely Benign | 0.18 | Likely Benign | 0.65 | Ambiguous | 0.088 | Likely Benign | -2.09 | Neutral | 0.784 | Possibly Damaging | 0.278 | Benign | 3.41 | Benign | 0.08 | Tolerated | 0.1365 | 0.2500 | 0 | -2 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||
| c.2102C>T | P701L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P701L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Foldetta, premPS, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and ESM1b. The remaining tools (FoldX, Rosetta, AlphaMissense‑Default) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions (7 benign vs. 3 pathogenic) and the two high‑accuracy benign calls suggest that the variant is most likely benign. This conclusion does not contradict any ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.404318 | Uncertain | 0.918 | 0.345 | 0.000 | -10.185 | Likely Pathogenic | 0.515 | Ambiguous | Likely Benign | 1.15 | Ambiguous | 0.0 | -0.68 | Ambiguous | 0.24 | Likely Benign | 0.12 | Likely Benign | 0.116 | Likely Benign | -3.04 | Deleterious | 0.642 | Possibly Damaging | 0.087 | Benign | 3.50 | Benign | 0.09 | Tolerated | 0.2018 | 0.5546 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||
| c.2104C>A | Q702K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q702K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.397258 | Uncertain | 0.907 | 0.345 | 0.000 | -8.750 | Likely Pathogenic | 0.338 | Likely Benign | Likely Benign | -0.23 | Likely Benign | 0.0 | 0.26 | Likely Benign | 0.02 | Likely Benign | 0.08 | Likely Benign | 0.224 | Likely Benign | -2.86 | Deleterious | 0.863 | Possibly Damaging | 0.773 | Possibly Damaging | 3.46 | Benign | 0.08 | Tolerated | 0.1346 | 0.2524 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||||
| c.2105A>C | Q702P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q702P is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect include Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; FoldX is uncertain and therefore not counted. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, more tools predict pathogenicity (7) than benignity (5), and the high‑accuracy pathogenic prediction from Foldetta further supports this. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.397258 | Uncertain | 0.907 | 0.345 | 0.000 | -10.361 | Likely Pathogenic | 0.324 | Likely Benign | Likely Benign | 1.76 | Ambiguous | 0.1 | 7.05 | Destabilizing | 4.41 | Destabilizing | -0.04 | Likely Benign | 0.369 | Likely Benign | -4.71 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.42 | Benign | 0.05 | Affected | 0.1877 | 0.3371 | 0 | -1 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||
| c.2105A>G | Q702R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q702R is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Predictions that remain inconclusive are Rosetta, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to equal benign and pathogenic signals. Overall, the majority of available predictions lean toward a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.397258 | Uncertain | 0.907 | 0.345 | 0.000 | Uncertain | 1 | -7.894 | In-Between | 0.348 | Ambiguous | Likely Benign | -0.31 | Likely Benign | 0.1 | 0.63 | Ambiguous | 0.16 | Likely Benign | 0.13 | Likely Benign | 0.294 | Likely Benign | -3.14 | Deleterious | 0.909 | Possibly Damaging | 0.889 | Possibly Damaging | 3.43 | Benign | 0.02 | Affected | 3.47 | 10 | 0.1225 | 0.0605 | 1 | 1 | -1.0 | 28.06 | 270.3 | -52.9 | 0.0 | 0.0 | 0.0 | 0.1 | X | Potentially Pathogenic | The carboxamide side chain of Gln702 is located at the end and outer surface of an α-helix (res. Leu685-Gln702), where it does not directly form hydrogen bonds with any residues in the WT simulations. In the variant simulations, the positively charged guanidinium group of Arg702 forms a salt bridge with the negatively charged carboxylate group of Glu698 on the same helix and/or hydrogen bonds with the backbone carbonyl group of Ala438 on an opposite α-helix (res. Tyr428-Glu436). Consequently, the residue swap could strengthen the tertiary structure assembly, which could have either positive or negative effects on its function. | |||||||||||||||||
| c.2105A>T | Q702L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q702L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the balance of evidence leans toward a benign effect. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.397258 | Uncertain | 0.907 | 0.345 | 0.000 | -9.954 | Likely Pathogenic | 0.149 | Likely Benign | Likely Benign | -0.13 | Likely Benign | 0.0 | 0.09 | Likely Benign | -0.02 | Likely Benign | 0.16 | Likely Benign | 0.392 | Likely Benign | -5.66 | Deleterious | 0.939 | Possibly Damaging | 0.838 | Possibly Damaging | 3.42 | Benign | 0.00 | Affected | 0.0584 | 0.3628 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||
| c.2106G>C | Q702H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q702H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, Rosetta, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Uncertain results come from FoldX and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign outcome (2 benign vs. 1 pathogenic vote), and Foldetta predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.397258 | Uncertain | 0.907 | 0.345 | 0.000 | -7.966 | In-Between | 0.211 | Likely Benign | Likely Benign | 0.55 | Ambiguous | 0.0 | 0.13 | Likely Benign | 0.34 | Likely Benign | 0.12 | Likely Benign | 0.233 | Likely Benign | -4.11 | Deleterious | 0.982 | Probably Damaging | 0.947 | Probably Damaging | 3.45 | Benign | 0.00 | Affected | 0.0811 | 0.2555 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.2106G>T | Q702H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q702H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, Rosetta, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Uncertain results come from FoldX and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign outcome (2 benign vs. 1 pathogenic vote), and Foldetta predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.397258 | Uncertain | 0.907 | 0.345 | 0.000 | -7.966 | In-Between | 0.211 | Likely Benign | Likely Benign | 0.55 | Ambiguous | 0.0 | 0.13 | Likely Benign | 0.34 | Likely Benign | 0.12 | Likely Benign | 0.233 | Likely Benign | -4.11 | Deleterious | 0.982 | Probably Damaging | 0.947 | Probably Damaging | 3.45 | Benign | 0.00 | Affected | 0.0811 | 0.2555 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||
| c.2107C>A | L703I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L703I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and Rosetta. Predictions that are uncertain or inconclusive (FoldX, Foldetta, premPS, AlphaMissense‑Default) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of high‑confidence tools predict a benign impact, and this conclusion does not contradict the ClinVar status, which has no pathogenic classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.144935 | Structured | 0.388282 | Uncertain | 0.929 | 0.353 | 0.000 | -9.332 | Likely Pathogenic | 0.345 | Ambiguous | Likely Benign | 1.44 | Ambiguous | 0.1 | 2.21 | Destabilizing | 1.83 | Ambiguous | 0.61 | Ambiguous | 0.108 | Likely Benign | -1.50 | Neutral | 0.982 | Probably Damaging | 0.758 | Possibly Damaging | 3.38 | Benign | 0.00 | Affected | 0.0909 | 0.3079 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2110A>T | S704C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S704C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. High‑accuracy assessments give a mixed picture: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Other stability‑based tools such as FoldX, Rosetta, and premPS also return uncertain results. Overall, the majority of individual predictors lean toward pathogenicity, and the high‑accuracy consensus also supports a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.096677 | Structured | 0.383620 | Uncertain | 0.928 | 0.363 | 0.000 | -10.438 | Likely Pathogenic | 0.423 | Ambiguous | Likely Benign | 1.56 | Ambiguous | 0.1 | 0.98 | Ambiguous | 1.27 | Ambiguous | 0.52 | Ambiguous | 0.251 | Likely Benign | -3.52 | Deleterious | 0.997 | Probably Damaging | 0.789 | Possibly Damaging | 3.40 | Benign | 0.01 | Affected | 0.0789 | 0.4612 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.2113A>G | K705E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K705E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, AlphaMissense‑Optimized, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta predicts a benign stability change. Overall, the majority of evidence (9 benign vs. 4 pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.134866 | Structured | 0.379324 | Uncertain | 0.922 | 0.364 | 0.000 | -9.371 | Likely Pathogenic | 0.704 | Likely Pathogenic | Likely Benign | 0.10 | Likely Benign | 0.0 | -0.09 | Likely Benign | 0.01 | Likely Benign | 0.47 | Likely Benign | 0.075 | Likely Benign | -1.53 | Neutral | 0.935 | Possibly Damaging | 0.537 | Possibly Damaging | 3.33 | Benign | 0.14 | Tolerated | 0.2601 | 0.0789 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||
| c.2116G>A | E706K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E706K is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM all classify the change as benign. In contrast, ESM1b and AlphaMissense‑Default predict a pathogenic impact. Tools that return uncertain results—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—do not provide decisive evidence. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign calls). High‑accuracy assessments are likewise ambiguous: AlphaMissense‑Optimized is uncertain, Foldetta is uncertain, and the SGM Consensus remains inconclusive. Overall, the preponderance of evidence points to a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.377033 | Uncertain | 0.929 | 0.363 | 0.000 | Uncertain | 1 | -10.519 | Likely Pathogenic | 0.833 | Likely Pathogenic | Ambiguous | 1.17 | Ambiguous | 0.1 | 0.51 | Ambiguous | 0.84 | Ambiguous | 0.08 | Likely Benign | 0.080 | Likely Benign | -1.51 | Neutral | 0.345 | Benign | 0.028 | Benign | 4.15 | Benign | 0.52 | Tolerated | 3.47 | 10 | 0.2065 | 0.4511 | 0 | 1 | -0.4 | -0.94 | 187.1 | 49.2 | 0.0 | 0.0 | 0.4 | 0.1 | X | Uncertain | The carboxylate side chain of Glu706, located at the end and outer surface of an α-helix (res. Thr704-Gly712), forms a salt bridge with Lys710 and a hydrogen bond with its own backbone amino group at the helix end in the WT simulations. Although Lys706 is unable to make these transient interactions in the variant simulations, there is no apparent negative effect on the protein structure due to the residue swap. However, because the model ends abruptly at the C-terminus, no definite conclusions can be drawn based on the simulations. | |||||||||||||||||
| c.2116G>C | E706Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E706Q missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are AlphaMissense‑Default and ESM1b. Rosetta and Foldetta provide uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta also yields an uncertain outcome. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.377033 | Uncertain | 0.929 | 0.363 | 0.000 | -8.171 | Likely Pathogenic | 0.588 | Likely Pathogenic | Likely Benign | 0.44 | Likely Benign | 0.0 | 0.61 | Ambiguous | 0.53 | Ambiguous | -0.14 | Likely Benign | 0.059 | Likely Benign | -1.00 | Neutral | 0.433 | Benign | 0.051 | Benign | 4.13 | Benign | 0.31 | Tolerated | 0.0941 | 0.3831 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.2120C>G | A707G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A707G missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. The remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b) give uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign, while Foldetta remains uncertain. Overall, the majority of definitive predictions lean toward a benign effect, and there is no ClinVar status to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.203355 | Structured | 0.371229 | Uncertain | 0.927 | 0.365 | 0.000 | -7.480 | In-Between | 0.224 | Likely Benign | Likely Benign | 1.45 | Ambiguous | 0.0 | 1.52 | Ambiguous | 1.49 | Ambiguous | 0.62 | Ambiguous | 0.198 | Likely Benign | -2.60 | Deleterious | 0.991 | Probably Damaging | 0.960 | Probably Damaging | 3.42 | Benign | 0.01 | Affected | 0.1678 | 0.2388 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||
| c.2122C>T | L708F 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant L708F is not reported in ClinVar and is present in gnomAD (ID 6‑33441587‑C‑T). Functional prediction tools that reach consensus classify the change as benign: REVEL, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Pathogenic predictions are limited to polyPhen‑2 HumDiv and ESM1b. The remaining tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Default) return uncertain or no result. High‑accuracy assessments give AlphaMissense‑Optimized a benign score, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, and Foldetta indicates no significant destabilization (uncertain). Overall, the preponderance of evidence supports a benign effect for L708F, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.250310 | Structured | 0.365875 | Uncertain | 0.931 | 0.378 | 0.000 | 6-33441587-C-T | 2 | 1.24e-6 | -9.154 | Likely Pathogenic | 0.436 | Ambiguous | Likely Benign | 1.48 | Ambiguous | 0.3 | 0.93 | Ambiguous | 1.21 | Ambiguous | 0.37 | Likely Benign | 0.110 | Likely Benign | -2.46 | Neutral | 0.931 | Possibly Damaging | 0.326 | Benign | 3.29 | Benign | 0.07 | Tolerated | 3.50 | 9 | 0.0497 | 0.2366 | 0 | 2 | -1.0 | 34.02 | |||||||||||||||||||||||||
| c.2126T>C | L709P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L709P is not reported in ClinVar and is absent from gnomAD. Benign predictions come from REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from FoldX, Rosetta, Foldetta, polyPhen2_HumDiv, polyPhen2_HumVar, and AlphaMissense‑Default. High‑accuracy tools give mixed results: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, FATHMM, PROVEAN) also predicts benign; Foldetta predicts pathogenic. Because the majority of standard predictors and the SGM Consensus favor benign, the variant is most likely benign, and this is consistent with the lack of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.243554 | Structured | 0.365830 | Uncertain | 0.934 | 0.379 | 0.000 | -7.517 | In-Between | 0.767 | Likely Pathogenic | Likely Benign | 2.81 | Destabilizing | 0.0 | 5.65 | Destabilizing | 4.23 | Destabilizing | 0.31 | Likely Benign | 0.167 | Likely Benign | -1.93 | Neutral | 1.000 | Probably Damaging | 0.975 | Probably Damaging | 3.47 | Benign | 0.37 | Tolerated | 0.3004 | 0.0825 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||
| c.2126T>G | L709R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L709R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. Tools with uncertain or inconclusive results are Rosetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts benign. Overall, the majority of evidence indicates a benign impact for the variant, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.243554 | Structured | 0.365830 | Uncertain | 0.934 | 0.379 | 0.000 | -8.636 | Likely Pathogenic | 0.482 | Ambiguous | Likely Benign | 0.22 | Likely Benign | 0.0 | 0.68 | Ambiguous | 0.45 | Likely Benign | 0.62 | Ambiguous | 0.061 | Likely Benign | -1.56 | Neutral | 0.906 | Possibly Damaging | 0.314 | Benign | 3.50 | Benign | 0.44 | Tolerated | 0.1127 | 0.0488 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||
| c.2128A>C | K710Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K710Q missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The high‑accuracy AlphaMissense‑Optimized score classifies the variant as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (uncertain), ESM1b (pathogenic), FATHMM (benign), and PROVEAN (pathogenic)—indicates pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, also predicts a benign effect. Overall, the majority of evidence points to a benign impact, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.321458 | Structured | 0.370438 | Uncertain | 0.949 | 0.368 | 0.000 | -8.920 | Likely Pathogenic | 0.372 | Ambiguous | Likely Benign | 0.12 | Likely Benign | 0.0 | 0.02 | Likely Benign | 0.07 | Likely Benign | 0.49 | Likely Benign | 0.183 | Likely Benign | -3.58 | Deleterious | 0.999 | Probably Damaging | 0.999 | Probably Damaging | 3.39 | Benign | 0.01 | Affected | 0.3043 | 0.1214 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||
| c.2131C>A | L711M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L711M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta reports an uncertain stability change, which is treated as unavailable evidence. Overall, the majority of individual predictors (six pathogenic vs. five benign) and the available high‑accuracy results lean toward a pathogenic interpretation. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.308712 | Structured | 0.377436 | Uncertain | 0.950 | 0.364 | 0.000 | -9.368 | Likely Pathogenic | 0.679 | Likely Pathogenic | Likely Benign | 0.39 | Likely Benign | 0.0 | 1.11 | Ambiguous | 0.75 | Ambiguous | 1.40 | Destabilizing | 0.216 | Likely Benign | -1.86 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.34 | Benign | 0.00 | Affected | 0.0755 | 0.2758 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||
| c.2134G>A | G712S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G712S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic impact comprise FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. Uncertain results come from premPS, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive and therefore unavailable. Overall, the majority of evidence points to a pathogenic effect for G712S. This conclusion does not contradict ClinVar, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.278302 | Structured | 0.384858 | Uncertain | 0.947 | 0.365 | 0.000 | -7.942 | In-Between | 0.422 | Ambiguous | Likely Benign | 2.30 | Destabilizing | 0.1 | 4.79 | Destabilizing | 3.55 | Destabilizing | 0.62 | Ambiguous | 0.282 | Likely Benign | -4.84 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.42 | Benign | 0.02 | Affected | 0.2862 | 0.4501 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||
| c.2135G>C | G712A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 G712A missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions arise from FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. Two tools report uncertainty: premPS and AlphaMissense‑Default. High‑accuracy assessments further refine the picture. AlphaMissense‑Optimized classifies the variant as benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also yields a benign verdict (2 benign vs. 1 pathogenic vote). Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a pathogenic effect. Because the majority of conventional predictors (7/11) indicate pathogenicity, the overall consensus leans toward a pathogenic impact, despite the mixed high‑accuracy results. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.278302 | Structured | 0.384858 | Uncertain | 0.947 | 0.365 | 0.000 | -6.444 | Likely Benign | 0.553 | Ambiguous | Likely Benign | 2.89 | Destabilizing | 0.1 | 4.98 | Destabilizing | 3.94 | Destabilizing | 0.70 | Ambiguous | 0.281 | Likely Benign | -4.99 | Deleterious | 0.999 | Probably Damaging | 0.991 | Probably Damaging | 3.44 | Benign | 0.02 | Affected | 0.3993 | 0.4105 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||
| c.2140C>G | L714V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L714V has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect include FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Overall, the majority of predictions (10 pathogenic vs. 4 benign) indicate a likely pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no record for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.314870 | Structured | 0.402311 | Uncertain | 0.961 | 0.369 | 0.000 | -4.966 | Likely Benign | 0.781 | Likely Pathogenic | Likely Benign | 3.91 | Destabilizing | 0.1 | 6.19 | Destabilizing | 5.05 | Destabilizing | 1.21 | Destabilizing | 0.259 | Likely Benign | -2.76 | Deleterious | 0.995 | Probably Damaging | 0.970 | Probably Damaging | 3.16 | Benign | 0.00 | Affected | 0.1421 | 0.2689 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||
| c.2143C>T | P715S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant P715S is listed in ClinVar as pathogenic (ClinVar ID 1804065.0) and is present in gnomAD (ID 6‑33441608‑C‑T). Functional prediction tools that agree on a benign effect are REVEL and FATHMM. Those that predict a pathogenic effect include FoldX, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. Predictions that are inconclusive are Rosetta, premPS, ESM1b, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of evidence points to a pathogenic impact, which is consistent with the ClinVar classification and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.243554 | Structured | 0.409757 | Uncertain | 0.956 | 0.362 | 0.000 | Likely Pathogenic | 1 | 6-33441608-C-T | 1 | 6.20e-7 | -7.635 | In-Between | 0.787 | Likely Pathogenic | Ambiguous | 3.54 | Destabilizing | 0.0 | 0.81 | Ambiguous | 2.18 | Destabilizing | 0.94 | Ambiguous | 0.277 | Likely Benign | -7.17 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.43 | Benign | 0.01 | Affected | 3.50 | 9 | 0.2977 | 0.3755 | 1 | -1 | 0.8 | -10.04 | 231.8 | -14.0 | -0.1 | 0.0 | -0.8 | 0.1 | X | Uncertain | Pro715, along with Gly712 and Pro713, are located in a hinge region of an α-helix making a ~90-degree turn (res. Lys705-Leu725). In the WT simulations, the pyrrolidine side chain of Pro715, lacking the backbone amide groups altogether, forces the tight helix turn to take place while also hydrophobically packing with nearby residues (e.g., Leu700, Leu708, Leu714, and Leu718). Leu715, with a normal amide backbone, could potentially affect protein folding and turn formation, although this was not observed in the variant simulations. Additionally, the hydroxyl group of the Ser715 side chain can form hydrogen bonds with the backbone carbonyl group of Gly712 and disrupt the hydrophobic packing arrangement of the leucine residues from the neighboring α-helices, impacting the GAP domain tertiary assembly. | ||||||||||||||
| c.2147G>A | R716Q 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant R716Q is listed in ClinVar with an uncertain significance (ClinVar ID 411585.0) and is present in gnomAD (ID 6‑33441612‑G‑A). Functional prediction tools that report a benign effect include REVEL, FoldX, Rosetta, Foldetta, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b, while premPS is inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the balance of evidence leans toward a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.247041 | Structured | 0.419135 | Uncertain | 0.962 | 0.379 | 0.000 | Conflicting | 2 | 6-33441612-G-A | 4 | 2.48e-6 | -8.338 | Likely Pathogenic | 0.308 | Likely Benign | Likely Benign | -0.01 | Likely Benign | 0.0 | 0.47 | Likely Benign | 0.23 | Likely Benign | 0.58 | Ambiguous | 0.210 | Likely Benign | -3.14 | Deleterious | 1.000 | Probably Damaging | 0.990 | Probably Damaging | 3.35 | Benign | 0.02 | Affected | 3.50 | 9 | 0.2834 | 0.2180 | 1 | 1 | 1.0 | -28.06 | 250.0 | 48.9 | 0.0 | 0.0 | -0.5 | 0.0 | X | Uncertain | The guanidinium group of Arg716, located on the outer surface of an α-helix (res. Leu714-Arg726), forms a salt bridge with the carboxylate group of Asp720. In the variant simulations, the carboxamide group of Gln716 also forms a hydrogen bond with the carboxylate group of Asp720, although this bond is weaker than the Arg716 salt bridge in the WT. Overall, no adverse effects on the protein structure are observed in the simulations. However, because the model ends abruptly at the C-terminus, no definite conclusions can be drawn based on the simulations. | ||||||||||||||
| c.2155A>G | N719D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N719D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FoldX, premPS, SIFT, FATHMM, AlphaMissense‑Optimized, and Foldetta. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools give uncertain results: Rosetta and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of individual predictors (seven benign vs. four pathogenic) support a benign effect, and this conclusion does not contradict the lack of ClinVar annotation. Thus, based on the available predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.384043 | Structured | 0.445381 | Uncertain | 0.961 | 0.386 | 0.000 | -9.016 | Likely Pathogenic | 0.416 | Ambiguous | Likely Benign | -0.03 | Likely Benign | 0.0 | 0.64 | Ambiguous | 0.31 | Likely Benign | 0.46 | Likely Benign | 0.106 | Likely Benign | -2.56 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.75 | Benign | 0.84 | Tolerated | 0.1581 | 0.2345 | 2 | 1 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.2155A>T | N719Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N719Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Rosetta and Foldetta provide uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is also uncertain. Overall, the majority of evaluated tools (7 benign vs 4 pathogenic) support a benign classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.384043 | Structured | 0.445381 | Uncertain | 0.961 | 0.386 | 0.000 | -11.005 | Likely Pathogenic | 0.302 | Likely Benign | Likely Benign | -0.38 | Likely Benign | 0.0 | -0.62 | Ambiguous | -0.50 | Ambiguous | 0.33 | Likely Benign | 0.187 | Likely Benign | -4.15 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.70 | Benign | 0.09 | Tolerated | 0.0412 | 0.3951 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||
| c.2156A>T | N719I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N719I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools report uncertainty: Rosetta and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of predictions lean toward a benign impact, with no conflict with ClinVar status. Thus, the variant is most likely benign based on the current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.384043 | Structured | 0.445381 | Uncertain | 0.961 | 0.386 | 0.000 | -10.794 | Likely Pathogenic | 0.399 | Ambiguous | Likely Benign | -0.19 | Likely Benign | 0.0 | -0.74 | Ambiguous | -0.47 | Likely Benign | 0.40 | Likely Benign | 0.146 | Likely Benign | -4.88 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.71 | Benign | 0.10 | Tolerated | 0.0408 | 0.4493 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||
| c.2157C>A | N719K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N719K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, ESM1b, and Foldetta) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of reliable predictions indicate a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.384043 | Structured | 0.445381 | Uncertain | 0.961 | 0.386 | 0.000 | -7.454 | In-Between | 0.651 | Likely Pathogenic | Likely Benign | -0.64 | Ambiguous | 0.0 | -0.62 | Ambiguous | -0.63 | Ambiguous | 0.42 | Likely Benign | 0.250 | Likely Benign | -1.84 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.80 | Benign | 0.55 | Tolerated | 0.1424 | 0.3112 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||
| c.2157C>G | N719K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N719K is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in silico predictors shows a predominance of benign calls: REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized all predict a neutral effect, whereas polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default predict pathogenicity. FoldX, Rosetta, ESM1b, and Foldetta are inconclusive. High‑accuracy assessment further supports a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign outcome; Foldetta remains uncertain. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.384043 | Structured | 0.445381 | Uncertain | 0.961 | 0.386 | 0.000 | -7.454 | In-Between | 0.651 | Likely Pathogenic | Likely Benign | -0.64 | Ambiguous | 0.0 | -0.62 | Ambiguous | -0.63 | Ambiguous | 0.42 | Likely Benign | 0.244 | Likely Benign | -1.84 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.80 | Benign | 0.55 | Tolerated | 0.1424 | 0.3112 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||
| c.2160C>A | D720E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D720E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority, and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.374039 | Structured | 0.450695 | Uncertain | 0.955 | 0.417 | 0.125 | -5.527 | Likely Benign | 0.391 | Ambiguous | Likely Benign | -0.31 | Likely Benign | 0.1 | -0.41 | Likely Benign | -0.36 | Likely Benign | 0.23 | Likely Benign | 0.131 | Likely Benign | -2.46 | Neutral | 0.975 | Probably Damaging | 0.969 | Probably Damaging | 2.46 | Pathogenic | 0.15 | Tolerated | 0.1310 | 0.5275 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||
| c.2160C>G | D720E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D720E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority, and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.374039 | Structured | 0.450695 | Uncertain | 0.955 | 0.417 | 0.125 | -5.527 | Likely Benign | 0.391 | Ambiguous | Likely Benign | -0.31 | Likely Benign | 0.1 | -0.41 | Likely Benign | -0.36 | Likely Benign | 0.23 | Likely Benign | 0.131 | Likely Benign | -2.46 | Neutral | 0.975 | Probably Damaging | 0.969 | Probably Damaging | 2.46 | Pathogenic | 0.15 | Tolerated | 0.1310 | 0.5275 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||
| c.2161A>C | I721L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I721L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. Two tools (premPS and ESM1b) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the predictions are split, but the majority of high‑confidence tools lean toward a benign interpretation. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.394753 | Structured | 0.454550 | Uncertain | 0.957 | 0.437 | 0.125 | -7.843 | In-Between | 0.757 | Likely Pathogenic | Likely Benign | 0.35 | Likely Benign | 0.0 | 0.34 | Likely Benign | 0.35 | Likely Benign | 0.65 | Ambiguous | 0.188 | Likely Benign | -1.83 | Neutral | 0.955 | Possibly Damaging | 0.985 | Probably Damaging | 2.37 | Pathogenic | 0.02 | Affected | 0.0627 | 0.3446 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2173C>A | L725M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L725M is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Uncertain or inconclusive predictions come from Foldetta, premPS, AlphaMissense‑Default, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans pathogenic (2 pathogenic vs. 1 benign). Foldetta remains uncertain and is not used as evidence. Overall, the majority of tools, including the high‑accuracy consensus, predict a pathogenic impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.557691 | Disordered | 0.455613 | Uncertain | 0.911 | 0.491 | 0.625 | -9.531 | Likely Pathogenic | 0.436 | Ambiguous | Likely Benign | 0.42 | Likely Benign | 0.1 | 1.05 | Ambiguous | 0.74 | Ambiguous | 0.80 | Ambiguous | 0.172 | Likely Benign | -1.81 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.32 | Pathogenic | 0.04 | Affected | 0.0854 | 0.4156 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||
| c.2177G>T | R726M 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R726M has no ClinVar entry and is not reported in gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, Rosetta, Foldetta, premPS, PROVEAN, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. FoldX is uncertain and is not counted as evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of tools and the two high‑accuracy methods predict a benign effect. Therefore, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.521092 | Disordered | 0.449098 | Uncertain | 0.888 | 0.513 | 0.625 | -8.611 | Likely Pathogenic | 0.750 | Likely Pathogenic | Likely Benign | 0.53 | Ambiguous | 0.1 | 0.31 | Likely Benign | 0.42 | Likely Benign | 0.20 | Likely Benign | 0.199 | Likely Benign | -2.02 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.59 | Benign | 0.03 | Affected | 0.1489 | 0.4051 | 0 | -1 | 6.4 | -24.99 | |||||||||||||||||||||||||||||||
| c.2179A>C | N727H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N727H is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Two tools (premPS and ESM1b) return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions (six benign vs. five pathogenic) lean toward a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.538167 | Disordered | 0.442107 | Uncertain | 0.843 | 0.542 | 0.625 | -7.308 | In-Between | 0.224 | Likely Benign | Likely Benign | 0.13 | Likely Benign | 0.0 | -0.02 | Likely Benign | 0.06 | Likely Benign | 0.51 | Ambiguous | 0.171 | Likely Benign | -3.18 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.13 | Pathogenic | 0.03 | Affected | 0.1320 | 0.7186 | 2 | 1 | 0.3 | 23.04 | |||||||||||||||||||||||||||||||
| c.2180A>C | N727T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N727T is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include REVEL, Rosetta, premPS, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. FoldX gives an uncertain result and is therefore treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta (combining FoldX‑MD and Rosetta outputs) as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.538167 | Disordered | 0.442107 | Uncertain | 0.843 | 0.542 | 0.625 | -6.900 | Likely Benign | 0.335 | Likely Benign | Likely Benign | 0.52 | Ambiguous | 0.1 | -0.18 | Likely Benign | 0.17 | Likely Benign | 0.04 | Likely Benign | 0.125 | Likely Benign | -3.08 | Deleterious | 0.987 | Probably Damaging | 0.980 | Probably Damaging | 2.25 | Pathogenic | 0.74 | Tolerated | 0.1315 | 0.7181 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||||
| c.2180A>G | N727S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N727S is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence supports a benign effect. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.538167 | Disordered | 0.442107 | Uncertain | 0.843 | 0.542 | 0.625 | -6.195 | Likely Benign | 0.184 | Likely Benign | Likely Benign | 0.32 | Likely Benign | 0.1 | 0.28 | Likely Benign | 0.30 | Likely Benign | 0.18 | Likely Benign | 0.118 | Likely Benign | -2.67 | Deleterious | 0.999 | Probably Damaging | 0.979 | Probably Damaging | 2.19 | Pathogenic | 0.23 | Tolerated | 0.3833 | 0.6680 | 1 | 1 | 2.7 | -27.03 | |||||||||||||||||||||||||||||||
| c.2189T>A | I730N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I730N is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar) and SIFT. The remaining tools—premPS, ESM1b, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts benign. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.733139 | Disordered | 0.420109 | Uncertain | 0.591 | 0.619 | 0.750 | -7.987 | In-Between | 0.369 | Ambiguous | Likely Benign | 0.09 | Likely Benign | 0.2 | 0.04 | Likely Benign | 0.07 | Likely Benign | 0.70 | Ambiguous | 0.080 | Likely Benign | -1.60 | Neutral | 1.000 | Probably Damaging | 0.986 | Probably Damaging | 3.46 | Benign | 0.02 | Affected | 0.0969 | 0.0533 | -2 | -3 | -8.0 | 0.94 | |||||||||||||||||||||||||||||||
| c.2194A>G | R732G 2D AIThe SynGAP1 R732G missense variant is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (ID 6‑33441659‑A‑G). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, whereas tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs. 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of standard predictors lean toward pathogenicity, while the most accurate single predictor (AlphaMissense‑Optimized) suggests a benign outcome. Given the lack of ClinVar evidence, there is no contradiction; the variant is most likely pathogenic based on the collective predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.846163 | Disordered | 0.412403 | Uncertain | 0.427 | 0.673 | 0.750 | 6-33441659-A-G | 1 | 6.20e-7 | -9.348 | Likely Pathogenic | 0.295 | Likely Benign | Likely Benign | 0.145 | Likely Benign | -2.98 | Deleterious | 1.000 | Probably Damaging | 0.982 | Probably Damaging | 2.56 | Benign | 0.03 | Affected | 3.59 | 7 | 0.3080 | 0.2942 | -2 | -3 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||
| c.2194A>T | R732W 2D AIThe SynGAP1 missense variant R732W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, five tools predict pathogenicity versus four predicting benign, with no ClinVar evidence to contradict these computational findings. Thus, the variant is most likely pathogenic based on the current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.846163 | Disordered | 0.412403 | Uncertain | 0.427 | 0.673 | 0.750 | -11.976 | Likely Pathogenic | 0.252 | Likely Benign | Likely Benign | 0.161 | Likely Benign | -3.57 | Deleterious | 1.000 | Probably Damaging | 0.973 | Probably Damaging | 2.53 | Benign | 0.01 | Affected | 0.1332 | 0.2748 | 2 | -3 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2195G>C | R732T 2D AISynGAP1 missense variant R732T is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign (REVEL, PROVEAN, SIFT, FATHMM, AlphaMissense‑Optimized) and pathogenic (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b). AlphaMissense‑Default remains uncertain. The high‑accuracy AlphaMissense‑Optimized predicts a benign effect, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors a benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a benign impact, which does not contradict the current ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.846163 | Disordered | 0.412403 | Uncertain | 0.427 | 0.673 | 0.750 | Uncertain | 1 | -8.545 | Likely Pathogenic | 0.434 | Ambiguous | Likely Benign | 0.075 | Likely Benign | -1.96 | Neutral | 0.999 | Probably Damaging | 0.892 | Possibly Damaging | 2.59 | Benign | 0.12 | Tolerated | 3.59 | 7 | 0.1915 | 0.3153 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||
| c.2195G>T | R732M 2D AIThe SynGAP1 missense variant R732M is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and ESM1b. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome. Foldetta results are unavailable. Overall, the balance of evidence (5 benign vs 3 pathogenic predictions) indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.846163 | Disordered | 0.412403 | Uncertain | 0.427 | 0.673 | 0.750 | -9.956 | Likely Pathogenic | 0.414 | Ambiguous | Likely Benign | 0.098 | Likely Benign | -1.42 | Neutral | 0.840 | Possibly Damaging | 0.357 | Benign | 2.55 | Benign | 0.01 | Affected | 0.1432 | 0.2980 | 0 | -1 | 6.4 | -24.99 | ||||||||||||||||||||||||||||||||||||||||
| c.2196G>C | R732S 2D AIThe SynGAP1 missense variant R732S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the balance of evidence (five benign versus four pathogenic predictions) favors a benign classification. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.846163 | Disordered | 0.412403 | Uncertain | 0.427 | 0.673 | 0.750 | -8.019 | Likely Pathogenic | 0.599 | Likely Pathogenic | Likely Benign | 0.115 | Likely Benign | -1.81 | Neutral | 1.000 | Probably Damaging | 0.982 | Probably Damaging | 2.61 | Benign | 0.14 | Tolerated | 0.2850 | 0.3129 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2196G>T | R732S 2D AIThe SynGAP1 missense variant R732S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign impact for R732S, and this conclusion does not contradict any ClinVar annotation because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.846163 | Disordered | 0.412403 | Uncertain | 0.427 | 0.673 | 0.750 | -8.019 | Likely Pathogenic | 0.599 | Likely Pathogenic | Likely Benign | 0.115 | Likely Benign | -1.81 | Neutral | 1.000 | Probably Damaging | 0.982 | Probably Damaging | 2.61 | Benign | 0.14 | Tolerated | 0.2850 | 0.3129 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2206C>T | R736C 2D  AISynGAP1 missense variant R736C is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33441671‑C‑T). Functional prediction tools cluster into two groups: benign predictions from REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM, while ESM1b remains uncertain. High‑accuracy assessments reinforce the benign trend: AlphaMissense‑Optimized scores benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also returns benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence indicates a benign effect, which does not conflict with the ClinVar uncertain designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.926919 | Disordered | 0.415259 | Uncertain | 0.305 | 0.771 | 0.875 | Conflicting | 3 | 6-33441671-C-T | 8 | 4.96e-6 | -7.113 | In-Between | 0.120 | Likely Benign | Likely Benign | 0.190 | Likely Benign | -2.06 | Neutral | 0.999 | Probably Damaging | 0.825 | Possibly Damaging | 2.48 | Pathogenic | 0.00 | Affected | 4.07 | 3 | 0.3740 | 0.1691 | -4 | -3 | 7.0 | -53.05 | |||||||||||||||||||||||||||||||||
| c.2216A>G | E739G 2D AIThe SynGAP1 missense variant E739G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of predictions (5 benign vs 4 pathogenic) lean toward a benign impact, and this is consistent with the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.456400 | Uncertain | 0.313 | 0.834 | 0.875 | -3.104 | Likely Benign | 0.154 | Likely Benign | Likely Benign | 0.095 | Likely Benign | -2.53 | Deleterious | 0.625 | Possibly Damaging | 0.252 | Benign | 2.49 | Pathogenic | 0.00 | Affected | 0.3335 | 0.5983 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2218C>T | R740W 2D AIThe SynGAP1 missense variant R740W is listed in ClinVar with an uncertain significance and is present in the gnomAD database (ID 6‑33441683‑C‑T). Prediction tools that classify the variant as benign include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while those that predict pathogenicity are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized predicting a benign effect; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two benign vs. two pathogenic calls) and is treated as unavailable, and no Foldetta stability data are reported. Overall, the majority of conventional tools (five pathogenic vs. four benign) suggest a pathogenic impact, whereas the single high‑accuracy tool indicates benign. Thus, the variant is most likely pathogenic based on the aggregate predictions, and this assessment does not contradict the ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.771762 | Disordered | 0.475392 | Uncertain | 0.269 | 0.849 | 0.875 | Uncertain | 2 | 6-33441683-C-T | 6 | 3.72e-6 | -8.561 | Likely Pathogenic | 0.168 | Likely Benign | Likely Benign | 0.180 | Likely Benign | -3.09 | Deleterious | 1.000 | Probably Damaging | 0.938 | Probably Damaging | 2.52 | Benign | 0.01 | Affected | 4.32 | 2 | 0.1566 | 0.3375 | 2 | -3 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||
| c.2237T>G | V746G 2D AIThe SynGAP1 missense variant V746G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized scores benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective predictions, V746G is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.808535 | Disordered | 0.576597 | Binding | 0.336 | 0.867 | 0.875 | -2.971 | Likely Benign | 0.113 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -0.89 | Neutral | 0.136 | Benign | 0.270 | Benign | 2.74 | Benign | 0.02 | Affected | 0.1897 | 0.2550 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2240T>G | V747G 2D AIThe SynGAP1 missense variant V747G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is benign. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as benign, and Foldetta results are unavailable. Overall, the consensus of the available predictions indicates that V747G is most likely benign, and this conclusion does not contradict any ClinVar status because the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.816150 | Disordered | 0.594069 | Binding | 0.343 | 0.873 | 0.750 | -3.883 | Likely Benign | 0.188 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -1.22 | Neutral | 0.589 | Possibly Damaging | 0.917 | Probably Damaging | 2.56 | Benign | 0.00 | Affected | 0.1960 | 0.2374 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2245C>T | R749W 2D AIThe SynGAP1 missense variant R749W is listed in ClinVar as benign and is observed in gnomAD (ID 6‑33441710‑C‑T). Prediction tools that classify the variant as benign include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, whereas pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also returns benign, and Foldetta stability analysis is unavailable. Overall, the majority of evidence, especially from high‑confidence methods, supports a benign effect. This consensus aligns with the ClinVar designation, so there is no contradiction between the predictions and the reported clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.675549 | Disordered | 0.626050 | Binding | 0.337 | 0.860 | 0.625 | Likely Benign | 1 | 6-33441710-C-T | 3 | 1.86e-6 | -7.647 | In-Between | 0.338 | Likely Benign | Likely Benign | 0.173 | Likely Benign | -2.62 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.59 | Benign | 0.00 | Affected | 4.32 | 2 | 0.1215 | 0.4088 | 2 | -3 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||
| c.2248G>A | G750R 2D AIThe SynGAP1 missense variant G750R is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33441713‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs four benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.618285 | Disordered | 0.646832 | Binding | 0.348 | 0.866 | 0.625 | 6-33441713-G-A | 17 | 1.05e-5 | -3.861 | Likely Benign | 0.619 | Likely Pathogenic | Likely Benign | 0.179 | Likely Benign | -2.09 | Neutral | 1.000 | Probably Damaging | 0.982 | Probably Damaging | 2.45 | Pathogenic | 0.00 | Affected | 3.99 | 5 | 0.0960 | 0.4015 | -2 | -3 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||
| c.2248G>C | G750R 2D AIThe SynGAP1 missense variant G750R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized predicts benign, the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta data are unavailable. Overall, the majority of standard predictors (5 pathogenic vs 4 benign) lean toward pathogenicity, and no ClinVar annotation contradicts this assessment. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.618285 | Disordered | 0.646832 | Binding | 0.348 | 0.866 | 0.625 | -3.861 | Likely Benign | 0.619 | Likely Pathogenic | Likely Benign | 0.179 | Likely Benign | -2.09 | Neutral | 1.000 | Probably Damaging | 0.982 | Probably Damaging | 2.45 | Pathogenic | 0.00 | Affected | 3.99 | 5 | 0.0960 | 0.4015 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||
| c.2249G>A | G750E 2D AISynGAP1 missense variant G750E is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.618285 | Disordered | 0.646832 | Binding | 0.348 | 0.866 | 0.625 | Uncertain | 1 | -2.618 | Likely Benign | 0.413 | Ambiguous | Likely Benign | 0.146 | Likely Benign | -2.27 | Neutral | 1.000 | Probably Damaging | 0.982 | Probably Damaging | 2.49 | Pathogenic | 0.01 | Affected | 3.99 | 5 | 0.1326 | 0.3768 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||||||||
| c.2249G>T | G750V 2D AIThe SynGAP1 missense variant G750V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions therefore show AlphaMissense‑Optimized as benign, while the remaining high‑confidence tools (PROVEAN, polyPhen‑2, SIFT, FATHMM) all indicate pathogenicity. Overall, the majority of evidence (five pathogenic vs four benign) points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.618285 | Disordered | 0.646832 | Binding | 0.348 | 0.866 | 0.625 | -4.512 | Likely Benign | 0.134 | Likely Benign | Likely Benign | 0.145 | Likely Benign | -2.54 | Deleterious | 0.984 | Probably Damaging | 0.850 | Possibly Damaging | 2.45 | Pathogenic | 0.01 | Affected | 0.1270 | 0.4212 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2260G>A | E754K 2D AIThe SynGAP1 missense variant E754K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Consensus among in‑silico predictors shows a predominance of benign calls: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized all predict a benign effect, whereas polyPhen‑2 HumDiv and AlphaMissense‑Default predict pathogenicity; ESM1b remains uncertain. High‑accuracy assessment further supports a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta data are unavailable. Consequently, the variant is most likely benign according to the aggregate predictions, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.750531 | Binding | 0.357 | 0.872 | 0.500 | -7.620 | In-Between | 0.610 | Likely Pathogenic | Likely Benign | 0.138 | Likely Benign | -1.33 | Neutral | 0.801 | Possibly Damaging | 0.412 | Benign | 2.50 | Benign | 0.26 | Tolerated | 0.2159 | 0.7136 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.2261A>C | E754A 2D AIThe SynGAP1 missense variant E754A is not reported in ClinVar (ClinVar status: not reported) and is absent from gnomAD (gnomAD: not present). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction (2 benign vs. 1 pathogenic votes). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.750531 | Binding | 0.357 | 0.872 | 0.500 | -4.688 | Likely Benign | 0.381 | Ambiguous | Likely Benign | 0.049 | Likely Benign | -1.56 | Neutral | 0.801 | Possibly Damaging | 0.412 | Benign | 2.49 | Pathogenic | 0.31 | Tolerated | 0.3549 | 0.6283 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2261A>T | E754V 2D  AIThe SynGAP1 missense variant E754V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence (five benign versus four pathogenic predictions) and the high‑accuracy benign call suggest that the variant is most likely benign. This conclusion does not contradict ClinVar status, as the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.750531 | Binding | 0.357 | 0.872 | 0.500 | -6.147 | Likely Benign | 0.601 | Likely Pathogenic | Likely Benign | 0.157 | Likely Benign | -1.86 | Neutral | 0.966 | Probably Damaging | 0.773 | Possibly Damaging | 2.45 | Pathogenic | 0.28 | Tolerated | 0.0730 | 0.7417 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2282G>T | R761L 2D AIThe SynGAP1 missense variant R761L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact. This consensus does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.538167 | Disordered | 0.900613 | Binding | 0.353 | 0.865 | 0.250 | -5.653 | Likely Benign | 0.718 | Likely Pathogenic | Likely Benign | 0.171 | Likely Benign | -2.51 | Deleterious | 0.992 | Probably Damaging | 0.900 | Possibly Damaging | 2.70 | Benign | 0.24 | Tolerated | 0.1786 | 0.4326 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2284G>A | D762N 2D AIThe SynGAP1 D762N missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign impact for D762N, and this conclusion does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.405110 | Structured | 0.910475 | Binding | 0.308 | 0.859 | 0.125 | -3.323 | Likely Benign | 0.640 | Likely Pathogenic | Likely Benign | 0.110 | Likely Benign | -1.51 | Neutral | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.13 | Pathogenic | 0.11 | Tolerated | 0.1698 | 0.8797 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2285A>C | D762A 2D AIThe SynGAP1 D762A missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool yields an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward a pathogenic impact. This assessment does not contradict any ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.405110 | Structured | 0.910475 | Binding | 0.308 | 0.859 | 0.125 | -4.510 | Likely Benign | 0.912 | Likely Pathogenic | Ambiguous | 0.178 | Likely Benign | -2.40 | Neutral | 0.994 | Probably Damaging | 0.900 | Possibly Damaging | 2.12 | Pathogenic | 0.05 | Affected | 0.4632 | 0.8428 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2285A>T | D762V 2D AIThe SynGAP1 D762V missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b. Tools that agree on a pathogenic effect include polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returned an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of consensus tools predict a pathogenic impact, and there is no ClinVar entry to contradict this assessment. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.405110 | Structured | 0.910475 | Binding | 0.308 | 0.859 | 0.125 | -6.122 | Likely Benign | 0.949 | Likely Pathogenic | Ambiguous | 0.229 | Likely Benign | -1.98 | Neutral | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.08 | Pathogenic | 0.01 | Affected | 0.1105 | 0.8785 | -2 | -3 | 7.7 | -15.96 | ||||||||||||||||||||||||||||||||||||||||
| c.2286C>A | D762E 2D AIThe SynGAP1 D762E missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, and Foldetta results are unavailable. Overall, the majority of high‑confidence predictions lean toward a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.405110 | Structured | 0.910475 | Binding | 0.308 | 0.859 | 0.125 | -4.332 | Likely Benign | 0.459 | Ambiguous | Likely Benign | 0.144 | Likely Benign | -1.59 | Neutral | 0.994 | Probably Damaging | 0.891 | Possibly Damaging | 2.18 | Pathogenic | 0.05 | Affected | 0.1911 | 0.8236 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2286C>G | D762E 2D AIThe SynGAP1 D762E missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, and Foldetta results are unavailable. Overall, the majority of high‑confidence predictions lean toward a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.405110 | Structured | 0.910475 | Binding | 0.308 | 0.859 | 0.125 | -4.332 | Likely Benign | 0.459 | Ambiguous | Likely Benign | 0.144 | Likely Benign | -1.59 | Neutral | 0.994 | Probably Damaging | 0.891 | Possibly Damaging | 2.18 | Pathogenic | 0.05 | Affected | 0.1911 | 0.8236 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2288T>A | L763H 2D  AIThe SynGAP1 missense variant L763H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on benign include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the predictions are mixed, with a slight tilt toward pathogenicity (5 pathogenic vs. 4 benign). Thus, the variant is most likely pathogenic according to the aggregate predictions, and this assessment does not contradict ClinVar, which currently has no entry for it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.380708 | Structured | 0.918636 | Binding | 0.351 | 0.865 | 0.125 | -6.681 | Likely Benign | 0.640 | Likely Pathogenic | Likely Benign | 0.180 | Likely Benign | -0.79 | Neutral | 1.000 | Probably Damaging | 0.992 | Probably Damaging | 2.36 | Pathogenic | 0.03 | Affected | 0.1021 | 0.0887 | -2 | -3 | -7.0 | 23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2288T>C | L763P 2D AIThe SynGAP1 missense variant L763P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. AlphaMissense‑Default is uncertain. For high‑accuracy assessment, AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also yields a benign prediction (2 benign vs. 1 pathogenic, with one uncertain). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.380708 | Structured | 0.918636 | Binding | 0.351 | 0.865 | 0.125 | -5.802 | Likely Benign | 0.550 | Ambiguous | Likely Benign | 0.158 | Likely Benign | -0.89 | Neutral | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.36 | Pathogenic | 0.12 | Tolerated | 0.3920 | 0.1182 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2288T>G | L763R 2D AIThe SynGAP1 missense variant L763R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.380708 | Structured | 0.918636 | Binding | 0.351 | 0.865 | 0.125 | -5.516 | Likely Benign | 0.643 | Likely Pathogenic | Likely Benign | 0.163 | Likely Benign | -1.66 | Neutral | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.38 | Pathogenic | 0.07 | Tolerated | 0.1199 | 0.0761 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2290A>T | N764Y 2D AIThe SynGAP1 missense variant N764Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy methods give mixed results: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta data are unavailable. Overall, the balance of evidence favors a pathogenic interpretation, with no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.380708 | Structured | 0.919527 | Binding | 0.305 | 0.861 | 0.250 | -5.914 | Likely Benign | 0.680 | Likely Pathogenic | Likely Benign | 0.158 | Likely Benign | -2.85 | Deleterious | 0.998 | Probably Damaging | 0.967 | Probably Damaging | 2.58 | Benign | 0.01 | Affected | 0.0552 | 0.4175 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||||||||||||
| c.2291A>T | N764I 2D AIThe SynGAP1 missense variant N764I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, more tools predict pathogenicity (five) than benignity (three), and no ClinVar evidence contradicts this assessment. Thus, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.380708 | Structured | 0.919527 | Binding | 0.305 | 0.861 | 0.250 | -6.879 | Likely Benign | 0.883 | Likely Pathogenic | Ambiguous | 0.115 | Likely Benign | -2.58 | Deleterious | 0.906 | Possibly Damaging | 0.679 | Possibly Damaging | 2.58 | Benign | 0.00 | Affected | 0.0581 | 0.4483 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.2296T>C | S766P 2D AIThe SynGAP1 missense variant S766P is reported in gnomAD (ID 6‑33442454‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while SIFT uniquely predicts it as pathogenic. The remaining tools, ESM1b and AlphaMissense‑Default, are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.405110 | Structured | 0.923125 | Binding | 0.338 | 0.874 | 0.250 | 6-33442454-T-C | 1 | 1.28e-6 | -7.343 | In-Between | 0.374 | Ambiguous | Likely Benign | 0.193 | Likely Benign | -0.91 | Neutral | 0.006 | Benign | 0.013 | Benign | 4.07 | Benign | 0.01 | Affected | 3.64 | 6 | 0.2216 | 0.5862 | -1 | 1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||
| c.2302G>C | D768H 2D AIThe SynGAP1 D768H missense variant is not reported in ClinVar (ClinVar status: not reported) and is absent from gnomAD (gnomAD: not present). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the majority of predictions (5 benign vs. 4 pathogenic) and the high‑accuracy benign call suggest the variant is most likely benign, with no ClinVar status to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.332115 | Structured | 0.928237 | Binding | 0.314 | 0.877 | 0.250 | -8.673 | Likely Pathogenic | 0.783 | Likely Pathogenic | Likely Benign | 0.160 | Likely Benign | -1.85 | Neutral | 0.966 | Probably Damaging | 0.737 | Possibly Damaging | 4.03 | Benign | 0.12 | Tolerated | 0.1450 | 0.8136 | 1 | -1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||||||||||
| c.2303A>C | D768A 2D AIThe SynGAP1 D768A variant is listed in gnomAD (ID 6‑33442461‑A‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM) and pathogenic predictions (ESM1b, AlphaMissense‑Default). AlphaMissense‑Optimized returns an uncertain result. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑vs‑2 tie and thus unavailable; Foldetta stability analysis is not reported. Overall, the preponderance of evidence (six benign vs two pathogenic) points to a benign effect. This conclusion does not contradict ClinVar, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.332115 | Structured | 0.928237 | Binding | 0.314 | 0.877 | 0.250 | 6-33442461-A-C | -8.153 | Likely Pathogenic | 0.786 | Likely Pathogenic | Ambiguous | 0.174 | Likely Benign | -1.84 | Neutral | 0.245 | Benign | 0.096 | Benign | 4.09 | Benign | 0.14 | Tolerated | 3.64 | 6 | 0.3924 | 0.7662 | -2 | 0 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||
| c.2303A>G | D768G 2D AIThe SynGAP1 D768G missense variant is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors shows a predominance of benign calls: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized all predict benign, whereas AlphaMissense‑Default predicts pathogenic. The high‑accuracy AlphaMissense‑Optimized result is benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign classification (2 benign vs. 1 pathogenic, with one uncertain). Foldetta, which would provide a protein‑folding stability assessment, has no available output for this variant. Overall, the preponderance of evidence points to a benign effect, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.332115 | Structured | 0.928237 | Binding | 0.314 | 0.877 | 0.250 | -7.150 | In-Between | 0.697 | Likely Pathogenic | Likely Benign | 0.184 | Likely Benign | 0.03 | Neutral | 0.393 | Benign | 0.131 | Benign | 4.09 | Benign | 0.72 | Tolerated | 0.4130 | 0.7476 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2312C>A | S771Y 2D AIThe SynGAP1 missense variant S771Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools, ESM1b and AlphaMissense‑Default, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it contains two benign and two uncertain calls, and Foldetta data are unavailable. Overall, the balance of evidence favors a benign interpretation, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.436924 | Structured | 0.922503 | Binding | 0.306 | 0.883 | 0.250 | -7.041 | In-Between | 0.397 | Ambiguous | Likely Benign | 0.176 | Likely Benign | -2.13 | Neutral | 0.990 | Probably Damaging | 0.892 | Possibly Damaging | 4.02 | Benign | 0.05 | Affected | 0.0707 | 0.5155 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2312C>T | S771F 2D AIThe SynGAP1 missense variant S771F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is uncertain, and Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a benign interpretation, and this conclusion does not conflict with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.436924 | Structured | 0.922503 | Binding | 0.306 | 0.883 | 0.250 | -7.988 | In-Between | 0.525 | Ambiguous | Likely Benign | 0.189 | Likely Benign | -2.29 | Neutral | 0.990 | Probably Damaging | 0.892 | Possibly Damaging | 4.02 | Benign | 0.04 | Affected | 0.0624 | 0.5428 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2326G>T | G776C 2D AIThe SynGAP1 missense variant G776C is not reported in ClinVar but is present in gnomAD (ID 6‑33442484‑G‑T). Prediction tools cluster into benign (REVEL, FATHMM, AlphaMissense‑Optimized) and pathogenic (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT). Two tools (ESM1b, AlphaMissense‑Default) return uncertain results. High‑accuracy assessments are limited: AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta data are unavailable. Overall, the majority of conventional predictors indicate pathogenicity, whereas the single high‑accuracy tool suggests benign. Given the preponderance of pathogenic predictions and the absence of a ClinVar entry, the variant is most likely pathogenic and does not contradict any existing ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.377384 | Structured | 0.886983 | Binding | 0.296 | 0.888 | 0.250 | 6-33442484-G-T | -7.974 | In-Between | 0.380 | Ambiguous | Likely Benign | 0.181 | Likely Benign | -2.59 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 4.15 | Benign | 0.01 | Affected | 3.64 | 6 | 0.1235 | 0.4403 | -3 | -3 | 2.9 | 46.09 | |||||||||||||||||||||||||||||||||||||
| c.2330T>A | L777H 2D AIThe SynGAP1 missense variant L777H has no ClinVar record and is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence—including the consensus of high‑accuracy tools—suggests that the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.408655 | Structured | 0.876129 | Binding | 0.336 | 0.882 | 0.250 | -6.719 | Likely Benign | 0.492 | Ambiguous | Likely Benign | 0.230 | Likely Benign | -2.51 | Deleterious | 0.997 | Probably Damaging | 0.993 | Probably Damaging | 3.95 | Benign | 0.00 | Affected | 0.1052 | 0.0972 | -2 | -3 | -7.0 | 23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2335A>C | S779R 2D AIThe SynGAP1 missense variant S779R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and ESM1b, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. High‑accuracy methods give mixed results: AlphaMissense‑Optimized is uncertain, and both the SGM Consensus and Foldetta (which would combine FoldX‑MD and Rosetta outputs) are unavailable. Consequently, the overall prediction is ambiguous. The variant is most likely benign based on the balance of evidence, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.509769 | Disordered | 0.834974 | Binding | 0.321 | 0.890 | 0.375 | -4.044 | Likely Benign | 0.950 | Likely Pathogenic | Ambiguous | 0.103 | Likely Benign | -0.74 | Neutral | 0.846 | Possibly Damaging | 0.627 | Possibly Damaging | 2.29 | Pathogenic | 0.12 | Tolerated | 0.0961 | 0.4046 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2336G>A | S779N 2D  AIThe SynGAP1 missense variant S779N is listed in gnomAD (ID 6‑33442494‑G‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. A high‑accuracy consensus (SGM Consensus) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a benign majority. The AlphaMissense‑Optimized score is benign, and no Foldetta stability assessment is available. Taken together, the majority of evidence points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.509769 | Disordered | 0.834974 | Binding | 0.321 | 0.890 | 0.375 | 6-33442494-G-A | -4.880 | Likely Benign | 0.384 | Ambiguous | Likely Benign | 0.087 | Likely Benign | -0.75 | Neutral | 0.021 | Benign | 0.026 | Benign | 2.30 | Pathogenic | 0.23 | Tolerated | 3.64 | 6 | 0.1327 | 0.4984 | 1 | 1 | -2.7 | 27.03 | |||||||||||||||||||||||||||||||||||||
| c.2336G>T | S779I 2D AIThe SynGAP1 missense variant S779I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy methods give a benign call from AlphaMissense‑Optimized; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the majority of tools (five pathogenic vs. four benign) suggest a pathogenic impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.509769 | Disordered | 0.834974 | Binding | 0.321 | 0.890 | 0.375 | -6.261 | Likely Benign | 0.578 | Likely Pathogenic | Likely Benign | 0.198 | Likely Benign | -2.10 | Neutral | 0.918 | Possibly Damaging | 0.827 | Possibly Damaging | 2.28 | Pathogenic | 0.05 | Affected | 0.1046 | 0.6248 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2337C>A | S779R 2D AIThe SynGAP1 missense variant S779R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and ESM1b, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. High‑accuracy methods give mixed results: AlphaMissense‑Optimized is uncertain, and both the SGM Consensus and Foldetta (which would combine FoldX‑MD and Rosetta outputs) are unavailable. Consequently, the overall prediction is ambiguous. The variant is most likely benign based on the balance of evidence, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.509769 | Disordered | 0.834974 | Binding | 0.321 | 0.890 | 0.375 | -4.044 | Likely Benign | 0.950 | Likely Pathogenic | Ambiguous | 0.119 | Likely Benign | -0.74 | Neutral | 0.846 | Possibly Damaging | 0.627 | Possibly Damaging | 2.29 | Pathogenic | 0.12 | Tolerated | 0.0961 | 0.4046 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2337C>G | S779R 2D AIThe SynGAP1 missense variant S779R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and ESM1b, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. High‑accuracy methods give mixed results: AlphaMissense‑Optimized is uncertain, and both the SGM Consensus and Foldetta (which would combine FoldX‑MD and Rosetta outputs) are unavailable. Consequently, the overall prediction is ambiguous. The variant is most likely benign based on the balance of evidence, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.509769 | Disordered | 0.834974 | Binding | 0.321 | 0.890 | 0.375 | -4.044 | Likely Benign | 0.950 | Likely Pathogenic | Ambiguous | 0.124 | Likely Benign | -0.74 | Neutral | 0.846 | Possibly Damaging | 0.627 | Possibly Damaging | 2.29 | Pathogenic | 0.12 | Tolerated | 0.0961 | 0.4046 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2339C>A | S780Y 2D AIThe SynGAP1 missense variant S780Y is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.812415 | Binding | 0.283 | 0.883 | 0.500 | -7.682 | In-Between | 0.656 | Likely Pathogenic | Likely Benign | 0.091 | Likely Benign | -1.71 | Neutral | 0.995 | Probably Damaging | 0.925 | Probably Damaging | 2.61 | Benign | 0.11 | Tolerated | 0.0810 | 0.6428 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2339C>T | S780F 2D AIThe SynGAP1 missense variant S780F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic) and therefore unavailable as evidence. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is also unavailable for this variant. Overall, the majority of available predictions (five benign vs. four pathogenic) lean toward a benign impact. There is no ClinVar annotation to contradict this assessment, so the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.812415 | Binding | 0.283 | 0.883 | 0.500 | -8.055 | Likely Pathogenic | 0.677 | Likely Pathogenic | Likely Benign | 0.104 | Likely Benign | -1.42 | Neutral | 0.995 | Probably Damaging | 0.925 | Probably Damaging | 2.61 | Benign | 0.10 | Tolerated | 0.0768 | 0.6706 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2344G>A | D782N 2D AIThe SynGAP1 missense variant D782N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs four benign) lean toward a pathogenic impact. This conclusion does not contradict ClinVar status, as the variant has no existing ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.768342 | Binding | 0.285 | 0.883 | 0.625 | -6.857 | Likely Benign | 0.766 | Likely Pathogenic | Likely Benign | 0.131 | Likely Benign | -2.26 | Neutral | 0.995 | Probably Damaging | 0.950 | Probably Damaging | 1.98 | Pathogenic | 0.02 | Affected | 0.1167 | 0.6884 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2346C>A | D782E 2D AIThe SynGAP1 missense variant D782E is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority (2 benign vs. 1 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact; this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.768342 | Binding | 0.285 | 0.883 | 0.625 | -4.447 | Likely Benign | 0.486 | Ambiguous | Likely Benign | 0.127 | Likely Benign | -1.75 | Neutral | 0.561 | Possibly Damaging | 0.207 | Benign | 2.14 | Pathogenic | 0.01 | Affected | 0.1351 | 0.7036 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2346C>G | D782E 2D AIThe SynGAP1 missense variant D782E is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a majority benign vote (2 benign vs. 1 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact; this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.768342 | Binding | 0.285 | 0.883 | 0.625 | -4.447 | Likely Benign | 0.486 | Ambiguous | Likely Benign | 0.127 | Likely Benign | -1.75 | Neutral | 0.561 | Possibly Damaging | 0.207 | Benign | 2.14 | Pathogenic | 0.01 | Affected | 0.1351 | 0.7036 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2348T>A | M783K 2D AIThe SynGAP1 missense variant M783K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.736850 | Disordered | 0.738119 | Binding | 0.331 | 0.889 | 0.625 | -4.923 | Likely Benign | 0.693 | Likely Pathogenic | Likely Benign | 0.206 | Likely Benign | -2.70 | Deleterious | 0.925 | Possibly Damaging | 0.424 | Benign | 2.66 | Benign | 0.01 | Affected | 0.1730 | 0.0912 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||||||||||||
| c.2348T>G | M783R 2D AIThe SynGAP1 missense variant M783R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs four benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.736850 | Disordered | 0.738119 | Binding | 0.331 | 0.889 | 0.625 | -3.849 | Likely Benign | 0.655 | Likely Pathogenic | Likely Benign | 0.208 | Likely Benign | -2.69 | Deleterious | 0.925 | Possibly Damaging | 0.529 | Possibly Damaging | 2.66 | Benign | 0.01 | Affected | 0.1815 | 0.0893 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||||||||||||
| c.2354G>A | R785H 2D AIThe SynGAP1 R785H missense variant (ClinVar ID 2321588.0) is listed as “Uncertain” in ClinVar and is present in gnomAD (ID 6‑33442906‑G‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, whereas the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a pathogenic outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, does not provide a result for this variant. Overall, the majority of computational predictions (five pathogenic versus three benign) lean toward a pathogenic interpretation. Thus, the variant is most likely pathogenic based on current predictions, and this conclusion does not contradict the ClinVar status, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.859585 | Disordered | 0.681730 | Binding | 0.325 | 0.896 | 0.625 | Uncertain | 2 | 6-33442906-G-A | 4 | 2.50e-6 | -4.782 | Likely Benign | 0.388 | Ambiguous | Likely Benign | 0.129 | Likely Benign | -2.61 | Deleterious | 0.999 | Probably Damaging | 0.947 | Probably Damaging | 2.25 | Pathogenic | 0.01 | Affected | 3.64 | 6 | 0.3260 | 0.1589 | 2 | 0 | 1.3 | -19.05 | ||||||||||||||||||||||||||||||||
| c.2359C>G | P787A 2D AIThe SynGAP1 P787A missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas a majority (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM) predict a pathogenic impact. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta results are unavailable. Overall, the balance of evidence—five pathogenic versus three benign predictions, with the SGM Consensus supporting pathogenicity—suggests that the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.901269 | Disordered | 0.613211 | Binding | 0.377 | 0.899 | 0.750 | -4.542 | Likely Benign | 0.451 | Ambiguous | Likely Benign | 0.242 | Likely Benign | -4.23 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.48 | Pathogenic | 0.02 | Affected | 0.3425 | 0.4444 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.2359C>T | P787S 2D AIThe SynGAP1 P787S variant is listed in ClinVar with an “Uncertain” status and is present in gnomAD (variant ID 6‑33442911‑C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized, while those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. The high‑accuracy consensus (SGM Consensus) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a pathogenic majority. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic effect, and this assessment does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.901269 | Disordered | 0.613211 | Binding | 0.377 | 0.899 | 0.750 | Uncertain | 1 | 6-33442911-C-T | 3 | 1.86e-6 | -4.203 | Likely Benign | 0.564 | Ambiguous | Likely Benign | 0.221 | Likely Benign | -3.81 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.48 | Pathogenic | 0.02 | Affected | 3.64 | 6 | 0.3420 | 0.4675 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||
| c.2360C>G | P787R 2D AIThe SynGAP1 missense variant P787R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of conventional tools lean toward pathogenicity, whereas the single high‑accuracy tool indicates benign and the consensus is unresolved. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.901269 | Disordered | 0.613211 | Binding | 0.377 | 0.899 | 0.750 | -5.013 | Likely Benign | 0.784 | Likely Pathogenic | Likely Benign | 0.268 | Likely Benign | -4.41 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.50 | Benign | 0.03 | Affected | 0.1373 | 0.2741 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.2362T>C | S788P 2D AIThe SynGAP1 missense variant S788P is catalogued in gnomAD (ID 6‑33442914‑T‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The AlphaMissense‑Default score is uncertain. A consensus derived from the SGM framework (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) is unavailable for this residue. Taken together, the preponderance of evidence from standard predictors and the SGM consensus points to a likely pathogenic effect, which does not contradict any existing ClinVar annotation because none is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.956248 | Disordered | 0.573557 | Binding | 0.349 | 0.895 | 0.750 | 6-33442914-T-C | -1.857 | Likely Benign | 0.435 | Ambiguous | Likely Benign | 0.236 | Likely Benign | -3.91 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 1.51 | Pathogenic | 0.01 | Affected | 3.64 | 6 | 0.2338 | 0.5537 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||
| c.2363C>G | S788C 2D AIThe SynGAP1 missense variant S788C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and AlphaMissense‑Optimized, while five tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM) predict a pathogenic outcome. Two tools (ESM1b and AlphaMissense‑Default) return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—classifies the variant as pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar status because the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.956248 | Disordered | 0.573557 | Binding | 0.349 | 0.895 | 0.750 | -7.935 | In-Between | 0.472 | Ambiguous | Likely Benign | 0.269 | Likely Benign | -3.90 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.1393 | 0.6073 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.2365C>A | P789T 2D AIThe SynGAP1 P789T missense variant has no ClinVar entry and is not present in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further clarify the variant’s likely effect: AlphaMissense‑Optimized predicts a benign outcome, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status is unavailable. Overall, the preponderance of evidence from both general and high‑accuracy tools indicates that P789T is most likely pathogenic, and this assessment does not contradict any ClinVar status, as none is reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.963420 | Disordered | 0.541575 | Binding | 0.398 | 0.903 | 0.750 | -5.242 | Likely Benign | 0.356 | Ambiguous | Likely Benign | 0.224 | Likely Benign | -4.77 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.06 | Pathogenic | 0.00 | Affected | 0.1371 | 0.3837 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2365C>G | P789A 2D AIThe SynGAP1 P789A missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a tie (2 benign, 2 pathogenic) and is therefore inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus remains inconclusive and Foldetta data are unavailable. Overall, the majority of tools (five out of nine) predict pathogenicity, but the presence of four benign predictions and the inconclusive high‑accuracy results suggest uncertainty. The variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.963420 | Disordered | 0.541575 | Binding | 0.398 | 0.903 | 0.750 | -4.777 | Likely Benign | 0.235 | Likely Benign | Likely Benign | 0.258 | Likely Benign | -4.92 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.11 | Pathogenic | 0.00 | Affected | 0.3120 | 0.3052 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.2365C>T | P789S 2D AIThe SynGAP1 P789S variant has no ClinVar entry (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify it as benign include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence leans toward pathogenicity, with five tools supporting a deleterious effect versus three supporting benignity. This prediction does not contradict ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.963420 | Disordered | 0.541575 | Binding | 0.398 | 0.903 | 0.750 | -4.793 | Likely Benign | 0.409 | Ambiguous | Likely Benign | 0.221 | Likely Benign | -4.64 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.19 | Pathogenic | 0.00 | Affected | 0.3100 | 0.3436 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2366C>A | P789Q 2D AIThe SynGAP1 P789Q missense change is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.963420 | Disordered | 0.541575 | Binding | 0.398 | 0.903 | 0.750 | -5.191 | Likely Benign | 0.465 | Ambiguous | Likely Benign | 0.235 | Likely Benign | -4.53 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.03 | Pathogenic | 0.00 | Affected | 0.1284 | 0.3215 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||||
| c.2366C>T | P789L 2D AIThe SynGAP1 P789L missense variant has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized predicts benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence (five pathogenic vs. three benign predictions, with the SGM Consensus supporting pathogenicity) indicates that P789L is most likely pathogenic, and this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.963420 | Disordered | 0.541575 | Binding | 0.398 | 0.903 | 0.750 | -4.623 | Likely Benign | 0.457 | Ambiguous | Likely Benign | 0.303 | Likely Benign | -5.91 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 0.1958 | 0.4866 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.2368A>C | T790P 2D AIThe SynGAP1 missense variant T790P has no ClinVar entry (ClinVar status: not reported) but is present in gnomAD (ID 6‑33442920‑A‑C). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of conventional tools (5 pathogenic vs 4 benign) lean toward a pathogenic interpretation, while the single high‑accuracy tool suggests benign. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this does not contradict the ClinVar status, which currently has no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.964893 | Disordered | 0.509280 | Binding | 0.385 | 0.896 | 0.875 | 6-33442920-A-C | -3.564 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.250 | Likely Benign | -3.55 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.25 | Pathogenic | 0.01 | Affected | 3.64 | 6 | 0.2147 | 0.4748 | -1 | 0 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||
| c.2368A>G | T790A 2D AIThe SynGAP1 T790A missense change is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized labeling the variant as benign, while the SGM Consensus remains inconclusive and Foldetta data are unavailable. Overall, the majority of standard predictors indicate pathogenicity, but the single high‑accuracy tool that is available suggests a benign effect, and no high‑accuracy tool provides a definitive pathogenic verdict. Consequently, the variant is most likely pathogenic according to the bulk of predictions, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.964893 | Disordered | 0.509280 | Binding | 0.385 | 0.896 | 0.875 | -4.337 | Likely Benign | 0.096 | Likely Benign | Likely Benign | 0.167 | Likely Benign | -2.64 | Deleterious | 0.992 | Probably Damaging | 0.989 | Probably Damaging | 2.35 | Pathogenic | 0.02 | Affected | 0.4157 | 0.4207 | 1 | 0 | 2.5 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2369C>A | T790N 2D  AIThe SynGAP1 missense variant T790N is listed in ClinVar with an “Uncertain” status and is present in the gnomAD database (ID 6‑33442921‑C‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive and therefore unavailable, and Foldetta results are not reported. Overall, the majority of conventional tools (5 pathogenic vs. 4 benign) lean toward a pathogenic interpretation, while the single high‑accuracy tool suggests benign. The variant’s ClinVar status remains uncertain, so there is no contradiction with the current clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.964893 | Disordered | 0.509280 | Binding | 0.385 | 0.896 | 0.875 | Conflicting | 3 | 6-33442921-C-A | 69 | 4.28e-5 | -5.243 | Likely Benign | 0.276 | Likely Benign | Likely Benign | 0.103 | Likely Benign | -2.54 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.27 | Pathogenic | 0.02 | Affected | 3.64 | 6 | 0.1446 | 0.4653 | 0 | 0 | -2.8 | 13.00 | ||||||||||||||||||||||||||||||||
| c.2369C>T | T790I 2D AIThe SynGAP1 missense variant T790I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic. Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact. This conclusion does not contradict the ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.964893 | Disordered | 0.509280 | Binding | 0.385 | 0.896 | 0.875 | -3.556 | Likely Benign | 0.482 | Ambiguous | Likely Benign | 0.190 | Likely Benign | -3.08 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.28 | Pathogenic | 0.01 | Affected | 0.0987 | 0.5431 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||||||||||||
| c.2375A>C | E792A 2D AIThe SynGAP1 missense variant E792A is catalogued in gnomAD (ID 6‑33442927‑A‑C) but has no ClinVar submission. Functional prediction tools cluster into two groups: benign (REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized) and pathogenic (PROVEAN, SIFT). The high‑accuracy AlphaMissense‑Optimized score is benign, and the SGM consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors benign. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Overall, the preponderance of evidence points to a benign effect. This conclusion is consistent with the absence of a ClinVar pathogenic classification; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.974374 | Disordered | 0.452261 | Uncertain | 0.352 | 0.896 | 0.875 | 6-33442927-A-C | 1 | 6.20e-7 | -3.248 | Likely Benign | 0.515 | Ambiguous | Likely Benign | 0.060 | Likely Benign | -3.35 | Deleterious | 0.000 | Benign | 0.001 | Benign | 3.92 | Benign | 0.01 | Affected | 3.64 | 6 | 0.4586 | 0.7544 | -1 | 0 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||
| c.2375A>G | E792G 2D AIThe SynGAP1 E792G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, favors a benign outcome (2 benign vs. 1 pathogenic vote). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as benign, and Foldetta results are unavailable. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.974374 | Disordered | 0.452261 | Uncertain | 0.352 | 0.896 | 0.875 | -3.925 | Likely Benign | 0.353 | Ambiguous | Likely Benign | 0.037 | Likely Benign | -3.77 | Deleterious | 0.000 | Benign | 0.000 | Benign | 3.88 | Benign | 0.01 | Affected | 0.3145 | 0.6036 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||||||
| c.2375A>T | E792V 2D AIThe SynGAP1 E792V missense change is not listed in ClinVar and has no allele record in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—yields a tie and is therefore inconclusive. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence leans toward a benign effect. This conclusion is consistent with the absence of a ClinVar assertion, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.974374 | Disordered | 0.452261 | Uncertain | 0.352 | 0.896 | 0.875 | -4.643 | Likely Benign | 0.640 | Likely Pathogenic | Likely Benign | 0.072 | Likely Benign | -3.85 | Deleterious | 0.000 | Benign | 0.001 | Benign | 3.83 | Benign | 0.00 | Affected | 0.0987 | 0.7772 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.239A>T | K80I 2D  AIThe SynGAP1 missense variant K80I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta data are unavailable. Consequently, the variant’s predicted impact is ambiguous, with an equal split between benign and pathogenic signals and no ClinVar entry to contradict the computational assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.637480 | Disordered | 0.477530 | Uncertain | 0.331 | 0.873 | 0.500 | -5.320 | Likely Benign | 0.947 | Likely Pathogenic | Ambiguous | 0.083 | Likely Benign | -2.54 | Deleterious | 0.939 | Possibly Damaging | 0.164 | Benign | 3.86 | Benign | 0.00 | Affected | 0.1095 | 0.2981 | -2 | -3 | 8.4 | -15.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2407A>G | K803E 2D AIThe SynGAP1 missense variant K803E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs 2 benign) and Foldetta results are unavailable. Overall, the majority of tools (five pathogenic vs. four benign) predict a pathogenic impact. Thus, the variant is most likely pathogenic, and this prediction does not contradict any ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.827927 | Disordered | 0.733908 | Binding | 0.349 | 0.900 | 0.625 | -3.889 | Likely Benign | 0.650 | Likely Pathogenic | Likely Benign | 0.172 | Likely Benign | -2.06 | Neutral | 0.896 | Possibly Damaging | 0.596 | Possibly Damaging | 2.38 | Pathogenic | 0.00 | Affected | 0.4043 | 0.1357 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.2409A>C | K803N 2D AIThe SynGAP1 K803N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool rates the variant as uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, more tools predict pathogenicity (five) than benignity (three), and the high‑accuracy assessments do not overturn this trend. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.827927 | Disordered | 0.733908 | Binding | 0.349 | 0.900 | 0.625 | -5.039 | Likely Benign | 0.807 | Likely Pathogenic | Ambiguous | 0.045 | Likely Benign | -2.13 | Neutral | 0.896 | Possibly Damaging | 0.673 | Possibly Damaging | 2.34 | Pathogenic | 0.00 | Affected | 0.3805 | 0.2035 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.2409A>T | K803N 2D AIThe SynGAP1 K803N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, more tools (five) predict pathogenicity than benign (three), and the high‑accuracy assessments do not overturn this trend. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.827927 | Disordered | 0.733908 | Binding | 0.349 | 0.900 | 0.625 | -5.039 | Likely Benign | 0.807 | Likely Pathogenic | Ambiguous | 0.045 | Likely Benign | -2.13 | Neutral | 0.896 | Possibly Damaging | 0.673 | Possibly Damaging | 2.34 | Pathogenic | 0.00 | Affected | 0.3805 | 0.2035 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.2410G>A | D804N 2D AIThe SynGAP1 D804N variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of conventional tools predict a benign impact, and this assessment does not contradict the lack of ClinVar annotation. Thus, based on the current predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.801317 | Disordered | 0.786762 | Binding | 0.294 | 0.900 | 0.625 | -4.681 | Likely Benign | 0.522 | Ambiguous | Likely Benign | 0.104 | Likely Benign | -2.62 | Deleterious | 0.400 | Benign | 0.212 | Benign | 1.22 | Pathogenic | 0.06 | Tolerated | 0.1528 | 0.7393 | 2 | 1 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.2414T>A | L805Q 2D AIThe SynGAP1 missense variant L805Q is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts a benign effect, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (2 pathogenic vs. 1 benign, 1 uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence (five pathogenic vs. three benign predictions, with a pathogenic SGM Consensus) indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.775545 | Disordered | 0.827669 | Binding | 0.341 | 0.903 | 0.625 | -6.244 | Likely Benign | 0.427 | Ambiguous | Likely Benign | 0.152 | Likely Benign | -2.66 | Deleterious | 0.927 | Possibly Damaging | 0.690 | Possibly Damaging | 2.37 | Pathogenic | 0.00 | Affected | 0.1215 | 0.1265 | -2 | -2 | -7.3 | 14.97 | |||||||||||||||||||||||||||||||||||||||
| c.2414T>C | L805P 2D AIThe SynGAP1 missense variant L805P is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments give a benign result from AlphaMissense‑Optimized, a pathogenic outcome from the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and no Foldetta data are available. Overall, the majority of predictions lean toward pathogenicity, and this conclusion does not conflict with the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.775545 | Disordered | 0.827669 | Binding | 0.341 | 0.903 | 0.625 | Uncertain | 1 | -4.661 | Likely Benign | 0.444 | Ambiguous | Likely Benign | 0.272 | Likely Benign | -3.40 | Deleterious | 0.975 | Probably Damaging | 0.767 | Possibly Damaging | 2.36 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3128 | 0.1650 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||
| c.2417T>A | F806Y 2D AIThe SynGAP1 missense variant F806Y is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM all predict a pathogenic outcome; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign, the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also classifies it as benign, and Foldetta results are unavailable. Taken together, the majority of evidence points to a benign effect, and this conclusion does not contradict the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.736850 | Disordered | 0.847454 | Binding | 0.276 | 0.904 | 0.500 | -5.229 | Likely Benign | 0.404 | Ambiguous | Likely Benign | 0.162 | Likely Benign | -1.51 | Neutral | 0.997 | Probably Damaging | 0.987 | Probably Damaging | 2.32 | Pathogenic | 0.03 | Affected | 0.1746 | 0.1437 | 7 | 3 | -4.1 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2419T>C | Y807H 2D AIThe SynGAP1 missense variant Y807H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that indicate a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as pathogenic. AlphaMissense‑Optimized remains benign, while Foldetta results are unavailable. Overall, the majority of high‑accuracy and consensus predictors (five out of six) suggest a pathogenic impact, whereas only three predict benign. Consequently, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.699094 | Disordered | 0.853760 | Binding | 0.336 | 0.901 | 0.500 | -5.879 | Likely Benign | 0.413 | Ambiguous | Likely Benign | 0.152 | Likely Benign | -2.74 | Deleterious | 0.989 | Probably Damaging | 0.913 | Probably Damaging | 2.44 | Pathogenic | 0.00 | Affected | 0.2530 | 0.0704 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||||||||||||
| c.2420A>C | Y807S 2D AIThe SynGAP1 Y807S variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on benign impact include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b, while those that predict pathogenicity are PROVEAN, SIFT, and FATHMM. AlphaMissense‑Default is uncertain, whereas AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, leans toward pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of conventional tools (four benign vs. three pathogenic) suggest a benign effect, and the high‑accuracy AlphaMissense‑Optimized also predicts benign, while the SGM Consensus predicts pathogenic. Based on the balance of evidence, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar reporting. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.699094 | Disordered | 0.853760 | Binding | 0.336 | 0.901 | 0.500 | -5.517 | Likely Benign | 0.384 | Ambiguous | Likely Benign | 0.093 | Likely Benign | -3.90 | Deleterious | 0.136 | Benign | 0.067 | Benign | 2.44 | Pathogenic | 0.01 | Affected | 0.4561 | 0.1884 | -3 | -2 | 0.5 | -76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2420A>G | Y807C 2D AIThe SynGAP1 missense variant Y807C is listed in ClinVar with an “Uncertain” status (ClinVar ID 2119812.0) and is present in gnomAD (ID 6‑33442972‑A‑G). Prediction tools that agree on a benign effect include REVEL, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. three benign) and the SGM Consensus support a pathogenic interpretation, whereas AlphaMissense‑Optimized alone suggests benign. The variant is most likely pathogenic based on the collective evidence, and this conclusion is not contradicted by the ClinVar “Uncertain” status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.699094 | Disordered | 0.853760 | Binding | 0.336 | 0.901 | 0.500 | Uncertain | 1 | 6-33442972-A-G | 1 | 6.20e-7 | -7.228 | In-Between | 0.204 | Likely Benign | Likely Benign | 0.243 | Likely Benign | -3.89 | Deleterious | 0.997 | Probably Damaging | 0.934 | Probably Damaging | 2.42 | Pathogenic | 0.01 | Affected | 3.77 | 5 | 0.3101 | 0.1907 | 0 | -2 | 3.8 | -60.04 | ||||||||||||||||||||||||||||||||
| c.2423T>C | V808A 2D AIThe SynGAP1 missense variant V808A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign verdict, while Foldetta results are unavailable. Taken together, the majority of evidence points to a benign impact, and this conclusion does not contradict the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.699094 | Disordered | 0.856438 | Binding | 0.289 | 0.903 | 0.500 | -5.091 | Likely Benign | 0.541 | Ambiguous | Likely Benign | 0.177 | Likely Benign | -1.96 | Neutral | 0.953 | Possibly Damaging | 0.621 | Possibly Damaging | 2.31 | Pathogenic | 0.00 | Affected | 0.2748 | 0.2809 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.2423T>G | V808G 2D AIThe SynGAP1 V808G missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, the majority of individual predictors (five benign vs. three pathogenic) support a benign classification, and this is not contradicted by any ClinVar annotation. Thus, based on the available predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.699094 | Disordered | 0.856438 | Binding | 0.289 | 0.903 | 0.500 | -6.635 | Likely Benign | 0.460 | Ambiguous | Likely Benign | 0.268 | Likely Benign | -2.94 | Deleterious | 0.069 | Benign | 0.135 | Benign | 2.27 | Pathogenic | 0.00 | Affected | 0.1925 | 0.3336 | -1 | -3 | -4.6 | -42.08 | |||||||||||||||||||||||||||||||||||||||
| c.2425A>C | S809R 2D AIThe SynGAP1 missense variant S809R is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools cluster into three groups: benign predictions from REVEL, PROVEAN, and FATHMM; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default; and uncertain predictions from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward benign. Protein‑folding stability analysis with Foldetta is unavailable. Overall, the balance of evidence, particularly the benign consensus from high‑accuracy tools, suggests the variant is most likely benign, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.626927 | Disordered | 0.853218 | Binding | 0.330 | 0.907 | 0.500 | -7.062 | In-Between | 0.935 | Likely Pathogenic | Ambiguous | 0.085 | Likely Benign | -1.50 | Neutral | 0.784 | Possibly Damaging | 0.472 | Possibly Damaging | 2.51 | Benign | 0.01 | Affected | 0.0954 | 0.3702 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.2425A>T | S809C 2D AIThe SynGAP1 missense variant S809C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The SGM Consensus, which is a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools specifically show AlphaMissense‑Optimized as benign, while SGM Consensus and Foldetta are unavailable. Overall, the majority of predictions (5 pathogenic vs. 4 benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.626927 | Disordered | 0.853218 | Binding | 0.330 | 0.907 | 0.500 | -8.186 | Likely Pathogenic | 0.221 | Likely Benign | Likely Benign | 0.145 | Likely Benign | -1.99 | Neutral | 0.975 | Probably Damaging | 0.766 | Possibly Damaging | 2.49 | Pathogenic | 0.01 | Affected | 0.1088 | 0.6174 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.2426G>T | S809I 2D AIThe SynGAP1 missense variant S809I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Consensus from standard in silico predictors shows a split: six tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized) predict a benign effect, while two (SIFT, AlphaMissense‑Default) predict pathogenicity; ESM1b is uncertain. High‑accuracy assessment further supports a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign classification, and Foldetta data are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.626927 | Disordered | 0.853218 | Binding | 0.330 | 0.907 | 0.500 | -7.708 | In-Between | 0.632 | Likely Pathogenic | Likely Benign | 0.087 | Likely Benign | -1.93 | Neutral | 0.065 | Benign | 0.022 | Benign | 2.50 | Benign | 0.01 | Affected | 0.1039 | 0.5984 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.2427C>A | S809R 2D AIThe SynGAP1 missense variant S809R is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools cluster into three groups: benign predictions from REVEL, PROVEAN, and FATHMM; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default; and uncertain predictions from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward benign. Protein‑folding stability analysis with Foldetta is unavailable. Overall, the balance of evidence, particularly the benign consensus from high‑accuracy tools, suggests the variant is most likely benign, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.626927 | Disordered | 0.853218 | Binding | 0.330 | 0.907 | 0.500 | -7.062 | In-Between | 0.935 | Likely Pathogenic | Ambiguous | 0.091 | Likely Benign | -1.50 | Neutral | 0.784 | Possibly Damaging | 0.472 | Possibly Damaging | 2.51 | Benign | 0.01 | Affected | 0.0954 | 0.3702 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.2427C>G | S809R 2D AIThe SynGAP1 missense variant S809R is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools cluster into three groups: benign predictions from REVEL, PROVEAN, and FATHMM; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default; and uncertain predictions from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward benign. Protein‑folding stability analysis with Foldetta is unavailable. Overall, the balance of evidence, particularly the benign consensus from high‑accuracy tools, suggests the variant is most likely benign, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.626927 | Disordered | 0.853218 | Binding | 0.330 | 0.907 | 0.500 | -7.062 | In-Between | 0.935 | Likely Pathogenic | Ambiguous | 0.091 | Likely Benign | -1.50 | Neutral | 0.784 | Possibly Damaging | 0.472 | Possibly Damaging | 2.51 | Benign | 0.01 | Affected | 0.0954 | 0.3702 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.2429G>A | R810H 2D AIThe SynGAP1 R810H missense variant is listed in gnomAD (ID 6‑33442981‑G‑A) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas a pathogenic consensus is reached by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as pathogenic, and Foldetta (FoldX‑MD/Rosetta stability analysis) is unavailable. Overall, the majority of tools predict pathogenicity, and the high‑accuracy subset is split, but the pathogenic signal dominates. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.486429 | Structured | 0.851848 | Binding | 0.263 | 0.907 | 0.375 | 6-33442981-G-A | 3 | 1.86e-6 | -6.554 | Likely Benign | 0.550 | Ambiguous | Likely Benign | 0.239 | Likely Benign | -2.80 | Deleterious | 1.000 | Probably Damaging | 0.980 | Probably Damaging | 2.35 | Pathogenic | 0.01 | Affected | 3.77 | 5 | 0.3149 | 0.2631 | 0 | 2 | 1.3 | -19.05 | ||||||||||||||||||||||||||||||||||
| c.2432C>G | P811R 2D AIThe SynGAP1 missense variant P811R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT; AlphaMissense‑Default remains uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, favors a benign outcome (2 benign vs. 1 pathogenic, with one uncertain). High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM Consensus as benign, while Foldetta (combining FoldX‑MD and Rosetta) has no available result. Overall, the preponderance of evidence points to a benign impact for P811R, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.411940 | Structured | 0.847064 | Binding | 0.382 | 0.910 | 0.250 | -5.273 | Likely Benign | 0.557 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -2.69 | Deleterious | 0.100 | Benign | 0.066 | Benign | 2.73 | Benign | 0.01 | Affected | 0.1448 | 0.3817 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.2432C>T | P811L 2D AIThe SynGAP1 P811L missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. AlphaMissense‑Default is uncertain. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, leans toward benign (2 benign vs. 1 pathogenic, 1 uncertain). AlphaMissense‑Optimized also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from high‑accuracy tools and consensus methods indicates that P811L is most likely benign. This assessment does not contradict ClinVar status, as the variant is not yet reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.411940 | Structured | 0.847064 | Binding | 0.382 | 0.910 | 0.250 | -6.184 | Likely Benign | 0.491 | Ambiguous | Likely Benign | 0.150 | Likely Benign | -3.98 | Deleterious | 0.982 | Probably Damaging | 0.824 | Possibly Damaging | 2.71 | Benign | 0.01 | Affected | 0.2327 | 0.6621 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.2435C>A | P812H 2D AIThe SynGAP1 missense variant P812H is listed in ClinVar with an uncertain significance and is present in the gnomAD database (ID 6‑33442987‑C‑A). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic outcome; ESM1b remains uncertain. High‑accuracy methods give a benign result from AlphaMissense‑Optimized, a pathogenic consensus from the SGM approach (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta data are unavailable. Overall, the balance of evidence points to a pathogenic effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.414856 | Structured | 0.842442 | Binding | 0.388 | 0.901 | 0.125 | Uncertain | 2 | 6-33442987-C-A | 3 | 1.86e-6 | -7.470 | In-Between | 0.698 | Likely Pathogenic | Likely Benign | 0.272 | Likely Benign | -2.81 | Deleterious | 1.000 | Probably Damaging | 0.995 | Probably Damaging | 2.68 | Benign | 0.00 | Affected | 4.32 | 4 | 0.1568 | 0.4923 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||
| c.2435C>G | P812R 2D AIThe SynGAP1 P812R missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of standard predictors indicate a pathogenic impact, while the most accurate tools provide no definitive evidence. Thus, the variant is most likely pathogenic, and this assessment does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.414856 | Structured | 0.842442 | Binding | 0.388 | 0.901 | 0.125 | -6.784 | Likely Benign | 0.799 | Likely Pathogenic | Ambiguous | 0.222 | Likely Benign | -2.70 | Deleterious | 0.999 | Probably Damaging | 0.985 | Probably Damaging | 2.77 | Benign | 0.01 | Affected | 0.1285 | 0.3525 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.2435C>T | P812L 2D AIThe SynGAP1 P812L missense variant is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (gnomAD ID: 6‑33442987‑C‑T). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic, and Foldetta’s protein‑folding stability analysis is unavailable. Overall, the majority of predictions lean toward pathogenicity, which is consistent with the lack of ClinVar reporting but does not contradict it. Thus, based on the available computational evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.414856 | Structured | 0.842442 | Binding | 0.388 | 0.901 | 0.125 | 6-33442987-C-T | 1 | 6.20e-7 | -7.121 | In-Between | 0.591 | Likely Pathogenic | Likely Benign | 0.172 | Likely Benign | -2.61 | Deleterious | 0.978 | Probably Damaging | 0.824 | Possibly Damaging | 2.76 | Benign | 0.01 | Affected | 4.32 | 4 | 0.2131 | 0.6634 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||
| c.2441C>A | A814D 2D AIThe SynGAP1 missense variant A814D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a tie and thus unavailable, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no reported result. Overall, the balance of evidence from the consensus of standard predictors points to a pathogenic effect. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.411940 | Structured | 0.814830 | Binding | 0.368 | 0.902 | 0.250 | -5.641 | Likely Benign | 0.939 | Likely Pathogenic | Ambiguous | 0.158 | Likely Benign | -2.56 | Deleterious | 0.968 | Probably Damaging | 0.810 | Possibly Damaging | 2.59 | Benign | 0.01 | Affected | 0.1740 | 0.2167 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.2443C>A | R815S 2D AISynGAP1 R815S is listed in ClinVar as Benign (ID 3645150.0) and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. Two tools report uncertainty: ESM1b and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM Consensus as Benign, and Foldetta (combining FoldX‑MD and Rosetta) has no available result. Overall, the majority of predictions lean toward pathogenicity, whereas the consensus and high‑accuracy tools suggest benignity. Thus, the variant is most likely pathogenic based on the prevailing predictions, contradicting its ClinVar benign designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.394753 | Structured | 0.780568 | Binding | 0.278 | 0.907 | 0.250 | Benign | 1 | -7.324 | In-Between | 0.950 | Likely Pathogenic | Ambiguous | 0.138 | Likely Benign | -1.86 | Neutral | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.67 | Benign | 0.02 | Affected | 0.2937 | 0.4164 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||
| c.2443C>G | R815G 2D AISynGAP1 missense variant R815G is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that agree on benign effect include REVEL and FATHMM, whereas pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. Uncertain calls are made by ESM1b and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.394753 | Structured | 0.780568 | Binding | 0.278 | 0.907 | 0.250 | Uncertain | 1 | -7.983 | In-Between | 0.854 | Likely Pathogenic | Ambiguous | 0.146 | Likely Benign | -3.22 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.62 | Benign | 0.02 | Affected | 4.32 | 4 | 0.3481 | 0.4015 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||
| c.2444G>A | R815H 2D AIThe SynGAP1 missense variant R815H (ClinVar ID 833773.0) is classified as benign in ClinVar and is present in gnomAD (6‑33442996‑G‑A). Functional prediction tools cluster into two groups: benign predictions from REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar) and SIFT. Two tools report uncertainty: ESM1b and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and Foldetta stability assessment are unavailable. Overall, the balance of evidence favors a benign effect, consistent with the ClinVar annotation and with no conflict regarding its status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.394753 | Structured | 0.780568 | Binding | 0.278 | 0.907 | 0.250 | Likely Benign | 2 | 6-33442996-G-A | 24 | 1.49e-5 | -7.474 | In-Between | 0.553 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -1.81 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.61 | Benign | 0.02 | Affected | 4.32 | 4 | 0.2944 | 0.2281 | 2 | 0 | 1.3 | -19.05 | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||||||||||||||||
| c.2444G>C | R815P 2D AIThe SynGAP1 missense variant R815P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default) predict a pathogenic impact. The remaining tools, ESM1b and AlphaMissense‑Optimized, return uncertain results. High‑accuracy assessments further support a deleterious interpretation: the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as pathogenic; AlphaMissense‑Optimized remains uncertain, and Foldetta data are unavailable. Overall, the preponderance of evidence indicates that R815P is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.394753 | Structured | 0.780568 | Binding | 0.278 | 0.907 | 0.250 | -7.495 | In-Between | 0.858 | Likely Pathogenic | Ambiguous | 0.153 | Likely Benign | -2.85 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.65 | Benign | 0.02 | Affected | 0.2113 | 0.4897 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.2447C>G | S816C 2D AIThe SynGAP1 missense variant S816C has no ClinVar record and is not present in gnomAD. Functional prediction tools fall into two groups: benign predictions come from REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar) and AlphaMissense‑Default; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the evidence is mixed, with an equal split between benign and pathogenic calls among general predictors and conflicting outcomes from the two high‑accuracy tools. The variant is most likely pathogenic based on the predominance of pathogenic predictions, and this assessment does not contradict ClinVar status, which currently has no entry for S816C. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.476583 | Structured | 0.747189 | Binding | 0.347 | 0.898 | 0.375 | -7.998 | In-Between | 0.605 | Likely Pathogenic | Likely Benign | 0.246 | Likely Benign | -2.75 | Deleterious | 1.000 | Probably Damaging | 0.992 | Probably Damaging | 2.59 | Benign | 0.06 | Tolerated | 0.1167 | 0.5472 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2449T>A | S817T 2D AIThe SynGAP1 missense variant S817T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also leans benign, and Foldetta results are unavailable. Overall, the balance of evidence from both general and high‑accuracy predictors points to a benign impact for S817T. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.490133 | Structured | 0.727082 | Binding | 0.314 | 0.901 | 0.625 | -6.544 | Likely Benign | 0.492 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -1.95 | Neutral | 0.990 | Probably Damaging | 0.846 | Possibly Damaging | 2.43 | Pathogenic | 0.00 | Affected | 0.1622 | 0.6482 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2449T>G | S817A 2D AIThe SynGAP1 missense variant S817A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also resolves to benign, while Foldetta results are unavailable. Overall, the majority of high‑confidence predictions indicate a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.490133 | Structured | 0.727082 | Binding | 0.314 | 0.901 | 0.625 | -6.321 | Likely Benign | 0.429 | Ambiguous | Likely Benign | 0.116 | Likely Benign | -1.86 | Neutral | 0.977 | Probably Damaging | 0.846 | Possibly Damaging | 2.45 | Pathogenic | 0.00 | Affected | 0.5207 | 0.5866 | Strenghten | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2455G>A | A819T 2D AIThe SynGAP1 missense variant A819T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs 2 benign) and Foldetta data are unavailable. Overall, the majority of conventional predictors lean toward pathogenicity, but the single high‑accuracy tool predicts benign and the consensus is unresolved. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.707644 | Binding | 0.317 | 0.892 | 0.625 | -3.421 | Likely Benign | 0.675 | Likely Pathogenic | Likely Benign | 0.287 | Likely Benign | -2.23 | Neutral | 0.998 | Probably Damaging | 0.963 | Probably Damaging | 2.29 | Pathogenic | 0.01 | Affected | 0.1443 | 0.7583 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2456C>G | A819G 2D AIThe SynGAP1 missense variant A819G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the majority of predictions (six benign vs three pathogenic) support a benign classification. Thus, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.707644 | Binding | 0.317 | 0.892 | 0.625 | -5.730 | Likely Benign | 0.714 | Likely Pathogenic | Likely Benign | 0.116 | Likely Benign | -2.17 | Neutral | 0.104 | Benign | 0.121 | Benign | 2.29 | Pathogenic | 0.03 | Affected | 0.2363 | 0.5396 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2456C>T | A819V 2D AIThe SynGAP1 missense variant A819V is catalogued in gnomAD (ID 6‑33443008‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas a majority of tools predict a pathogenic outcome: polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic), and Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a pathogenic classification for A819V, and this assessment does not contradict any ClinVar status because the variant is not yet reported in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.707644 | Binding | 0.317 | 0.892 | 0.625 | 6-33443008-C-T | 1 | 6.19e-7 | -4.944 | Likely Benign | 0.797 | Likely Pathogenic | Ambiguous | 0.246 | Likely Benign | -2.40 | Neutral | 0.998 | Probably Damaging | 0.963 | Probably Damaging | 2.19 | Pathogenic | 0.01 | Affected | 3.77 | 5 | 0.1098 | 0.6664 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||||||||
| c.2458T>A | Y820N 2D AIThe SynGAP1 Y820N variant is listed in ClinVar with an “Uncertain” significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default all predict a pathogenic outcome. AlphaMissense‑Optimized returns an “Uncertain” result. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the predictions are evenly split between benign and pathogenic, with no high‑confidence pathogenic or benign signal. Thus, the variant is most likely of uncertain significance, which is consistent with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.733139 | Disordered | 0.695550 | Binding | 0.293 | 0.883 | 0.625 | Uncertain | 1 | -9.032 | Likely Pathogenic | 0.842 | Likely Pathogenic | Ambiguous | 0.143 | Likely Benign | -1.53 | Neutral | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.74 | Benign | 0.20 | Tolerated | 0.2352 | 0.0704 | -2 | -2 | -2.2 | -49.07 | ||||||||||||||||||||||||||||||||||||||
| c.2458T>C | Y820H 2D AIThe SynGAP1 missense variant Y820H is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443010‑T‑C). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Uncertain predictions come from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.733139 | Disordered | 0.695550 | Binding | 0.293 | 0.883 | 0.625 | 6-33443010-T-C | 5 | 3.10e-6 | -7.432 | In-Between | 0.928 | Likely Pathogenic | Ambiguous | 0.129 | Likely Benign | -0.13 | Neutral | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 2.70 | Benign | 0.24 | Tolerated | 3.77 | 5 | 0.2498 | 0.0704 | 2 | 0 | -1.9 | -26.03 | |||||||||||||||||||||||||||||||||||
| c.2459A>C | Y820S 2D AIThe SynGAP1 missense variant Y820S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign (2 benign vs. 1 pathogenic). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for Y820S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.733139 | Disordered | 0.695550 | Binding | 0.293 | 0.883 | 0.625 | -7.094 | In-Between | 0.723 | Likely Pathogenic | Likely Benign | 0.101 | Likely Benign | -1.79 | Neutral | 0.999 | Probably Damaging | 0.951 | Probably Damaging | 2.80 | Benign | 0.22 | Tolerated | 0.5166 | 0.2291 | Weaken | -3 | -2 | 0.5 | -76.10 | |||||||||||||||||||||||||||||||||||||||
| c.245T>A | L82Q 2D AIThe SynGAP1 missense variant L82Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, whereas the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign prediction. Foldetta results are unavailable. Overall, the majority of conventional tools lean toward benign, and the SGM Consensus supports this, but the AlphaMissense‑Optimized prediction introduces a pathogenic signal. Consequently, the variant is most likely benign based on the prevailing evidence, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.637480 | Disordered | 0.517720 | Binding | 0.284 | 0.890 | 0.375 | -7.576 | In-Between | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.079 | Likely Benign | -2.16 | Neutral | 0.939 | Possibly Damaging | 0.114 | Benign | 3.71 | Benign | 0.00 | Affected | 0.1150 | 0.0790 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||||
| c.245T>C | L82P 2D AIThe SynGAP1 missense variant L82P is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33425853‑T‑C). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors pathogenicity. Foldetta, a protein‑folding stability method, did not provide a result for this variant. Overall, the majority of evidence points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.637480 | Disordered | 0.517720 | Binding | 0.284 | 0.890 | 0.375 | 6-33425853-T-C | 1 | 6.20e-7 | -7.667 | In-Between | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.125 | Likely Benign | -2.73 | Deleterious | 0.939 | Possibly Damaging | 0.162 | Benign | 3.64 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3477 | 0.1118 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||
| c.2461T>C | C821R 2D AIThe SynGAP1 missense variant C821R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy predictors (six out of nine) indicate a pathogenic impact, while three suggest benign. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.745909 | Disordered | 0.672821 | Binding | 0.351 | 0.883 | 0.750 | -3.997 | Likely Benign | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.183 | Likely Benign | -2.93 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.69 | Benign | 0.03 | Affected | 0.1944 | 0.1746 | -4 | -3 | -7.0 | 53.05 | ||||||||||||||||||||||||||||||||||||||||
| c.2461T>G | C821G 2D AIThe SynGAP1 missense variant C821G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.745909 | Disordered | 0.672821 | Binding | 0.351 | 0.883 | 0.750 | -4.613 | Likely Benign | 0.910 | Likely Pathogenic | Ambiguous | 0.187 | Likely Benign | -2.94 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.68 | Benign | 0.05 | Affected | 0.3287 | 0.2747 | -3 | -3 | -2.9 | -46.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2462G>A | C821Y 2D AIThe SynGAP1 missense variant C821Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, seven tools support pathogenicity while three support benignity, and no high‑accuracy consensus contradicts this trend. Therefore, the variant is most likely pathogenic based on the available predictions, and this assessment does not conflict with the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.745909 | Disordered | 0.672821 | Binding | 0.351 | 0.883 | 0.750 | -1.007 | Likely Benign | 0.982 | Likely Pathogenic | Likely Pathogenic | 0.342 | Likely Benign | -3.11 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.66 | Benign | 0.01 | Affected | 0.1288 | 0.3187 | 0 | -2 | -3.8 | 60.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2462G>T | C821F 2D AIThe SynGAP1 missense variant C821F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any existing ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.745909 | Disordered | 0.672821 | Binding | 0.351 | 0.883 | 0.750 | 0.971 | Likely Benign | 0.924 | Likely Pathogenic | Ambiguous | 0.338 | Likely Benign | -3.33 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.66 | Benign | 0.01 | Affected | 0.1504 | 0.3505 | -4 | -2 | 0.3 | 44.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2463C>G | C821W 2D  AIThe SynGAP1 missense variant C821W is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated predictors (six pathogenic vs three benign) indicate a pathogenic impact. This prediction is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.745909 | Disordered | 0.672821 | Binding | 0.351 | 0.883 | 0.750 | -4.843 | Likely Benign | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.201 | Likely Benign | -3.30 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.64 | Benign | 0.01 | Affected | 0.1742 | 0.3268 | -8 | -2 | -3.4 | 83.07 | ||||||||||||||||||||||||||||||||||||||||
| c.2465C>G | T822R 2D AIThe SynGAP1 missense variant T822R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑vs‑2 tie and therefore unavailable; Foldetta predictions are not provided. Overall, the balance of evidence (five pathogenic versus three benign predictions) suggests the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.724957 | Disordered | 0.651624 | Binding | 0.295 | 0.881 | 0.750 | 0.414 | Likely Benign | 0.952 | Likely Pathogenic | Ambiguous | 0.237 | Likely Benign | -2.74 | Deleterious | 1.000 | Probably Damaging | 0.992 | Probably Damaging | 2.50 | Benign | 0.05 | Affected | 0.1182 | 0.3439 | -1 | -1 | -3.8 | 55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2465C>T | T822M 2D AIThe SynGAP1 missense variant T822M is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443017‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.724957 | Disordered | 0.651624 | Binding | 0.295 | 0.881 | 0.750 | 6-33443017-C-T | 5 | 3.10e-6 | -4.525 | Likely Benign | 0.833 | Likely Pathogenic | Ambiguous | 0.239 | Likely Benign | -2.11 | Neutral | 1.000 | Probably Damaging | 0.989 | Probably Damaging | 2.48 | Pathogenic | 0.03 | Affected | 3.77 | 5 | 0.1580 | 0.6379 | -1 | -1 | 2.6 | 30.09 | |||||||||||||||||||||||||||||||||||
| c.2468G>A | S823N 2D AIThe SynGAP1 missense variant S823N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic, two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the available predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.685117 | Disordered | 0.627336 | Binding | 0.358 | 0.884 | 0.750 | -6.880 | Likely Benign | 0.974 | Likely Pathogenic | Likely Pathogenic | 0.170 | Likely Benign | -1.76 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 1.93 | Pathogenic | 0.00 | Affected | 0.1370 | 0.5172 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2468G>C | S823T 2D AIThe SynGAP1 missense variant S823T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy consensus (SGM Consensus) derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN is inconclusive (two benign, two pathogenic). AlphaMissense‑Optimized predicts benign, while Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the majority of standard predictors indicate a pathogenic effect, and this assessment does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.685117 | Disordered | 0.627336 | Binding | 0.358 | 0.884 | 0.750 | -6.036 | Likely Benign | 0.717 | Likely Pathogenic | Likely Benign | 0.165 | Likely Benign | -2.02 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 2.05 | Pathogenic | 0.00 | Affected | 0.1453 | 0.6580 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2473T>A | S825T 2D AIThe SynGAP1 missense variant S825T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy consensus (SGM Consensus) derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN is inconclusive (two benign, two pathogenic). AlphaMissense‑Optimized, a high‑accuracy model, predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of standard tools (5 pathogenic vs. 4 benign) suggest a pathogenic impact, while the single high‑accuracy model indicates benign. Thus, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.675549 | Disordered | 0.618614 | Binding | 0.384 | 0.886 | 0.750 | -4.659 | Likely Benign | 0.615 | Likely Pathogenic | Likely Benign | 0.155 | Likely Benign | -2.26 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 2.07 | Pathogenic | 0.05 | Affected | 0.1432 | 0.6658 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2473T>G | S825A 2D AIThe SynGAP1 missense variant S825A is not reported in ClinVar (no entry) and is absent from gnomAD. Prediction tools that agree on benign include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on pathogenic are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of standard predictors indicate a pathogenic effect, while the high‑accuracy AlphaMissense‑Optimized suggests benign and the consensus is neutral. Given the preponderance of pathogenic predictions and the lack of conflicting evidence from ClinVar or population databases, the variant is most likely pathogenic, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.675549 | Disordered | 0.618614 | Binding | 0.384 | 0.886 | 0.750 | -4.878 | Likely Benign | 0.569 | Likely Pathogenic | Likely Benign | 0.135 | Likely Benign | -2.30 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 2.04 | Pathogenic | 0.05 | Affected | 0.4794 | 0.5749 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.2477A>C | D826A 2D AIThe SynGAP1 D826A missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated predictors (six pathogenic vs. three benign) indicate a pathogenic impact. This prediction is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.627309 | Binding | 0.327 | 0.886 | 0.625 | -5.590 | Likely Benign | 0.984 | Likely Pathogenic | Likely Pathogenic | 0.307 | Likely Benign | -3.77 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.53 | Benign | 0.02 | Affected | 0.4463 | 0.7867 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2477A>G | D826G 2D AIThe SynGAP1 D826G missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated predictors (six pathogenic vs. three benign) indicate a pathogenic impact. This prediction is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.627309 | Binding | 0.327 | 0.886 | 0.625 | -6.310 | Likely Benign | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.323 | Likely Benign | -2.94 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.51 | Benign | 0.01 | Affected | 0.4251 | 0.7480 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.247A>G | R83G 2D AIThe SynGAP1 R83G missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes) and is therefore treated as unavailable. Foldetta stability analysis is not provided and is likewise unavailable. Overall, the preponderance of evidence (six pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic, with no contradiction to ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.637480 | Disordered | 0.522784 | Binding | 0.275 | 0.895 | 0.250 | -4.708 | Likely Benign | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.111 | Likely Benign | -2.60 | Deleterious | 0.909 | Possibly Damaging | 0.587 | Possibly Damaging | 3.18 | Benign | 0.00 | Affected | 0.3514 | 0.2537 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.2482A>C | T828P 2D AIThe SynGAP1 missense variant T828P is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain. The SGM Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to benign (2 benign vs. 1 pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) is unavailable. Overall, the preponderance of evidence points to a benign impact for T828P, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.631236 | Binding | 0.321 | 0.879 | 0.500 | -3.902 | Likely Benign | 0.466 | Ambiguous | Likely Benign | 0.280 | Likely Benign | -2.53 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.61 | Benign | 0.12 | Tolerated | 0.1898 | 0.4424 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.2483C>T | T828I 2D AISynGAP1 missense variant T828I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM, whereas tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑to‑2 split and therefore unavailable; Foldetta predictions are not provided. Consequently, the computational evidence is evenly divided between benign and pathogenic, with no clear direction. The variant is therefore not clearly benign or pathogenic based on current predictions, and this lack of consensus does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.631236 | Binding | 0.321 | 0.879 | 0.500 | -4.971 | Likely Benign | 0.893 | Likely Pathogenic | Ambiguous | 0.139 | Likely Benign | -3.40 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.61 | Benign | 0.06 | Tolerated | 0.0767 | 0.5262 | 0 | -1 | 5.2 | 12.05 | ||||||||||||||||||||||||||||||||||||||||
| c.2485G>C | E829Q 2D AIThe SynGAP1 missense variant E829Q is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of reliable tools predict a benign impact, and there is no ClinVar entry to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.562014 | Disordered | 0.626045 | Binding | 0.326 | 0.882 | 0.375 | -4.985 | Likely Benign | 0.531 | Ambiguous | Likely Benign | 0.225 | Likely Benign | -1.83 | Neutral | 0.994 | Probably Damaging | 0.946 | Probably Damaging | 2.26 | Pathogenic | 0.00 | Affected | 0.1321 | 0.7248 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2489C>G | P830R 2D AIThe SynGAP1 missense variant P830R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of reliable predictors and the high‑accuracy consensus indicate a benign impact. This conclusion is not contradicted by any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.590140 | Disordered | 0.618152 | Binding | 0.333 | 0.874 | 0.500 | -5.919 | Likely Benign | 0.515 | Ambiguous | Likely Benign | 0.227 | Likely Benign | -3.51 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.72 | Benign | 0.00 | Affected | 0.1277 | 0.2984 | 0 | -2 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||||||||||||
| c.2489C>T | P830L 2D AIThe SynGAP1 missense variant P830L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign. Foldetta results are unavailable. Overall, the majority of reliable predictors lean toward a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.590140 | Disordered | 0.618152 | Binding | 0.333 | 0.874 | 0.500 | -3.990 | Likely Benign | 0.362 | Ambiguous | Likely Benign | 0.269 | Likely Benign | -5.31 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.65 | Benign | 0.00 | Affected | 0.2138 | 0.6631 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.248G>T | R83I 2D  AIThe SynGAP1 missense variant R83I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign). Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the preponderance of evidence (seven pathogenic vs. three benign predictions) indicates that R83I is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.637480 | Disordered | 0.522784 | Binding | 0.275 | 0.895 | 0.250 | -4.021 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.183 | Likely Benign | -3.15 | Deleterious | 0.972 | Probably Damaging | 0.766 | Possibly Damaging | 3.17 | Benign | 0.00 | Affected | 0.1450 | 0.3219 | -2 | -3 | 9.0 | -43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2491G>A | E831K 2D AIThe SynGAP1 missense variant E831K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, FATHMM, and AlphaMissense‑Default; ESM1b is uncertain. High‑accuracy assessment shows AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of conventional tools lean benign, but the high‑accuracy consensus and several individual pathogenic predictions suggest a pathogenic likelihood. Thus, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.626927 | Disordered | 0.617732 | Binding | 0.319 | 0.874 | 0.375 | -7.447 | In-Between | 0.636 | Likely Pathogenic | Likely Benign | 0.167 | Likely Benign | -1.43 | Neutral | 0.625 | Possibly Damaging | 0.252 | Benign | 2.37 | Pathogenic | 0.07 | Tolerated | 0.1989 | 0.6995 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.2491G>C | E831Q 2D AIThe SynGAP1 missense variant E831Q is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, favors a benign outcome (2 benign vs. 1 pathogenic). High‑accuracy methods give a benign prediction from AlphaMissense‑Optimized and a benign consensus from SGM; Foldetta results are unavailable. Overall, the available computational evidence points to a benign impact for E831Q, and this assessment does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.626927 | Disordered | 0.617732 | Binding | 0.319 | 0.874 | 0.375 | -5.604 | Likely Benign | 0.349 | Ambiguous | Likely Benign | 0.147 | Likely Benign | -1.23 | Neutral | 0.891 | Possibly Damaging | 0.492 | Possibly Damaging | 2.36 | Pathogenic | 0.09 | Tolerated | 0.1022 | 0.7052 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2492A>C | E831A 2D AIThe SynGAP1 missense variant E831A is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, SIFT, polyPhen‑2 HumVar, and ESM1b, while pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, and FATHMM. AlphaMissense‑Default is uncertain, whereas AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a pathogenic verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools therefore give conflicting signals: AlphaMissense‑Optimized benign versus SGM Consensus pathogenic, with no Foldetta data. Overall, the bulk of predictions lean toward a benign effect, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.626927 | Disordered | 0.617732 | Binding | 0.319 | 0.874 | 0.375 | -4.780 | Likely Benign | 0.429 | Ambiguous | Likely Benign | 0.115 | Likely Benign | -2.56 | Deleterious | 0.625 | Possibly Damaging | 0.315 | Benign | 2.36 | Pathogenic | 0.07 | Tolerated | 0.3845 | 0.6868 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2501T>A | M834K 2D AIThe SynGAP1 missense variant M834K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.585406 | Disordered | 0.640801 | Binding | 0.258 | 0.863 | 0.375 | -3.154 | Likely Benign | 0.453 | Ambiguous | Likely Benign | 0.154 | Likely Benign | -2.21 | Neutral | 0.369 | Benign | 0.150 | Benign | 2.43 | Pathogenic | 0.00 | Affected | 0.1131 | 0.0688 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||||||||||||
| c.2501T>G | M834R 2D AIThe SynGAP1 missense variant M834R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) leans toward benign (2 benign vs. 1 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence indicates a benign impact, and this conclusion does not contradict any ClinVar annotation, as none exists for M834R. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.585406 | Disordered | 0.640801 | Binding | 0.258 | 0.863 | 0.375 | -2.621 | Likely Benign | 0.449 | Ambiguous | Likely Benign | 0.148 | Likely Benign | -2.44 | Neutral | 0.812 | Possibly Damaging | 0.284 | Benign | 2.43 | Pathogenic | 0.00 | Affected | 0.1310 | 0.0837 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||||||||||||
| c.2506A>T | S836C 2D AIThe SynGAP1 missense variant S836C is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT; ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields benign, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.634582 | Binding | 0.269 | 0.859 | 0.250 | -7.859 | In-Between | 0.208 | Likely Benign | Likely Benign | 0.184 | Likely Benign | -2.66 | Deleterious | 0.997 | Probably Damaging | 0.923 | Probably Damaging | 2.50 | Benign | 0.04 | Affected | 0.0834 | 0.5468 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2507G>T | S836I 2D AIThe SynGAP1 missense variant S836I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Consensus from standard in silico predictors shows five tools (REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized) predict a benign effect, while two tools (PROVEAN, SIFT) predict pathogenicity. Two additional predictors (ESM1b, AlphaMissense‑Default) return uncertain results. High‑accuracy assessment further indicates a benign prediction from AlphaMissense‑Optimized; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (one benign, one pathogenic, two uncertain). Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a benign classification, and this conclusion does not conflict with the absence of a ClinVar annotation. Therefore, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.634582 | Binding | 0.269 | 0.859 | 0.250 | -7.751 | In-Between | 0.517 | Ambiguous | Likely Benign | 0.149 | Likely Benign | -3.33 | Deleterious | 0.057 | Benign | 0.053 | Benign | 2.51 | Benign | 0.03 | Affected | 0.0756 | 0.5023 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.250C>G | R84G 2D AIThe SynGAP1 missense variant R84G is listed in ClinVar with an “Uncertain” significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the majority of available predictions, the variant is most likely pathogenic, which does not contradict the current ClinVar status of “Uncertain.” Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.529205 | Binding | 0.298 | 0.888 | 0.500 | Uncertain | 1 | -6.627 | Likely Benign | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.139 | Likely Benign | -2.64 | Deleterious | 0.962 | Probably Damaging | 0.726 | Possibly Damaging | 3.68 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3387 | 0.3391 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||
| c.2513A>C | N838T 2D AIThe SynGAP1 missense variant N838T is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, favors a benign outcome (2 benign vs. 1 pathogenic, with one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions therefore point to a benign classification: AlphaMissense‑Optimized is benign, and the SGM Consensus is benign. Consequently, the variant is most likely benign based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.613320 | Binding | 0.276 | 0.861 | 0.250 | -4.847 | Likely Benign | 0.373 | Ambiguous | Likely Benign | 0.091 | Likely Benign | -2.59 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.69 | Benign | 0.12 | Tolerated | 0.1279 | 0.5586 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||||||||||||
| c.2519G>C | S840T 2D AIThe SynGAP1 missense variant S840T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic. Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.611356 | Binding | 0.259 | 0.865 | 0.250 | -7.243 | In-Between | 0.583 | Likely Pathogenic | Likely Benign | 0.170 | Likely Benign | -2.30 | Neutral | 0.951 | Possibly Damaging | 0.729 | Possibly Damaging | 1.55 | Pathogenic | 0.00 | Affected | 0.1068 | 0.5494 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.251G>A | R84H 2D AIThe SynGAP1 missense variant R84H is not reported in ClinVar (ClinVar status: not reported) and is absent from gnomAD (gnomAD: not present). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. Two tools give uncertain results: ESM1b and AlphaMissense‑Optimized. High‑accuracy assessment shows that AlphaMissense‑Optimized remains uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves as benign, and Foldetta (combining FoldX‑MD and Rosetta) has no available output. Overall, the majority of standard predictors (four pathogenic vs. three benign) lean toward a pathogenic interpretation, and the high‑accuracy consensus does not overturn this trend. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.529205 | Binding | 0.298 | 0.888 | 0.500 | -7.333 | In-Between | 0.935 | Likely Pathogenic | Ambiguous | 0.147 | Likely Benign | -1.77 | Neutral | 0.995 | Probably Damaging | 0.840 | Possibly Damaging | 3.68 | Benign | 0.00 | Affected | 0.2739 | 0.2426 | 2 | 0 | 1.3 | -19.05 | ||||||||||||||||||||||||||||||||||||||||
| c.251G>C | R84P 2D AIThe SynGAP1 missense variant R84P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. ESM1b is uncertain and does not influence the consensus. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a pathogenic prediction. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that R84P is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.529205 | Binding | 0.298 | 0.888 | 0.500 | -7.959 | In-Between | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.157 | Likely Benign | -2.50 | Deleterious | 0.989 | Probably Damaging | 0.859 | Possibly Damaging | 3.67 | Benign | 0.00 | Affected | 0.1978 | 0.4231 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||||||||||
| c.251G>T | R84L 2D AIThe SynGAP1 missense variant R84L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and FATHMM, while the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. ESM1b remains uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that R84L is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.529205 | Binding | 0.298 | 0.888 | 0.500 | -7.579 | In-Between | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.120 | Likely Benign | -2.75 | Deleterious | 0.962 | Probably Damaging | 0.726 | Possibly Damaging | 3.70 | Benign | 0.00 | Affected | 0.1595 | 0.4653 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2521G>A | V841M 2D AISynGAP1 variant V841M is listed in ClinVar with an uncertain significance and is present in gnomAD (6-33443073-G-A). Functional prediction tools cluster into two groups: benign predictions from REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The ESM1b score is inconclusive. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, while Foldetta stability analysis is unavailable. Taken together, the majority of evidence, including the high‑accuracy tools, points to a benign effect for V841M. This conclusion does not conflict with the ClinVar uncertain status, which reflects the current lack of definitive clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.616495 | Binding | 0.261 | 0.873 | 0.125 | Uncertain | 1 | 6-33443073-G-A | 3 | 1.86e-6 | -7.000 | In-Between | 0.651 | Likely Pathogenic | Likely Benign | 0.119 | Likely Benign | -0.74 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.54 | Benign | 0.02 | Affected | 3.77 | 5 | 0.0707 | 0.3937 | 1 | 2 | -2.3 | 32.06 | |||||||||||||||||||||||||||||||||
| c.2521G>C | V841L 2D AIThe SynGAP1 missense variant V841L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.616495 | Binding | 0.261 | 0.873 | 0.125 | -7.924 | In-Between | 0.514 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -0.01 | Neutral | 0.992 | Probably Damaging | 0.989 | Probably Damaging | 2.85 | Benign | 0.56 | Tolerated | 0.0850 | 0.4233 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2521G>T | V841L 2D AIThe SynGAP1 missense variant V841L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta results are unavailable. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.616495 | Binding | 0.261 | 0.873 | 0.125 | -7.924 | In-Between | 0.514 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -0.01 | Neutral | 0.992 | Probably Damaging | 0.989 | Probably Damaging | 2.85 | Benign | 0.56 | Tolerated | 0.0850 | 0.4233 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2522T>C | V841A 2D AIThe SynGAP1 missense variant V841A (ClinVar ID 1395978.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33443074‑T‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool reports an uncertain outcome, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—yields a tie (two pathogenic, two benign) and is therefore inconclusive. No Foldetta stability assessment is available for this variant. Overall, the balance of evidence favors a pathogenic interpretation, which does not contradict the current ClinVar designation of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.616495 | Binding | 0.261 | 0.873 | 0.125 | Uncertain | 1 | 6-33443074-T-C | 3 | 1.86e-6 | -8.152 | Likely Pathogenic | 0.901 | Likely Pathogenic | Ambiguous | 0.183 | Likely Benign | -2.13 | Neutral | 0.992 | Probably Damaging | 0.989 | Probably Damaging | 2.57 | Benign | 0.02 | Affected | 3.77 | 5 | 0.3069 | 0.2106 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||
| c.2522T>G | V841G 2D AIThe SynGAP1 missense variant V841G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and FATHMM, while the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—consistently predict a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, whereas the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for V841G, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.616495 | Binding | 0.261 | 0.873 | 0.125 | -10.054 | Likely Pathogenic | 0.913 | Likely Pathogenic | Ambiguous | 0.288 | Likely Benign | -4.11 | Deleterious | 0.997 | Probably Damaging | 0.999 | Probably Damaging | 2.51 | Benign | 0.00 | Affected | 0.2351 | 0.2422 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2528T>C | M843T 2D AIThe SynGAP1 missense variant M843T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL and FATHMM, and pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b remains uncertain. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic majority, and Foldetta data are unavailable. Overall, the preponderance of evidence indicates that M843T is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.585406 | Disordered | 0.617934 | Binding | 0.327 | 0.854 | 0.375 | -7.297 | In-Between | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.295 | Likely Benign | -3.22 | Deleterious | 0.968 | Probably Damaging | 0.954 | Probably Damaging | 2.62 | Benign | 0.00 | Affected | 0.2235 | 0.2037 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2531T>C | L844P 2D AIThe SynGAP1 missense variant L844P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Consensus from standard in‑silico predictors shows a split: benign calls come from REVEL and FATHMM, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b remains uncertain. High‑accuracy assessment further supports a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence from both conventional and high‑accuracy tools indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.595080 | Disordered | 0.611301 | Binding | 0.304 | 0.835 | 0.375 | -7.425 | In-Between | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.319 | Likely Benign | -3.18 | Deleterious | 0.986 | Probably Damaging | 0.876 | Possibly Damaging | 2.58 | Benign | 0.01 | Affected | 0.3528 | 0.1458 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2536T>A | L846I 2D AIThe SynGAP1 missense variant L846I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Two tools, ESM1b and AlphaMissense‑Default, return uncertain results. For high‑accuracy assessment, AlphaMissense‑Optimized classifies the variant as benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because the votes are split (one pathogenic, one benign, two uncertain). Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, has no available output for this variant. Overall, the balance of evidence from the majority of prediction tools suggests the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.653063 | Disordered | 0.589606 | Binding | 0.349 | 0.825 | 0.500 | -7.882 | In-Between | 0.474 | Ambiguous | Likely Benign | 0.151 | Likely Benign | -1.38 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.18 | Pathogenic | 0.00 | Affected | 0.0973 | 0.3475 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||||
| c.2539C>G | Q847E 2D AISynGAP1 missense variant Q847E is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The remaining tools, ESM1b and AlphaMissense‑Default, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is unavailable due to no majority, and Foldetta stability analysis is also unavailable. Overall, the predictions are split, providing no clear bias toward benign or pathogenic. Thus the variant is most likely of uncertain significance, which does not contradict its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.653063 | Disordered | 0.577677 | Binding | 0.282 | 0.818 | 0.500 | Uncertain | 1 | -7.864 | In-Between | 0.377 | Ambiguous | Likely Benign | 0.140 | Likely Benign | -2.12 | Neutral | 0.649 | Possibly Damaging | 0.535 | Possibly Damaging | 2.31 | Pathogenic | 0.00 | Affected | 0.1376 | 0.2083 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||||||||||
| c.253A>C | T85P 2D AIThe SynGAP1 missense variant T85P is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments are mixed: AlphaMissense‑Optimized predicts pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields no clear majority and is therefore unavailable; Foldetta results are not provided. Overall, the majority of available predictions (five pathogenic vs. four benign) indicate a pathogenic effect. There is no ClinVar entry to contradict this assessment, so the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.542004 | Binding | 0.288 | 0.888 | 0.500 | -8.406 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.125 | Likely Benign | -2.28 | Neutral | 0.813 | Possibly Damaging | 0.053 | Benign | 3.80 | Benign | 0.00 | Affected | 0.1499 | 0.4485 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.2540A>C | Q847P 2D AIThe SynGAP1 missense variant Q847P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑vs‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy predictors (AlphaMissense‑Optimized and AlphaMissense‑Default) indicate a benign outcome, while the consensus of other tools is mixed. Thus, the variant is most likely benign based on current predictions, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.653063 | Disordered | 0.577677 | Binding | 0.282 | 0.818 | 0.500 | -6.742 | Likely Benign | 0.320 | Likely Benign | Likely Benign | 0.331 | Likely Benign | -3.83 | Deleterious | 0.990 | Probably Damaging | 0.892 | Possibly Damaging | 2.26 | Pathogenic | 0.00 | Affected | 0.2290 | 0.4831 | 0 | -1 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2548G>T | G850W 2D AIThe SynGAP1 missense variant G850W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign, two uncertain), and Foldetta data are unavailable. Overall, the balance of evidence (four benign vs. three pathogenic predictions, with a benign high‑accuracy result and no contradictory ClinVar annotation) indicates that the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.648219 | Disordered | 0.540897 | Binding | 0.312 | 0.820 | 0.500 | -7.826 | In-Between | 0.351 | Ambiguous | Likely Benign | 0.170 | Likely Benign | -1.94 | Neutral | 0.996 | Probably Damaging | 0.933 | Probably Damaging | 4.16 | Benign | 0.00 | Affected | 0.0734 | 0.4002 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.254C>A | T85K 2D AIThe SynGAP1 missense variant T85K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments are mixed: AlphaMissense‑Optimized predicts pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields an equal split (2 pathogenic, 2 benign) and is therefore considered unavailable; Foldetta results are not provided and are likewise unavailable. Overall, the majority of available predictions (5 pathogenic vs. 4 benign) indicate that the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.542004 | Binding | 0.288 | 0.888 | 0.500 | -9.278 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.127 | Likely Benign | -2.32 | Neutral | 0.588 | Possibly Damaging | 0.036 | Benign | 3.88 | Benign | 0.00 | Affected | 0.0823 | 0.2796 | 0 | -1 | -3.2 | 27.07 | ||||||||||||||||||||||||||||||||||||||||
| c.254C>G | T85R 2D AIThe SynGAP1 missense variant T85R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools show a split: benign calls come from REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls come from polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b is uncertain. High‑accuracy assessment gives AlphaMissense‑Optimized a pathogenic score, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (benign), and PROVEAN (benign)—leans toward benign. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Overall, the balance of evidence tilts toward a benign interpretation, with no ClinVar entry to contradict this assessment. Thus, the variant is most likely benign, though the presence of pathogenic predictions from several tools indicates that further functional validation would be prudent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.542004 | Binding | 0.288 | 0.888 | 0.500 | -7.936 | In-Between | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.116 | Likely Benign | -2.32 | Neutral | 0.813 | Possibly Damaging | 0.072 | Benign | 3.91 | Benign | 0.00 | Affected | 0.0754 | 0.2482 | -1 | -1 | -3.8 | 55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2575A>C | S859R 2D AIThe SynGAP1 missense variant S859R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Uncertain predictions come from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessment shows that AlphaMissense‑Optimized is uncertain, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward benign (two benign versus one pathogenic vote). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.648219 | Disordered | 0.497075 | Uncertain | 0.288 | 0.819 | 0.375 | -7.810 | In-Between | 0.844 | Likely Pathogenic | Ambiguous | 0.127 | Likely Benign | -1.42 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 4.00 | Benign | 0.06 | Tolerated | 0.0945 | 0.4061 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2576G>T | S859I 2D AIThe SynGAP1 missense variant S859I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of computational evidence indicates a benign impact, and this conclusion does not contradict any ClinVar annotation because no ClinVar record exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.648219 | Disordered | 0.497075 | Uncertain | 0.288 | 0.819 | 0.375 | -8.342 | Likely Pathogenic | 0.351 | Ambiguous | Likely Benign | 0.256 | Likely Benign | -1.94 | Neutral | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 3.99 | Benign | 0.02 | Affected | 0.1094 | 0.5867 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2577T>A | S859R 2D AIThe SynGAP1 missense variant S859R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Uncertain predictions come from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessment shows that the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans benign (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact; this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.648219 | Disordered | 0.497075 | Uncertain | 0.288 | 0.819 | 0.375 | -7.810 | In-Between | 0.844 | Likely Pathogenic | Ambiguous | 0.120 | Likely Benign | -1.42 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 4.00 | Benign | 0.06 | Tolerated | 0.0945 | 0.4061 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2577T>G | S859R 2D AIThe SynGAP1 missense variant S859R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Uncertain predictions come from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessment shows that AlphaMissense‑Optimized remains uncertain, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward benign (two benign versus one pathogenic vote). Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.648219 | Disordered | 0.497075 | Uncertain | 0.288 | 0.819 | 0.375 | -7.810 | In-Between | 0.844 | Likely Pathogenic | Ambiguous | 0.120 | Likely Benign | -1.42 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 4.00 | Benign | 0.06 | Tolerated | 0.0945 | 0.4061 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2579T>G | V860G 2D AIThe SynGAP1 missense variant V860G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools cluster into two groups: the benign group includes REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; the pathogenic group contains only SIFT. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized is benign, the SGM Consensus (derived from the majority of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign, while Foldetta results are unavailable. Overall, the collective evidence indicates that V860G is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.545602 | Disordered | 0.518121 | Binding | 0.269 | 0.803 | 0.250 | -3.471 | Likely Benign | 0.173 | Likely Benign | Likely Benign | 0.198 | Likely Benign | -1.54 | Neutral | 0.012 | Benign | 0.009 | Benign | 4.11 | Benign | 0.00 | Affected | 0.2312 | 0.2547 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.257T>A | V86D 2D  AIThe SynGAP1 missense variant V86D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on benign impact include REVEL, PROVEAN, and FATHMM, whereas those that agree on pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b is uncertain. High‑accuracy assessment shows AlphaMissense‑Optimized predicts pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans benign (two benign, one pathogenic, one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of conventional tools predict pathogenicity, whereas the high‑accuracy consensus suggests benign. Based on the aggregate predictions, the variant is most likely pathogenic, and this assessment does not contradict ClinVar status, which currently has no entry for V86D. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.685117 | Disordered | 0.552911 | Binding | 0.295 | 0.887 | 0.500 | -7.141 | In-Between | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.147 | Likely Benign | -2.38 | Neutral | 0.824 | Possibly Damaging | 0.485 | Possibly Damaging | 3.73 | Benign | 0.00 | Affected | 0.1590 | 0.1047 | -2 | -3 | -7.7 | 15.96 | ||||||||||||||||||||||||||||||||||||||||
| c.257T>G | V86G 2D AIThe SynGAP1 missense variant V86G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. Foldetta, which evaluates protein‑folding stability via FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of evidence (five pathogenic versus four benign predictions) indicates that V86G is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.685117 | Disordered | 0.552911 | Binding | 0.295 | 0.887 | 0.500 | -4.212 | Likely Benign | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.130 | Likely Benign | -2.50 | Deleterious | 0.307 | Benign | 0.824 | Possibly Damaging | 3.74 | Benign | 0.00 | Affected | 0.2321 | 0.2547 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2584A>T | N862Y 2D AIThe SynGAP1 missense variant N862Y is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence—especially the high‑accuracy consensus—suggests the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.564559 | Binding | 0.257 | 0.791 | 0.250 | -8.200 | Likely Pathogenic | 0.485 | Ambiguous | Likely Benign | 0.216 | Likely Benign | -3.06 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 4.01 | Benign | 0.06 | Tolerated | 0.0871 | 0.6246 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||||||||||||
| c.2585A>T | N862I 2D AIThe SynGAP1 missense variant N862I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; AlphaMissense‑Default is uncertain. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a pathogenic majority (2 pathogenic vs. 1 benign, 1 uncertain). High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM Consensus as pathogenic, and Foldetta results are unavailable. Overall, the majority of predictions support a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.564559 | Binding | 0.257 | 0.791 | 0.250 | -8.702 | Likely Pathogenic | 0.561 | Ambiguous | Likely Benign | 0.195 | Likely Benign | -3.19 | Deleterious | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 4.03 | Benign | 0.03 | Affected | 0.0844 | 0.6443 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.2586C>A | N862K 2D AIThe SynGAP1 missense variant N862K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools cluster into two groups: benign predictions from REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. ESM1b is uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.564559 | Binding | 0.257 | 0.791 | 0.250 | -7.000 | In-Between | 0.766 | Likely Pathogenic | Likely Benign | 0.084 | Likely Benign | -1.87 | Neutral | 0.995 | Probably Damaging | 0.946 | Probably Damaging | 4.10 | Benign | 0.20 | Tolerated | 0.2313 | 0.5469 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.2586C>G | N862K 2D AIThe SynGAP1 missense variant N862K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools cluster into two groups: benign predictions from REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. ESM1b is uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.564559 | Binding | 0.257 | 0.791 | 0.250 | -7.000 | In-Between | 0.766 | Likely Pathogenic | Likely Benign | 0.084 | Likely Benign | -1.87 | Neutral | 0.995 | Probably Damaging | 0.946 | Probably Damaging | 4.10 | Benign | 0.20 | Tolerated | 0.2313 | 0.5469 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.2591C>A | A864E 2D AIThe SynGAP1 missense variant A864E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors a benign outcome. Foldetta, which evaluates protein‑folding stability, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.549308 | Disordered | 0.611966 | Binding | 0.269 | 0.788 | 0.250 | -5.508 | Likely Benign | 0.350 | Ambiguous | Likely Benign | 0.153 | Likely Benign | 0.10 | Neutral | 0.138 | Benign | 0.070 | Benign | 2.45 | Pathogenic | 0.04 | Affected | 0.1204 | 0.2037 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2597T>G | V866G 2D AIThe SynGAP1 missense variant V866G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign impact for V866G, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.638070 | Binding | 0.266 | 0.788 | 0.250 | -1.519 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -2.30 | Neutral | 0.912 | Possibly Damaging | 0.993 | Probably Damaging | 2.69 | Benign | 0.07 | Tolerated | 0.2548 | 0.2157 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.259T>C | S87P 2D AIThe SynGAP1 missense variant S87P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Based on the available predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.550904 | Binding | 0.302 | 0.878 | 0.500 | -8.566 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.073 | Likely Benign | -2.04 | Neutral | 0.676 | Possibly Damaging | 0.307 | Benign | 3.75 | Benign | 0.00 | Affected | 0.1639 | 0.4260 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||||
| c.259T>G | S87A 2D AIThe SynGAP1 missense variant S87A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default, while ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence (six benign predictions versus two pathogenic) indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.550904 | Binding | 0.302 | 0.878 | 0.500 | -7.817 | In-Between | 0.676 | Likely Pathogenic | Likely Benign | 0.039 | Likely Benign | -1.24 | Neutral | 0.140 | Benign | 0.097 | Benign | 3.84 | Benign | 0.00 | Affected | 0.4272 | 0.3540 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.2600G>T | G867V 2D AIThe SynGAP1 missense variant G867V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split between benign and uncertain calls. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence (five benign predictions versus two pathogenic predictions, with no decisive high‑accuracy support for pathogenicity) indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.517562 | Disordered | 0.657954 | Binding | 0.285 | 0.801 | 0.250 | -7.049 | In-Between | 0.441 | Ambiguous | Likely Benign | 0.111 | Likely Benign | -2.23 | Neutral | 0.999 | Probably Damaging | 0.958 | Probably Damaging | 2.70 | Benign | 0.10 | Tolerated | 0.1125 | 0.4613 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2602G>C | D868H 2D AIThe SynGAP1 missense variant D868H is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (variant ID 6‑33443154‑G‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, and this is not contradicted by ClinVar status. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.676362 | Binding | 0.262 | 0.815 | 0.250 | 6-33443154-G-C | 1 | 6.20e-7 | -3.358 | Likely Benign | 0.763 | Likely Pathogenic | Likely Benign | 0.212 | Likely Benign | -2.34 | Neutral | 1.000 | Probably Damaging | 0.983 | Probably Damaging | 2.49 | Pathogenic | 0.08 | Tolerated | 3.82 | 4 | 0.2258 | 0.7837 | -1 | 1 | 0.3 | 22.05 | |||||||||||||||||||||||||||||||||||
| c.2603A>C | D868A 2D AIThe SynGAP1 D868A variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the balance of evidence—including the benign prediction from the most accurate AlphaMissense‑Optimized model and the majority of benign calls—suggests that the variant is most likely benign. This conclusion does not contradict any ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.676362 | Binding | 0.262 | 0.815 | 0.250 | -2.764 | Likely Benign | 0.702 | Likely Pathogenic | Likely Benign | 0.139 | Likely Benign | -2.75 | Deleterious | 0.972 | Probably Damaging | 0.760 | Possibly Damaging | 2.54 | Benign | 0.21 | Tolerated | 0.4590 | 0.6990 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2603A>G | D868G 2D AIThe SynGAP1 missense variant D868G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain. The high‑accuracy consensus (SGM Consensus) – derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN – is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the available predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.525368 | Disordered | 0.676362 | Binding | 0.262 | 0.815 | 0.250 | -2.218 | Likely Benign | 0.552 | Ambiguous | Likely Benign | 0.113 | Likely Benign | -2.71 | Deleterious | 0.986 | Probably Damaging | 0.860 | Possibly Damaging | 2.51 | Benign | 0.21 | Tolerated | 0.4658 | 0.6414 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.260C>A | S87Y 2D AIThe SynGAP1 missense variant S87Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy predictors (six out of nine) indicate a pathogenic impact, whereas three predict benign. Therefore, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.550904 | Binding | 0.302 | 0.878 | 0.500 | -10.410 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.052 | Likely Benign | -2.20 | Neutral | 0.880 | Possibly Damaging | 0.608 | Possibly Damaging | 3.75 | Benign | 0.00 | Affected | 0.0522 | 0.4798 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.260C>G | S87C 2D AIThe SynGAP1 missense variant S87C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy consensus methods give the following results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is uncertain due to a 2‑to‑2 split; and Foldetta, which assesses protein‑folding stability, is unavailable for this variant. Overall, the majority of available predictions (five pathogenic versus three benign) indicate a likely pathogenic impact. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.550904 | Binding | 0.302 | 0.878 | 0.500 | -8.769 | Likely Pathogenic | 0.915 | Likely Pathogenic | Ambiguous | 0.095 | Likely Benign | -2.14 | Neutral | 0.880 | Possibly Damaging | 0.700 | Possibly Damaging | 3.74 | Benign | 0.00 | Affected | 0.0794 | 0.4849 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.260C>T | S87F 2D AIThe SynGAP1 missense variant S87F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictors (six out of nine) indicate a pathogenic impact, whereas three predict benign. Therefore, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.550904 | Binding | 0.302 | 0.878 | 0.500 | -9.673 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.054 | Likely Benign | -2.34 | Neutral | 0.676 | Possibly Damaging | 0.485 | Possibly Damaging | 3.74 | Benign | 0.00 | Affected | 0.0493 | 0.4937 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2617A>C | S873R 2D AIThe SynGAP1 missense variant S873R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the balance of evidence (five pathogenic vs four benign predictions) suggests the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.414856 | Structured | 0.649816 | Binding | 0.283 | 0.866 | 0.125 | -5.856 | Likely Benign | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.218 | Likely Benign | -2.74 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.67 | Benign | 0.06 | Tolerated | 3.77 | 5 | 0.0970 | 0.3667 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||
| c.2617A>T | S873C 2D AIThe SynGAP1 missense variant S873C is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b) predict a pathogenic impact; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a pathogenic view: AlphaMissense‑Optimized predicts benign, while the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence leans toward pathogenicity, and this conclusion does not contradict the ClinVar status, which currently contains no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.414856 | Structured | 0.649816 | Binding | 0.283 | 0.866 | 0.125 | -8.293 | Likely Pathogenic | 0.502 | Ambiguous | Likely Benign | 0.224 | Likely Benign | -2.71 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.62 | Benign | 0.04 | Affected | 0.1124 | 0.5887 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2619C>A | S873R 2D AIThe SynGAP1 missense variant S873R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the balance of evidence (five pathogenic vs. four benign predictions, with a pathogenic high‑accuracy tool) suggests the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.414856 | Structured | 0.649816 | Binding | 0.283 | 0.866 | 0.125 | -5.856 | Likely Benign | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.192 | Likely Benign | -2.74 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.67 | Benign | 0.06 | Tolerated | 3.77 | 5 | 0.0970 | 0.3667 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||
| c.2619C>G | S873R 2D AIThe SynGAP1 missense variant S873R is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33443171‑C‑G). Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign votes), and Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a pathogenic effect, which contrasts with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.414856 | Structured | 0.649816 | Binding | 0.283 | 0.866 | 0.125 | Uncertain | 1 | 6-33443171-C-G | 1 | 6.20e-7 | -5.856 | Likely Benign | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.192 | Likely Benign | -2.74 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.67 | Benign | 0.06 | Tolerated | 3.77 | 5 | 0.0970 | 0.3667 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||
| c.2621A>T | Q874L 2D AIThe SynGAP1 missense variant Q874L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs four benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.490133 | Structured | 0.635258 | Binding | 0.289 | 0.873 | 0.250 | -6.084 | Likely Benign | 0.608 | Likely Pathogenic | Likely Benign | 0.199 | Likely Benign | -3.39 | Deleterious | 0.985 | Probably Damaging | 0.982 | Probably Damaging | 2.67 | Benign | 0.00 | Affected | 0.0828 | 0.6595 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||||||||||||
| c.2622G>C | Q874H 2D AIThe SynGAP1 missense variant Q874H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign, while Foldetta results are unavailable. Overall, the balance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.490133 | Structured | 0.635258 | Binding | 0.289 | 0.873 | 0.250 | -4.658 | Likely Benign | 0.543 | Ambiguous | Likely Benign | 0.236 | Likely Benign | -2.90 | Deleterious | 0.996 | Probably Damaging | 0.995 | Probably Damaging | 2.65 | Benign | 0.00 | Affected | 0.1588 | 0.5088 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2622G>T | Q874H 2D AIThe SynGAP1 missense variant Q874H has no ClinVar record and is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, while PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT all predict a pathogenic outcome; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority, and Foldetta data are unavailable. Overall, the balance of evidence leans toward a benign impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.490133 | Structured | 0.635258 | Binding | 0.289 | 0.873 | 0.250 | -4.658 | Likely Benign | 0.543 | Ambiguous | Likely Benign | 0.236 | Likely Benign | -2.90 | Deleterious | 0.996 | Probably Damaging | 0.995 | Probably Damaging | 2.65 | Benign | 0.00 | Affected | 0.1588 | 0.5088 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2624C>A | A875D 2D AIThe SynGAP1 missense variant A875D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2) and Foldetta data are unavailable. Overall, the majority of standard tools (5 pathogenic vs 4 benign) lean toward a pathogenic interpretation, but the high‑accuracy predictions are mixed or missing. Thus, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.545602 | Disordered | 0.632173 | Binding | 0.273 | 0.872 | 0.250 | -3.268 | Likely Benign | 0.645 | Likely Pathogenic | Likely Benign | 0.149 | Likely Benign | -2.78 | Deleterious | 0.992 | Probably Damaging | 0.913 | Probably Damaging | 2.68 | Benign | 0.02 | Affected | 0.1648 | 0.2035 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2627C>T | S876L 2D AISynGAP1 variant S876L is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect; the SGM Consensus, derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive and therefore unavailable; Foldetta stability analysis is also unavailable. Overall, the majority of available predictions favor a benign impact, suggesting the variant is most likely benign. This conclusion does not conflict with the ClinVar uncertain status, which reflects the current lack of definitive evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.549308 | Disordered | 0.631130 | Binding | 0.280 | 0.872 | 0.250 | Uncertain | 2 | -5.856 | Likely Benign | 0.489 | Ambiguous | Likely Benign | 0.249 | Likely Benign | -3.56 | Deleterious | 0.998 | Probably Damaging | 0.992 | Probably Damaging | 2.57 | Benign | 0.05 | Affected | 3.77 | 5 | 0.1268 | 0.6053 | -2 | -3 | 4.6 | 26.08 | ||||||||||||||||||||||||||||||||||||
| c.263T>A | V88E 2D  AIThe SynGAP1 missense variant V88E is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) favors a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of predictions (five benign vs. three pathogenic) and the SGM Consensus support a benign classification, whereas AlphaMissense‑Optimized alone suggests pathogenicity. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.703578 | Disordered | 0.552910 | Binding | 0.323 | 0.870 | 0.500 | -7.978 | In-Between | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.095 | Likely Benign | -1.95 | Neutral | 0.001 | Benign | 0.000 | Benign | 3.68 | Benign | 0.00 | Affected | 0.1444 | 0.1725 | -2 | -2 | -7.7 | 29.98 | ||||||||||||||||||||||||||||||||||||||||
| c.263T>G | V88G 2D AIThe SynGAP1 missense variant V88G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2) and Foldetta results are unavailable. Overall, the majority of predictions (5 benign vs. 4 pathogenic) and the lack of a ClinVar pathogenic classification suggest that V88G is most likely benign, with no contradiction to existing ClinVar data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.703578 | Disordered | 0.552910 | Binding | 0.323 | 0.870 | 0.500 | -8.588 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.084 | Likely Benign | -2.40 | Neutral | 0.024 | Benign | 0.208 | Benign | 3.68 | Benign | 0.00 | Affected | 0.2608 | 0.2609 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2644G>A | G882R 2D AIThe SynGAP1 missense variant G882R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.632888 | Binding | 0.306 | 0.878 | 0.250 | -4.715 | Likely Benign | 0.758 | Likely Pathogenic | Likely Benign | 0.051 | Likely Benign | -1.48 | Neutral | 0.001 | Benign | 0.003 | Benign | 2.04 | Pathogenic | 0.01 | Affected | 0.0905 | 0.4126 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.2644G>C | G882R 2D AIThe SynGAP1 missense variant G882R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the majority of predictions (six benign vs three pathogenic) support a benign classification. This consensus does not contradict any ClinVar status, as the variant is not yet catalogued there. Thus, the variant is most likely benign based on current predictive evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.632888 | Binding | 0.306 | 0.878 | 0.250 | -4.715 | Likely Benign | 0.758 | Likely Pathogenic | Likely Benign | 0.051 | Likely Benign | -1.48 | Neutral | 0.001 | Benign | 0.003 | Benign | 2.04 | Pathogenic | 0.01 | Affected | 0.0905 | 0.4126 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.2645G>A | G882E 2D AIThe SynGAP1 missense variant G882E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split; Foldetta results are unavailable. Overall, more tools (six) predict benign than pathogenic (three), and no ClinVar evidence contradicts this assessment. Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.632888 | Binding | 0.306 | 0.878 | 0.250 | -4.246 | Likely Benign | 0.613 | Likely Pathogenic | Likely Benign | 0.079 | Likely Benign | -1.16 | Neutral | 0.004 | Benign | 0.005 | Benign | 2.04 | Pathogenic | 0.01 | Affected | 0.1263 | 0.3244 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2657C>A | A886E 2D AIThe SynGAP1 missense variant A886E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome. Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.716283 | Disordered | 0.619166 | Binding | 0.359 | 0.922 | 0.500 | -3.747 | Likely Benign | 0.422 | Ambiguous | Likely Benign | 0.096 | Likely Benign | -1.31 | Neutral | 0.423 | Benign | 0.230 | Benign | 2.17 | Pathogenic | 0.00 | Affected | 0.1483 | 0.2008 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2665G>A | G889R 2D AIThe SynGAP1 missense variant G889R is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (ID 6‑33443217‑G‑A). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized; pathogenic predictions come from SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta results are unavailable. Overall, the majority of evidence (six benign vs three pathogenic) supports a benign impact. This conclusion does not contradict ClinVar, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.552581 | Binding | 0.331 | 0.928 | 0.625 | 6-33443217-G-A | 1 | 6.20e-7 | -3.357 | Likely Benign | 0.685 | Likely Pathogenic | Likely Benign | 0.070 | Likely Benign | -1.96 | Neutral | 0.027 | Benign | 0.009 | Benign | 2.38 | Pathogenic | 0.02 | Affected | 4.32 | 4 | 0.0925 | 0.4165 | -2 | -3 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||
| c.2665G>C | G889R 2D AIThe SynGAP1 missense variant G889R is not reported in ClinVar (ClinVar status: not present) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.552581 | Binding | 0.331 | 0.928 | 0.625 | -3.357 | Likely Benign | 0.685 | Likely Pathogenic | Likely Benign | 0.072 | Likely Benign | -1.96 | Neutral | 0.027 | Benign | 0.009 | Benign | 2.38 | Pathogenic | 0.02 | Affected | 4.32 | 4 | 0.0925 | 0.4165 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||
| c.2666G>A | G889E 2D AIThe SynGAP1 missense variant G889E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions lean toward a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.552581 | Binding | 0.331 | 0.928 | 0.625 | -5.352 | Likely Benign | 0.572 | Likely Pathogenic | Likely Benign | 0.083 | Likely Benign | -1.96 | Neutral | 0.611 | Possibly Damaging | 0.187 | Benign | 2.41 | Pathogenic | 0.02 | Affected | 0.1442 | 0.3719 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2668C>T | R890C 2D AIThe SynGAP1 missense variant R890C is listed in ClinVar as benign and is present in gnomAD (6-33443220-C‑T). Functional prediction tools show mixed results: benign predictions come from REVEL, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Pathogenic predictions are reported by PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus also indicates benign; Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence leans toward a benign effect, which is consistent with the ClinVar classification and does not contradict the reported status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.720929 | Disordered | 0.531156 | Binding | 0.284 | 0.928 | 0.625 | Benign | 1 | 6-33443220-C-T | 9 | 5.58e-6 | -5.786 | Likely Benign | 0.402 | Ambiguous | Likely Benign | 0.200 | Likely Benign | -3.38 | Deleterious | 1.000 | Probably Damaging | 0.971 | Probably Damaging | 3.94 | Benign | 0.04 | Affected | 4.32 | 4 | 0.3626 | 0.2206 | -4 | -3 | 7.0 | -53.05 | |||||||||||||||||||||||||||||||||
| c.2669G>T | R890L 2D AIThe SynGAP1 missense variant R890L is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a benign prediction (2 benign vs. 1 pathogenic, with one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy and consensus predictions indicate a benign impact. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.720929 | Disordered | 0.531156 | Binding | 0.284 | 0.928 | 0.625 | -2.387 | Likely Benign | 0.389 | Ambiguous | Likely Benign | 0.213 | Likely Benign | -2.74 | Deleterious | 0.990 | Probably Damaging | 0.921 | Probably Damaging | 3.98 | Benign | 0.20 | Tolerated | 0.1876 | 0.3406 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.266C>A | P89Q 2D AIThe SynGAP1 missense variant P89Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the balance of evidence (five pathogenic vs. four benign predictions) indicates the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.703578 | Disordered | 0.545797 | Binding | 0.316 | 0.865 | 0.500 | -4.779 | Likely Benign | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.107 | Likely Benign | -2.63 | Deleterious | 0.642 | Possibly Damaging | 0.038 | Benign | 3.74 | Benign | 0.00 | Affected | 0.1633 | 0.4310 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||||||
| c.266C>G | P89R 2D AIThe SynGAP1 missense variant P89R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the balance of evidence (five pathogenic vs. four benign predictions) indicates the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.703578 | Disordered | 0.545797 | Binding | 0.316 | 0.865 | 0.500 | -3.636 | Likely Benign | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.116 | Likely Benign | -3.00 | Deleterious | 0.642 | Possibly Damaging | 0.097 | Benign | 3.83 | Benign | 0.00 | Affected | 0.1720 | 0.3362 | 0 | -2 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||||||||||||
| c.266C>T | P89L 2D AISynGAP1 missense variant P89L is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 HumVar, ESM1b, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments indicate that AlphaMissense‑Optimized predicts pathogenicity, whereas the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of tools favor a pathogenic effect, but the evidence is not unanimous. Therefore, the variant is most likely pathogenic according to the current predictions, and this assessment does not contradict its ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.703578 | Disordered | 0.545797 | Binding | 0.316 | 0.865 | 0.500 | Uncertain | 2 | -6.775 | Likely Benign | 0.982 | Likely Pathogenic | Likely Pathogenic | 0.119 | Likely Benign | -3.29 | Deleterious | 0.889 | Possibly Damaging | 0.058 | Benign | 3.73 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2399 | 0.5638 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||
| c.2675C>T | S892F 2D AIThe SynGAP1 missense variant S892F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2). Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs four benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.754692 | Disordered | 0.473390 | Uncertain | 0.319 | 0.926 | 0.875 | -4.709 | Likely Benign | 0.605 | Likely Pathogenic | Likely Benign | 0.090 | Likely Benign | -3.07 | Deleterious | 0.998 | Probably Damaging | 0.959 | Probably Damaging | 2.55 | Benign | 0.00 | Affected | 0.0715 | 0.5124 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2680G>T | G894W 2D AIThe SynGAP1 missense variant G894W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs. two benign votes). Foldetta, which assesses protein‑folding stability, has no available output for this variant. Overall, the majority of evaluated predictors (five pathogenic vs. three benign) lean toward a pathogenic interpretation. Because there is no ClinVar annotation to contradict this assessment, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.788093 | Disordered | 0.425700 | Uncertain | 0.310 | 0.925 | 0.750 | -6.927 | Likely Benign | 0.814 | Likely Pathogenic | Ambiguous | 0.177 | Likely Benign | -2.70 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.59 | Benign | 0.00 | Affected | 0.0698 | 0.4184 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.2681G>T | G894V 2D AIThe SynGAP1 missense variant G894V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also leans benign, and Foldetta results are unavailable. Overall, the majority of high‑confidence tools predict a benign impact, and there is no conflict with the ClinVar status. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.788093 | Disordered | 0.425700 | Uncertain | 0.310 | 0.925 | 0.750 | -5.047 | Likely Benign | 0.395 | Ambiguous | Likely Benign | 0.188 | Likely Benign | -2.61 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.64 | Benign | 0.02 | Affected | 0.1187 | 0.4091 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2686G>C | G896R 2D AIThe SynGAP1 missense variant G896R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and ESM1b, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Consequently, the evidence is evenly split between benign and pathogenic predictions, with no decisive support from the high‑accuracy or folding‑stability analyses. The variant is therefore most likely of uncertain significance; it does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.675549 | Disordered | 0.412816 | Uncertain | 0.314 | 0.923 | 0.625 | -4.511 | Likely Benign | 0.897 | Likely Pathogenic | Ambiguous | 0.218 | Likely Benign | -2.45 | Neutral | 0.999 | Probably Damaging | 0.967 | Probably Damaging | 2.44 | Pathogenic | 0.14 | Tolerated | 0.0931 | 0.4228 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.2687G>A | G896D 2D AIThe SynGAP1 missense variant G896D is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and ESM1b, whereas polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default all predict a pathogenic outcome. The high‑accuracy AlphaMissense‑Optimized score is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive, and Foldetta stability analysis is unavailable. Consequently, the evidence is evenly split between benign and pathogenic predictions, with no decisive support from the most reliable methods. The variant is therefore classified as of uncertain significance; it does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.675549 | Disordered | 0.412816 | Uncertain | 0.314 | 0.923 | 0.625 | -4.500 | Likely Benign | 0.905 | Likely Pathogenic | Ambiguous | 0.159 | Likely Benign | -2.39 | Neutral | 0.997 | Probably Damaging | 0.934 | Probably Damaging | 2.44 | Pathogenic | 0.16 | Tolerated | 0.1707 | 0.1842 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2693C>G | S898C 2D AIThe SynGAP1 missense variant S898C is catalogued in gnomAD (ID 6‑33443245‑C‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM, while ESM1b remains uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus also indicates benign. Foldetta, a protein‑folding stability predictor that integrates FoldX‑MD and Rosetta outputs, did not return a result for this variant, so its stability impact is unavailable. Overall, the preponderance of evidence points to a benign effect, and this assessment does not conflict with ClinVar, which currently has no classification for S898C. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.426070 | Uncertain | 0.305 | 0.922 | 0.500 | 6-33443245-C-G | 1 | 6.20e-7 | -7.007 | In-Between | 0.257 | Likely Benign | Likely Benign | 0.146 | Likely Benign | -2.43 | Neutral | 0.999 | Probably Damaging | 0.986 | Probably Damaging | 2.43 | Pathogenic | 0.01 | Affected | 4.32 | 4 | 0.1487 | 0.6166 | -1 | 0 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||
| c.269T>G | V90G 2D AIThe SynGAP1 missense variant V90G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign verdict. Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.570702 | Disordered | 0.542047 | Binding | 0.343 | 0.873 | 0.500 | -2.617 | Likely Benign | 0.358 | Ambiguous | Likely Benign | 0.094 | Likely Benign | -0.55 | Neutral | 0.024 | Benign | 0.208 | Benign | 4.00 | Benign | 0.00 | Affected | 0.2047 | 0.2936 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2707G>C | G903R 2D AIThe SynGAP1 missense variant G903R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of standard predictors lean toward a benign impact, and no ClinVar annotation contradicts this assessment. Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.549818 | Binding | 0.291 | 0.917 | 0.375 | -3.503 | Likely Benign | 0.865 | Likely Pathogenic | Ambiguous | 0.119 | Likely Benign | -2.02 | Neutral | 0.241 | Benign | 0.244 | Benign | 2.33 | Pathogenic | 0.02 | Affected | 0.0872 | 0.4064 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.2707G>T | G903C 2D AIThe SynGAP1 missense variant G903C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain and does not influence the consensus. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, which would provide a protein‑folding stability estimate, is unavailable for this variant. Overall, the majority of evidence (five pathogenic versus three benign predictions) points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.549818 | Binding | 0.291 | 0.917 | 0.375 | -6.593 | Likely Benign | 0.471 | Ambiguous | Likely Benign | 0.151 | Likely Benign | -3.28 | Deleterious | 1.000 | Probably Damaging | 0.990 | Probably Damaging | 2.32 | Pathogenic | 0.02 | Affected | 0.1158 | 0.4420 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2708G>A | G903D 2D AIThe SynGAP1 missense variant G903D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward a pathogenic interpretation. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.549818 | Binding | 0.291 | 0.917 | 0.375 | -4.481 | Likely Benign | 0.849 | Likely Pathogenic | Ambiguous | 0.137 | Likely Benign | -1.85 | Neutral | 0.997 | Probably Damaging | 0.939 | Probably Damaging | 2.33 | Pathogenic | 0.01 | Affected | 0.1535 | 0.1321 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2708G>T | G903V 2D AIThe SynGAP1 missense variant G903V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, while Foldetta results are unavailable. Overall, the balance of evidence (five benign predictions versus three pathogenic predictions, with no conflicting ClinVar annotation) indicates that the variant is most likely benign. This conclusion does not contradict any ClinVar status, as the variant is not currently catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.549818 | Binding | 0.291 | 0.917 | 0.375 | -4.750 | Likely Benign | 0.491 | Ambiguous | Likely Benign | 0.168 | Likely Benign | -1.93 | Neutral | 0.997 | Probably Damaging | 0.959 | Probably Damaging | 2.44 | Pathogenic | 0.95 | Tolerated | 0.1269 | 0.4266 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2713C>T | R905C 2D AIThe SynGAP1 missense variant R905C (ClinVar ID 469152.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33443265‑C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of standard predictors indicate a pathogenic impact, whereas the high‑accuracy AlphaMissense‑Optimized tool suggests a benign effect. Consequently, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict the ClinVar status of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.671169 | Disordered | 0.618085 | Binding | 0.291 | 0.920 | 0.250 | Conflicting | 2 | 6-33443265-C-T | 15 | 9.31e-6 | -5.578 | Likely Benign | 0.723 | Likely Pathogenic | Likely Benign | 0.194 | Likely Benign | -3.14 | Deleterious | 1.000 | Probably Damaging | 0.980 | Probably Damaging | 2.57 | Benign | 0.01 | Affected | 3.77 | 5 | 0.3231 | 0.3642 | -4 | -3 | 7.0 | -53.05 | |||||||||||||||||||||||||||||||||
| c.2714G>T | R905L 2D AIThe SynGAP1 missense variant R905L is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (allele ID 6‑33443266‑G‑T). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of predictions leans toward a benign interpretation, and this conclusion does not contradict the ClinVar status, which currently has no assertion for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.671169 | Disordered | 0.618085 | Binding | 0.291 | 0.920 | 0.250 | 6-33443266-G-T | -3.284 | Likely Benign | 0.709 | Likely Pathogenic | Likely Benign | 0.242 | Likely Benign | -2.55 | Deleterious | 0.963 | Probably Damaging | 0.753 | Possibly Damaging | 2.61 | Benign | 0.10 | Tolerated | 3.77 | 5 | 0.1676 | 0.4905 | -2 | -3 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||
| c.2717T>A | L906H 2D AIThe SynGAP1 missense variant L906H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes) and Foldetta results are unavailable. Overall, the majority of standard predictors lean toward pathogenicity, but the most reliable high‑accuracy tool suggests a benign outcome and the consensus is unresolved. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.644316 | Binding | 0.315 | 0.920 | 0.250 | -4.367 | Likely Benign | 0.732 | Likely Pathogenic | Likely Benign | 0.124 | Likely Benign | -1.01 | Neutral | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 2.18 | Pathogenic | 0.02 | Affected | 0.1125 | 0.1073 | -2 | -3 | -7.0 | 23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2717T>C | L906P 2D AIThe SynGAP1 missense variant L906P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors a benign outcome. Foldetta, which would provide a protein‑folding stability estimate, is unavailable for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.644316 | Binding | 0.315 | 0.920 | 0.250 | -3.662 | Likely Benign | 0.478 | Ambiguous | Likely Benign | 0.150 | Likely Benign | 0.08 | Neutral | 1.000 | Probably Damaging | 0.993 | Probably Damaging | 2.18 | Pathogenic | 0.10 | Tolerated | 0.3430 | 0.1458 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2717T>G | L906R 2D AIThe SynGAP1 missense variant L906R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of standard predictors (5 vs 4) lean toward pathogenicity, while the high‑accuracy AlphaMissense‑Optimized predicts benign and the consensus remains unresolved. Thus, the variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.604312 | Disordered | 0.644316 | Binding | 0.315 | 0.920 | 0.250 | -3.440 | Likely Benign | 0.769 | Likely Pathogenic | Likely Benign | 0.160 | Likely Benign | -0.26 | Neutral | 1.000 | Probably Damaging | 0.990 | Probably Damaging | 2.18 | Pathogenic | 0.04 | Affected | 0.1228 | 0.0947 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2728G>T | G910C 2D AIThe SynGAP1 missense variant G910C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta data are unavailable. Overall, the majority of tools (five pathogenic vs four benign) lean toward a pathogenic interpretation, and the high‑accuracy benign prediction from AlphaMissense‑Optimized does not override this trend. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.707319 | Binding | 0.264 | 0.917 | 0.250 | -6.359 | Likely Benign | 0.723 | Likely Pathogenic | Likely Benign | 0.252 | Likely Benign | -3.11 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.69 | Benign | 0.01 | Affected | 0.1179 | 0.4194 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2729G>T | G910V 2D AIThe SynGAP1 missense variant G910V is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443281‑G‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of tools (five pathogenic vs. four benign) predict a pathogenic impact. This prediction does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.707319 | Binding | 0.264 | 0.917 | 0.250 | 6-33443281-G-T | 1 | 6.20e-7 | -4.362 | Likely Benign | 0.724 | Likely Pathogenic | Likely Benign | 0.237 | Likely Benign | -2.69 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.73 | Benign | 0.02 | Affected | 3.77 | 5 | 0.1179 | 0.3662 | -3 | -1 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||
| c.2732T>G | V911G 2D AIThe SynGAP1 missense variant V911G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). All available in silico predictors classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity, so the benign group includes every listed predictor, while the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign; the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.685117 | Disordered | 0.724137 | Binding | 0.327 | 0.914 | 0.375 | -2.754 | Likely Benign | 0.222 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -0.04 | Neutral | 0.004 | Benign | 0.003 | Benign | 2.74 | Benign | 0.16 | Tolerated | 0.2307 | 0.3360 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2740G>T | D914Y 2D AIThe SynGAP1 missense variant D914Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of tools (five pathogenic vs. four benign) and the lack of a benign consensus from the high‑accuracy methods suggest that D914Y is most likely pathogenic. This prediction does not contradict ClinVar status, as the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.699094 | Disordered | 0.785987 | Binding | 0.320 | 0.892 | 0.250 | -4.768 | Likely Benign | 0.740 | Likely Pathogenic | Likely Benign | 0.239 | Likely Benign | -2.65 | Deleterious | 1.000 | Probably Damaging | 0.993 | Probably Damaging | 2.62 | Benign | 0.01 | Affected | 0.0907 | 0.7583 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2747T>G | V916G 2D AIThe SynGAP1 missense variant V916G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.642678 | Disordered | 0.835395 | Binding | 0.308 | 0.879 | 0.250 | -1.950 | Likely Benign | 0.091 | Likely Benign | Likely Benign | 0.093 | Likely Benign | -0.63 | Neutral | 0.666 | Possibly Damaging | 0.917 | Probably Damaging | 2.69 | Benign | 0.05 | Affected | 0.2420 | 0.2681 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.274G>T | G92W 2D AIThe SynGAP1 missense variant G92W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) suggest a pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.575842 | Disordered | 0.537848 | Binding | 0.337 | 0.874 | 0.625 | -6.502 | Likely Benign | 0.789 | Likely Pathogenic | Ambiguous | 0.224 | Likely Benign | -2.61 | Deleterious | 1.000 | Probably Damaging | 0.979 | Probably Damaging | 3.95 | Benign | 0.00 | Affected | 0.0640 | 0.4546 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.2762T>C | L921P 2D AIThe SynGAP1 missense variant L921P is recorded in gnomAD (ID 6‑33443314‑T‑C) but has no ClinVar submission. Functional prediction tools cluster into two groups: benign calls from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized; pathogenic calls from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because the votes are split (two benign, one pathogenic, one uncertain). Foldetta, a protein‑folding stability predictor that combines FoldX‑MD and Rosetta outputs, has no reported result for this variant. Consequently, the evidence is evenly divided, with high‑accuracy tools not providing a decisive verdict. The variant is therefore inconclusive; it does not contradict ClinVar status, which currently has no entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.608892 | Disordered | 0.943282 | Binding | 0.311 | 0.845 | 0.375 | 6-33443314-T-C | 1 | 6.20e-7 | -2.087 | Likely Benign | 0.393 | Ambiguous | Likely Benign | 0.215 | Likely Benign | -1.83 | Neutral | 0.998 | Probably Damaging | 0.958 | Probably Damaging | 2.39 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3581 | 0.1343 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||
| c.2762T>G | L921R 2D AIThe SynGAP1 missense variant L921R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, and Foldetta results are unavailable. Overall, the majority of high‑confidence tools predict a benign impact, and there is no conflict with ClinVar status. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.608892 | Disordered | 0.943282 | Binding | 0.311 | 0.845 | 0.375 | -2.205 | Likely Benign | 0.563 | Ambiguous | Likely Benign | 0.217 | Likely Benign | -1.19 | Neutral | 0.994 | Probably Damaging | 0.912 | Probably Damaging | 2.41 | Pathogenic | 0.00 | Affected | 0.1306 | 0.0761 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2773C>A | L925I 2D AIThe SynGAP1 missense variant L925I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of consensus tools (five pathogenic vs. three benign) lean toward a pathogenic interpretation. This prediction does not contradict ClinVar status, as the variant is not yet catalogued there. Thus, based on current computational evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.483068 | Structured | 0.977963 | Binding | 0.290 | 0.852 | 0.125 | -5.266 | Likely Benign | 0.828 | Likely Pathogenic | Ambiguous | 0.205 | Likely Benign | -1.38 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.37 | Pathogenic | 0.00 | Affected | 0.1192 | 0.3405 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||||
| c.2773C>G | L925V 2D AIThe SynGAP1 missense variant L925V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of consensus tools predict a pathogenic impact, and this prediction does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.483068 | Structured | 0.977963 | Binding | 0.290 | 0.852 | 0.125 | -3.977 | Likely Benign | 0.831 | Likely Pathogenic | Ambiguous | 0.244 | Likely Benign | -2.09 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.36 | Pathogenic | 0.00 | Affected | 0.1848 | 0.3230 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2776T>A | S926T 2D AIThe SynGAP1 missense variant S926T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy consensus (SGM Consensus) is inconclusive, with two pathogenic and two benign votes. AlphaMissense‑Optimized, a high‑accuracy model, predicts benign, while Foldetta stability analysis is unavailable. Overall, the majority of standard tools favor a pathogenic effect, whereas the single high‑accuracy model suggests benign. Thus, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.461924 | Structured | 0.981753 | Binding | 0.295 | 0.854 | 0.250 | -2.917 | Likely Benign | 0.651 | Likely Pathogenic | Likely Benign | 0.226 | Likely Benign | -2.34 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 1.55 | Pathogenic | 0.00 | Affected | 0.1174 | 0.5663 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2776T>G | S926A 2D AIThe SynGAP1 missense variant S926A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of reliable predictions indicate a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.461924 | Structured | 0.981753 | Binding | 0.295 | 0.854 | 0.250 | -3.042 | Likely Benign | 0.463 | Ambiguous | Likely Benign | 0.186 | Likely Benign | -2.13 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 1.59 | Pathogenic | 0.00 | Affected | 0.4657 | 0.4488 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.2780T>A | F927Y 2D AIThe SynGAP1 missense variant F927Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas tools that agree on a pathogenic effect include polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (5 pathogenic vs. 3 benign) lean toward a pathogenic interpretation. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.529623 | Disordered | 0.985043 | Binding | 0.324 | 0.854 | 0.250 | -5.244 | Likely Benign | 0.954 | Likely Pathogenic | Ambiguous | 0.291 | Likely Benign | -2.29 | Neutral | 0.997 | Probably Damaging | 0.987 | Probably Damaging | 1.40 | Pathogenic | 0.00 | Affected | 0.1551 | 0.1518 | 7 | 3 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.2782C>G | Q928E 2D AIThe SynGAP1 missense variant Q928E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2) and Foldetta results are unavailable. Based on the overall distribution of predictions, the variant is most likely pathogenic; this assessment does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.497853 | Structured | 0.986260 | Binding | 0.324 | 0.852 | 0.250 | -5.168 | Likely Benign | 0.590 | Likely Pathogenic | Likely Benign | 0.283 | Likely Benign | -2.06 | Neutral | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 1.58 | Pathogenic | 0.00 | Affected | 0.1368 | 0.3477 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2791C>T | L931F 2D AIThe SynGAP1 missense variant L931F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and ESM1b, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. A high‑accuracy consensus (SGM) that aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a 2‑to‑2 split, leaving the consensus inconclusive. Foldetta, which assesses protein‑folding stability via FoldX‑MD and Rosetta, has no available result for this variant. Consequently, the overall evidence leans toward pathogenicity, driven by the majority of standard predictors, and does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.549308 | Disordered | 0.989212 | Binding | 0.335 | 0.856 | 0.375 | -5.101 | Likely Benign | 0.790 | Likely Pathogenic | Ambiguous | 0.167 | Likely Benign | -2.00 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.41 | Pathogenic | 0.04 | Affected | 0.0718 | 0.3208 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.2797C>A | H933N 2D AIThe SynGAP1 missense variant H933N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions show AlphaMissense‑Optimized as benign, while SGM Consensus and Foldetta are unavailable. Overall, five tools predict pathogenicity versus four predicting benignity, suggesting the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.987531 | Binding | 0.305 | 0.862 | 0.625 | -4.333 | Likely Benign | 0.226 | Likely Benign | Likely Benign | 0.261 | Likely Benign | -3.65 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.39 | Pathogenic | 0.04 | Affected | 0.1897 | 0.3439 | 2 | 1 | -0.3 | -23.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2797C>T | H933Y 2D AIThe SynGAP1 H933Y variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic prediction. Foldetta results are unavailable. Overall, the majority of evidence (five pathogenic vs. three benign predictions) points to a pathogenic impact for H933Y. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.987531 | Binding | 0.305 | 0.862 | 0.625 | -3.952 | Likely Benign | 0.525 | Ambiguous | Likely Benign | 0.314 | Likely Benign | -3.49 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.36 | Pathogenic | 0.01 | Affected | 0.0968 | 0.4815 | 0 | 2 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2798A>C | H933P 2D AIThe SynGAP1 missense variant H933P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, six tools predict pathogenicity versus three predicting benign, and the high‑accuracy benign prediction is outweighed by the majority of pathogenic calls. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.987531 | Binding | 0.305 | 0.862 | 0.625 | -3.890 | Likely Benign | 0.332 | Likely Benign | Likely Benign | 0.508 | Likely Pathogenic | -5.39 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.36 | Pathogenic | 0.02 | Affected | 0.1821 | 0.4400 | 0 | -2 | 1.6 | -40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.2798A>T | H933L 2D AIThe SynGAP1 H933L missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.987531 | Binding | 0.305 | 0.862 | 0.625 | -0.858 | Likely Benign | 0.470 | Ambiguous | Likely Benign | 0.388 | Likely Benign | -6.26 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.38 | Pathogenic | 0.02 | Affected | 0.1003 | 0.5749 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2799C>A | H933Q 2D  AIThe SynGAP1 missense variant H933Q has no ClinVar record and is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to benign (two benign versus one pathogenic vote). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) is unavailable for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.987531 | Binding | 0.305 | 0.862 | 0.625 | -3.042 | Likely Benign | 0.410 | Ambiguous | Likely Benign | 0.211 | Likely Benign | -2.98 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.54 | Benign | 0.53 | Tolerated | 0.1654 | 0.3938 | 3 | 0 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2799C>G | H933Q 2D AIThe SynGAP1 H933Q missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction between the predictions and ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.987531 | Binding | 0.305 | 0.862 | 0.625 | -3.042 | Likely Benign | 0.410 | Ambiguous | Likely Benign | 0.210 | Likely Benign | -2.98 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.54 | Benign | 0.53 | Tolerated | 0.1654 | 0.3938 | 3 | 0 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2802G>A | M934I 2D AIThe SynGAP1 missense variant M934I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are unavailable. Overall, the majority of evidence (seven benign versus two pathogenic predictions) indicates that M934I is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.984677 | Binding | 0.290 | 0.867 | 0.625 | -4.582 | Likely Benign | 0.757 | Likely Pathogenic | Likely Benign | 0.086 | Likely Benign | -1.78 | Neutral | 0.316 | Benign | 0.101 | Benign | 2.49 | Pathogenic | 0.27 | Tolerated | 0.1686 | 0.3100 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2802G>C | M934I 2D AIThe SynGAP1 missense variant M934I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are unavailable. Overall, the majority of evidence (seven benign versus two pathogenic predictions) indicates that M934I is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.984677 | Binding | 0.290 | 0.867 | 0.625 | -4.582 | Likely Benign | 0.757 | Likely Pathogenic | Likely Benign | 0.085 | Likely Benign | -1.78 | Neutral | 0.316 | Benign | 0.101 | Benign | 2.49 | Pathogenic | 0.27 | Tolerated | 0.1686 | 0.3100 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2802G>T | M934I 2D AIThe SynGAP1 missense variant M934I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are unavailable. Overall, the majority of evidence (seven benign versus two pathogenic predictions) indicates that M934I is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.984677 | Binding | 0.290 | 0.867 | 0.625 | -4.582 | Likely Benign | 0.757 | Likely Pathogenic | Likely Benign | 0.085 | Likely Benign | -1.78 | Neutral | 0.316 | Benign | 0.101 | Benign | 2.49 | Pathogenic | 0.27 | Tolerated | 0.1686 | 0.3100 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2803G>C | A935P 2D AIThe SynGAP1 missense variant A935P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, and Foldetta results are unavailable. Overall, the majority of high‑confidence tools predict a benign impact, and there is no conflict with ClinVar status. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.736850 | Disordered | 0.980490 | Binding | 0.286 | 0.865 | 0.625 | -3.239 | Likely Benign | 0.371 | Ambiguous | Likely Benign | 0.269 | Likely Benign | -2.08 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.29 | Pathogenic | 0.00 | Affected | 0.2062 | 0.5466 | 1 | -1 | -3.4 | 26.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2804C>T | A935V 2D AIThe SynGAP1 missense variant A935V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also leans benign (2 benign vs. 1 pathogenic votes). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for A935V, and this conclusion does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.736850 | Disordered | 0.980490 | Binding | 0.286 | 0.865 | 0.625 | -4.750 | Likely Benign | 0.397 | Ambiguous | Likely Benign | 0.194 | Likely Benign | -2.06 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1340 | 0.5941 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||||||
| c.2807C>A | A936D 2D AIThe SynGAP1 missense variant A936D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact. This conclusion does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.812494 | Disordered | 0.973218 | Binding | 0.319 | 0.874 | 0.625 | -4.162 | Likely Benign | 0.686 | Likely Pathogenic | Likely Benign | 0.115 | Likely Benign | -1.63 | Neutral | 0.801 | Possibly Damaging | 0.339 | Benign | 2.48 | Pathogenic | 0.02 | Affected | 0.1778 | 0.1660 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2809G>T | D937Y 2D AIThe SynGAP1 missense variant D937Y is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic versus four benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic based on current computational evidence, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.819762 | Disordered | 0.963385 | Binding | 0.348 | 0.883 | 0.625 | -4.720 | Likely Benign | 0.711 | Likely Pathogenic | Likely Benign | 0.137 | Likely Benign | -2.52 | Deleterious | 1.000 | Probably Damaging | 0.983 | Probably Damaging | 2.67 | Benign | 0.05 | Affected | 0.1305 | 0.7494 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2812G>T | G938W 2D AIThe SynGAP1 missense variant G938W is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar annotation exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.905695 | Disordered | 0.949795 | Binding | 0.318 | 0.883 | 0.625 | -8.763 | Likely Pathogenic | 0.604 | Likely Pathogenic | Likely Benign | 0.194 | Likely Benign | -1.71 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.70 | Benign | 0.04 | Affected | 0.0762 | 0.4008 | -7 | -2 | -0.5 | 129.16 | ||||||||||||||||||||||||||||||||||||||||
| c.2815C>A | P939T 2D AIThe SynGAP1 missense variant P939T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions show AlphaMissense‑Optimized as benign, while the SGM Consensus remains inconclusive and Foldetta is unavailable. Overall, more tools (five) predict pathogenicity than benign (four), suggesting the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.894241 | Disordered | 0.935841 | Binding | 0.397 | 0.897 | 0.625 | -5.739 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.139 | Likely Benign | -3.34 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.22 | Pathogenic | 0.00 | Affected | 0.1788 | 0.5310 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.2815C>G | P939A 2D AIThe SynGAP1 missense variant P939A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign versus two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools therefore provide a benign prediction from AlphaMissense‑Optimized, an inconclusive SGM Consensus, and no Foldetta data. Overall, the majority of individual predictors (five pathogenic versus four benign) lean toward a pathogenic interpretation, and this does not contradict the lack of ClinVar annotation. **Thus, the variant is most likely pathogenic based on the current computational predictions.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.894241 | Disordered | 0.935841 | Binding | 0.397 | 0.897 | 0.625 | -4.614 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.111 | Likely Benign | -3.57 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.3565 | 0.4336 | 1 | -1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2815C>T | P939S 2D AIThe SynGAP1 missense variant P939S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools specifically show AlphaMissense‑Optimized as benign, while SGM Consensus and Foldetta remain unavailable. Overall, the majority of predictions (5 pathogenic vs. 4 benign) indicate that P939S is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.894241 | Disordered | 0.935841 | Binding | 0.397 | 0.897 | 0.625 | -4.331 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.101 | Likely Benign | -3.44 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.24 | Pathogenic | 0.00 | Affected | 0.3372 | 0.4517 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2816C>A | P939Q 2D AIThe SynGAP1 missense variant P939Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus remains inconclusive and Foldetta is unavailable. Overall, five of the nine evaluated tools predict pathogenicity versus four predicting benignity, giving a slight tilt toward a pathogenic interpretation. This prediction does not contradict ClinVar status, as no ClinVar entry exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.894241 | Disordered | 0.935841 | Binding | 0.397 | 0.897 | 0.625 | -4.950 | Likely Benign | 0.131 | Likely Benign | Likely Benign | 0.156 | Likely Benign | -3.45 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.18 | Pathogenic | 0.00 | Affected | 0.1551 | 0.4342 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||||||
| c.2816C>G | P939R 2D AIThe SynGAP1 missense variant P939R is reported in gnomAD (ID 6‑33443368‑C‑G) but has no ClinVar entry. Functional prediction tools show mixed results: benign calls come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Grouping by consensus, the benign‑predicted tools outnumbered the pathogenic ones by one, but the overall tally favors pathogenicity (5 pathogenic vs 4 benign). High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is benign, but the SGM Consensus (a 2‑vs‑2 majority among AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields no clear verdict, and Foldetta data are unavailable. Consequently, the variant is most likely pathogenic according to the available predictions, and this assessment does not contradict any ClinVar status because none is reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.894241 | Disordered | 0.935841 | Binding | 0.397 | 0.897 | 0.625 | 6-33443368-C-G | 1 | 6.20e-7 | -4.863 | Likely Benign | 0.260 | Likely Benign | Likely Benign | 0.199 | Likely Benign | -3.79 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.18 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1360 | 0.3336 | -2 | 0 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||
| c.2816C>T | P939L 2D AIThe SynGAP1 missense variant P939L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions show AlphaMissense‑Optimized as benign, while SGM Consensus and Foldetta are unavailable. Overall, five tools predict pathogenicity versus four predicting benign, so the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.894241 | Disordered | 0.935841 | Binding | 0.397 | 0.897 | 0.625 | -4.191 | Likely Benign | 0.138 | Likely Benign | Likely Benign | 0.159 | Likely Benign | -3.69 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.18 | Pathogenic | 0.00 | Affected | 0.2113 | 0.5142 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2819G>A | G940D 2D AIThe SynGAP1 missense variant G940D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443371‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. Two tools, AlphaMissense‑Default and ESM1b, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it receives two benign and two uncertain votes, and Foldetta’s protein‑folding stability analysis is unavailable. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.889439 | Disordered | 0.920635 | Binding | 0.383 | 0.902 | 0.625 | 6-33443371-G-A | 6 | 3.72e-6 | -7.311 | In-Between | 0.397 | Ambiguous | Likely Benign | 0.076 | Likely Benign | -0.68 | Neutral | 0.770 | Possibly Damaging | 0.583 | Possibly Damaging | 2.72 | Benign | 0.17 | Tolerated | 3.77 | 5 | 0.1837 | 0.2252 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||
| c.2839G>A | G947R 2D AIThe SynGAP1 missense variant G947R is listed in gnomAD (6-33443391-G-A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized) and pathogenic (SIFT). Two tools remain uncertain (ESM1b, AlphaMissense‑Default). High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—does not reach a definitive conclusion (two benign, two uncertain). Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no reported result for this variant. Overall, the preponderance of evidence points to a benign impact, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988695 | Disordered | 0.850554 | Binding | 0.358 | 0.919 | 0.750 | 6-33443391-G-A | 6 | 3.72e-6 | -7.481 | In-Between | 0.343 | Ambiguous | Likely Benign | 0.273 | Likely Benign | -0.60 | Neutral | 0.411 | Benign | 0.140 | Benign | 4.94 | Benign | 0.04 | Affected | 4.32 | 4 | 0.1034 | 0.4733 | -2 | -3 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||
| c.2839G>C | G947R 2D AIThe SynGAP1 missense variant G947R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988695 | Disordered | 0.850554 | Binding | 0.358 | 0.919 | 0.750 | -7.481 | In-Between | 0.343 | Ambiguous | Likely Benign | 0.273 | Likely Benign | -0.60 | Neutral | 0.411 | Benign | 0.140 | Benign | 4.94 | Benign | 0.04 | Affected | 4.32 | 4 | 0.1034 | 0.4733 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||
| c.2845G>C | G949R 2D AIThe SynGAP1 missense variant G949R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Consensus from standard in silico predictors shows a split: benign calls come from REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic calls come from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; ESM1b is uncertain. High‑accuracy assessment further supports a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign outcome, and Foldetta data are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this conclusion does not conflict with any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988861 | Disordered | 0.874971 | Binding | 0.365 | 0.923 | 0.750 | -7.650 | In-Between | 0.301 | Likely Benign | Likely Benign | 0.317 | Likely Benign | -0.14 | Neutral | 0.997 | Probably Damaging | 0.934 | Probably Damaging | 2.21 | Pathogenic | 0.01 | Affected | 0.1095 | 0.4569 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.2845G>T | G949C 2D AIThe SynGAP1 missense variant G949C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. four benign) indicate that the variant is most likely pathogenic. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988861 | Disordered | 0.874971 | Binding | 0.365 | 0.923 | 0.750 | -10.580 | Likely Pathogenic | 0.091 | Likely Benign | Likely Benign | 0.345 | Likely Benign | -1.02 | Neutral | 1.000 | Probably Damaging | 0.975 | Probably Damaging | 2.21 | Pathogenic | 0.01 | Affected | 0.1480 | 0.4097 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2846G>A | G949D 2D AIThe SynGAP1 missense variant G949D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictors (5 pathogenic vs. 4 benign) indicate a pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988861 | Disordered | 0.874971 | Binding | 0.365 | 0.923 | 0.750 | -8.692 | Likely Pathogenic | 0.239 | Likely Benign | Likely Benign | 0.316 | Likely Benign | 0.25 | Neutral | 0.945 | Possibly Damaging | 0.753 | Possibly Damaging | 2.21 | Pathogenic | 0.02 | Affected | 0.1927 | 0.2826 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2846G>T | G949V 2D AIThe SynGAP1 missense variant G949V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988861 | Disordered | 0.874971 | Binding | 0.365 | 0.923 | 0.750 | -7.364 | In-Between | 0.102 | Likely Benign | Likely Benign | 0.328 | Likely Benign | -0.55 | Neutral | 0.992 | Probably Damaging | 0.834 | Possibly Damaging | 2.21 | Pathogenic | 0.01 | Affected | 0.1493 | 0.3165 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2848G>T | G950C 2D AIThe SynGAP1 missense variant G950C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictors (five pathogenic vs. four benign) indicate a pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.987317 | Disordered | 0.888649 | Binding | 0.368 | 0.923 | 0.750 | -9.589 | Likely Pathogenic | 0.092 | Likely Benign | Likely Benign | 0.395 | Likely Benign | -0.35 | Neutral | 0.983 | Probably Damaging | 0.750 | Possibly Damaging | 2.26 | Pathogenic | 0.01 | Affected | 0.1406 | 0.4439 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||||||||||||
| c.2849G>A | G950D 2D AIThe SynGAP1 missense variant G950D is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM, while ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.987317 | Disordered | 0.888649 | Binding | 0.368 | 0.923 | 0.750 | -7.515 | In-Between | 0.245 | Likely Benign | Likely Benign | 0.402 | Likely Benign | -0.61 | Neutral | 0.411 | Benign | 0.131 | Benign | 2.26 | Pathogenic | 0.02 | Affected | 0.1874 | 0.2826 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2849G>T | G950V 2D AIThe SynGAP1 missense variant G950V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM, while ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign prediction. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that G950V is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.987317 | Disordered | 0.888649 | Binding | 0.368 | 0.923 | 0.750 | -7.796 | In-Between | 0.093 | Likely Benign | Likely Benign | 0.428 | Likely Benign | -0.91 | Neutral | 0.411 | Benign | 0.239 | Benign | 2.26 | Pathogenic | 0.01 | Affected | 0.1392 | 0.3507 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2864C>G | S955C 2D AIThe SynGAP1 missense variant S955C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (5 pathogenic vs. 4 benign) indicate a likely pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.984871 | Disordered | 0.945325 | Binding | 0.350 | 0.924 | 0.750 | -8.675 | Likely Pathogenic | 0.117 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -1.48 | Neutral | 0.977 | Probably Damaging | 0.796 | Possibly Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.1741 | 0.5470 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2864C>T | S955F 2D AISynGAP1 missense variant S955F is listed in ClinVar as uncertain and is present in gnomAD (ID 6‑33443416‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM; ESM1b is inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also returns benign, and Foldetta results are unavailable. Overall, the majority of high‑confidence predictions favor a benign impact, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.984871 | Disordered | 0.945325 | Binding | 0.350 | 0.924 | 0.750 | Conflicting | 4 | 6-33443416-C-T | 95 | 5.89e-5 | -7.374 | In-Between | 0.176 | Likely Benign | Likely Benign | 0.093 | Likely Benign | -1.73 | Neutral | 0.977 | Probably Damaging | 0.721 | Possibly Damaging | 2.32 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1246 | 0.4943 | -3 | -2 | 3.6 | 60.10 | |||||||||||||||||||||||||||||||||
| c.2867C>G | S956C 2D AIThe SynGAP1 missense variant S956C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. four benign) indicate a likely pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.984871 | Disordered | 0.957345 | Binding | 0.364 | 0.917 | 0.750 | -9.292 | Likely Pathogenic | 0.108 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -0.34 | Neutral | 0.938 | Possibly Damaging | 0.665 | Possibly Damaging | 1.94 | Pathogenic | 0.03 | Affected | 0.1833 | 0.5470 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.2869C>G | H957D 2D AIThe SynGAP1 missense variant H957D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence (7 benign vs 2 pathogenic) supports a benign classification. This prediction is consistent with the lack of ClinVar annotation and gnomAD presence, indicating no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.985964 | Disordered | 0.968874 | Binding | 0.362 | 0.915 | 0.750 | -8.777 | Likely Pathogenic | 0.237 | Likely Benign | Likely Benign | 0.149 | Likely Benign | -0.77 | Neutral | 0.144 | Benign | 0.058 | Benign | 2.45 | Pathogenic | 0.44 | Tolerated | 0.2646 | 0.3066 | 1 | -1 | -0.3 | -22.05 | ||||||||||||||||||||||||||||||||||||||||
| c.2947A>G | S983G 2D AIThe SynGAP1 missense variant S983G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2). High‑accuracy methods show AlphaMissense‑Optimized as benign; the SGM Consensus is unavailable, and Foldetta results are not provided, so no folding‑stability evidence is available. Overall, the balance of evidence (five pathogenic vs four benign predictions) suggests the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.707965 | Disordered | 0.960212 | Binding | 0.277 | 0.889 | 0.625 | -5.206 | Likely Benign | 0.638 | Likely Pathogenic | Likely Benign | 0.109 | Likely Benign | -2.22 | Neutral | 0.979 | Probably Damaging | 0.982 | Probably Damaging | 2.17 | Pathogenic | 0.00 | Affected | 0.2521 | 0.4480 | 1 | 0 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2948G>A | S983N 2D AISynGAP1 missense variant S983N is listed as Benign in ClinVar (ID 469153) and is present in gnomAD (6‑33443500‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive; Foldetta results are not available. Overall, the majority of available predictions (five pathogenic vs. three benign) suggest a pathogenic impact, which contradicts the ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.707965 | Disordered | 0.960212 | Binding | 0.277 | 0.889 | 0.625 | Likely Benign | 1 | 6-33443500-G-A | 6 | 3.72e-6 | -5.604 | Likely Benign | 0.909 | Likely Pathogenic | Ambiguous | 0.136 | Likely Benign | -1.78 | Neutral | 0.991 | Probably Damaging | 0.988 | Probably Damaging | 2.04 | Pathogenic | 0.00 | Affected | 4.32 | 1 | 0.1933 | 0.4069 | 1 | 1 | -2.7 | 27.03 | |||||||||||||||||||||||||||||||||
| c.2948G>C | S983T 2D AIThe SynGAP1 missense variant S983T is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus and Foldetta (which would combine FoldX‑MD and Rosetta outputs) are unavailable. Overall, the balance of evidence leans toward pathogenicity, with five tools supporting a deleterious effect versus four supporting benign. This prediction does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.707965 | Disordered | 0.960212 | Binding | 0.277 | 0.889 | 0.625 | -4.493 | Likely Benign | 0.588 | Likely Pathogenic | Likely Benign | 0.172 | Likely Benign | -1.65 | Neutral | 0.979 | Probably Damaging | 0.982 | Probably Damaging | 2.08 | Pathogenic | 0.00 | Affected | 0.2042 | 0.5517 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2954G>T | S985I 2D AIThe SynGAP1 missense variant S985I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. ESM1b remains uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that S985I is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.720929 | Disordered | 0.941547 | Binding | 0.302 | 0.896 | 0.750 | -7.858 | In-Between | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.131 | Likely Benign | -2.78 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.1367 | 0.5206 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2956G>A | E986K 2D AIThe SynGAP1 missense variant E986K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and PROVEAN, while those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The remaining tools, ESM1b and AlphaMissense‑Optimized, return uncertain results and are not considered evidence for either side. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a pathogenic verdict (2 pathogenic vs. 1 benign, with one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy predictors classify the variant as pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.750527 | Disordered | 0.929726 | Binding | 0.349 | 0.902 | 0.750 | -7.174 | In-Between | 0.950 | Likely Pathogenic | Ambiguous | 0.164 | Likely Benign | -2.19 | Neutral | 0.924 | Possibly Damaging | 0.722 | Possibly Damaging | 2.15 | Pathogenic | 0.00 | Affected | 0.2760 | 0.7973 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.2956G>C | E986Q 2D AIThe SynGAP1 missense variant E986Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward a pathogenic interpretation. This assessment does not contradict ClinVar status, as the variant is not yet catalogued there. Thus, based on current computational evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.750527 | Disordered | 0.929726 | Binding | 0.349 | 0.902 | 0.750 | -4.471 | Likely Benign | 0.839 | Likely Pathogenic | Ambiguous | 0.160 | Likely Benign | -1.66 | Neutral | 0.974 | Probably Damaging | 0.842 | Possibly Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.1700 | 0.7589 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2957A>C | E986A 2D AIThe SynGAP1 missense variant E986A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and ESM1b, whereas tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta stability analysis is unavailable. Overall, the evidence is evenly divided, with no clear majority. Based on the current predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.750527 | Disordered | 0.929726 | Binding | 0.349 | 0.902 | 0.750 | -4.653 | Likely Benign | 0.895 | Likely Pathogenic | Ambiguous | 0.160 | Likely Benign | -2.32 | Neutral | 0.552 | Possibly Damaging | 0.388 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 0.4207 | 0.7733 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2958G>C | E986D 2D AIThe SynGAP1 missense variant E986D is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs two benign votes) and Foldetta results are unavailable. Overall, the majority of conventional tools lean toward pathogenicity, but the single high‑accuracy benign prediction and the inconclusive consensus leave the variant’s impact uncertain. There is no contradiction with ClinVar status, as the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.750527 | Disordered | 0.929726 | Binding | 0.349 | 0.902 | 0.750 | -4.001 | Likely Benign | 0.628 | Likely Pathogenic | Likely Benign | 0.129 | Likely Benign | -1.22 | Neutral | 0.974 | Probably Damaging | 0.715 | Possibly Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.2081 | 0.5364 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2958G>T | E986D 2D AIThe SynGAP1 missense variant E986D is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs two benign votes) and Foldetta results are unavailable. Overall, the majority of conventional tools lean toward pathogenicity, but the single high‑accuracy benign prediction and the inconclusive consensus leave the variant’s impact uncertain. There is no contradiction with ClinVar status, as the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.750527 | Disordered | 0.929726 | Binding | 0.349 | 0.902 | 0.750 | -4.001 | Likely Benign | 0.628 | Likely Pathogenic | Likely Benign | 0.129 | Likely Benign | -1.22 | Neutral | 0.974 | Probably Damaging | 0.715 | Possibly Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.2081 | 0.5364 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2959G>A | D987N 2D AIThe SynGAP1 D987N missense variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.919118 | Binding | 0.299 | 0.903 | 0.750 | -5.035 | Likely Benign | 0.698 | Likely Pathogenic | Likely Benign | 0.126 | Likely Benign | -1.42 | Neutral | 0.943 | Possibly Damaging | 0.755 | Possibly Damaging | 2.41 | Pathogenic | 0.49 | Tolerated | 0.1331 | 0.7427 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2963T>A | L988H 2D AIThe SynGAP1 missense variant L988H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2). Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs four benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.827927 | Disordered | 0.918781 | Binding | 0.360 | 0.913 | 0.750 | -4.779 | Likely Benign | 0.733 | Likely Pathogenic | Likely Benign | 0.179 | Likely Benign | -2.70 | Deleterious | 0.998 | Probably Damaging | 0.947 | Probably Damaging | 2.62 | Benign | 0.00 | Affected | 0.1179 | 0.1414 | -2 | -3 | -7.0 | 23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.2963T>C | L988P 2D AIThe SynGAP1 missense variant L988P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.827927 | Disordered | 0.918781 | Binding | 0.360 | 0.913 | 0.750 | -3.848 | Likely Benign | 0.703 | Likely Pathogenic | Likely Benign | 0.216 | Likely Benign | -2.96 | Deleterious | 0.977 | Probably Damaging | 0.900 | Possibly Damaging | 2.62 | Benign | 0.00 | Affected | 0.3282 | 0.1698 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2966C>A | S989Y 2D AIThe SynGAP1 missense variant S989Y is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. High‑accuracy predictions therefore indicate a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus is benign, and Foldetta data are unavailable. Consequently, the variant is most likely benign based on the current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.795062 | Disordered | 0.908835 | Binding | 0.296 | 0.911 | 0.750 | -4.774 | Likely Benign | 0.471 | Ambiguous | Likely Benign | 0.103 | Likely Benign | -3.32 | Deleterious | 0.986 | Probably Damaging | 0.876 | Possibly Damaging | 2.60 | Benign | 0.00 | Affected | 0.0611 | 0.4815 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2966C>T | S989F 2D AIThe SynGAP1 missense variant S989F is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, and Foldetta results are unavailable. Overall, the majority of reliable predictors and the consensus high‑accuracy tools indicate a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.795062 | Disordered | 0.908835 | Binding | 0.296 | 0.911 | 0.750 | -4.115 | Likely Benign | 0.435 | Ambiguous | Likely Benign | 0.109 | Likely Benign | -3.35 | Deleterious | 0.986 | Probably Damaging | 0.876 | Possibly Damaging | 2.60 | Benign | 0.00 | Affected | 0.0598 | 0.5103 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.2975T>G | V992G 2D AIThe SynGAP1 missense variant V992G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). All available in silico predictors classify the variant as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity, so the benign group includes every listed predictor, while the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign; the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.754692 | Disordered | 0.921728 | Binding | 0.331 | 0.917 | 0.750 | -1.906 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.067 | Likely Benign | -0.28 | Neutral | 0.056 | Benign | 0.086 | Benign | 4.21 | Benign | 0.16 | Tolerated | 0.2114 | 0.2612 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3005A>T | H1002L 2D AIThe SynGAP1 H1002L missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and the absence of the variant in population databases, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.953758 | Binding | 0.285 | 0.900 | 0.500 | -6.448 | Likely Benign | 0.556 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -3.12 | Deleterious | 0.801 | Possibly Damaging | 0.602 | Possibly Damaging | 2.79 | Benign | 0.13 | Tolerated | 0.1296 | 0.5088 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3007A>C | S1003R 2D AIThe SynGAP1 missense variant S1003R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of evaluated tools (six pathogenic vs. three benign) indicate a pathogenic impact. This prediction aligns with the lack of ClinVar annotation and does not contradict any existing clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.947349 | Binding | 0.272 | 0.901 | 0.625 | -5.113 | Likely Benign | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.110 | Likely Benign | -1.88 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.48 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1151 | 0.3746 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3007A>G | S1003G 2D AIThe SynGAP1 missense variant S1003G is catalogued in gnomAD (ID 6‑33443559‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect for S1003G, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.947349 | Binding | 0.272 | 0.901 | 0.625 | 6-33443559-A-G | 1 | 6.20e-7 | -5.888 | Likely Benign | 0.542 | Ambiguous | Likely Benign | 0.088 | Likely Benign | -1.72 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 2.48 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2730 | 0.4789 | 0 | 1 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||
| c.3008G>A | S1003N 2D AIThe SynGAP1 missense variant S1003N is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, more tools (five) predict pathogenicity than benign (three), and the high‑accuracy methods do not overturn this trend. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.947349 | Binding | 0.272 | 0.901 | 0.625 | -5.175 | Likely Benign | 0.889 | Likely Pathogenic | Ambiguous | 0.122 | Likely Benign | -1.37 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.48 | Pathogenic | 0.00 | Affected | 0.1798 | 0.5029 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3008G>T | S1003I 2D AIThe SynGAP1 missense variant S1003I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy consensus methods give mixed results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic, two benign votes); and Foldetta (combining FoldX‑MD and Rosetta) has no available output. Based on the overall distribution of predictions, the variant is most likely pathogenic. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.947349 | Binding | 0.272 | 0.901 | 0.625 | -8.952 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.189 | Likely Benign | -2.31 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.1294 | 0.5735 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3009C>A | S1003R 2D AIThe SynGAP1 missense variant S1003R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of evaluated tools (six pathogenic vs. three benign) indicate a pathogenic impact. This prediction aligns with the lack of ClinVar annotation and does not contradict any existing clinical classification. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.947349 | Binding | 0.272 | 0.901 | 0.625 | -5.113 | Likely Benign | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.141 | Likely Benign | -1.88 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.48 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1151 | 0.3746 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3009C>G | S1003R 2D AIThe SynGAP1 missense variant S1003R (ClinVar ID 1798770.0) is listed as Uncertain in ClinVar and is not present in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and ESM1b, while pathogenic predictions are reported by polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessment shows AlphaMissense‑Optimized classifying the variant as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic), and Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a pathogenic effect, which contrasts with the ClinVar designation of Uncertain. Thus, the variant is most likely pathogenic, contradicting the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.947349 | Binding | 0.272 | 0.901 | 0.625 | Uncertain | 1 | -5.113 | Likely Benign | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.141 | Likely Benign | -1.88 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.48 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1151 | 0.3746 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.3011A>C | H1004P 2D AIThe SynGAP1 missense variant H1004P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and the high‑accuracy AlphaMissense‑Optimized model. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a benign majority vote. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction between the predictions and ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.808535 | Disordered | 0.943707 | Binding | 0.271 | 0.901 | 0.750 | -3.686 | Likely Benign | 0.460 | Ambiguous | Likely Benign | 0.236 | Likely Benign | -2.69 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.72 | Benign | 0.18 | Tolerated | 0.2076 | 0.4643 | 0 | -2 | 1.6 | -40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3011A>T | H1004L 2D AIThe SynGAP1 missense variant H1004L is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs two benign votes). Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the balance of evidence leans toward a benign interpretation, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.808535 | Disordered | 0.943707 | Binding | 0.271 | 0.901 | 0.750 | -5.179 | Likely Benign | 0.716 | Likely Pathogenic | Likely Benign | 0.220 | Likely Benign | -3.14 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.75 | Benign | 0.55 | Tolerated | 0.1223 | 0.6026 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3013A>T | S1005C 2D AIThe SynGAP1 missense variant S1005C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy consensus (SGM Consensus) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of standard predictors lean toward pathogenicity, while the single high‑accuracy tool that is available (AlphaMissense‑Optimized) predicts benign, and the consensus tool is inconclusive. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.812494 | Disordered | 0.936602 | Binding | 0.261 | 0.897 | 0.750 | -8.519 | Likely Pathogenic | 0.640 | Likely Pathogenic | Likely Benign | 0.173 | Likely Benign | -2.20 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.59 | Benign | 0.00 | Affected | 0.1246 | 0.4492 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.3019A>T | S1007C 2D AIThe SynGAP1 missense variant S1007C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. Two tools—ESM1b and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is uncertain, and Foldetta’s stability prediction is unavailable. Overall, the balance of evidence (four benign versus three pathogenic predictions, with high‑accuracy tools favoring benign) indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.671169 | Disordered | 0.925648 | Binding | 0.295 | 0.899 | 0.750 | -7.399 | In-Between | 0.487 | Ambiguous | Likely Benign | 0.132 | Likely Benign | -1.91 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.61 | Benign | 0.01 | Affected | 0.1405 | 0.5142 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.3020G>T | S1007I 2D AIThe SynGAP1 missense variant S1007I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default) predict a pathogenic impact. The remaining tools, ESM1b and AlphaMissense‑Optimized, return uncertain results. High‑accuracy assessments further support a deleterious interpretation: the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as pathogenic; AlphaMissense‑Optimized remains uncertain, and Foldetta data are unavailable. Overall, the preponderance of evidence from both conventional and high‑accuracy predictors indicates that the S1007I variant is most likely pathogenic, with no conflict with ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.671169 | Disordered | 0.925648 | Binding | 0.295 | 0.899 | 0.750 | -7.800 | In-Between | 0.920 | Likely Pathogenic | Ambiguous | 0.126 | Likely Benign | -2.55 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.65 | Benign | 0.00 | Affected | 0.1324 | 0.4769 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3022G>T | D1008Y 2D AIThe SynGAP1 missense variant D1008Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a tie and thus unavailable, and Foldetta results are not provided. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.919416 | Binding | 0.280 | 0.899 | 0.625 | -5.371 | Likely Benign | 0.928 | Likely Pathogenic | Ambiguous | 0.237 | Likely Benign | -3.71 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.62 | Benign | 0.00 | Affected | 0.1043 | 0.6293 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||||
| c.3023A>C | D1008A 2D AIThe SynGAP1 D1008A variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, more tools (five) predict pathogenicity than benign (three), and no ClinVar evidence contradicts this assessment. Thus, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.919416 | Binding | 0.280 | 0.899 | 0.625 | -3.210 | Likely Benign | 0.861 | Likely Pathogenic | Ambiguous | 0.209 | Likely Benign | -2.65 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.69 | Benign | 0.03 | Affected | 0.4014 | 0.6444 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.3023A>G | D1008G 2D AIThe SynGAP1 D1008G missense variant (ClinVar ID 2963386.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33443575‑A‑G). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the balance of evidence leans toward a pathogenic interpretation, which does not contradict the current ClinVar designation of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.919416 | Binding | 0.280 | 0.899 | 0.625 | Uncertain | 1 | 6-33443575-A-G | 1 | 6.20e-7 | -3.213 | Likely Benign | 0.742 | Likely Pathogenic | Likely Benign | 0.203 | Likely Benign | -2.84 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.65 | Benign | 0.01 | Affected | 3.77 | 5 | 0.3660 | 0.6073 | -1 | 1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||
| c.3023A>T | D1008V 2D AIThe SynGAP1 D1008V variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, more tools (five) predict pathogenicity than benign (three), and the high‑accuracy assessments do not overturn this trend. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.919416 | Binding | 0.280 | 0.899 | 0.625 | -4.828 | Likely Benign | 0.944 | Likely Pathogenic | Ambiguous | 0.242 | Likely Benign | -3.61 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.64 | Benign | 0.01 | Affected | 0.1447 | 0.6608 | -2 | -3 | 7.7 | -15.96 | ||||||||||||||||||||||||||||||||||||||||
| c.3025G>A | E1009K 2D AIThe SynGAP1 missense variant E1009K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of consensus tools (five pathogenic vs. three benign) indicate a pathogenic effect. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.728858 | Disordered | 0.914552 | Binding | 0.325 | 0.885 | 0.500 | -3.419 | Likely Benign | 0.897 | Likely Pathogenic | Ambiguous | 0.061 | Likely Benign | -1.90 | Neutral | 0.961 | Probably Damaging | 0.630 | Possibly Damaging | 2.41 | Pathogenic | 0.01 | Affected | 0.2511 | 0.7625 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
Found 8751 rows. Show 800 rows per page. Page 1/11 | Next »