
Table of SynGAP1 Isoform α2 (UniProt Q96PV0-1) Missense Variants.
| c.dna | Variant | SGM Consensus | Domain and Structure information: based on WT protein | Annotated databases | Deep learning-based pathogenicity predictions | Folding stability-based pathogenicity predictions | Sequence/structure-based pathogenicity predictions | Phase Separation | Evolutionary/physical properties | Molecular Dynamics-based analysis | DOI | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Domain | IUPred2 | ANCHOR2 | AlphaFold | MobiDB | PhosphoSitePlus | ClinVar | gnomAD | ESM1b | AlphaMissense | FoldX | Rosetta | Foldetta | PremPS | REVEL | PROVEAN | PolyPhen-2 HumDiv | PolyPhen-2 HumVar | FATHMM | SIFT | PSMutPred | PAM | Physical | SASA | Normalized B-factor backbone | Normalized B-factor sidechain | SynGAP Structural Annotation | |||||||||||||||||||||||||||||||||||||||||||||
| Score | Prediction | Score | Prediction | pLDDT | disorder | disorder | LTP | HTP | KL | PTM | Clinical Status | Review | Subm. | ID | Allele count | Allele freq. | LLR score | Prediction | Pathogenicity | Class | Optimized | Average ΔΔG | Prediction | StdDev | ΔΔG | Prediction | ΔΔG | Prediction | ΔΔG | Prediction | Score | Prediction | Score | Prediction | pph2_prob | Prediction | pph2_prob | Prediction | Nervous System Score | Prediction | Prediction | Status | Conservation | Sequences | IP RF | SP RF | Prediction | PAM250 | PAM120 | Hydropathy Δ | MW Δ | Average | Δ | Δ | StdDev | Δ | StdDev | Secondary | Tertiary bonds | Inside out | GAP-Ras interface | At membrane | No effect | MD Alert | Verdict | Description | |||||
| c.3697A>C | I1233L 2D  AIThe SynGAP1 missense variant I1233L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the substitution as benign, and AlphaMissense‑Optimized also predicts a benign outcome. No tool predicts pathogenicity; AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also indicates likely benign; Foldetta data are not available. Overall, the consensus of available predictions points to a benign effect, and this is consistent with the lack of ClinVar evidence or gnomAD observation. Thus, the variant is most likely benign, and this conclusion does not contradict ClinVar status, which has no entry for it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -3.031 | Likely Benign | 0.382 | Ambiguous | Likely Benign | 0.113 | Likely Benign | -0.01 | Neutral | 0.211 | Benign | 0.108 | Benign | 2.95 | Benign | 1.00 | Tolerated | 0.0631 | 0.3246 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||
| c.40C>A | P14T 2D  AIThe SynGAP1 missense variant P14T is reported in gnomAD (ID 6‑33420304‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also reports likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for P14T, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | 6-33420304-C-A | -3.261 | Likely Benign | 0.150 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -0.27 | Neutral | 0.212 | Benign | 0.014 | Benign | 4.20 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1912 | 0.6658 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.40C>T | P14S 2D  AIThe SynGAP1 missense variant P14S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | -2.352 | Likely Benign | 0.123 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -0.02 | Neutral | 0.212 | Benign | 0.014 | Benign | 4.23 | Benign | 0.00 | Affected | 0.3935 | 0.5938 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.49T>C | S17P 2D  AIThe SynGAP1 missense variant S17P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a benign effect. AlphaMissense‑Optimized independently scores the variant as benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | -3.251 | Likely Benign | 0.256 | Likely Benign | Likely Benign | 0.210 | Likely Benign | -0.65 | Neutral | 0.212 | Benign | 0.010 | Benign | 4.01 | Benign | 0.00 | Affected | 0.2406 | 0.6473 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.5G>T | S2I 2D  AIThe SynGAP1 missense variant S2I is catalogued in gnomAD (6‑33420269‑G‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all report benign or likely benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments confirm the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus, derived from the majority of the high‑confidence predictors, is benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that S2I is most likely benign, and this assessment does not contradict any ClinVar status, as none is reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.543646 | Binding | 0.382 | 0.922 | 0.750 | 6-33420269-G-T | -4.947 | Likely Benign | 0.439 | Ambiguous | Likely Benign | 0.031 | Likely Benign | -0.59 | Neutral | 0.212 | Benign | 0.020 | Benign | 4.04 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1021 | 0.5980 | -2 | -1 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||
| c.1111A>G | S371G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S371G is not reported in ClinVar and is absent from gnomAD. Across the available in‑silico predictors, every tool examined—REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized—classifies the substitution as benign. No pathogenic predictions are present. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also reports a benign effect. Consequently, the variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.444081 | Structured | 0.432086 | Uncertain | 0.294 | 0.746 | 0.375 | -2.073 | Likely Benign | 0.053 | Likely Benign | Likely Benign | 0.47 | Likely Benign | 0.3 | 0.49 | Likely Benign | 0.48 | Likely Benign | 0.35 | Likely Benign | 0.164 | Likely Benign | -1.32 | Neutral | 0.213 | Benign | 0.067 | Benign | 4.63 | Benign | 0.17 | Tolerated | 0.3192 | 0.5295 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||
| c.1112G>C | S371T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S371T is not reported in ClinVar and is absent from gnomAD. All available in silico predictors classify the substitution as benign: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). No tool predicts pathogenicity. High‑accuracy assessments concur: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields Likely Benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also reports a benign effect. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.444081 | Structured | 0.432086 | Uncertain | 0.294 | 0.746 | 0.375 | -4.512 | Likely Benign | 0.072 | Likely Benign | Likely Benign | 0.38 | Likely Benign | 0.1 | -0.27 | Likely Benign | 0.06 | Likely Benign | 0.05 | Likely Benign | 0.173 | Likely Benign | -0.65 | Neutral | 0.213 | Benign | 0.067 | Benign | 4.64 | Benign | 0.22 | Tolerated | 0.2245 | 0.6462 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||
| c.362C>T | A121V 2D  AIThe SynGAP1 missense variant A121V is reported in gnomAD (ID 6‑33432227‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence supports a benign classification for A121V, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.779859 | Disordered | 0.661304 | Binding | 0.362 | 0.888 | 0.750 | 6-33432227-C-T | 3 | 1.86e-6 | -3.818 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.046 | Likely Benign | -1.44 | Neutral | 0.213 | Benign | 0.050 | Benign | 4.04 | Benign | 0.02 | Affected | 3.61 | 5 | 0.1450 | 0.6898 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||
| c.520A>G | M174V 2D  AIThe SynGAP1 missense variant M174V is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (gnomAD ID: 6‑33435162‑A‑G). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM; pathogenic predictions come from SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and no Foldetta stability data are available. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a 2‑vs‑2 split and is therefore inconclusive. With five benign versus three pathogenic calls and no contradictory ClinVar evidence, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.661982 | Disordered | 0.485854 | Uncertain | 0.373 | 0.620 | 0.375 | 6-33435162-A-G | 2 | 1.24e-6 | -8.604 | Likely Pathogenic | 0.897 | Likely Pathogenic | Ambiguous | 0.108 | Likely Benign | -1.76 | Neutral | 0.213 | Benign | 0.067 | Benign | 4.12 | Benign | 0.04 | Affected | 3.61 | 5 | 0.2866 | 0.3841 | 1 | 2 | 2.3 | -32.06 | |||||||||||||||||||||||||||||||||||
| c.522G>A | M174I 2D  AIThe SynGAP1 missense variant M174I is listed in gnomAD (ID 6‑33435164‑G‑A) but has no ClinVar record. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; pathogenic predictions come from ESM1b and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, whereas the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2) and Foldetta results are unavailable. Overall, six tools predict benign while only two predict pathogenic, and the only high‑accuracy pathogenic call is from AlphaMissense‑Optimized. Thus, the variant is most likely benign based on the preponderance of evidence, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.661982 | Disordered | 0.485854 | Uncertain | 0.373 | 0.620 | 0.375 | 6-33435164-G-A | 4 | 2.48e-6 | -8.732 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.120 | Likely Benign | -1.63 | Neutral | 0.213 | Benign | 0.067 | Benign | 4.10 | Benign | 0.07 | Tolerated | 3.61 | 5 | 0.1191 | 0.3469 | 1 | 2 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||
| c.522G>C | M174I 2D AIThe SynGAP1 missense variant M174I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. High‑accuracy methods give mixed results: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs 2 benign), and Foldetta’s stability assessment is unavailable. Overall, the majority of standard predictors favor a benign outcome, and the high‑accuracy predictions do not override this trend. Thus, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar reporting. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.661982 | Disordered | 0.485854 | Uncertain | 0.373 | 0.620 | 0.375 | -8.732 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.120 | Likely Benign | -1.63 | Neutral | 0.213 | Benign | 0.067 | Benign | 4.10 | Benign | 0.07 | Tolerated | 3.61 | 5 | 0.1191 | 0.3469 | 1 | 2 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||
| c.522G>T | M174I 2D AIThe SynGAP1 missense variant M174I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. High‑accuracy methods give mixed results: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta data are unavailable. Overall, the majority of standard predictors indicate a benign impact, while the single high‑accuracy tool suggests pathogenicity but is not supported by consensus or folding‑stability evidence. Thus, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.661982 | Disordered | 0.485854 | Uncertain | 0.373 | 0.620 | 0.375 | -8.732 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.120 | Likely Benign | -1.63 | Neutral | 0.213 | Benign | 0.067 | Benign | 4.10 | Benign | 0.07 | Tolerated | 3.61 | 5 | 0.1191 | 0.3469 | 1 | 2 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||
| c.1895A>C | N632T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N632T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, PROVEAN, and FATHMM. The remaining tools—FoldX, Rosetta, and AlphaMissense‑Default—return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of predictions lean toward a benign impact, and this does not contradict any ClinVar annotation (none is available). Thus, based on the current computational evidence, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.041437 | Uncertain | 0.938 | 0.254 | 0.000 | -6.797 | Likely Benign | 0.371 | Ambiguous | Likely Benign | 1.14 | Ambiguous | 0.2 | -0.58 | Ambiguous | 0.28 | Likely Benign | 0.39 | Likely Benign | 0.541 | Likely Pathogenic | -4.48 | Deleterious | 0.214 | Benign | 0.062 | Benign | -1.49 | Pathogenic | 0.13 | Tolerated | 0.1295 | 0.6809 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||
| c.2487G>C | E829D 2D  AIThe SynGAP1 missense variant E829D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence from multiple independent predictors indicates that E829D is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.626045 | Binding | 0.326 | 0.882 | 0.375 | -4.015 | Likely Benign | 0.091 | Likely Benign | Likely Benign | 0.066 | Likely Benign | -1.89 | Neutral | 0.217 | Benign | 0.121 | Benign | 2.40 | Pathogenic | 0.00 | Affected | 0.1939 | 0.4964 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2487G>T | E829D 2D AIThe SynGAP1 missense variant E829D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence from multiple independent predictors indicates that E829D is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.626045 | Binding | 0.326 | 0.882 | 0.375 | -4.015 | Likely Benign | 0.091 | Likely Benign | Likely Benign | 0.066 | Likely Benign | -1.89 | Neutral | 0.217 | Benign | 0.121 | Benign | 2.40 | Pathogenic | 0.00 | Affected | 0.1939 | 0.4964 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2596G>C | V866L 2D  AIThe SynGAP1 missense variant V866L is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification—there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.599170 | Disordered | 0.638070 | Binding | 0.266 | 0.788 | 0.250 | -3.352 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.046 | Likely Benign | -0.97 | Neutral | 0.217 | Benign | 0.229 | Benign | 2.71 | Benign | 0.21 | Tolerated | 3.82 | 4 | 0.0836 | 0.4660 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||||||||||
| c.2596G>T | V866L 2D AIThe SynGAP1 missense variant V866L is listed in ClinVar (ID 469150.0) with an “Uncertain” clinical significance and is present in gnomAD (6‑33443148‑G‑T). All evaluated in‑silico predictors classify the substitution as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool reports a pathogenic outcome. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant. Overall, the computational evidence strongly supports a benign effect, and this conclusion does not contradict the current ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.599170 | Disordered | 0.638070 | Binding | 0.266 | 0.788 | 0.250 | Uncertain | 1 | 6-33443148-G-T | 1 | 6.20e-7 | -3.352 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.046 | Likely Benign | -0.97 | Neutral | 0.217 | Benign | 0.229 | Benign | 2.71 | Benign | 0.21 | Tolerated | 3.82 | 4 | 0.0836 | 0.4660 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||
| c.2734A>C | T912P 2D  AIThe SynGAP1 missense variant T912P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.740671 | Binding | 0.285 | 0.909 | 0.250 | -2.955 | Likely Benign | 0.111 | Likely Benign | Likely Benign | 0.087 | Likely Benign | -0.78 | Neutral | 0.217 | Benign | 0.121 | Benign | 4.00 | Benign | 0.00 | Affected | 0.1832 | 0.4608 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3600G>C | E1200D 2D AIThe SynGAP1 missense variant E1200D is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the preponderance of evidence supports a benign classification for E1200D, and this conclusion is consistent with the lack of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.545602 | Disordered | 0.458056 | Uncertain | 0.889 | 0.596 | 0.250 | -4.731 | Likely Benign | 0.172 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -1.88 | Neutral | 0.217 | Benign | 0.121 | Benign | 2.68 | Benign | 0.04 | Affected | 0.1495 | 0.2475 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3600G>T | E1200D 2D AIThe SynGAP1 missense variant E1200D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is also benign. Foldetta results are not available, so they do not influence the assessment. Overall, the preponderance of evidence indicates that E1200D is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.545602 | Disordered | 0.458056 | Uncertain | 0.889 | 0.596 | 0.250 | -4.731 | Likely Benign | 0.172 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -1.88 | Neutral | 0.217 | Benign | 0.121 | Benign | 2.68 | Benign | 0.04 | Affected | 0.1495 | 0.2475 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.106C>T | H36Y 2D  AIThe SynGAP1 missense variant H36Y is listed in ClinVar with an uncertain significance (ClinVar ID 2089635.0) and is present in the gnomAD database (gnomAD ID 6‑33423515‑C‑T). Functional prediction tools largely agree that the substitution is benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report a benign effect. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled Likely Benign. No Foldetta stability prediction is available. Overall, the computational evidence overwhelmingly supports a benign classification, which is consistent with the ClinVar designation of uncertain significance rather than a pathogenic claim. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.433974 | Uncertain | 0.334 | 0.834 | 0.375 | Uncertain | 1 | 6-33423515-C-T | 2 | 1.24e-6 | -3.461 | Likely Benign | 0.139 | Likely Benign | Likely Benign | 0.023 | Likely Benign | -1.03 | Neutral | 0.219 | Benign | 0.066 | Benign | 4.16 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1265 | 0.5024 | 0 | 2 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||||
| c.283C>A | H95N 2D  AIThe SynGAP1 missense variant H95N is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) indicates likely benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the computational evidence overwhelmingly supports a benign classification for H95N, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.590542 | Binding | 0.335 | 0.875 | 0.625 | -3.454 | Likely Benign | 0.069 | Likely Benign | Likely Benign | 0.089 | Likely Benign | -1.12 | Neutral | 0.219 | Benign | 0.009 | Benign | 4.20 | Benign | 0.00 | Affected | 0.1821 | 0.2491 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.283C>T | H95Y 2D AIThe SynGAP1 missense variant H95Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta results are unavailable. Overall, the majority of evidence indicates that H95Y is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.590542 | Binding | 0.335 | 0.875 | 0.625 | -3.610 | Likely Benign | 0.112 | Likely Benign | Likely Benign | 0.090 | Likely Benign | -1.45 | Neutral | 0.219 | Benign | 0.014 | Benign | 4.14 | Benign | 0.00 | Affected | 0.0880 | 0.4122 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||||||
| c.3746G>A | R1249K 2D 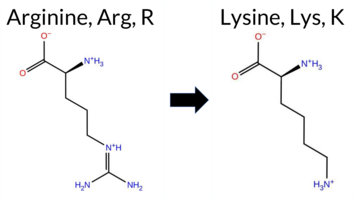 AIThe SynGAP1 missense variant R1249K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, SIFT and FATHMM predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; no Foldetta stability data are available, so it does not influence the overall assessment. Overall, the preponderance of evidence points to a benign effect for R1249K, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -5.782 | Likely Benign | 0.205 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -1.99 | Neutral | 0.219 | Benign | 0.191 | Benign | 1.80 | Pathogenic | 0.00 | Affected | 0.3343 | 0.2413 | 3 | 2 | 0.6 | -28.01 | ||||||||||||||||||||||||||||||||||||||
| c.2198A>C | Q733P 2D 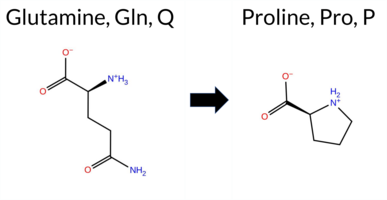 AIThe SynGAP1 missense variant Q733P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.874069 | Disordered | 0.410831 | Uncertain | 0.331 | 0.686 | 0.875 | -4.249 | Likely Benign | 0.075 | Likely Benign | Likely Benign | 0.172 | Likely Benign | -1.91 | Neutral | 0.220 | Benign | 0.308 | Benign | 2.52 | Benign | 0.04 | Affected | 0.2182 | 0.3894 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||||||||||||||
| c.2318T>G | M773R 2D 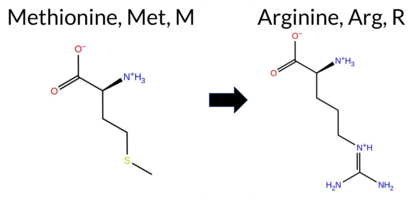 AIThe SynGAP1 missense variant M773R is listed in gnomAD (ID 6‑33442476‑T‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign. No Foldetta (FoldX‑MD/Rosetta stability) result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.408655 | Structured | 0.916222 | Binding | 0.325 | 0.893 | 0.250 | 6-33442476-T-G | 1 | 1.28e-6 | -4.340 | Likely Benign | 0.597 | Likely Pathogenic | Likely Benign | 0.183 | Likely Benign | -1.87 | Neutral | 0.220 | Benign | 0.471 | Possibly Damaging | 4.18 | Benign | 0.02 | Affected | 3.64 | 6 | 0.1778 | 0.0800 | -1 | 0 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||||||
| c.2695A>C | I899L 2D AIThe SynGAP1 missense variant I899L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.443727 | Uncertain | 0.292 | 0.928 | 0.375 | -1.137 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.055 | Likely Benign | -0.36 | Neutral | 0.220 | Benign | 0.078 | Benign | 2.74 | Benign | 0.04 | Affected | 0.0863 | 0.2840 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.2695A>G | I899V 2D 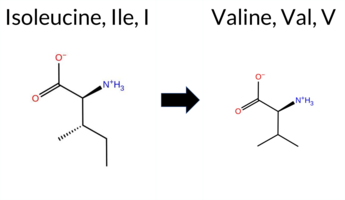 AIThe SynGAP1 missense variant I899V is listed in ClinVar as a benign alteration (ClinVar ID 1003653.0) and is present in the gnomAD database (gnomAD ID 6‑33443247‑A‑G). All evaluated in‑silico predictors classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity, so the pathogenic‑prediction group is empty. High‑accuracy assessments further support a benign effect: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant. Overall, the computational evidence strongly suggests the variant is benign, consistent with its ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.443727 | Uncertain | 0.292 | 0.928 | 0.375 | Benign | 1 | 6-33443247-A-G | 6 | 3.72e-6 | -2.569 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.040 | Likely Benign | 0.09 | Neutral | 0.220 | Benign | 0.078 | Benign | 2.75 | Benign | 0.92 | Tolerated | 4.32 | 4 | 0.1169 | 0.2864 | 4 | 3 | -0.3 | -14.03 | ||||||||||||||||||||||||||||||||
| c.3097T>G | S1033A 2D  AIThe SynGAP1 missense variant S1033A is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are unavailable. Overall, the consensus of all available predictions indicates that the variant is most likely benign, and this conclusion is consistent with the lack of a ClinVar classification, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.993473 | Binding | 0.294 | 0.737 | 0.625 | -3.107 | Likely Benign | 0.121 | Likely Benign | Likely Benign | 0.033 | Likely Benign | 0.06 | Neutral | 0.220 | Benign | 0.085 | Benign | 2.81 | Benign | 1.00 | Tolerated | 0.4387 | 0.5282 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2186A>G | N729S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N729S is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Across the available in‑silico predictors, the majority (REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) classify the change as benign, while Rosetta and Foldetta return uncertain results. No tool predicts pathogenicity. High‑accuracy assessments reinforce this benign trend: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign,” and Foldetta’s stability prediction is inconclusive. Overall, the computational evidence strongly supports a benign effect, and this does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.750527 | Disordered | 0.426547 | Uncertain | 0.651 | 0.583 | 0.625 | Conflicting | 2 | -1.578 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 0.14 | Likely Benign | 0.1 | 1.34 | Ambiguous | 0.74 | Ambiguous | -0.36 | Likely Benign | 0.063 | Likely Benign | -0.42 | Neutral | 0.221 | Benign | 0.027 | Benign | 3.38 | Benign | 0.93 | Tolerated | 3.59 | 7 | 0.3411 | 0.4854 | 1 | 1 | 2.7 | -27.03 | ||||||||||||||||||||||||||
| c.2351C>G | A784G 2D  AIThe SynGAP1 missense variant A784G is reported in gnomAD (ID 6‑33442903‑C‑G) and has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are unavailable. Overall, the evidence strongly suggests the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.801317 | Disordered | 0.708872 | Binding | 0.314 | 0.896 | 0.625 | 6-33442903-C-G | 1 | 6.20e-7 | -3.370 | Likely Benign | 0.158 | Likely Benign | Likely Benign | 0.056 | Likely Benign | -1.24 | Neutral | 0.224 | Benign | 0.138 | Benign | 2.67 | Benign | 0.26 | Tolerated | 3.64 | 6 | 0.2303 | 0.4418 | 0 | 1 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||||||
| c.2501T>C | M834T 2D  AIThe SynGAP1 missense variant M834T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for M834T, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.585406 | Disordered | 0.640801 | Binding | 0.258 | 0.863 | 0.375 | -1.525 | Likely Benign | 0.332 | Likely Benign | Likely Benign | 0.137 | Likely Benign | -2.03 | Neutral | 0.224 | Benign | 0.085 | Benign | 2.45 | Pathogenic | 0.00 | Affected | 0.1757 | 0.1799 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.2590G>A | A864T 2D  AIThe SynGAP1 missense variant A864T is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.549308 | Disordered | 0.611966 | Binding | 0.269 | 0.788 | 0.250 | -4.314 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.016 | Likely Benign | -1.00 | Neutral | 0.224 | Benign | 0.113 | Benign | 2.55 | Benign | 0.10 | Tolerated | 0.1504 | 0.7485 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2645G>T | G882V 2D  AIThe SynGAP1 missense variant G882V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation—there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.690604 | Disordered | 0.632888 | Binding | 0.306 | 0.878 | 0.250 | -5.893 | Likely Benign | 0.257 | Likely Benign | Likely Benign | 0.115 | Likely Benign | -2.41 | Neutral | 0.224 | Benign | 0.066 | Benign | 2.06 | Pathogenic | 0.02 | Affected | 0.1178 | 0.4717 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.2656G>T | A886S 2D  AIThe SynGAP1 missense variant A886S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.716283 | Disordered | 0.619166 | Binding | 0.359 | 0.922 | 0.500 | -2.366 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.055 | Likely Benign | -0.50 | Neutral | 0.224 | Benign | 0.185 | Benign | 2.24 | Pathogenic | 0.00 | Affected | 0.2755 | 0.5639 | 1 | 1 | -2.6 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2698A>T | T900S 2D  AIThe SynGAP1 missense variant T900S is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.465347 | Uncertain | 0.289 | 0.924 | 0.375 | -2.031 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.056 | Likely Benign | -0.17 | Neutral | 0.224 | Benign | 0.171 | Benign | 2.70 | Benign | 0.71 | Tolerated | 0.3311 | 0.4301 | 1 | 1 | -0.1 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2824C>A | P942T 2D AIThe SynGAP1 missense variant P942T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for P942T, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.878102 | Binding | 0.365 | 0.915 | 0.625 | -5.745 | Likely Benign | 0.067 | Likely Benign | Likely Benign | 0.027 | Likely Benign | -1.23 | Neutral | 0.224 | Benign | 0.096 | Benign | 2.49 | Pathogenic | 0.00 | Affected | 0.1683 | 0.5599 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2828G>A | G943D 2D  AIThe SynGAP1 missense variant G943D is not reported in ClinVar and is absent from gnomAD. In silico predictors that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; ESM1b is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Taken together, the overwhelming majority of predictions indicate a benign effect, and this conclusion does not contradict any ClinVar status (none is assigned). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.973328 | Disordered | 0.860437 | Binding | 0.372 | 0.910 | 0.750 | -7.951 | In-Between | 0.304 | Likely Benign | Likely Benign | 0.296 | Likely Benign | -0.28 | Neutral | 0.224 | Benign | 0.113 | Benign | 2.85 | Benign | 0.11 | Tolerated | 0.1904 | 0.2826 | 1 | -1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||||||
| c.2828G>T | G943V 2D AIThe SynGAP1 missense variant G943V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. The only tool with an uncertain outcome is ESM1b, which does not alter the overall consensus. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign classification. High‑accuracy predictors corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are not available. Taken together, the evidence overwhelmingly supports a benign interpretation, and there is no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.973328 | Disordered | 0.860437 | Binding | 0.372 | 0.910 | 0.750 | -7.658 | In-Between | 0.092 | Likely Benign | Likely Benign | 0.265 | Likely Benign | -0.43 | Neutral | 0.224 | Benign | 0.062 | Benign | 2.85 | Benign | 0.16 | Tolerated | 0.1399 | 0.3507 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.2831G>A | G944D 2D AIThe SynGAP1 missense variant G944D is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools largely support a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the majority‑vote SGM‑Consensus also indicates a likely benign outcome. Only SIFT predicts a pathogenic effect, and ESM1b remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a likely benign classification. Foldetta stability analysis is not available for this residue. Overall, the preponderance of evidence points to a benign impact, and this assessment does not conflict with the absence of a ClinVar claim. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.977651 | Disordered | 0.852408 | Binding | 0.360 | 0.923 | 0.750 | -7.684 | In-Between | 0.296 | Likely Benign | Likely Benign | 0.421 | Likely Benign | -1.42 | Neutral | 0.224 | Benign | 0.131 | Benign | 3.70 | Benign | 0.00 | Affected | 0.1833 | 0.2826 | 1 | -1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||||||
| c.2840G>C | G947A 2D  AIThe SynGAP1 missense variant G947A is listed in ClinVar (ID 1595137) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443392‑G‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool in the dataset predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Overall, the computational evidence strongly favors a benign impact, which does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988695 | Disordered | 0.850554 | Binding | 0.358 | 0.919 | 0.750 | Conflicting | 3 | 6-33443392-G-C | 28 | 1.73e-5 | -6.511 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.156 | Likely Benign | -0.41 | Neutral | 0.224 | Benign | 0.131 | Benign | 4.97 | Benign | 0.10 | Tolerated | 4.32 | 4 | 0.3375 | 0.4957 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||||
| c.2888A>T | H963L 2D  AIThe SynGAP1 missense variant H963L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only ESM1b predicts a pathogenic outcome, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized indicates benign, and the SGM‑Consensus likewise suggests a benign effect; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact for H963L, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.983973 | Binding | 0.325 | 0.886 | 0.750 | -8.110 | Likely Pathogenic | 0.109 | Likely Benign | Likely Benign | 0.172 | Likely Benign | -1.58 | Neutral | 0.224 | Benign | 0.091 | Benign | 4.13 | Benign | 0.43 | Tolerated | 0.1626 | 0.5680 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||||||||||||
| c.2918G>T | G973V 2D AIThe SynGAP1 missense variant G973V is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. Grouping by consensus, the benign‑predicting tools outnumber the single pathogenic prediction. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign classification. Foldetta results are unavailable, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.959498 | Binding | 0.390 | 0.897 | 0.625 | -4.688 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.092 | Likely Benign | -0.76 | Neutral | 0.224 | Benign | 0.091 | Benign | 4.18 | Benign | 0.01 | Affected | 0.1392 | 0.3684 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.2978C>T | P993L 2D  AIThe SynGAP1 missense variant P993L is reported in ClinVar as “Not listed” and is not present in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which contains no pathogenic assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.923979 | Binding | 0.319 | 0.908 | 0.750 | -3.581 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.028 | Likely Benign | -1.37 | Neutral | 0.224 | Benign | 0.138 | Benign | 4.14 | Benign | 0.01 | Affected | 0.2299 | 0.6697 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.2983C>A | P995T 2D AIThe SynGAP1 missense variant P995T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect for P995T, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.935305 | Binding | 0.338 | 0.902 | 0.750 | -5.148 | Likely Benign | 0.058 | Likely Benign | Likely Benign | 0.034 | Likely Benign | -1.15 | Neutral | 0.224 | Benign | 0.096 | Benign | 4.19 | Benign | 0.00 | Affected | 0.1467 | 0.5667 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2989G>A | A997T 2D AIThe SynGAP1 missense variant A997T is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (derived from the four high‑accuracy predictors) is benign; Foldetta results are unavailable. Taken together, the majority of computational evidence indicates a benign effect, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.948624 | Binding | 0.273 | 0.901 | 0.500 | Uncertain | 1 | -4.102 | Likely Benign | 0.071 | Likely Benign | Likely Benign | 0.085 | Likely Benign | -0.62 | Neutral | 0.224 | Benign | 0.120 | Benign | 4.17 | Benign | 0.00 | Affected | 4.32 | 4 | 0.1582 | 0.7182 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||
| c.2989G>T | A997S 2D AIThe SynGAP1 missense variant A997S is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic call comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.948624 | Binding | 0.273 | 0.901 | 0.500 | -3.362 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -0.63 | Neutral | 0.224 | Benign | 0.066 | Benign | 4.18 | Benign | 0.00 | Affected | 0.2661 | 0.5694 | 1 | 1 | -2.6 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.3034C>T | P1012S 2D AIThe SynGAP1 missense variant P1012S is catalogued in gnomAD (ID 6‑33443586‑C‑T) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Overall, the evidence strongly supports a benign classification, and this conclusion is consistent with the absence of a ClinVar pathogenic designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.894674 | Binding | 0.319 | 0.866 | 0.625 | 6-33443586-C-T | 2 | 1.24e-6 | -3.342 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.23 | Neutral | 0.224 | Benign | 0.131 | Benign | 2.81 | Benign | 0.22 | Tolerated | 3.77 | 5 | 0.3217 | 0.5300 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3035C>T | P1012L 2D AIThe SynGAP1 missense variant P1012L is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which contains no pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.894674 | Binding | 0.319 | 0.866 | 0.625 | -4.363 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.014 | Likely Benign | -0.90 | Neutral | 0.224 | Benign | 0.131 | Benign | 2.76 | Benign | 0.12 | Tolerated | 0.2155 | 0.6388 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.3089A>G | H1030R 2D  AIThe SynGAP1 missense variant H1030R is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. The high‑accuracy consensus methods likewise support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.849326 | Disordered | 0.995856 | Binding | 0.375 | 0.735 | 0.500 | -2.312 | Likely Benign | 0.340 | Likely Benign | Likely Benign | 0.031 | Likely Benign | -1.08 | Neutral | 0.224 | Benign | 0.066 | Benign | 2.85 | Benign | 0.06 | Tolerated | 0.1872 | 0.2955 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.3089A>T | H1030L 2D AIThe SynGAP1 missense variant H1030L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign effect. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that H1030L is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.849326 | Disordered | 0.995856 | Binding | 0.375 | 0.735 | 0.500 | -3.513 | Likely Benign | 0.270 | Likely Benign | Likely Benign | 0.108 | Likely Benign | -2.35 | Neutral | 0.224 | Benign | 0.120 | Benign | 2.76 | Benign | 0.02 | Affected | 0.0968 | 0.5710 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||||||||||||
| c.3124C>A | Q1042K 2D  AIThe SynGAP1 missense variant Q1042K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective predictions strongly suggest that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.959333 | Binding | 0.310 | 0.846 | 0.625 | -4.331 | Likely Benign | 0.498 | Ambiguous | Likely Benign | 0.304 | Likely Benign | -1.52 | Neutral | 0.224 | Benign | 0.091 | Benign | 5.44 | Benign | 0.13 | Tolerated | 0.2338 | 0.5202 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.3124C>G | Q1042E 2D  AIThe SynGAP1 missense variant Q1042E is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.959333 | Binding | 0.310 | 0.846 | 0.625 | -4.231 | Likely Benign | 0.262 | Likely Benign | Likely Benign | 0.267 | Likely Benign | -1.12 | Neutral | 0.224 | Benign | 0.077 | Benign | 5.44 | Benign | 0.15 | Tolerated | 0.2042 | 0.3276 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3134C>G | A1045G 2D AIThe SynGAP1 missense variant A1045G is reported in ClinVar as Benign (ClinVar ID 416778.0) and is present in the gnomAD database (gnomAD ID 6‑33443686‑C‑G). Prediction tools that assess pathogenicity all converge on a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. No tool in the dataset predicts pathogenicity. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Overall, the variant is most likely benign, and this prediction aligns with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.975609 | Disordered | 0.948874 | Binding | 0.352 | 0.882 | 0.750 | Benign/Likely benign | 7 | 6-33443686-C-G | 1407 | 8.72e-4 | -3.246 | Likely Benign | 0.075 | Likely Benign | Likely Benign | 0.024 | Likely Benign | -1.21 | Neutral | 0.224 | Benign | 0.066 | Benign | 2.64 | Benign | 0.33 | Tolerated | 3.77 | 5 | 0.2246 | 0.4798 | 1 | 0 | -2.2 | -14.03 | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||||||||||||||||
| c.3182G>A | G1061D 2D AIThe SynGAP1 missense variant G1061D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign prediction (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta results are unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.978672 | Disordered | 0.926729 | Binding | 0.394 | 0.923 | 0.875 | -9.481 | Likely Pathogenic | 0.346 | Ambiguous | Likely Benign | 0.375 | Likely Benign | -1.32 | Neutral | 0.224 | Benign | 0.120 | Benign | 4.01 | Benign | 0.00 | Affected | 0.1671 | 0.2024 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3182G>T | G1061V 2D AIThe SynGAP1 missense variant G1061V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta results are unavailable. Overall, the majority of evidence indicates that G1061V is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978672 | Disordered | 0.926729 | Binding | 0.394 | 0.923 | 0.875 | -6.709 | Likely Benign | 0.106 | Likely Benign | Likely Benign | 0.307 | Likely Benign | -1.41 | Neutral | 0.224 | Benign | 0.066 | Benign | 3.98 | Benign | 0.00 | Affected | 0.1431 | 0.3684 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3190C>A | Q1064K 2D AIThe SynGAP1 missense variant Q1064K is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the collective predictions strongly support a benign classification, and this conclusion is consistent with the lack of any ClinVar pathogenic annotation, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.953106 | Binding | 0.378 | 0.914 | 0.875 | -3.592 | Likely Benign | 0.234 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.32 | Neutral | 0.224 | Benign | 0.120 | Benign | 4.23 | Benign | 0.24 | Tolerated | 0.2758 | 0.4192 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.3191A>T | Q1064L 2D  AIThe SynGAP1 missense variant Q1064L is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the consensus of all available predictions strongly supports a benign classification, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.953106 | Binding | 0.378 | 0.914 | 0.875 | -3.492 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.133 | Likely Benign | -1.16 | Neutral | 0.224 | Benign | 0.091 | Benign | 4.20 | Benign | 0.13 | Tolerated | 0.1817 | 0.5485 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3244C>A | Q1082K 2D AIThe SynGAP1 missense variant Q1082K is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score benign, and AlphaMissense‑Optimized also predicts benign. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. No tools predict pathogenicity, and AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is Likely Benign, while Foldetta results are unavailable. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.819762 | Disordered | 0.979325 | Binding | 0.347 | 0.896 | 0.875 | -4.488 | Likely Benign | 0.460 | Ambiguous | Likely Benign | 0.087 | Likely Benign | -1.13 | Neutral | 0.224 | Benign | 0.058 | Benign | 4.19 | Benign | 0.12 | Tolerated | 0.2008 | 0.4972 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.3245A>G | Q1082R 2D  AIThe SynGAP1 missense variant Q1082R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.819762 | Disordered | 0.979325 | Binding | 0.347 | 0.896 | 0.875 | -3.584 | Likely Benign | 0.418 | Ambiguous | Likely Benign | 0.076 | Likely Benign | -0.96 | Neutral | 0.224 | Benign | 0.058 | Benign | 4.14 | Benign | 0.10 | Tolerated | 0.1590 | 0.3014 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3245A>T | Q1082L 2D AIThe SynGAP1 missense variant Q1082L is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.819762 | Disordered | 0.979325 | Binding | 0.347 | 0.896 | 0.875 | -3.284 | Likely Benign | 0.171 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -1.30 | Neutral | 0.224 | Benign | 0.058 | Benign | 4.12 | Benign | 1.00 | Tolerated | 0.0979 | 0.6824 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3277C>A | Q1093K 2D AIThe SynGAP1 missense variant Q1093K is reported in gnomAD (variant ID 6‑33443829‑C‑A) but has no ClinVar entry. Functional prediction tools uniformly favor a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all classify the change as benign, and AlphaMissense‑Optimized also predicts benign. No tool predicts pathogenicity; AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.983312 | Binding | 0.351 | 0.886 | 1.000 | 6-33443829-C-A | -3.919 | Likely Benign | 0.558 | Ambiguous | Likely Benign | 0.061 | Likely Benign | -0.92 | Neutral | 0.224 | Benign | 0.091 | Benign | 2.78 | Benign | 0.06 | Tolerated | 3.77 | 5 | 0.1926 | 0.5861 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||||||||||
| c.3278A>G | Q1093R 2D AIThe SynGAP1 missense variant Q1093R is reported in gnomAD (ID 6‑33443830‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic prediction comes from SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.983312 | Binding | 0.351 | 0.886 | 1.000 | 6-33443830-A-G | 1 | 6.40e-7 | -3.681 | Likely Benign | 0.483 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -1.06 | Neutral | 0.224 | Benign | 0.091 | Benign | 2.73 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1537 | 0.3904 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||
| c.3278A>T | Q1093L 2D AIThe SynGAP1 missense variant Q1093L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.983312 | Binding | 0.351 | 0.886 | 1.000 | -3.242 | Likely Benign | 0.165 | Likely Benign | Likely Benign | 0.055 | Likely Benign | -1.10 | Neutral | 0.224 | Benign | 0.091 | Benign | 2.72 | Benign | 0.03 | Affected | 0.0899 | 0.6837 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3293G>C | S1098T 2D AIThe SynGAP1 missense variant S1098T is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.973030 | Binding | 0.337 | 0.855 | 1.000 | -4.144 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.051 | Likely Benign | -0.18 | Neutral | 0.224 | Benign | 0.120 | Benign | 2.76 | Benign | 0.88 | Tolerated | 0.1706 | 0.6653 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3380G>A | G1127E 2D  AIThe SynGAP1 missense variant G1127E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; the remaining tools (ESM1b and AlphaMissense‑Default) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a tie between benign and uncertain calls. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the evidence strongly points to a benign effect, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.852422 | Binding | 0.344 | 0.915 | 0.875 | -7.359 | In-Between | 0.422 | Ambiguous | Likely Benign | 0.314 | Likely Benign | -0.56 | Neutral | 0.224 | Benign | 0.091 | Benign | 4.88 | Benign | 0.15 | Tolerated | 0.1395 | 0.4244 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.3403A>G | K1135E 2D  AIThe SynGAP1 missense variant K1135E is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33443955‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Only AlphaMissense‑Default predicts a pathogenic outcome, while AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and Foldetta (combining FoldX‑MD and Rosetta) results are unavailable. Overall, the preponderance of evidence points to a benign impact for K1135E, and this assessment does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.790969 | Binding | 0.303 | 0.889 | 0.875 | 6-33443955-A-G | -6.499 | Likely Benign | 0.924 | Likely Pathogenic | Ambiguous | 0.239 | Likely Benign | -0.76 | Neutral | 0.224 | Benign | 0.237 | Benign | 5.45 | Benign | 0.12 | Tolerated | 4.32 | 2 | 0.4330 | 0.1476 | 1 | 0 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||||||||
| c.3416A>T | Q1139L 2D AIThe SynGAP1 missense variant Q1139L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -1.689 | Likely Benign | 0.153 | Likely Benign | Likely Benign | 0.472 | Likely Benign | -3.75 | Deleterious | 0.224 | Benign | 0.237 | Benign | 5.30 | Benign | 0.00 | Affected | 0.0745 | 0.5599 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3829C>A | H1277N 2D AIThe SynGAP1 missense variant H1277N is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447877‑C‑A). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | 6-33447877-C-A | -3.347 | Likely Benign | 0.193 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -4.96 | Deleterious | 0.224 | Benign | 0.120 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1562 | 0.1250 | 1 | 2 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||
| c.3830A>T | H1277L 2D AIThe SynGAP1 missense variant H1277L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | -2.949 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.215 | Likely Benign | -7.93 | Deleterious | 0.224 | Benign | 0.091 | Benign | 2.17 | Pathogenic | 0.00 | Affected | 0.0822 | 0.4158 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3867G>C | K1289N 2D  AIThe SynGAP1 missense variant K1289N is reported in ClinVar as “Not submitted” and is not present in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic call comes from SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, SGM‑Consensus also predicts likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the collective predictions, K1289N is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.828700 | Binding | 0.548 | 0.787 | 0.625 | -4.244 | Likely Benign | 0.481 | Ambiguous | Likely Benign | 0.043 | Likely Benign | -1.56 | Neutral | 0.224 | Benign | 0.120 | Benign | 2.59 | Benign | 0.02 | Affected | 0.3442 | 0.0940 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3867G>T | K1289N 2D AIThe SynGAP1 missense variant K1289N is reported in ClinVar as “Not submitted” and is not present in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic call comes from SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, SGM‑Consensus also predicts likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the collective predictions, K1289N is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.828700 | Binding | 0.548 | 0.787 | 0.625 | -4.244 | Likely Benign | 0.481 | Ambiguous | Likely Benign | 0.043 | Likely Benign | -1.56 | Neutral | 0.224 | Benign | 0.120 | Benign | 2.59 | Benign | 0.02 | Affected | 0.3442 | 0.0940 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3889A>G | R1297G 2D  AIThe SynGAP1 missense variant R1297G is reported in gnomAD (ID 6‑33451763‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus itself is Benign; Foldetta results are unavailable. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that R1297G is most likely benign, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.859585 | Disordered | 0.895222 | Binding | 0.511 | 0.817 | 0.625 | 6-33451763-A-G | 3 | 1.86e-6 | -2.833 | Likely Benign | 0.168 | Likely Benign | Likely Benign | 0.215 | Likely Benign | -3.65 | Deleterious | 0.224 | Benign | 0.171 | Benign | 2.56 | Benign | 0.01 | Affected | 3.77 | 5 | 0.3089 | 0.2400 | -2 | -3 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||
| c.3890G>C | R1297T 2D  AIThe SynGAP1 missense variant R1297T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Functional prediction tools cluster into two groups: seven tools (REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a benign effect, while two tools (PROVEAN, FATHMM) predict pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.859585 | Disordered | 0.895222 | Binding | 0.511 | 0.817 | 0.625 | -3.941 | Likely Benign | 0.287 | Likely Benign | Likely Benign | 0.151 | Likely Benign | -2.95 | Deleterious | 0.224 | Benign | 0.171 | Benign | 2.49 | Pathogenic | 0.08 | Tolerated | 0.1634 | 0.2972 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3891G>C | R1297S 2D  AIThe SynGAP1 missense variant R1297S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the consensus of available predictions indicates that R1297S is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.859585 | Disordered | 0.895222 | Binding | 0.511 | 0.817 | 0.625 | -3.468 | Likely Benign | 0.401 | Ambiguous | Likely Benign | 0.139 | Likely Benign | -2.39 | Neutral | 0.224 | Benign | 0.096 | Benign | 2.50 | Benign | 0.07 | Tolerated | 0.2863 | 0.2347 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||||
| c.3891G>T | R1297S 2D AIThe SynGAP1 missense variant R1297S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions strongly suggests that R1297S is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.859585 | Disordered | 0.895222 | Binding | 0.511 | 0.817 | 0.625 | -3.468 | Likely Benign | 0.401 | Ambiguous | Likely Benign | 0.139 | Likely Benign | -2.39 | Neutral | 0.224 | Benign | 0.096 | Benign | 2.50 | Benign | 0.07 | Tolerated | 0.2863 | 0.2347 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||||
| c.3893A>T | Q1298L 2D AIThe SynGAP1 missense variant Q1298L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.895297 | Binding | 0.410 | 0.821 | 0.750 | -2.251 | Likely Benign | 0.147 | Likely Benign | Likely Benign | 0.221 | Likely Benign | -3.04 | Deleterious | 0.224 | Benign | 0.078 | Benign | 2.79 | Benign | 0.04 | Affected | 0.0667 | 0.4164 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.238A>G | K80E 2D AIThe SynGAP1 K80E missense variant has no ClinVar record and is not listed in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The high‑accuracy consensus (SGM‑Consensus) is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yielding a “Likely Benign” classification. AlphaMissense‑Optimized returns an uncertain result, and Foldetta (which would combine FoldX‑MD and Rosetta outputs) has no available data for this variant. Overall, the balance of evidence favors a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar entry exists for K80E. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.477530 | Uncertain | 0.331 | 0.873 | 0.500 | -5.605 | Likely Benign | 0.943 | Likely Pathogenic | Ambiguous | 0.034 | Likely Benign | -0.91 | Neutral | 0.225 | Benign | 0.027 | Benign | 3.92 | Benign | 0.00 | Affected | 0.3850 | 0.0952 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.262G>C | V88L 2D AIThe SynGAP1 V88L variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome; Foldetta results are unavailable. Overall, the majority of tools suggest a benign impact, but the high‑accuracy predictions are conflicting. The variant is most likely benign based on the collective evidence, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.552910 | Binding | 0.323 | 0.870 | 0.500 | -5.808 | Likely Benign | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.066 | Likely Benign | -0.79 | Neutral | 0.225 | Benign | 0.027 | Benign | 3.74 | Benign | 0.00 | Affected | 0.1457 | 0.4562 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.262G>T | V88L 2D AIThe SynGAP1 missense variant V88L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM, whereas SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized predict a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, while the SGM Consensus (a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect; a Foldetta stability analysis is unavailable. Overall, the majority of predictions lean toward a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign based on the current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.552910 | Binding | 0.323 | 0.870 | 0.500 | -5.808 | Likely Benign | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.066 | Likely Benign | -0.79 | Neutral | 0.225 | Benign | 0.027 | Benign | 3.74 | Benign | 0.00 | Affected | 0.1457 | 0.4562 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.265C>G | P89A 2D  AIThe SynGAP1 missense variant P89A is listed in ClinVar (ID 1031674) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and Foldetta results are unavailable. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments therefore indicate a benign or uncertain impact: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus is likely benign, and Foldetta data are missing. Overall, the balance of evidence points to the variant being most likely benign, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.545797 | Binding | 0.316 | 0.865 | 0.500 | Conflicting | 3 | -5.778 | Likely Benign | 0.920 | Likely Pathogenic | Ambiguous | 0.095 | Likely Benign | -2.47 | Neutral | 0.225 | Benign | 0.020 | Benign | 3.77 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3407 | 0.3768 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||
| c.265C>T | P89S 2D AIThe SynGAP1 missense variant P89S (ClinVar ID 4801335) is listed as “Uncertain” in ClinVar and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are AlphaMissense‑Default, AlphaMissense‑Optimized, and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus (majority vote) remains likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (six benign vs. three pathogenic) and the SGM‑Consensus support a benign interpretation, which does not contradict the ClinVar “Uncertain” status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.545797 | Binding | 0.316 | 0.865 | 0.500 | Uncertain | 1 | -5.198 | Likely Benign | 0.979 | Likely Pathogenic | Likely Pathogenic | 0.099 | Likely Benign | -2.40 | Neutral | 0.225 | Benign | 0.020 | Benign | 3.76 | Benign | 0.00 | Affected | 0.3459 | 0.4191 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||
| c.3946A>G | N1316D 2D  AIThe SynGAP1 missense variant N1316D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451820‑A‑G). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also leans toward benign, with two benign and one pathogenic prediction among the four considered. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence indicates a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451820-A-G | -2.717 | Likely Benign | 0.364 | Ambiguous | Likely Benign | 0.057 | Likely Benign | -2.80 | Deleterious | 0.225 | Benign | 0.084 | Benign | 3.96 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2095 | 0.3745 | 1 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||
| c.3970C>T | P1324S 2D AIThe SynGAP1 missense variant P1324S is listed in ClinVar as Benign (ClinVar ID 1137951.0) and is present in gnomAD (ID 6‑33451844‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote) also benign; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, which is consistent with its ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | Likely Benign | 1 | 6-33451844-C-T | 5 | 3.26e-6 | -5.451 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.049 | Likely Benign | 0.35 | Neutral | 0.225 | Benign | 0.092 | Benign | 4.33 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3505 | 0.4450 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||
| c.4025A>G | D1342G 2D  AIThe SynGAP1 missense variant D1342G is reported in gnomAD (variant ID 6‑33451899‑A‑G) but has no ClinVar entry. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.981682 | Binding | 0.316 | 0.678 | 0.875 | 6-33451899-A-G | 1 | 7.29e-7 | -3.227 | Likely Benign | 0.129 | Likely Benign | Likely Benign | 0.021 | Likely Benign | -0.89 | Neutral | 0.225 | Benign | 0.045 | Benign | 4.05 | Benign | 0.09 | Tolerated | 4.32 | 4 | 0.3633 | 0.5601 | -1 | 1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||
| c.4026C>A | D1342E 2D  AIThe SynGAP1 missense variant D1342E is catalogued in gnomAD (ID 6‑33451900‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes. Only SIFT predicts a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that D1342E is most likely benign, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.981682 | Binding | 0.316 | 0.678 | 0.875 | 6-33451900-C-A | -3.169 | Likely Benign | 0.134 | Likely Benign | Likely Benign | 0.026 | Likely Benign | -0.88 | Neutral | 0.225 | Benign | 0.084 | Benign | 4.16 | Benign | 0.04 | Affected | 4.32 | 4 | 0.2130 | 0.5720 | 2 | 3 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||
| c.4026C>G | D1342E 2D AIThe SynGAP1 missense variant D1342E is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the consensus of available predictions points to a benign effect for D1342E, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.981682 | Binding | 0.316 | 0.678 | 0.875 | -3.169 | Likely Benign | 0.134 | Likely Benign | Likely Benign | 0.026 | Likely Benign | -0.88 | Neutral | 0.225 | Benign | 0.084 | Benign | 4.16 | Benign | 0.04 | Affected | 4.32 | 4 | 0.2130 | 0.5720 | 2 | 3 | 0.0 | 14.03 | |||||||||||||||||||||||||||||||||||||
| c.55G>T | A19S 2D AIThe SynGAP1 missense variant A19S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.443393 | Uncertain | 0.378 | 0.906 | 0.500 | -2.905 | Likely Benign | 0.107 | Likely Benign | Likely Benign | 0.029 | Likely Benign | -0.21 | Neutral | 0.225 | Benign | 0.027 | Benign | 4.17 | Benign | 0.00 | Affected | 0.3273 | 0.6260 | 1 | 1 | -2.6 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.1957C>G | L653V 2D  AIThe SynGAP1 missense variant L653V is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that classify the variant as benign include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX, Rosetta, and premPS, while ESM1b is inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of evidence points to a benign impact, and this does not contradict the ClinVar “Uncertain” classification. Thus, based on current predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.049374 | Structured | 0.335213 | Uncertain | 0.963 | 0.332 | 0.000 | Uncertain | 1 | -7.050 | In-Between | 0.301 | Likely Benign | Likely Benign | 3.28 | Destabilizing | 0.3 | 2.18 | Destabilizing | 2.73 | Destabilizing | 1.32 | Destabilizing | 0.146 | Likely Benign | -2.25 | Neutral | 0.227 | Benign | 0.039 | Benign | 3.28 | Benign | 0.08 | Tolerated | 0.1436 | 0.3557 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||
| c.1318A>G | N440D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N440D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while FoldX, Foldetta, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -9.335 | Likely Pathogenic | 0.407 | Ambiguous | Likely Benign | -0.62 | Ambiguous | 0.0 | -0.41 | Likely Benign | -0.52 | Ambiguous | 0.47 | Likely Benign | 0.074 | Likely Benign | -1.71 | Neutral | 0.229 | Benign | 0.045 | Benign | 3.43 | Benign | 0.43 | Tolerated | 0.1544 | 0.2025 | 2 | 1 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.1919C>A | T640N 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T640N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as tolerated or benign. Only the protein‑stability predictors FoldX and Rosetta returned uncertain results, which are treated as unavailable evidence. High‑accuracy assessments corroborate the benign prediction: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also reports a benign effect. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.066181 | Structured | 0.137043 | Uncertain | 0.893 | 0.284 | 0.000 | -4.258 | Likely Benign | 0.107 | Likely Benign | Likely Benign | -0.50 | Ambiguous | 0.2 | 0.53 | Ambiguous | 0.02 | Likely Benign | 0.03 | Likely Benign | 0.097 | Likely Benign | 0.51 | Neutral | 0.229 | Benign | 0.084 | Benign | 3.45 | Benign | 0.25 | Tolerated | 0.1518 | 0.4079 | 0 | 0 | -2.8 | 13.00 | |||||||||||||||||||||||||||||
| c.3343A>T | I1115F 2D  AIThe SynGAP1 missense variant I1115F is reported in gnomAD (ID 6‑33443895‑A‑T) and has no ClinVar entry. All available in silico predictors classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are not available. Based on the unanimous benign predictions and the lack of a ClinVar pathogenic classification, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.889439 | Disordered | 0.892339 | Binding | 0.308 | 0.912 | 0.750 | 6-33443895-A-T | -3.426 | Likely Benign | 0.095 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -0.98 | Neutral | 0.230 | Benign | 0.098 | Benign | 2.71 | Benign | 0.19 | Tolerated | 4.32 | 2 | 0.0769 | 0.4338 | 0 | 1 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||
| c.321G>C | R107S 2D AIThe SynGAP1 missense variant R107S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome; Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, and this is not contradicted by any ClinVar annotation. Thus, the variant is most likely benign, although high‑accuracy tools provide conflicting evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.666105 | Disordered | 0.663448 | Binding | 0.331 | 0.863 | 0.875 | -1.162 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.147 | Likely Benign | -2.37 | Neutral | 0.231 | Benign | 0.037 | Benign | 2.99 | Benign | 0.00 | Affected | 0.2771 | 0.4148 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||||
| c.321G>T | R107S 2D AIThe SynGAP1 missense variant R107S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome; Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, but the high‑accuracy tools provide conflicting evidence. Thus, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.666105 | Disordered | 0.663448 | Binding | 0.331 | 0.863 | 0.875 | -1.162 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.147 | Likely Benign | -2.37 | Neutral | 0.231 | Benign | 0.037 | Benign | 2.99 | Benign | 0.00 | Affected | 0.2771 | 0.4148 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||||
| c.326G>C | S109T 2D AIThe SynGAP1 missense variant S109T is catalogued in gnomAD (6‑33432191‑G‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all report benign or likely benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments confirm the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus, derived from the majority of the high‑accuracy predictors, is benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that S109T is most likely benign, and this assessment does not contradict any ClinVar status (none). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.622677 | Disordered | 0.669335 | Binding | 0.328 | 0.864 | 0.750 | 6-33432191-G-C | 2 | 1.24e-6 | -4.065 | Likely Benign | 0.449 | Ambiguous | Likely Benign | 0.090 | Likely Benign | -1.19 | Neutral | 0.231 | Benign | 0.050 | Benign | 3.59 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1519 | 0.5212 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.335G>C | G112A 2D AIThe SynGAP1 missense variant G112A is listed in ClinVar with an uncertain significance (ClinVar ID 1425533.0) and is present in gnomAD (6‑33432200‑G‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also benign. Foldetta results are unavailable. Overall, the majority of computational evidence indicates that the variant is most likely benign, which is consistent with its ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.728858 | Disordered | 0.640153 | Binding | 0.332 | 0.867 | 0.750 | Uncertain | 1 | 6-33432200-G-C | 15 | 9.30e-6 | -2.456 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -2.34 | Neutral | 0.231 | Benign | 0.054 | Benign | 4.07 | Benign | 0.00 | Affected | 3.61 | 5 | 0.3817 | 0.4429 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||||
| c.356A>C | E119A 2D  AIThe SynGAP1 missense variant E119A is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM, while those that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs. 2 pathogenic), and Foldetta results are unavailable. Overall, the balance of evidence (five benign versus three pathogenic predictions) suggests the variant is most likely benign. This conclusion does not contradict ClinVar status, as the variant has no existing ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.661946 | Binding | 0.346 | 0.881 | 0.750 | -4.881 | Likely Benign | 0.647 | Likely Pathogenic | Likely Benign | 0.108 | Likely Benign | -2.52 | Deleterious | 0.231 | Benign | 0.074 | Benign | 3.84 | Benign | 0.01 | Affected | 0.4374 | 0.7514 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.544T>A | S182T 2D AIThe SynGAP1 missense variant S182T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of available predictions (five benign vs three pathogenic) suggest a benign impact. This consensus does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Thus, based on current computational evidence, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.501700 | Disordered | 0.436016 | Uncertain | 0.368 | 0.619 | 0.625 | -11.247 | Likely Pathogenic | 0.857 | Likely Pathogenic | Ambiguous | 0.128 | Likely Benign | -2.29 | Neutral | 0.231 | Benign | 0.081 | Benign | 3.68 | Benign | 0.00 | Affected | 0.1624 | 0.5657 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.553T>G | S185A 2D AIThe SynGAP1 missense variant S185A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. AlphaMissense‑Optimized is uncertain, and no Foldetta stability result is available. High‑accuracy assessments therefore show a likely pathogenic consensus from SGM‑Consensus, with no contradictory evidence from ClinVar or population databases. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any existing ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.545602 | Disordered | 0.430485 | Uncertain | 0.365 | 0.623 | 0.500 | -11.839 | Likely Pathogenic | 0.868 | Likely Pathogenic | Ambiguous | 0.203 | Likely Benign | -2.57 | Deleterious | 0.231 | Benign | 0.107 | Benign | 3.65 | Benign | 0.00 | Affected | 0.5147 | 0.4472 | Weaken | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||
| c.572G>C | S191T 2D AIThe SynGAP1 missense variant S191T is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools overwhelmingly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score benign, while the majority‑vote SGM‑Consensus also classifies it as likely benign. Only SIFT predicts a pathogenic outcome, and ESM1b remains uncertain. High‑accuracy assessments corroborate the benign consensus: AlphaMissense‑Optimized returns a benign prediction and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports likely benign. Foldetta stability analysis is not available for this variant. Overall, the preponderance of evidence supports a benign classification, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.428475 | Uncertain | 0.322 | 0.615 | 0.125 | -7.315 | In-Between | 0.319 | Likely Benign | Likely Benign | 0.150 | Likely Benign | -2.34 | Neutral | 0.231 | Benign | 0.081 | Benign | 3.81 | Benign | 0.03 | Affected | 0.1466 | 0.7405 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.662A>C | E221A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E221A missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta as benign. Overall, the predictions are split, but the two high‑confidence pathogenic calls outweigh the single high‑confidence benign call, suggesting the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.127496 | Structured | 0.413334 | Uncertain | 0.891 | 0.283 | 0.000 | -12.906 | Likely Pathogenic | 0.964 | Likely Pathogenic | Likely Pathogenic | 0.31 | Likely Benign | 0.0 | -0.11 | Likely Benign | 0.10 | Likely Benign | 0.52 | Ambiguous | 0.796 | Likely Pathogenic | -4.70 | Deleterious | 0.231 | Benign | 0.081 | Benign | 5.82 | Benign | 0.01 | Affected | 0.3486 | 0.7335 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.667A>G | T223A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T223A is listed in ClinVar with an “Uncertain” status and is present in the gnomAD database (ID 6‑33435518‑A‑G). Functional prediction tools that agree on a benign effect include FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL and PROVEAN. Predictions that are inconclusive are Rosetta, Foldetta, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also resolves to benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains uncertain. Overall, the majority of evidence points to a benign impact, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.070400 | Structured | 0.382605 | Uncertain | 0.867 | 0.316 | 0.125 | Uncertain | 1 | 6-33435518-A-G | 3 | 1.86e-6 | -7.076 | In-Between | 0.316 | Likely Benign | Likely Benign | 0.30 | Likely Benign | 0.1 | 0.77 | Ambiguous | 0.54 | Ambiguous | 0.74 | Ambiguous | 0.574 | Likely Pathogenic | -3.36 | Deleterious | 0.231 | Benign | 0.058 | Benign | 5.74 | Benign | 0.09 | Tolerated | 3.41 | 13 | 0.2982 | 0.3031 | 1 | 0 | 2.5 | -30.03 | 186.4 | 44.0 | 0.0 | 0.0 | 0.0 | 0.0 | X | X | Uncertain | The introduced residue Ala223 is located on the outer surface of an anti-parallel β sheet strand (res. Cys219-Thr224). Unlike the hydroxyl group of the Thr223 side chain in the WT protein, the methyl side chain of Ala223 cannot form hydrogen bonds with nearby residues Thr228 and Lys207. Without these hydrogen-bonding interactions at the β sheet surface, the secondary structure element becomes unstable and partially unfolds in the variant simulations. However, since the model ends abruptly at the N-terminus, no definite conclusions can be drawn from the simulations. | |||||||||||||
| c.2629C>G | L877V 2D AIThe SynGAP1 missense variant L877V is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence strongly supports a benign classification, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.653063 | Disordered | 0.634010 | Binding | 0.265 | 0.875 | 0.250 | -5.063 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.022 | Likely Benign | -0.10 | Neutral | 0.232 | Benign | 0.109 | Benign | 2.71 | Benign | 0.26 | Tolerated | 0.1726 | 0.3178 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.440A>C | Q147P 2D AIThe SynGAP1 missense variant Q147P is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. Grouping by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Overall, the balance of evidence, especially the SGM Consensus and the majority of individual predictors, indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.541878 | Disordered | 0.503877 | Binding | 0.349 | 0.840 | 0.625 | -14.132 | Likely Pathogenic | 0.797 | Likely Pathogenic | Ambiguous | 0.264 | Likely Benign | -3.03 | Deleterious | 0.232 | Benign | 0.147 | Benign | 3.87 | Benign | 0.01 | Affected | 0.2350 | 0.4402 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||||||||||||||
| c.14G>C | R5P 2D  AIThe SynGAP1 missense variant R5P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that R5P is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.595080 | Disordered | 0.547847 | Binding | 0.363 | 0.920 | 0.750 | -3.438 | Likely Benign | 0.189 | Likely Benign | Likely Benign | 0.286 | Likely Benign | -0.35 | Neutral | 0.233 | Benign | 0.013 | Benign | 4.10 | Benign | 0.00 | Affected | 0.2294 | 0.5530 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.29G>C | R10P 2D AIThe SynGAP1 missense variant R10P is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33420293‑G‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority of the four high‑accuracy tools) is benign; Foldetta results are unavailable. Overall, the collective evidence points to a benign effect for R10P, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.534167 | Disordered | 0.513657 | Binding | 0.330 | 0.915 | 0.625 | Uncertain | 2 | 6-33420293-G-C | 2 | 1.30e-6 | -3.772 | Likely Benign | 0.162 | Likely Benign | Likely Benign | 0.220 | Likely Benign | -0.05 | Neutral | 0.233 | Benign | 0.026 | Benign | 4.13 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2261 | 0.5245 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||
| c.457A>T | T153S 2D AIThe SynGAP1 missense variant T153S is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.501700 | Disordered | 0.502105 | Binding | 0.297 | 0.818 | 0.625 | -1.890 | Likely Benign | 0.118 | Likely Benign | Likely Benign | 0.118 | Likely Benign | -0.50 | Neutral | 0.235 | Benign | 0.039 | Benign | 4.19 | Benign | 0.14 | Tolerated | 0.3756 | 0.2886 | 1 | 1 | -0.1 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.458C>G | T153S 2D AIThe SynGAP1 missense variant T153S is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.501700 | Disordered | 0.502105 | Binding | 0.297 | 0.818 | 0.625 | -1.890 | Likely Benign | 0.118 | Likely Benign | Likely Benign | 0.066 | Likely Benign | -0.50 | Neutral | 0.235 | Benign | 0.039 | Benign | 4.19 | Benign | 0.14 | Tolerated | 0.3756 | 0.2886 | 1 | 1 | -0.1 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3919C>T | P1307S 2D AIThe SynGAP1 missense variant P1307S is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. No pathogenic predictions are present among the evaluated tools. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.798249 | Disordered | 0.913511 | Binding | 0.491 | 0.901 | 0.875 | -3.700 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.049 | Likely Benign | 0.45 | Neutral | 0.239 | Benign | 0.157 | Benign | 3.09 | Benign | 0.48 | Tolerated | 0.3652 | 0.4846 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2707G>C | G903R 2D  AIThe SynGAP1 missense variant G903R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of standard predictors lean toward a benign impact, and no ClinVar annotation contradicts this assessment. Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.680603 | Disordered | 0.549818 | Binding | 0.291 | 0.917 | 0.375 | -3.503 | Likely Benign | 0.865 | Likely Pathogenic | Ambiguous | 0.119 | Likely Benign | -2.02 | Neutral | 0.241 | Benign | 0.244 | Benign | 2.33 | Pathogenic | 0.02 | Affected | 0.0872 | 0.4064 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.3502A>C | I1168L 2D AIThe SynGAP1 missense variant I1168L is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors indicates a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the substitution as tolerated, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also classifies it as likely benign. Only AlphaMissense‑Default predicts a pathogenic outcome, but this is an outlier relative to the other tools. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus (majority vote) concurs. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so it does not influence the overall assessment. Overall, the computational evidence overwhelmingly favors a benign classification, and this is consistent with the absence of a ClinVar pathogenic report. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.763262 | Binding | 0.423 | 0.796 | 0.500 | -1.855 | Likely Benign | 0.604 | Likely Pathogenic | Likely Benign | 0.361 | Likely Benign | -0.60 | Neutral | 0.241 | Benign | 0.286 | Benign | 5.48 | Benign | 0.20 | Tolerated | 0.0956 | 0.4465 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3508A>G | S1170G 2D AIThe SynGAP1 missense variant S1170G is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools that agree on benign impact include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool in the dataset predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign, while Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.622677 | Disordered | 0.719138 | Binding | 0.417 | 0.767 | 0.500 | Uncertain | 1 | -4.288 | Likely Benign | 0.221 | Likely Benign | Likely Benign | 0.349 | Likely Benign | -0.81 | Neutral | 0.241 | Benign | 0.229 | Benign | 5.31 | Benign | 0.54 | Tolerated | 4.32 | 4 | 0.2705 | 0.4558 | 1 | 0 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||||||
| c.3886G>A | E1296K 2D  AIThe SynGAP1 missense variant E1296K is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (no gnomAD ID). Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM; pathogenic predictions come from PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact, with no conflict with ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.837511 | Disordered | 0.894444 | Binding | 0.530 | 0.809 | 0.625 | -3.435 | Likely Benign | 0.713 | Likely Pathogenic | Likely Benign | 0.136 | Likely Benign | -2.80 | Deleterious | 0.241 | Benign | 0.210 | Benign | 2.65 | Benign | 0.05 | Affected | 0.1951 | 0.6138 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.568A>G | S190G 2D AIThe SynGAP1 missense variant S190G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Overall, the consensus of all available predictions is that the variant is most likely benign, and this conclusion is consistent with the lack of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.422041 | Structured | 0.428613 | Uncertain | 0.338 | 0.615 | 0.250 | -5.440 | Likely Benign | 0.321 | Likely Benign | Likely Benign | 0.048 | Likely Benign | -1.26 | Neutral | 0.243 | Benign | 0.079 | Benign | 4.11 | Benign | 0.19 | Tolerated | 0.2780 | 0.4765 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.586T>G | L196V 2D AIThe SynGAP1 missense variant L196V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of evidence (five benign vs three pathogenic) points to a benign impact. This conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.284882 | Structured | 0.429452 | Uncertain | 0.489 | 0.521 | 0.125 | -8.702 | Likely Pathogenic | 0.898 | Likely Pathogenic | Ambiguous | 0.106 | Likely Benign | -2.31 | Neutral | 0.243 | Benign | 0.097 | Benign | 3.71 | Benign | 0.01 | Affected | 0.1436 | 0.2850 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.670A>G | T224A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant T224A is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33435521‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and AlphaMissense‑Default. Predictions that are inconclusive are Rosetta, Foldetta, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.104810 | Structured | 0.360921 | Uncertain | 0.848 | 0.315 | 0.125 | Conflicting | 4 | 6-33435521-A-G | 2 | 1.24e-6 | -7.379 | In-Between | 0.651 | Likely Pathogenic | Likely Benign | 0.33 | Likely Benign | 0.1 | 1.05 | Ambiguous | 0.69 | Ambiguous | 0.91 | Ambiguous | 0.464 | Likely Benign | -2.96 | Deleterious | 0.243 | Benign | 0.079 | Benign | 5.57 | Benign | 0.57 | Tolerated | 3.41 | 13 | 0.4253 | 0.5053 | 1 | 0 | 2.5 | -30.03 | 169.0 | 41.4 | -0.5 | 1.1 | -0.4 | 0.0 | X | X | Uncertain | The introduced residue Ala224 is located on the outer surface of an anti-parallel β sheet strand (res. Cys219-Thr224). Unlike the hydroxyl group of the Thr224 side chain in the WT model, the methyl side chain of Ala224 cannot form hydrogen bonds with nearby residues Ser204, Ser226, and Gly227. Without these hydrogen-bonding interactions at the β sheet surface, the secondary structure element becomes unstable and unfolds during the variant simulations. However, since the model ends abruptly at the N-terminus, no definite conclusions can be drawn from the simulations. | |||||||||||||
| c.1106C>G | T369R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T369R is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438011‑C‑G). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. Rosetta and Foldetta are uncertain, so their results are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is also uncertain. Overall, the majority of evidence (nine benign vs two pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | 6-33438011-C-G | 3 | 1.93e-6 | -6.772 | Likely Benign | 0.571 | Likely Pathogenic | Likely Benign | -0.27 | Likely Benign | 0.1 | 1.48 | Ambiguous | 0.61 | Ambiguous | 0.29 | Likely Benign | 0.148 | Likely Benign | -2.15 | Neutral | 0.244 | Benign | 0.107 | Benign | 1.72 | Pathogenic | 0.32 | Tolerated | 3.42 | 19 | 0.1217 | 0.3737 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||
| c.1951G>C | E651Q 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E651Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM—all classifying the change as benign. No tool predicts a pathogenic outcome. The high‑accuracy assessments are consistent: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign effect; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also reports a benign result. Consequently, the variant is most likely benign based on the available predictions, and this assessment does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.088832 | Structured | 0.365409 | Uncertain | 0.955 | 0.340 | 0.000 | -6.308 | Likely Benign | 0.476 | Ambiguous | Likely Benign | 0.13 | Likely Benign | 0.1 | 0.15 | Likely Benign | 0.14 | Likely Benign | -0.13 | Likely Benign | 0.158 | Likely Benign | -1.23 | Neutral | 0.244 | Benign | 0.075 | Benign | 3.34 | Benign | 0.58 | Tolerated | 0.1438 | 0.5893 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||
| c.1960G>C | E654Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E654Q missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), and FATHMM, while those that predict a pathogenic outcome are SGM‑Consensus, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicating likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicting a benign effect. Overall, the majority of tools (nine benign vs. five pathogenic) suggest the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar assertion is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.026892 | Structured | 0.303029 | Uncertain | 0.957 | 0.311 | 0.000 | -10.368 | Likely Pathogenic | 0.699 | Likely Pathogenic | Likely Benign | 0.00 | Likely Benign | 0.0 | 0.18 | Likely Benign | 0.09 | Likely Benign | -0.12 | Likely Benign | 0.298 | Likely Benign | -2.79 | Deleterious | 0.244 | Benign | 0.075 | Benign | 3.33 | Benign | 0.02 | Affected | 0.1140 | 0.3913 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||
| c.2747T>C | V916A 2D  AIThe SynGAP1 missense variant V916A is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity all converge on a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. No tool in the dataset indicates a pathogenic effect, so the pathogenic‑prediction group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign,” and the Foldetta stability analysis is not available. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.835395 | Binding | 0.308 | 0.879 | 0.250 | -2.524 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.116 | Likely Benign | -0.18 | Neutral | 0.244 | Benign | 0.171 | Benign | 2.91 | Benign | 1.00 | Tolerated | 0.3455 | 0.2841 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.362C>A | A121D 2D  AIThe SynGAP1 missense variant A121D is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.779859 | Disordered | 0.661304 | Binding | 0.362 | 0.888 | 0.750 | -3.626 | Likely Benign | 0.595 | Likely Pathogenic | Likely Benign | 0.096 | Likely Benign | -0.89 | Neutral | 0.244 | Benign | 0.050 | Benign | 4.06 | Benign | 0.03 | Affected | 0.1876 | 0.1945 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.512C>A | A171D 2D AIThe SynGAP1 missense variant A171D is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments therefore lean toward a benign interpretation: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus is benign, and Foldetta data are missing. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.492272 | Uncertain | 0.358 | 0.652 | 0.375 | -6.977 | Likely Benign | 0.908 | Likely Pathogenic | Ambiguous | 0.144 | Likely Benign | -1.45 | Neutral | 0.244 | Benign | 0.037 | Benign | 4.15 | Benign | 0.01 | Affected | 0.1634 | 0.2462 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.521T>C | M174T 2D AIThe SynGAP1 missense variant M174T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.661982 | Disordered | 0.485854 | Uncertain | 0.373 | 0.620 | 0.375 | -9.174 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.286 | Likely Benign | -3.12 | Deleterious | 0.244 | Benign | 0.049 | Benign | 4.08 | Benign | 0.01 | Affected | 0.1949 | 0.2334 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.2303A>C | D768A 2D  AIThe SynGAP1 D768A variant is listed in gnomAD (ID 6‑33442461‑A‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM) and pathogenic predictions (ESM1b, AlphaMissense‑Default). AlphaMissense‑Optimized returns an uncertain result. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑vs‑2 tie and thus unavailable; Foldetta stability analysis is not reported. Overall, the preponderance of evidence (six benign vs two pathogenic) points to a benign effect. This conclusion does not contradict ClinVar, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.332115 | Structured | 0.928237 | Binding | 0.314 | 0.877 | 0.250 | 6-33442461-A-C | -8.153 | Likely Pathogenic | 0.786 | Likely Pathogenic | Ambiguous | 0.174 | Likely Benign | -1.84 | Neutral | 0.245 | Benign | 0.096 | Benign | 4.09 | Benign | 0.14 | Tolerated | 3.64 | 6 | 0.3924 | 0.7662 | -2 | 0 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||
| c.2380C>A | P794T 2D AIThe SynGAP1 missense variant P794T is catalogued in gnomAD (ID 6‑33442932‑C‑A) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the dataset predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign prediction. The Foldetta stability analysis is not available for this variant. Overall, the consensus of all available predictions is benign, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.979741 | Disordered | 0.408951 | Uncertain | 0.550 | 0.898 | 0.875 | 6-33442932-C-A | 1 | 6.20e-7 | -4.838 | Likely Benign | 0.060 | Likely Benign | Likely Benign | 0.046 | Likely Benign | -0.34 | Neutral | 0.245 | Benign | 0.138 | Benign | 4.25 | Benign | 0.66 | Tolerated | 4.07 | 3 | 0.1739 | 0.6207 | -1 | 0 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||
| c.2666G>C | G889A 2D AIThe SynGAP1 missense variant G889A is listed in gnomAD (6‑33443218‑G‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign or likely benign. Only FATHMM predicts a pathogenic outcome, representing a single dissenting signal. High‑accuracy assessments confirm the benign consensus: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates a likely benign status. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the preponderance of evidence supports a benign classification, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.690604 | Disordered | 0.552581 | Binding | 0.331 | 0.928 | 0.625 | 6-33443218-G-C | 1 | 6.20e-7 | -4.580 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.063 | Likely Benign | -1.49 | Neutral | 0.245 | Benign | 0.096 | Benign | 2.47 | Pathogenic | 0.76 | Tolerated | 4.32 | 4 | 0.3838 | 0.4902 | 0 | 1 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.2696T>C | I899T 2D  AIThe SynGAP1 missense variant I899T is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33443248‑T‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.443727 | Uncertain | 0.292 | 0.928 | 0.375 | 6-33443248-T-C | 2 | 1.24e-6 | -2.472 | Likely Benign | 0.521 | Ambiguous | Likely Benign | 0.103 | Likely Benign | -0.25 | Neutral | 0.245 | Benign | 0.096 | Benign | 2.75 | Benign | 0.01 | Affected | 4.32 | 4 | 0.1006 | 0.1014 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||
| c.2701G>A | A901T 2D AIThe SynGAP1 missense variant A901T is reported as “Likely Benign” in SGM‑Consensus and has no ClinVar entry or gnomAD allele. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool in the dataset predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also benign. Foldetta results are not available. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.595080 | Disordered | 0.489838 | Uncertain | 0.306 | 0.917 | 0.375 | -4.708 | Likely Benign | 0.116 | Likely Benign | Likely Benign | 0.046 | Likely Benign | -0.95 | Neutral | 0.245 | Benign | 0.096 | Benign | 2.70 | Benign | 0.16 | Tolerated | 0.1587 | 0.7680 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2903G>C | G968A 2D AIThe SynGAP1 missense variant G968A is predicted to be benign by all available in‑silico tools. Consensus predictors (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign. No tools predict pathogenicity, so the pathogenic group is empty. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized reports benign, and the SGM‑Consensus indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Based on the unanimous benign predictions and the absence of ClinVar evidence, the variant is most likely benign and does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.961360 | Binding | 0.327 | 0.896 | 0.750 | -4.721 | Likely Benign | 0.070 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -0.33 | Neutral | 0.245 | Benign | 0.140 | Benign | 4.24 | Benign | 0.68 | Tolerated | 0.3401 | 0.5154 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2993C>T | A998V 2D AIThe SynGAP1 missense variant A998V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also reports a benign prediction. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.759478 | Disordered | 0.951758 | Binding | 0.318 | 0.902 | 0.500 | -4.795 | Likely Benign | 0.101 | Likely Benign | Likely Benign | 0.051 | Likely Benign | -1.09 | Neutral | 0.245 | Benign | 0.138 | Benign | 4.09 | Benign | 0.00 | Affected | 0.1281 | 0.6147 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||||||||||||
| c.3151G>A | G1051S 2D  AIThe SynGAP1 missense variant G1051S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only FATHMM predicts a pathogenic outcome. When the predictions are grouped, the benign consensus includes eight tools, whereas the pathogenic consensus contains a single tool. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates a likely benign outcome. Foldetta data are unavailable, so no stability inference can be drawn. Overall, the computational evidence overwhelmingly supports a benign classification for G1051S, and this conclusion is consistent with the absence of any ClinVar annotation. Thus, the variant is most likely benign, and this assessment does not contradict ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987317 | Disordered | 0.900141 | Binding | 0.358 | 0.936 | 0.875 | -4.742 | Likely Benign | 0.079 | Likely Benign | Likely Benign | 0.445 | Likely Benign | 0.10 | Neutral | 0.245 | Benign | 0.096 | Benign | -0.74 | Pathogenic | 0.61 | Tolerated | 0.2446 | 0.4911 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3152G>T | G1051V 2D AIThe SynGAP1 missense variant G1051V is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443704‑G‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign prediction (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for G1051V, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.987317 | Disordered | 0.900141 | Binding | 0.358 | 0.936 | 0.875 | 6-33443704-G-T | 1 | 6.20e-7 | -7.098 | In-Between | 0.102 | Likely Benign | Likely Benign | 0.460 | Likely Benign | -0.62 | Neutral | 0.245 | Benign | 0.096 | Benign | -0.74 | Pathogenic | 0.17 | Tolerated | 3.77 | 5 | 0.1274 | 0.3680 | -3 | -1 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||
| c.3289C>G | P1097A 2D AIThe SynGAP1 missense variant P1097A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts pathogenicity. The high‑accuracy consensus (SGM‑Consensus, derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the unanimous benign predictions and the lack of any pathogenic evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.974957 | Binding | 0.384 | 0.858 | 1.000 | -4.315 | Likely Benign | 0.072 | Likely Benign | Likely Benign | 0.059 | Likely Benign | -1.77 | Neutral | 0.245 | Benign | 0.140 | Benign | 2.71 | Benign | 0.26 | Tolerated | 0.3199 | 0.5381 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3334G>A | E1112K 2D AIThe SynGAP1 missense variant E1112K is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools largely agree on a benign effect: SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized all predict benign. Only two tools predict pathogenicity: SIFT and AlphaMissense‑Default. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available, so it does not influence the conclusion. Overall, the majority of predictions, including the high‑accuracy tools, suggest the variant is most likely benign, and this is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.909381 | Binding | 0.335 | 0.902 | 0.875 | -3.772 | Likely Benign | 0.684 | Likely Pathogenic | Likely Benign | 0.210 | Likely Benign | 0.15 | Neutral | 0.245 | Benign | 0.096 | Benign | 2.82 | Benign | 0.02 | Affected | 0.3002 | 0.7704 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3391C>A | P1131T 2D AIThe SynGAP1 missense variant P1131T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | -4.766 | Likely Benign | 0.219 | Likely Benign | Likely Benign | 0.313 | Likely Benign | -2.93 | Deleterious | 0.245 | Benign | 0.096 | Benign | 5.30 | Benign | 0.00 | Affected | 0.1353 | 0.6315 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3511G>A | A1171T 2D AIThe SynGAP1 missense variant A1171T is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. All available in‑silico predictors classify the change as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a “Likely Benign” outcome. No tool predicts pathogenicity. High‑accuracy assessments confirm the benign prediction: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.702689 | Binding | 0.472 | 0.775 | 0.500 | Uncertain | 1 | -3.658 | Likely Benign | 0.149 | Likely Benign | Likely Benign | 0.201 | Likely Benign | -0.48 | Neutral | 0.245 | Benign | 0.138 | Benign | 5.45 | Benign | 0.07 | Tolerated | 4.32 | 4 | 0.1421 | 0.5465 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||||||
| c.3512C>G | A1171G 2D AIThe SynGAP1 missense variant A1171G (Ala→Gly at residue 1171) is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a benign effect, and no tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome, while Foldetta results are unavailable. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.702689 | Binding | 0.472 | 0.775 | 0.500 | -3.487 | Likely Benign | 0.286 | Likely Benign | Likely Benign | 0.141 | Likely Benign | -0.60 | Neutral | 0.245 | Benign | 0.138 | Benign | 5.38 | Benign | 0.08 | Tolerated | 0.1957 | 0.3201 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3512C>T | A1171V 2D AIThe SynGAP1 missense variant A1171V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a benign effect, and no tool predicts pathogenicity. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign; Foldetta results are not available. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.702689 | Binding | 0.472 | 0.775 | 0.500 | -3.516 | Likely Benign | 0.325 | Likely Benign | Likely Benign | 0.152 | Likely Benign | -0.72 | Neutral | 0.245 | Benign | 0.138 | Benign | 5.34 | Benign | 0.11 | Tolerated | 0.1019 | 0.4611 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||||
| c.1181A>C | K394T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 K394T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. Predictions that are uncertain or inconclusive are FoldX, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, the balance of evidence (five benign versus three pathogenic predictions) suggests the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | -6.487 | Likely Benign | 0.599 | Likely Pathogenic | Likely Benign | 0.50 | Ambiguous | 0.1 | 2.46 | Destabilizing | 1.48 | Ambiguous | 0.57 | Ambiguous | 0.482 | Likely Benign | -3.35 | Deleterious | 0.247 | Benign | 0.166 | Benign | 4.61 | Benign | 0.01 | Affected | 0.2727 | 0.4453 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||
| c.1313C>G | A438G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A438G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only PROVEAN predicts a pathogenic outcome. Predictions that are inconclusive or unavailable are FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as Likely Benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact for A438G, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.147574 | Structured | 0.290154 | Uncertain | 0.929 | 0.293 | 0.000 | -5.790 | Likely Benign | 0.182 | Likely Benign | Likely Benign | 1.08 | Ambiguous | 0.1 | 1.78 | Ambiguous | 1.43 | Ambiguous | 0.80 | Ambiguous | 0.034 | Likely Benign | -2.51 | Deleterious | 0.247 | Benign | 0.037 | Benign | 4.12 | Benign | 0.11 | Tolerated | 0.1788 | 0.2719 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||
| c.397C>G | L133V 2D AIThe SynGAP1 missense variant L133V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are limited: AlphaMissense‑Optimized remains uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes); Foldetta results are unavailable. Overall, the majority of evidence (five benign vs. three pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.422041 | Structured | 0.718429 | Binding | 0.320 | 0.896 | 0.250 | -8.429 | Likely Pathogenic | 0.848 | Likely Pathogenic | Ambiguous | 0.082 | Likely Benign | -1.27 | Neutral | 0.247 | Benign | 0.163 | Benign | 3.61 | Benign | 0.03 | Affected | 0.1561 | 0.1883 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.412A>G | K138E 2D AIThe SynGAP1 missense variant K138E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2). AlphaMissense‑Optimized specifically predicts pathogenicity, while Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the majority of predictions lean toward a benign outcome (5 benign vs 4 pathogenic), and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.590140 | Disordered | 0.619482 | Binding | 0.349 | 0.901 | 0.375 | -11.902 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.212 | Likely Benign | -2.13 | Neutral | 0.247 | Benign | 0.125 | Benign | 3.60 | Benign | 0.01 | Affected | 0.3611 | 0.0974 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.413A>G | K138R 2D  AIThe SynGAP1 missense variant K138R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta results are unavailable. Taken together, the preponderance of evidence points to a benign impact for K138R, and this conclusion does not conflict with the absence of a ClinVar assertion. The variant is most likely benign, and this assessment does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.619482 | Binding | 0.349 | 0.901 | 0.375 | -5.135 | Likely Benign | 0.406 | Ambiguous | Likely Benign | 0.115 | Likely Benign | -1.58 | Neutral | 0.247 | Benign | 0.163 | Benign | 3.66 | Benign | 0.02 | Affected | 0.4555 | 0.1036 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.424A>G | K142E 2D AIThe SynGAP1 missense variant K142E is not reported in ClinVar (no ClinVar entry) and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions point to a pathogenic effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -14.450 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.249 | Likely Benign | -2.59 | Deleterious | 0.247 | Benign | 0.125 | Benign | 3.49 | Benign | 0.00 | Affected | 0.3644 | 0.0952 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.425A>C | K142T 2D AIThe SynGAP1 missense variant K142T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -12.013 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.251 | Likely Benign | -3.77 | Deleterious | 0.247 | Benign | 0.166 | Benign | 3.47 | Benign | 0.00 | Affected | 0.2034 | 0.2708 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.433A>G | K145E 2D AIThe SynGAP1 missense variant K145E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions arise from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of high‑confidence tools predict a pathogenic impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -12.571 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.245 | Likely Benign | -2.51 | Deleterious | 0.247 | Benign | 0.125 | Benign | 3.68 | Benign | 0.00 | Affected | 0.3638 | 0.1179 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.434A>C | K145T 2D AIThe SynGAP1 missense variant K145T is not reported in ClinVar and is absent from gnomAD. Computational assessment shows a split: benign predictions from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, whereas pathogenic predictions come from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN, classifies the change as Likely Pathogenic. High‑accuracy tools further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus itself is a high‑confidence prediction; Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic impact for K145T. Thus, the variant is most likely pathogenic, and there is no ClinVar annotation to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -8.519 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.224 | Likely Benign | -3.54 | Deleterious | 0.247 | Benign | 0.166 | Benign | 3.63 | Benign | 0.00 | Affected | 0.1965 | 0.3611 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.446A>G | K149R 2D AIThe SynGAP1 missense variant K149R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that K149R is most likely benign, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -8.237 | Likely Pathogenic | 0.406 | Ambiguous | Likely Benign | 0.143 | Likely Benign | -1.70 | Neutral | 0.247 | Benign | 0.125 | Benign | 3.70 | Benign | 0.00 | Affected | 0.5639 | 0.1169 | Weaken | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.478C>G | L160V 2D AIThe SynGAP1 missense variant L160V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (six benign vs. three pathogenic) indicate that the variant is most likely benign. This conclusion is consistent with the lack of ClinVar evidence and gnomAD presence, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.526760 | Binding | 0.275 | 0.728 | 0.125 | -12.506 | Likely Pathogenic | 0.729 | Likely Pathogenic | Likely Benign | 0.055 | Likely Benign | -1.44 | Neutral | 0.247 | Benign | 0.113 | Benign | 3.90 | Benign | 0.00 | Affected | 0.1553 | 0.3319 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.481C>G | P161A 2D AIThe SynGAP1 missense variant P161A is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. When predictions are grouped by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Overall, the balance of evidence, especially the SGM Consensus and the majority of individual predictors, indicates that P161A is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -9.012 | Likely Pathogenic | 0.926 | Likely Pathogenic | Ambiguous | 0.079 | Likely Benign | -3.52 | Deleterious | 0.247 | Benign | 0.091 | Benign | 3.95 | Benign | 0.00 | Affected | 0.3600 | 0.4123 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.2254T>A | S752T 2D AIThe SynGAP1 missense variant S752T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.604312 | Disordered | 0.690594 | Binding | 0.365 | 0.877 | 0.625 | -4.040 | Likely Benign | 0.101 | Likely Benign | Likely Benign | 0.029 | Likely Benign | -1.33 | Neutral | 0.248 | Benign | 0.137 | Benign | 1.55 | Pathogenic | 0.07 | Tolerated | 0.1633 | 0.6342 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3670C>G | L1224V 2D AIThe SynGAP1 missense variant L1224V is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. No Foldetta (FoldX‑MD/Rosetta) stability prediction is available, so it does not influence the assessment. Overall, the preponderance of evidence from multiple prediction algorithms and consensus methods indicates that the variant is most likely benign, and this conclusion does not contradict the current ClinVar status of “Uncertain.” Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.525368 | Disordered | 0.441554 | Uncertain | 0.871 | 0.543 | 0.500 | Uncertain | 1 | -5.796 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.093 | Likely Benign | -1.41 | Neutral | 0.248 | Benign | 0.112 | Benign | 2.42 | Pathogenic | 0.18 | Tolerated | 0.1397 | 0.2289 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||
| c.1832T>A | M611K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 M611K variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that classify it as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. The majority of other in silico predictors (SGM‑Consensus, REVEL, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) indicate a pathogenic effect; FoldX is uncertain and therefore not considered evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) and Foldetta (combining FoldX‑MD and Rosetta outputs) both predict pathogenicity. Based on the preponderance of pathogenic predictions and the high‑accuracy consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.236433 | Structured | 0.210791 | Uncertain | 0.870 | 0.253 | 0.000 | -13.021 | Likely Pathogenic | 0.630 | Likely Pathogenic | Likely Benign | 1.86 | Ambiguous | 0.6 | 2.65 | Destabilizing | 2.26 | Destabilizing | 1.65 | Destabilizing | 0.665 | Likely Pathogenic | -4.10 | Deleterious | 0.250 | Benign | 0.120 | Benign | -1.26 | Pathogenic | 0.03 | Affected | 0.1193 | 0.0688 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1833G>A | M611I 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant M611I is reported in gnomAD (ID 6‑33440885‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized; pathogenic predictions arise from SGM‑Consensus, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments further clarify the variant’s likely effect: AlphaMissense‑Optimized classifies it as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates pathogenicity, and Foldetta, which integrates FoldX‑MD and Rosetta outputs, yields an uncertain stability change. No folding‑stability method provides definitive evidence. Overall, the majority of predictions lean toward pathogenicity, and this conclusion does not conflict with ClinVar status, which lacks an entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.236433 | Structured | 0.210791 | Uncertain | 0.870 | 0.253 | 0.000 | 6-33440885-G-A | 1 | 6.19e-7 | -8.552 | Likely Pathogenic | 0.736 | Likely Pathogenic | Likely Benign | 1.45 | Ambiguous | 0.4 | 1.36 | Ambiguous | 1.41 | Ambiguous | 0.72 | Ambiguous | 0.292 | Likely Benign | -2.10 | Neutral | 0.250 | Benign | 0.091 | Benign | -1.14 | Pathogenic | 0.38 | Tolerated | 3.37 | 35 | 0.1009 | 0.2302 | 1 | 2 | 2.6 | -18.03 | ||||||||||||||||||||||||
| c.1833G>C | M611I 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant M611I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are SGM‑Consensus, ESM1b, FATHMM, and AlphaMissense‑Default. Predictions that are inconclusive are FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. Overall, the majority of evidence points toward a pathogenic classification, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.236433 | Structured | 0.210791 | Uncertain | 0.870 | 0.253 | 0.000 | -8.552 | Likely Pathogenic | 0.736 | Likely Pathogenic | Likely Benign | 1.45 | Ambiguous | 0.4 | 1.36 | Ambiguous | 1.41 | Ambiguous | 0.72 | Ambiguous | 0.292 | Likely Benign | -2.10 | Neutral | 0.250 | Benign | 0.091 | Benign | -1.14 | Pathogenic | 0.38 | Tolerated | 3.37 | 35 | 0.1009 | 0.2302 | 1 | 2 | 2.6 | -18.03 | |||||||||||||||||||||||||||
| c.1833G>T | M611I 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant M611I is not reported in ClinVar and is absent from gnomAD. In silico predictors that classify the variant as benign include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Predictors that classify it as pathogenic are SGM‑Consensus, ESM1b, FATHMM, and AlphaMissense‑Default. Four tools (FoldX, Rosetta, premPS, and Foldetta) provide uncertain or unavailable results. High‑accuracy assessment shows AlphaMissense‑Optimized predicts benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic; Foldetta’s stability output is unavailable. Overall, the majority of predictions lean toward a benign effect, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.236433 | Structured | 0.210791 | Uncertain | 0.870 | 0.253 | 0.000 | -8.552 | Likely Pathogenic | 0.736 | Likely Pathogenic | Likely Benign | 1.45 | Ambiguous | 0.4 | 1.36 | Ambiguous | 1.41 | Ambiguous | 0.72 | Ambiguous | 0.292 | Likely Benign | -2.10 | Neutral | 0.250 | Benign | 0.091 | Benign | -1.14 | Pathogenic | 0.38 | Tolerated | 3.37 | 35 | 0.1009 | 0.2302 | 1 | 2 | 2.6 | -18.03 | |||||||||||||||||||||||||||
| c.3238G>A | A1080T 2D AIThe SynGAP1 missense variant A1080T (ClinVar ID 1473274.0) is listed as “Uncertain” in ClinVar and is present in gnomAD (ID 6‑33443790‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool in the dataset predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence strongly suggests the variant is most likely benign, which does not contradict the current ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.981457 | Binding | 0.303 | 0.900 | 0.750 | Conflicting | 2 | 6-33443790-G-A | 17 | 1.06e-5 | -3.928 | Likely Benign | 0.133 | Likely Benign | Likely Benign | 0.144 | Likely Benign | -0.19 | Neutral | 0.253 | Benign | 0.042 | Benign | 4.10 | Benign | 0.60 | Tolerated | 3.77 | 5 | 0.1564 | 0.7103 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||||
| c.1430T>A | M477K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS and FATHMM. Predictions that are uncertain or inconclusive are FoldX, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is unavailable due to mixed or uncertain inputs. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the current ClinVar status, which contains no report of pathogenicity. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -7.519 | In-Between | 0.553 | Ambiguous | Likely Benign | 0.54 | Ambiguous | 0.1 | 0.37 | Likely Benign | 0.46 | Likely Benign | 1.12 | Destabilizing | 0.371 | Likely Benign | -1.32 | Neutral | 0.254 | Benign | 0.122 | Benign | -1.15 | Pathogenic | 0.14 | Tolerated | 0.1817 | 0.0847 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||
| c.2669G>A | R890H 2D  AIThe SynGAP1 missense variant R890H is listed in ClinVar as a benign alteration (ClinVar ID 1037885.0) and is observed in gnomAD (6‑33443221‑G‑A). All evaluated in‑silico predictors agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores, and no tool predicts pathogenicity. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign,” while Foldetta’s protein‑folding stability analysis is unavailable. Overall, the computational evidence strongly supports a benign classification, which is consistent with the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.720929 | Disordered | 0.531156 | Binding | 0.284 | 0.928 | 0.625 | Benign | 1 | 6-33443221-G-A | 19 | 1.18e-5 | -3.600 | Likely Benign | 0.198 | Likely Benign | Likely Benign | 0.056 | Likely Benign | -1.29 | Neutral | 0.254 | Benign | 0.134 | Benign | 3.97 | Benign | 0.15 | Tolerated | 4.32 | 4 | 0.3032 | 0.1235 | 2 | 0 | 1.3 | -19.05 | ||||||||||||||||||||||||||||||||
| c.1055C>A | T352N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T352N is listed in ClinVar as Benign (ClinVar ID 590151.0) and is present in the gnomAD database (gnomAD ID 6‑33437960‑C‑A). Across the broad panel of in‑silico predictors, 13 tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) uniformly report a benign effect, whereas only FATHMM predicts pathogenicity. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is also benign. No predictions or stability analyses are missing or inconclusive. Overall, the computational evidence strongly supports a benign classification, consistent with the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.137348 | Structured | 0.367886 | Uncertain | 0.926 | 0.329 | 0.000 | Likely Benign | 1 | 6-33437960-C-A | 2 | 1.24e-6 | -4.817 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.20 | Likely Benign | 0.0 | -0.04 | Likely Benign | 0.08 | Likely Benign | 0.45 | Likely Benign | 0.027 | Likely Benign | -0.92 | Neutral | 0.255 | Benign | 0.057 | Benign | 1.75 | Pathogenic | 0.19 | Tolerated | 3.37 | 25 | 0.1311 | 0.4358 | 0 | 0 | -2.8 | 13.00 | 208.4 | -14.5 | -0.2 | 0.1 | -0.1 | 0.0 | X | Potentially Benign | Thr352 is located in a short α helical section within a loop connecting two β strands (res. Gly341-Pro349, res. Thr359-Pro364) originating from two different anti-parallel β sheets of the C2 domain. In the WT simulations, the side chain hydroxyl and backbone amide groups of Thr354 form hydrogen bonds with the backbone carbonyl group of Pro349 at the end of the preceding β strand. This arrangement likely stabilizes the α helical section and aids in folding, keeping the short secondary structure element intact in the variant simulations. However, the carboxamide group of the Asn352 side chain does not form hydrogen bonds with the backbone carbonyl group of Pro349. Instead, it packs against the cyclic ring and forms hydrogen bonds with the phenol group of the Tyr363 side chain in the other β strand. | |||||||||||||
| c.1061C>A | A354D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A354D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. Those that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. Four tools (Foldetta, premPS, ESM1b, and Rosetta) return uncertain results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors a pathogenic interpretation. Foldetta’s stability prediction is uncertain. Overall, the majority of consensus tools lean toward a benign effect, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.381329 | Uncertain | 0.882 | 0.335 | 0.125 | -7.914 | In-Between | 0.725 | Likely Pathogenic | Likely Benign | 0.22 | Likely Benign | 0.1 | 1.10 | Ambiguous | 0.66 | Ambiguous | 0.57 | Ambiguous | 0.087 | Likely Benign | -1.49 | Neutral | 0.255 | Benign | 0.053 | Benign | 1.75 | Pathogenic | 0.07 | Tolerated | 0.1759 | 0.1345 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1076C>A | T359K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 T359K missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. The remaining tools—FoldX, Rosetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions (seven benign vs. two pathogenic) support a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.281712 | Structured | 0.414952 | Uncertain | 0.939 | 0.480 | 0.250 | -10.822 | Likely Pathogenic | 0.517 | Ambiguous | Likely Benign | -0.65 | Ambiguous | 0.1 | 0.66 | Ambiguous | 0.01 | Likely Benign | 0.75 | Ambiguous | 0.163 | Likely Benign | -2.23 | Neutral | 0.255 | Benign | 0.045 | Benign | 1.79 | Pathogenic | 0.24 | Tolerated | 0.1618 | 0.3483 | 0 | -1 | -3.2 | 27.07 | ||||||||||||||||||||||||||||||
| c.2033G>C | S678T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678T is not reported in ClinVar and is absent from gnomAD. All available in‑silico predictors classify the change as benign: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). No tool predicts pathogenicity. High‑accuracy assessments likewise support a benign effect: AlphaMissense‑Optimized is benign, the SGM Consensus is “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign stability. Based on the unanimous computational evidence, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -5.389 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.32 | Likely Benign | 0.1 | 0.41 | Likely Benign | 0.37 | Likely Benign | 0.14 | Likely Benign | 0.053 | Likely Benign | -1.82 | Neutral | 0.255 | Benign | 0.053 | Benign | 3.41 | Benign | 0.11 | Tolerated | 0.1494 | 0.6351 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||
| c.2613C>A | H871Q 2D  AIThe SynGAP1 missense variant H871Q is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the collective evidence strongly supports a benign classification, and this conclusion is consistent with the lack of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.679301 | Binding | 0.279 | 0.858 | 0.250 | -4.049 | Likely Benign | 0.140 | Likely Benign | Likely Benign | 0.061 | Likely Benign | -0.67 | Neutral | 0.255 | Benign | 0.113 | Benign | 2.69 | Benign | 0.17 | Tolerated | 3.88 | 3 | 0.1384 | 0.3357 | 0 | 3 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||
| c.2613C>G | H871Q 2D AIThe SynGAP1 missense variant H871Q is reported in gnomAD (ID 6‑33443165‑C‑G) and has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability predictor, was not available for this variant. Overall, the evidence strongly supports a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.679301 | Binding | 0.279 | 0.858 | 0.250 | 6-33443165-C-G | 1 | 6.20e-7 | -4.049 | Likely Benign | 0.140 | Likely Benign | Likely Benign | 0.061 | Likely Benign | -0.67 | Neutral | 0.255 | Benign | 0.113 | Benign | 2.69 | Benign | 0.17 | Tolerated | 3.88 | 3 | 0.1384 | 0.3357 | 0 | 3 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||
| c.2871T>A | H957Q 2D AIThe SynGAP1 missense variant H957Q is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.968874 | Binding | 0.362 | 0.915 | 0.750 | -6.304 | Likely Benign | 0.130 | Likely Benign | Likely Benign | 0.113 | Likely Benign | -0.87 | Neutral | 0.255 | Benign | 0.105 | Benign | 2.54 | Benign | 0.56 | Tolerated | 0.2227 | 0.4114 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2871T>G | H957Q 2D AIThe SynGAP1 missense variant H957Q is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.968874 | Binding | 0.362 | 0.915 | 0.750 | -6.304 | Likely Benign | 0.130 | Likely Benign | Likely Benign | 0.113 | Likely Benign | -0.87 | Neutral | 0.255 | Benign | 0.105 | Benign | 2.54 | Benign | 0.56 | Tolerated | 0.2227 | 0.4114 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2877C>A | H959Q 2D AIThe SynGAP1 missense variant H959Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as likely benign. Only ESM1b predicts a pathogenic outcome, representing the sole discordant signal. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that H959Q is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.980566 | Binding | 0.333 | 0.905 | 0.750 | -8.657 | Likely Pathogenic | 0.126 | Likely Benign | Likely Benign | 0.182 | Likely Benign | -0.77 | Neutral | 0.255 | Benign | 0.105 | Benign | 4.15 | Benign | 0.10 | Tolerated | 0.2264 | 0.3936 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2877C>G | H959Q 2D AIThe SynGAP1 missense variant H959Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign, while the single pathogenic prediction comes from ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that H959Q is most likely benign, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.980566 | Binding | 0.333 | 0.905 | 0.750 | -8.657 | Likely Pathogenic | 0.126 | Likely Benign | Likely Benign | 0.182 | Likely Benign | -0.77 | Neutral | 0.255 | Benign | 0.105 | Benign | 4.15 | Benign | 0.10 | Tolerated | 0.2264 | 0.3936 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2883C>A | H961Q 2D AIThe SynGAP1 missense variant H961Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only SIFT and ESM1b predict pathogenicity. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict (3 benign vs. 1 pathogenic). High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.989835 | Disordered | 0.984562 | Binding | 0.323 | 0.893 | 0.750 | -8.368 | Likely Pathogenic | 0.118 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.49 | Neutral | 0.255 | Benign | 0.105 | Benign | 4.17 | Benign | 0.02 | Affected | 0.2022 | 0.3743 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2883C>G | H961Q 2D AIThe SynGAP1 missense variant H961Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT and ESM1b predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a majority‑benign vote and is reported as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the available predictions indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.989835 | Disordered | 0.984562 | Binding | 0.323 | 0.893 | 0.750 | -8.368 | Likely Pathogenic | 0.118 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.49 | Neutral | 0.255 | Benign | 0.105 | Benign | 4.17 | Benign | 0.02 | Affected | 0.2022 | 0.3743 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2982G>C | K994N 2D AIThe SynGAP1 missense variant K994N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic call comes from SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, SGM‑Consensus confirms a Likely Benign status, and Foldetta data are unavailable. Taken together, the preponderance of evidence indicates that K994N is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.930054 | Binding | 0.289 | 0.912 | 0.750 | -3.724 | Likely Benign | 0.519 | Ambiguous | Likely Benign | 0.011 | Likely Benign | -0.73 | Neutral | 0.255 | Benign | 0.113 | Benign | 4.09 | Benign | 0.01 | Affected | 0.4010 | 0.2376 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.2982G>T | K994N 2D AIThe SynGAP1 missense variant K994N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic call comes from SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, SGM‑Consensus confirms a Likely Benign status, and Foldetta data are unavailable. Taken together, the preponderance of evidence indicates that K994N is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.930054 | Binding | 0.289 | 0.912 | 0.750 | -3.724 | Likely Benign | 0.519 | Ambiguous | Likely Benign | 0.009 | Likely Benign | -0.73 | Neutral | 0.255 | Benign | 0.113 | Benign | 4.09 | Benign | 0.01 | Affected | 0.4010 | 0.2376 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3443T>G | M1148R 2D AIThe SynGAP1 missense variant M1148R is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. No Foldetta stability analysis is available for this variant. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -2.303 | Likely Benign | 0.669 | Likely Pathogenic | Likely Benign | 0.129 | Likely Benign | -1.77 | Neutral | 0.255 | Benign | 0.113 | Benign | 2.54 | Benign | 0.00 | Affected | 0.1624 | 0.0888 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.1598C>G | A533G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A533G resides in the GAP domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome; Rosetta is uncertain and therefore not counted as evidence. High‑accuracy assessments—all of which are available—show AlphaMissense‑Optimized as Benign, SGM‑Consensus as Likely Benign, and Foldetta as Benign, reinforcing the benign consensus. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.179055 | Structured | 0.026324 | Uncertain | 0.843 | 0.393 | 0.000 | -3.412 | Likely Benign | 0.122 | Likely Benign | Likely Benign | 0.32 | Likely Benign | 0.1 | 0.64 | Ambiguous | 0.48 | Likely Benign | 0.33 | Likely Benign | 0.185 | Likely Benign | -1.94 | Neutral | 0.258 | Benign | 0.099 | Benign | -1.23 | Pathogenic | 0.23 | Tolerated | 0.2348 | 0.4737 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||
| c.1595C>A | T532K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T532K missense variant is not reported in ClinVar and has no entries in gnomAD. Prediction tools that classify the change as benign include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict pathogenicity are ESM1b, FATHMM, AlphaMissense‑Default, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. With ten benign versus four pathogenic predictions and two of the three high‑accuracy tools supporting a benign outcome, the variant is most likely benign. This assessment does not contradict the ClinVar status, which currently has no classification for T532K. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.021478 | Uncertain | 0.889 | 0.385 | 0.000 | -8.460 | Likely Pathogenic | 0.644 | Likely Pathogenic | Likely Benign | -0.44 | Likely Benign | 0.2 | 0.24 | Likely Benign | -0.10 | Likely Benign | 0.34 | Likely Benign | 0.376 | Likely Benign | -1.97 | Neutral | 0.259 | Benign | 0.033 | Benign | -1.21 | Pathogenic | 0.29 | Tolerated | 0.0943 | 0.1992 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||
| c.1598C>A | A533E 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A533E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.026324 | Uncertain | 0.843 | 0.393 | 0.000 | -10.810 | Likely Pathogenic | 0.553 | Ambiguous | Likely Benign | -0.13 | Likely Benign | 0.0 | -0.26 | Likely Benign | -0.20 | Likely Benign | 0.43 | Likely Benign | 0.314 | Likely Benign | -1.89 | Neutral | 0.259 | Benign | 0.107 | Benign | -1.18 | Pathogenic | 0.18 | Tolerated | 0.1147 | 0.1498 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||
| c.2404G>C | G802R 2D AIThe SynGAP1 missense variant G802R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. Only SIFT predicts a pathogenic effect, and AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign interpretation: AlphaMissense‑Optimized reports benign, and the SGM‑Consensus (majority vote) likewise predicts likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence supports a benign classification for G802R, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.894241 | Disordered | 0.681966 | Binding | 0.294 | 0.898 | 0.625 | -4.756 | Likely Benign | 0.504 | Ambiguous | Likely Benign | 0.072 | Likely Benign | -1.05 | Neutral | 0.259 | Benign | 0.196 | Benign | 2.68 | Benign | 0.01 | Affected | 0.0995 | 0.4514 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||
| c.2572A>G | S858G 2D AIThe SynGAP1 missense variant S858G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while only polyPhen‑2 HumVar indicates a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also predicts benign. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.482724 | Uncertain | 0.305 | 0.833 | 0.375 | -2.181 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.143 | Likely Benign | -0.18 | Neutral | 0.259 | Benign | 0.786 | Possibly Damaging | 4.13 | Benign | 0.11 | Tolerated | 0.3084 | 0.5116 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2573G>C | S858T 2D AIThe SynGAP1 missense variant S858T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumVar predicts it as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that S858T is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.482724 | Uncertain | 0.305 | 0.833 | 0.375 | -4.960 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -0.92 | Neutral | 0.259 | Benign | 0.786 | Possibly Damaging | 4.14 | Benign | 0.06 | Tolerated | 0.1660 | 0.6489 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3131C>G | P1044R 2D  AIThe SynGAP1 missense variant P1044R is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the collective evidence strongly supports a benign classification, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.979741 | Disordered | 0.952126 | Binding | 0.331 | 0.855 | 0.750 | -3.969 | Likely Benign | 0.242 | Likely Benign | Likely Benign | 0.368 | Likely Benign | 0.79 | Neutral | 0.259 | Benign | 0.140 | Benign | 5.45 | Benign | 1.00 | Tolerated | 0.1451 | 0.4270 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3176G>T | G1059V 2D AIThe SynGAP1 missense variant G1059V is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in silico predictors indicates a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score benign, while the majority‑vote SGM‑Consensus also classifies it as likely benign. Only SIFT predicts a pathogenic outcome, and ESM1b remains uncertain. High‑accuracy tools corroborate the benign prediction: AlphaMissense‑Optimized is benign and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely benign; Foldetta results are not available. Overall, the preponderance of evidence supports a benign classification for G1059V, and this assessment does not conflict with ClinVar, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.983019 | Disordered | 0.898939 | Binding | 0.399 | 0.926 | 0.875 | -7.242 | In-Between | 0.106 | Likely Benign | Likely Benign | 0.478 | Likely Benign | -0.82 | Neutral | 0.259 | Benign | 0.066 | Benign | 2.54 | Benign | 0.00 | Affected | 0.1462 | 0.3494 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3185G>T | G1062V 2D AIThe SynGAP1 missense variant G1062V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.976962 | Disordered | 0.936972 | Binding | 0.368 | 0.917 | 0.875 | -6.598 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 0.377 | Likely Benign | -0.78 | Neutral | 0.259 | Benign | 0.066 | Benign | 4.12 | Benign | 0.01 | Affected | 0.1441 | 0.3694 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3293G>T | S1098I 2D AIThe SynGAP1 missense variant S1098I is catalogued in gnomAD (ID 6‑33443845‑G‑T) and has no ClinVar entry. All evaluated in‑silico predictors classify it as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool reports a pathogenic outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.973030 | Binding | 0.337 | 0.855 | 1.000 | 6-33443845-G-T | -4.497 | Likely Benign | 0.182 | Likely Benign | Likely Benign | 0.139 | Likely Benign | -0.92 | Neutral | 0.259 | Benign | 0.066 | Benign | 2.67 | Benign | 0.15 | Tolerated | 3.77 | 5 | 0.1219 | 0.5601 | -2 | -1 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||
| c.3440C>T | T1147I 2D  AIThe SynGAP1 missense variant T1147I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.831250 | Disordered | 0.746520 | Binding | 0.345 | 0.839 | 0.875 | -3.552 | Likely Benign | 0.151 | Likely Benign | Likely Benign | 0.423 | Likely Benign | -2.03 | Neutral | 0.259 | Benign | 0.059 | Benign | 5.42 | Benign | 0.01 | Affected | 0.1029 | 0.4968 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||||||||||||
| c.3830A>G | H1277R 2D AIThe SynGAP1 missense variant H1277R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | -2.940 | Likely Benign | 0.306 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -5.60 | Deleterious | 0.259 | Benign | 0.066 | Benign | 2.15 | Pathogenic | 0.00 | Affected | 0.1710 | 0.1562 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||||||
| c.563G>A | S188N 2D  AIThe SynGAP1 missense variant S188N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of available predictions (five benign vs three pathogenic) suggest a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.490133 | Structured | 0.428502 | Uncertain | 0.298 | 0.603 | 0.500 | -10.307 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 0.095 | Likely Benign | -2.34 | Neutral | 0.259 | Benign | 0.048 | Benign | 3.92 | Benign | 0.01 | Affected | 0.1420 | 0.5658 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||||||||
| c.709G>T | A237S 2D AIThe SynGAP1 missense variant A237S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, AlphaMissense‑Optimized, and FATHMM. No tool predicts a pathogenic outcome; the only inconclusive results come from premPS and ESM1b, which are treated as unavailable. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.200174 | Structured | 0.334699 | Uncertain | 0.719 | 0.352 | 0.000 | -7.696 | In-Between | 0.112 | Likely Benign | Likely Benign | 0.48 | Likely Benign | 0.6 | 0.34 | Likely Benign | 0.41 | Likely Benign | 0.61 | Ambiguous | 0.421 | Likely Benign | -1.41 | Neutral | 0.259 | Benign | 0.048 | Benign | 5.82 | Benign | 0.31 | Tolerated | 0.2209 | 0.4357 | 1 | 1 | -2.6 | 16.00 | |||||||||||||||||||||||||||||
| c.716G>C | R239T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R239T is recorded in gnomAD (ID 6‑33435567‑G‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; pathogenic predictions arise from REVEL, Rosetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). FoldX reports an uncertain effect and is therefore not considered. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenicity. Consequently, the variant is most likely pathogenic according to the available computational evidence, and this assessment does not contradict any ClinVar classification because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.170161 | Structured | 0.336504 | Uncertain | 0.854 | 0.319 | 0.000 | 6-33435567-G-C | 1 | 6.19e-7 | -14.792 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 1.96 | Ambiguous | 0.3 | 2.44 | Destabilizing | 2.20 | Destabilizing | 1.21 | Destabilizing | 0.869 | Likely Pathogenic | -5.35 | Deleterious | 0.259 | Benign | 0.064 | Benign | 5.66 | Benign | 0.01 | Affected | 3.40 | 14 | 0.1805 | 0.4414 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||
| c.1853A>C | Q618P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q618P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are SGM Consensus, ESM1b, FATHMM, PROVEAN, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta stability outputs) as uncertain. Overall, the majority of predictions lean toward a benign effect, though a subset of high‑confidence tools suggest pathogenicity. The variant’s status is not contradicted by ClinVar, which has no entry for this change. Based on the collective evidence, Q618P is most likely benign, albeit with some conflicting pathogenic signals from high‑accuracy predictors. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | -8.273 | Likely Pathogenic | 0.248 | Likely Benign | Likely Benign | -0.11 | Likely Benign | 0.1 | 3.41 | Destabilizing | 1.65 | Ambiguous | 0.23 | Likely Benign | 0.449 | Likely Benign | -3.32 | Deleterious | 0.261 | Benign | 0.246 | Benign | -1.35 | Pathogenic | 0.08 | Tolerated | 0.1990 | 0.4011 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||||
| c.1904A>G | N635S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant N635S is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6-33440956-A-G). Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and ESM1b. Predictions that are inconclusive or unavailable are FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is also inconclusive. Overall, the majority of available predictions lean toward a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.060246 | Uncertain | 0.900 | 0.252 | 0.000 | Conflicting | 4 | 6-33440956-A-G | 10 | 6.20e-6 | -9.002 | Likely Pathogenic | 0.101 | Likely Benign | Likely Benign | 0.80 | Ambiguous | 0.1 | 0.67 | Ambiguous | 0.74 | Ambiguous | 0.95 | Ambiguous | 0.104 | Likely Benign | -4.45 | Deleterious | 0.261 | Benign | 0.044 | Benign | 3.06 | Benign | 0.05 | Affected | 3.37 | 34 | 0.2816 | 0.4279 | 1 | 1 | 2.7 | -27.03 | 196.0 | 30.9 | 0.1 | 0.0 | -0.3 | 0.2 | X | Uncertain | In the WT simulations, the carboxamide side chain of Asn635, located on the outer surface of an α helix (res. Glu617-Asn635), forms hydrogen bonds with Gln631 on the same α helix and with the hydroxyl side chain of Ser590 on an opposing α helix (res. Glu582-Met603).In the variant simulations, the side chain of Ser635 is shorter than asparagine and thus prefers to hydrogen bond with the carbonyl group of Gln631 on the same helix and, to a lesser extent, with Ser590 compared to Asn635 in the WT. Ser635 forms hydrogen bonds with the backbone atoms of the same helix, which may destabilize the helix, although this is not clearly evident in the simulations. The weakening of the hydrogen bond between Ser635 and Ser590 in the variant may also weaken the tertiary structure assembly between the helices.Additionally, Asn635 is at the GTPase interface. However, the implication of the residue swap on the complex formation with the GTPase cannot be investigated using solvent-only simulations. | ||||||||||||||
| c.1042G>C | V348L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V348L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign; and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No other high‑confidence tools provide conflicting evidence. Based on the preponderance of predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.170161 | Structured | 0.346556 | Uncertain | 0.951 | 0.414 | 0.000 | -4.842 | Likely Benign | 0.270 | Likely Benign | Likely Benign | -0.58 | Ambiguous | 0.0 | 0.04 | Likely Benign | -0.27 | Likely Benign | 0.24 | Likely Benign | 0.072 | Likely Benign | -0.75 | Neutral | 0.264 | Benign | 0.030 | Benign | 1.89 | Pathogenic | 0.62 | Tolerated | 0.1088 | 0.4953 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.1042G>T | V348L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V348L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign; and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No other high‑confidence tools provide conflicting evidence. Based on the preponderance of predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.170161 | Structured | 0.346556 | Uncertain | 0.951 | 0.414 | 0.000 | -4.842 | Likely Benign | 0.270 | Likely Benign | Likely Benign | -0.58 | Ambiguous | 0.0 | 0.04 | Likely Benign | -0.27 | Likely Benign | 0.24 | Likely Benign | 0.072 | Likely Benign | -0.75 | Neutral | 0.264 | Benign | 0.030 | Benign | 1.89 | Pathogenic | 0.62 | Tolerated | 0.1088 | 0.4953 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.2732T>C | V911A 2D AIThe SynGAP1 missense variant V911A is not reported in ClinVar and is absent from gnomAD. All evaluated in silico predictors classify the change as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. Foldetta results are unavailable. Based on the consensus of all predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.724137 | Binding | 0.327 | 0.914 | 0.375 | -3.030 | Likely Benign | 0.249 | Likely Benign | Likely Benign | 0.118 | Likely Benign | 0.08 | Neutral | 0.264 | Benign | 0.102 | Benign | 2.77 | Benign | 0.41 | Tolerated | 0.3069 | 0.3336 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.2926T>C | F976L 2D  AIThe SynGAP1 missense variant F976L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), which classifies the variant as “Likely Benign.” In contrast, AlphaMissense‑Default predicts a pathogenic effect, while AlphaMissense‑Optimized is uncertain. No Foldetta (FoldX‑MD/ Rosetta) stability result is available, so it does not contribute to the assessment. Overall, the majority of high‑confidence predictors indicate a benign impact, and this is consistent with the lack of ClinVar evidence. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.975061 | Binding | 0.311 | 0.894 | 0.625 | -2.432 | Likely Benign | 0.825 | Likely Pathogenic | Ambiguous | 0.237 | Likely Benign | -0.87 | Neutral | 0.264 | Benign | 0.102 | Benign | 4.20 | Benign | 0.53 | Tolerated | 4.32 | 2 | 0.2829 | 0.3082 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||||
| c.2928T>A | F976L 2D AIThe SynGAP1 missense variant F976L is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33443480‑T‑A). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Only AlphaMissense‑Default predicts a pathogenic outcome, while AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from high‑accuracy predictors and consensus analysis indicates that F976L is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.975061 | Binding | 0.311 | 0.894 | 0.625 | 6-33443480-T-A | 2 | 1.24e-6 | -2.432 | Likely Benign | 0.825 | Likely Pathogenic | Ambiguous | 0.212 | Likely Benign | -0.87 | Neutral | 0.264 | Benign | 0.102 | Benign | 4.20 | Benign | 0.53 | Tolerated | 4.32 | 2 | 0.2829 | 0.3082 | 2 | 0 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||
| c.2928T>G | F976L 2D AIThe SynGAP1 missense variant F976L is listed in ClinVar (ID 624245.0) with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Only AlphaMissense‑Default predicts a pathogenic outcome. The high‑accuracy consensus predictor SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. AlphaMissense‑Optimized is inconclusive, and no Foldetta (FoldX‑MD/Rosetta stability) result is available. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar designation of uncertainty rather than a definitive pathogenic claim. Thus, the variant is most likely benign, and its predictions do not contradict the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.975061 | Binding | 0.311 | 0.894 | 0.625 | Uncertain | 1 | -2.432 | Likely Benign | 0.825 | Likely Pathogenic | Ambiguous | 0.212 | Likely Benign | -0.87 | Neutral | 0.264 | Benign | 0.102 | Benign | 4.20 | Benign | 0.53 | Tolerated | 4.32 | 2 | 0.2829 | 0.3082 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||
| c.3106C>G | Q1036E 2D AIThe SynGAP1 missense variant Q1036E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. Only SIFT predicts a pathogenic outcome. When predictions are grouped by consensus, the benign group contains nine tools, while the pathogenic group contains one (SIFT). High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are unavailable, so they do not influence the assessment. Overall, the evidence strongly suggests the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.987955 | Binding | 0.275 | 0.765 | 0.625 | -3.797 | Likely Benign | 0.313 | Likely Benign | Likely Benign | 0.056 | Likely Benign | -1.22 | Neutral | 0.264 | Benign | 0.062 | Benign | 2.59 | Benign | 0.03 | Affected | 0.1629 | 0.3484 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3115A>C | I1039L 2D AIThe SynGAP1 missense variant I1039L is not reported in ClinVar and is absent from gnomAD, indicating no documented clinical or population evidence. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the consensus of all available predictions strongly supports a benign classification, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.979204 | Binding | 0.292 | 0.806 | 0.625 | -2.035 | Likely Benign | 0.269 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -0.14 | Neutral | 0.264 | Benign | 0.048 | Benign | 2.82 | Benign | 0.98 | Tolerated | 0.1057 | 0.4605 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3115A>G | I1039V 2D AIThe SynGAP1 missense variant I1039V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.979204 | Binding | 0.292 | 0.806 | 0.625 | -2.455 | Likely Benign | 0.164 | Likely Benign | Likely Benign | 0.060 | Likely Benign | 0.21 | Neutral | 0.264 | Benign | 0.048 | Benign | 2.76 | Benign | 0.41 | Tolerated | 0.1405 | 0.4112 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3167G>C | G1056A 2D AIThe SynGAP1 missense variant G1056A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign consensus (2 benign vs. 1 pathogenic, with one uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988291 | Disordered | 0.868632 | Binding | 0.402 | 0.935 | 0.875 | -7.458 | In-Between | 0.086 | Likely Benign | Likely Benign | 0.325 | Likely Benign | -0.25 | Neutral | 0.264 | Benign | 0.097 | Benign | 1.85 | Pathogenic | 0.49 | Tolerated | 0.3325 | 0.4957 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3220C>G | Q1074E 2D AIThe SynGAP1 missense variant Q1074E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions strongly suggests that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.987006 | Binding | 0.339 | 0.897 | 0.750 | -2.815 | Likely Benign | 0.419 | Ambiguous | Likely Benign | 0.096 | Likely Benign | -0.79 | Neutral | 0.264 | Benign | 0.103 | Benign | 2.79 | Benign | 0.24 | Tolerated | 0.1386 | 0.2856 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3359G>C | G1120A 2D AIThe SynGAP1 missense variant G1120A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; ESM1b is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign likelihood. Foldetta results are unavailable. Overall, the consensus of available predictions indicates that G1120A is most likely benign, and this conclusion does not contradict any ClinVar status because none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.871313 | Disordered | 0.804931 | Binding | 0.335 | 0.925 | 0.875 | -7.192 | In-Between | 0.082 | Likely Benign | Likely Benign | 0.243 | Likely Benign | -0.39 | Neutral | 0.264 | Benign | 0.139 | Benign | 3.64 | Benign | 0.28 | Tolerated | 0.3435 | 0.4944 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3368G>C | G1123A 2D AIThe SynGAP1 missense variant G1123A is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. No pathogenic predictions are present among the evaluated methods. High‑accuracy tools confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.837511 | Disordered | 0.827246 | Binding | 0.346 | 0.934 | 0.875 | -6.720 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.240 | Likely Benign | -0.55 | Neutral | 0.264 | Benign | 0.103 | Benign | 4.38 | Benign | 1.00 | Tolerated | 0.3544 | 0.4944 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3853C>G | P1285A 2D AIThe SynGAP1 missense variant P1285A is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.821643 | Binding | 0.557 | 0.759 | 0.750 | -3.902 | Likely Benign | 0.060 | Likely Benign | Likely Benign | 0.052 | Likely Benign | -0.52 | Neutral | 0.264 | Benign | 0.070 | Benign | 4.27 | Benign | 0.71 | Tolerated | 0.2944 | 0.2974 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.1585A>T | I529F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I529F is not reported in ClinVar and is present in gnomAD (variant ID 6‑33438828‑A‑T). Consensus from most in‑silico predictors classifies the change as benign: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic effect. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus is Likely Benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports a benign effect. Overall, the preponderance of evidence indicates that I529F is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.318242 | Structured | 0.019545 | Uncertain | 0.901 | 0.403 | 0.000 | 6-33438828-A-T | 4 | 2.48e-6 | -5.669 | Likely Benign | 0.235 | Likely Benign | Likely Benign | -0.21 | Likely Benign | 0.0 | 0.06 | Likely Benign | -0.08 | Likely Benign | 0.06 | Likely Benign | 0.320 | Likely Benign | -0.97 | Neutral | 0.266 | Benign | 0.054 | Benign | -1.30 | Pathogenic | 0.23 | Tolerated | 3.37 | 35 | 0.0522 | 0.2713 | 0 | 1 | -1.7 | 34.02 | ||||||||||||||||||||||||
| c.1019C>G | A340G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A340G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only FATHMM predicts a pathogenic outcome. Stability‑based methods (FoldX, Rosetta, Foldetta) are inconclusive, so they provide no evidence for or against pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as likely benign, and Foldetta as uncertain. Overall, the majority of reliable predictors indicate a benign effect, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.390993 | Structured | 0.410781 | Uncertain | 0.558 | 0.485 | 0.250 | -3.763 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.66 | Ambiguous | 0.2 | 1.44 | Ambiguous | 1.05 | Ambiguous | 0.50 | Likely Benign | 0.044 | Likely Benign | -0.34 | Neutral | 0.267 | Benign | 0.127 | Benign | 1.92 | Pathogenic | 0.42 | Tolerated | 0.2355 | 0.4470 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||
| c.104T>C | V35A 2D AIThe SynGAP1 missense variant V35A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools, polyPhen‑2 HumVar and SIFT, predict pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.613573 | Disordered | 0.434838 | Uncertain | 0.360 | 0.851 | 0.375 | -3.552 | Likely Benign | 0.120 | Likely Benign | Likely Benign | 0.098 | Likely Benign | -0.05 | Neutral | 0.267 | Benign | 0.481 | Possibly Damaging | 4.25 | Benign | 0.00 | Affected | 0.2340 | 0.2144 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.183G>C | E61D 2D AIThe SynGAP1 missense variant E61D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumVar and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. The variant’s predicted benign status does not contradict ClinVar, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | -4.394 | Likely Benign | 0.231 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -0.29 | Neutral | 0.267 | Benign | 0.585 | Possibly Damaging | 4.11 | Benign | 0.00 | Affected | 0.1895 | 0.3851 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.183G>T | E61D 2D AIThe SynGAP1 missense variant E61D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumVar and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. The variant’s predicted benign status does not contradict ClinVar, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | -4.394 | Likely Benign | 0.231 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -0.29 | Neutral | 0.267 | Benign | 0.585 | Possibly Damaging | 4.11 | Benign | 0.00 | Affected | 0.1895 | 0.3851 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.189G>C | E63D 2D AIThe SynGAP1 missense variant E63D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus also reports it as likely benign. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a benign effect, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.474807 | Uncertain | 0.494 | 0.739 | 0.125 | -3.821 | Likely Benign | 0.594 | Likely Pathogenic | Likely Benign | 0.066 | Likely Benign | -0.83 | Neutral | 0.267 | Benign | 0.585 | Possibly Damaging | 3.98 | Benign | 0.00 | Affected | 0.1524 | 0.4130 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.189G>T | E63D 2D AIThe SynGAP1 missense variant E63D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority of the high‑accuracy tools) also indicates benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective evidence points to a benign effect for E63D, and this conclusion does not conflict with ClinVar, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.474807 | Uncertain | 0.494 | 0.739 | 0.125 | -3.821 | Likely Benign | 0.594 | Likely Pathogenic | Likely Benign | 0.066 | Likely Benign | -0.83 | Neutral | 0.267 | Benign | 0.585 | Possibly Damaging | 3.98 | Benign | 0.00 | Affected | 0.1524 | 0.4130 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.19T>C | S7P 2D AIThe SynGAP1 missense variant S7P is reported in gnomAD (ID 6‑33420283‑T‑C) but has no ClinVar entry. In silico prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority‑vote) is benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools, including the high‑accuracy methods, points to a benign effect. This prediction is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.548467 | Binding | 0.386 | 0.922 | 0.750 | 6-33420283-T-C | -3.798 | Likely Benign | 0.111 | Likely Benign | Likely Benign | 0.210 | Likely Benign | -0.23 | Neutral | 0.267 | Benign | 0.029 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2474 | 0.4925 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||
| c.205A>C | I69L 2D AIThe SynGAP1 missense variant I69L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic call comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign; Foldetta results are not available. Overall, the majority of evidence supports a benign impact for I69L, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.575842 | Disordered | 0.466129 | Uncertain | 0.437 | 0.786 | 0.375 | -2.065 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.067 | Likely Benign | -0.32 | Neutral | 0.267 | Benign | 0.141 | Benign | 4.27 | Benign | 0.00 | Affected | 0.0636 | 0.3447 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.205A>G | I69V 2D AIThe SynGAP1 missense variant I69V is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic call comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” status. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign; Foldetta results are not available. Overall, the consensus of the available predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.575842 | Disordered | 0.466129 | Uncertain | 0.437 | 0.786 | 0.375 | -2.809 | Likely Benign | 0.176 | Likely Benign | Likely Benign | 0.080 | Likely Benign | 0.06 | Neutral | 0.267 | Benign | 0.141 | Benign | 4.24 | Benign | 0.00 | Affected | 0.0972 | 0.3142 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2270G>C | G757A 2D AIThe SynGAP1 missense change G757A is catalogued in ClinVar (ID 3635272.0) with an uncertain significance designation and is not reported in gnomAD. Functional prediction algorithms uniformly classify the variant as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores. No tool in the dataset predicts pathogenicity. High‑accuracy consensus methods corroborate this view: the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect, and AlphaMissense‑Optimized also predicts benign. The Foldetta stability assessment is unavailable for this variant. Taken together, the evidence overwhelmingly supports a benign interpretation, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.830995 | Binding | 0.310 | 0.869 | 0.375 | Uncertain | 1 | -2.626 | Likely Benign | 0.091 | Likely Benign | Likely Benign | 0.066 | Likely Benign | -0.45 | Neutral | 0.267 | Benign | 0.127 | Benign | 2.73 | Benign | 0.35 | Tolerated | 0.3690 | 0.3842 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||
| c.2275A>G | M759V 2D AIThe SynGAP1 missense variant M759V is catalogued in gnomAD (ID 6‑33441740‑A‑G) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available for this variant. Taken together, the computational evidence overwhelmingly supports a benign effect, and this conclusion does not conflict with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.525368 | Disordered | 0.879389 | Binding | 0.299 | 0.864 | 0.375 | 6-33441740-A-G | 20 | 1.24e-5 | -3.985 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.030 | Likely Benign | -1.00 | Neutral | 0.267 | Benign | 0.127 | Benign | 2.67 | Benign | 0.23 | Tolerated | 3.99 | 5 | 0.2926 | 0.3041 | 1 | 2 | 2.3 | -32.06 | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||||||||||||||||||
| c.256G>A | V86I 2D  AIThe SynGAP1 missense variant V86I is listed in ClinVar (ID 588267.0) with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the variant as benign or likely benign. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is “Likely Benign.” No Foldetta (FoldX‑MD/Rosetta stability) result is available, so it does not influence the assessment. Overall, the majority of predictions indicate that V86I is most likely benign, which is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.552911 | Binding | 0.295 | 0.887 | 0.500 | Uncertain | 1 | -4.726 | Likely Benign | 0.338 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -0.31 | Neutral | 0.267 | Benign | 0.097 | Benign | 3.94 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0887 | 0.4417 | 4 | 3 | 0.3 | 14.03 | |||||||||||||||||||||||||||||||||||
| c.256G>C | V86L 2D AIThe SynGAP1 missense variant V86L is reported in ClinVar as “not listed” and is present in the gnomAD database (ID 6‑33425864‑G‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains benign; Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a benign interpretation, and this conclusion does not contradict the ClinVar status, which currently contains no pathogenic assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.552911 | Binding | 0.295 | 0.887 | 0.500 | 6-33425864-G-C | 1 | 6.20e-7 | -3.658 | Likely Benign | 0.965 | Likely Pathogenic | Likely Pathogenic | 0.081 | Likely Benign | -0.84 | Neutral | 0.267 | Benign | 0.097 | Benign | 3.85 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1121 | 0.5386 | 1 | 2 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.257T>C | V86A 2D AIThe SynGAP1 missense variant V86A has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and the SGM‑Consensus (Likely Benign). Pathogenic predictions arise from SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further highlight a split: AlphaMissense‑Optimized classifies the variant as pathogenic, whereas the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates it is likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of tools (six benign vs. three pathogenic) suggest the variant is most likely benign, and this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.552911 | Binding | 0.295 | 0.887 | 0.500 | -3.022 | Likely Benign | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.059 | Likely Benign | -1.35 | Neutral | 0.267 | Benign | 0.153 | Benign | 3.80 | Benign | 0.00 | Affected | 0.3205 | 0.2723 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.326G>T | S109I 2D AIThe SynGAP1 missense variant S109I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta data are unavailable. Overall, the majority of evidence (five benign vs. three pathogenic predictions) points toward a benign impact. This conclusion does not contradict ClinVar, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.669335 | Binding | 0.328 | 0.864 | 0.750 | -5.195 | Likely Benign | 0.927 | Likely Pathogenic | Ambiguous | 0.200 | Likely Benign | -2.56 | Deleterious | 0.267 | Benign | 0.039 | Benign | 3.47 | Benign | 0.00 | Affected | 0.0910 | 0.4930 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.337G>A | G113R 2D AIThe SynGAP1 missense variant G113R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default, while AlphaMissense‑Optimized returns an uncertain result. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.639486 | Binding | 0.350 | 0.870 | 0.750 | -3.904 | Likely Benign | 0.791 | Likely Pathogenic | Ambiguous | 0.087 | Likely Benign | -1.50 | Neutral | 0.267 | Benign | 0.080 | Benign | 4.16 | Benign | 0.03 | Affected | 0.1025 | 0.4138 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||||||
| c.337G>C | G113R 2D AIThe SynGAP1 missense variant G113R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM all classify it as benign. In contrast, SIFT and AlphaMissense‑Default predict a pathogenic impact. The high‑accuracy AlphaMissense‑Optimized score is uncertain, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as likely benign. Protein‑folding stability analysis via Foldetta is unavailable for this residue. Overall, the preponderance of evidence points to a benign effect, and this assessment does not conflict with the absence of a ClinVar classification. Thus, the variant is most likely benign, and this conclusion is consistent with the lack of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.639486 | Binding | 0.350 | 0.870 | 0.750 | -3.904 | Likely Benign | 0.791 | Likely Pathogenic | Ambiguous | 0.088 | Likely Benign | -1.50 | Neutral | 0.267 | Benign | 0.080 | Benign | 4.16 | Benign | 0.03 | Affected | 0.1025 | 0.4138 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||||||
| c.542A>T | H181L 2D AIThe SynGAP1 H181L variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans pathogenic (2 pathogenic vs 1 benign). Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the majority of standard predictors classify the variant as benign, and there is no ClinVar entry to contradict this assessment. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.439530 | Uncertain | 0.294 | 0.616 | 0.500 | -11.014 | Likely Pathogenic | 0.561 | Ambiguous | Likely Benign | 0.212 | Likely Benign | -3.51 | Deleterious | 0.267 | Benign | 0.039 | Benign | 4.17 | Benign | 0.04 | Affected | 0.0734 | 0.4395 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.668C>A | T223K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T223K missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are REVEL, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The remaining tools, premPS and AlphaMissense‑Optimized, are uncertain and therefore treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Benign. Overall, the balance of evidence leans toward a benign impact, with no contradiction to the ClinVar status (which is currently unreported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.070400 | Structured | 0.382605 | Uncertain | 0.867 | 0.316 | 0.125 | -12.084 | Likely Pathogenic | 0.853 | Likely Pathogenic | Ambiguous | -0.30 | Likely Benign | 0.1 | 0.42 | Likely Benign | 0.06 | Likely Benign | 0.93 | Ambiguous | 0.810 | Likely Pathogenic | -4.60 | Deleterious | 0.267 | Benign | 0.086 | Benign | 5.78 | Benign | 0.01 | Affected | 0.0793 | 0.2462 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||
| c.2221C>T | P741S 2D AIThe SynGAP1 missense variant P741S is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33441686‑C‑T). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. Grouping by consensus, the benign‑predicting tools outnumber the pathogenic one. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” No Foldetta stability data are available, so it does not influence the conclusion. Overall, the computational evidence indicates that the variant is most likely benign, and this assessment does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.885302 | Disordered | 0.493550 | Uncertain | 0.354 | 0.859 | 0.875 | Uncertain | 2 | 6-33441686-C-T | 3 | 1.86e-6 | -3.700 | Likely Benign | 0.063 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -0.27 | Neutral | 0.270 | Benign | 0.136 | Benign | 2.92 | Benign | 0.00 | Affected | 4.32 | 2 | 0.2888 | 0.4278 | 1 | -1 | 0.8 | -10.04 | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||||||||||||||||
| c.2425A>G | S809G 2D AIThe SynGAP1 missense variant S809G is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is also benign. Foldetta results are not available, so they do not influence the conclusion. Overall, the preponderance of evidence indicates that S809G is most likely benign, and this assessment does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.626927 | Disordered | 0.853218 | Binding | 0.330 | 0.907 | 0.500 | -6.100 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.085 | Likely Benign | -1.80 | Neutral | 0.270 | Benign | 0.136 | Benign | 2.61 | Benign | 0.02 | Affected | 0.2743 | 0.5369 | 1 | 0 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||||||||||
| c.3704T>A | M1235K 2D AIThe SynGAP1 missense variant M1235K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of tools, including the high‑accuracy methods, supports a benign classification. This prediction is consistent with the lack of ClinVar evidence and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.690604 | Disordered | 0.577958 | Binding | 0.872 | 0.532 | 0.125 | -6.348 | Likely Benign | 0.269 | Likely Benign | Likely Benign | 0.109 | Likely Benign | -2.37 | Neutral | 0.270 | Benign | 0.089 | Benign | 2.66 | Benign | 0.00 | Affected | 0.1282 | 0.0656 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||||||||||
| c.3704T>G | M1235R 2D AIThe SynGAP1 missense variant M1235R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.690604 | Disordered | 0.577958 | Binding | 0.872 | 0.532 | 0.125 | -5.410 | Likely Benign | 0.246 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -2.37 | Neutral | 0.270 | Benign | 0.089 | Benign | 2.64 | Benign | 0.00 | Affected | 0.1505 | 0.0757 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||||||||||
| c.3715G>A | A1239T 2D AIThe SynGAP1 missense variant A1239T is not reported in ClinVar and has no gnomAD entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the consensus score from SGM‑Consensus is “Likely Benign.” In contrast, two sequence‑based predictors flag it as pathogenic: SIFT and FATHMM. When considering the high‑accuracy subset, AlphaMissense‑Optimized remains benign and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a benign outcome; Foldetta data are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -3.570 | Likely Benign | 0.070 | Likely Benign | Likely Benign | 0.030 | Likely Benign | -1.62 | Neutral | 0.270 | Benign | 0.136 | Benign | 2.37 | Pathogenic | 0.00 | Affected | 0.0957 | 0.5384 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.3716C>A | A1239D 2D AIThe SynGAP1 missense variant A1239D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes). Foldetta results are unavailable. Overall, the majority of evidence (six benign vs. three pathogenic predictions) supports a benign classification. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -6.177 | Likely Benign | 0.326 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -2.60 | Deleterious | 0.270 | Benign | 0.136 | Benign | 2.36 | Pathogenic | 0.00 | Affected | 0.1521 | 0.2062 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.965C>A | A322D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A322D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The remaining tools—FoldX, Rosetta, ESM1b, and AlphaMissense‑Default—return uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta indicates no significant stability change (uncertain). Overall, the preponderance of evidence (seven benign predictions versus one pathogenic) suggests the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.175930 | Structured | 0.425745 | Uncertain | 0.938 | 0.334 | 0.000 | -7.184 | In-Between | 0.411 | Ambiguous | Likely Benign | 0.65 | Ambiguous | 0.1 | 1.35 | Ambiguous | 1.00 | Ambiguous | 0.34 | Likely Benign | 0.246 | Likely Benign | -0.95 | Neutral | 0.270 | Benign | 0.136 | Benign | 1.96 | Pathogenic | 0.32 | Tolerated | 0.1646 | 0.1660 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.965C>T | A322V 2D AIThe SynGAP1 missense variant A322V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign; and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No predictions or stability results are missing or inconclusive. Based on the overwhelming consensus of the available tools, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.175930 | Structured | 0.425745 | Uncertain | 0.938 | 0.334 | 0.000 | -4.905 | Likely Benign | 0.121 | Likely Benign | Likely Benign | 0.02 | Likely Benign | 1.0 | -0.42 | Likely Benign | -0.20 | Likely Benign | 0.19 | Likely Benign | 0.052 | Likely Benign | -1.63 | Neutral | 0.270 | Benign | 0.136 | Benign | 1.96 | Pathogenic | 0.22 | Tolerated | 0.1279 | 0.6126 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.992C>T | S331L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S331L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate benign or likely benign. Only FATHMM predicts pathogenicity, while FoldX, Rosetta, and Foldetta provide uncertain or inconclusive stability results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta as uncertain. Overall, the consensus of the majority of tools supports a benign classification, and this is consistent with the lack of ClinVar evidence; there is no contradiction with ClinVar status. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.433034 | Structured | 0.346458 | Uncertain | 0.658 | 0.475 | 0.250 | -6.570 | Likely Benign | 0.219 | Likely Benign | Likely Benign | 0.54 | Ambiguous | 0.0 | 1.06 | Ambiguous | 0.80 | Ambiguous | 0.07 | Likely Benign | 0.090 | Likely Benign | -1.72 | Neutral | 0.270 | Benign | 0.136 | Benign | 1.87 | Pathogenic | 0.58 | Tolerated | 0.0953 | 0.4124 | -3 | -2 | 4.6 | 26.08 | |||||||||||||||||||||||||||||
| c.3178G>A | G1060S 2D AIThe SynGAP1 missense variant G1060S is listed in ClinVar with an uncertain significance (ClinVar ID 1512003.0) and is present in gnomAD (variant ID 6‑33443730‑G‑A). All evaluated in‑silico predictors classify the change as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments reinforce this benign view: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.979242 | Disordered | 0.913048 | Binding | 0.407 | 0.928 | 0.875 | Uncertain | 1 | 6-33443730-G-A | -4.759 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.376 | Likely Benign | -0.08 | Neutral | 0.271 | Benign | 0.054 | Benign | 2.69 | Benign | 0.49 | Tolerated | 4.32 | 2 | 0.2468 | 0.5311 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||||
| c.3199C>T | P1067S 2D AIThe SynGAP1 missense variant P1067S is not reported in ClinVar and is absent from gnomAD, indicating no documented clinical or population evidence. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Based on the collective predictions, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.966441 | Disordered | 0.975099 | Binding | 0.459 | 0.907 | 0.875 | -4.673 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.079 | Likely Benign | -1.43 | Neutral | 0.271 | Benign | 0.054 | Benign | 2.91 | Benign | 0.48 | Tolerated | 0.3086 | 0.5753 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.395T>A | F132Y 2D  AIThe SynGAP1 missense variant F132Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of available predictions (five benign vs three pathogenic) suggest a benign impact. This consensus does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -8.220 | Likely Pathogenic | 0.839 | Likely Pathogenic | Ambiguous | 0.165 | Likely Benign | -1.98 | Neutral | 0.272 | Benign | 0.126 | Benign | 3.39 | Benign | 0.00 | Affected | 0.1635 | 0.1526 | 7 | 3 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.422T>C | I141T 2D AIThe SynGAP1 missense variant I141T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its contribution is unavailable. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.465241 | Structured | 0.577021 | Binding | 0.367 | 0.877 | 0.500 | -10.580 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.251 | Likely Benign | -2.81 | Deleterious | 0.272 | Benign | 0.167 | Benign | 3.57 | Benign | 0.00 | Affected | 0.1165 | 0.1214 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||||||||||||
| c.493A>G | S165G 2D AIThe SynGAP1 missense variant S165G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable. Overall, the preponderance of evidence points to the variant being most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -7.691 | In-Between | 0.223 | Likely Benign | Likely Benign | 0.182 | Likely Benign | -1.17 | Neutral | 0.272 | Benign | 0.086 | Benign | 3.99 | Benign | 0.00 | Affected | 0.2622 | 0.4441 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.164A>T | Q55L 2D AIThe SynGAP1 missense variant Q55L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign based on current predictive data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.332115 | Structured | 0.470108 | Uncertain | 0.461 | 0.657 | 0.000 | -5.823 | Likely Benign | 0.844 | Likely Pathogenic | Ambiguous | 0.173 | Likely Benign | -1.99 | Neutral | 0.273 | Benign | 0.275 | Benign | 3.83 | Benign | 0.00 | Affected | 0.0776 | 0.5982 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.193C>A | H65N 2D AIThe SynGAP1 missense variant H65N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are AlphaMissense‑Default and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely benign; a Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | -2.788 | Likely Benign | 0.771 | Likely Pathogenic | Likely Benign | 0.038 | Likely Benign | -1.42 | Neutral | 0.273 | Benign | 0.107 | Benign | 4.18 | Benign | 0.00 | Affected | 0.1280 | 0.2200 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.193C>T | H65Y 2D AIThe SynGAP1 missense variant H65Y has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic designation and the lack of population frequency data. The variant is most likely benign based on predictions, and this does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | -3.644 | Likely Benign | 0.852 | Likely Pathogenic | Ambiguous | 0.037 | Likely Benign | -1.11 | Neutral | 0.273 | Benign | 0.152 | Benign | 4.15 | Benign | 0.00 | Affected | 0.0788 | 0.3628 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||||||
| c.62T>A | F21Y 2D AIThe SynGAP1 missense variant F21Y is listed in gnomAD (ID 6‑33420326‑T‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus also indicates a likely benign classification. No Foldetta (FoldX‑MD/Rosetta stability) result is available, so it does not influence the assessment. Overall, the preponderance of evidence points to a benign effect for F21Y, and this conclusion is not contradicted by any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.534167 | Disordered | 0.443284 | Uncertain | 0.369 | 0.897 | 0.500 | 6-33420326-T-A | -3.712 | Likely Benign | 0.352 | Ambiguous | Likely Benign | 0.088 | Likely Benign | -0.23 | Neutral | 0.273 | Benign | 0.153 | Benign | 4.15 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1698 | 0.2608 | 3 | 7 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||||||||
| c.88C>A | H30N 2D AIThe SynGAP1 missense variant H30N is not reported in ClinVar and is absent from gnomAD. Consensus among most in silico predictors is benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a benign effect, while only SIFT predicts pathogenicity. Grouping the tools, the benign‑predicting set (nine tools) overwhelmingly outweighs the single pathogenic prediction. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. No Foldetta stability analysis is available, so it does not influence the conclusion. Overall, the computational evidence strongly supports a benign classification, and this is consistent with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | -3.096 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.052 | Likely Benign | -1.91 | Neutral | 0.273 | Benign | 0.380 | Benign | 3.92 | Benign | 0.00 | Affected | 0.2699 | 0.3818 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.88C>T | H30Y 2D AIThe SynGAP1 H30Y missense variant (ClinVar ID 972248.0) is listed as “Uncertain” and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumVar and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as “Likely Benign”; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | Uncertain | 1 | -3.047 | Likely Benign | 0.115 | Likely Benign | Likely Benign | 0.082 | Likely Benign | -1.84 | Neutral | 0.273 | Benign | 0.478 | Possibly Damaging | 3.99 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1572 | 0.4856 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||
| c.1169G>A | G390E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G390E is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that classify the variant as benign include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that predict pathogenicity are REVEL, FoldX, Rosetta, Foldetta, SIFT, FATHMM, and AlphaMissense‑Default. A high‑accuracy assessment shows AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of predictions support a pathogenic effect, and this aligns with the ClinVar designation of uncertain significance rather than contradicting it. Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | Uncertain | 1 | -7.913 | In-Between | 0.646 | Likely Pathogenic | Likely Benign | 2.61 | Destabilizing | 0.9 | 4.28 | Destabilizing | 3.45 | Destabilizing | 0.47 | Likely Benign | 0.575 | Likely Pathogenic | -0.87 | Neutral | 0.276 | Benign | 0.045 | Benign | 1.32 | Pathogenic | 0.05 | Affected | 4.32 | 8 | 0.1595 | 0.4309 | 0 | -2 | -3.1 | 72.06 | 241.5 | -108.4 | 0.6 | 0.5 | -0.1 | 0.1 | Uncertain | Gly390 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364 and res. Ala399-Ile411). The Ω loop is assumed to directly interact with the membrane, and it is observed to move arbitrarily throughout the WT solvent simulations. This loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play significant roles in protein functions that require flexibility, and so they are rich in glycine residues, prolines, and to a lesser extent, small hydrophilic residues to ensure maximum flexibility. Thus, the variant’s Glu390 may not be as well tolerated in the Ω loop. Additionally, the carboxylate group of Glu390 occasionally forms H-bonds with other loop residues in the variant simulations. The interaction between the acidic carboxylate side chain and the acidic membrane lipids may further influence the SynGAP-membrane complex. However, since the effects on the Gly-rich Ω loop dynamics can only be well studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||||||
| c.1174A>G | K392E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and the SGM‑Consensus score. Tools that predict a pathogenic effect are REVEL, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, while the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) and Foldetta (combining FoldX‑MD and Rosetta) both indicate a benign outcome. Overall, the majority of evidence supports a benign classification, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.392 | Likely Benign | 0.850 | Likely Pathogenic | Ambiguous | 0.09 | Likely Benign | 0.0 | -0.04 | Likely Benign | 0.03 | Likely Benign | 0.28 | Likely Benign | 0.529 | Likely Pathogenic | -1.92 | Neutral | 0.276 | Benign | 0.083 | Benign | 4.60 | Benign | 0.02 | Affected | 0.4812 | 0.1916 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1175A>C | K392T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, PROVEAN, and SIFT. Two tools—FoldX and AlphaMissense‑Default—return uncertain results and are treated as unavailable. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign; and Foldetta, a protein‑folding stability method, also predicts benign. No ClinVar entry exists to contradict these predictions. Based on the collective evidence, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -3.224 | Likely Benign | 0.481 | Ambiguous | Likely Benign | 0.52 | Ambiguous | 0.1 | -0.29 | Likely Benign | 0.12 | Likely Benign | 0.05 | Likely Benign | 0.512 | Likely Pathogenic | -3.14 | Deleterious | 0.276 | Benign | 0.045 | Benign | 4.60 | Benign | 0.02 | Affected | 0.2807 | 0.4053 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||
| c.1176G>C | K392N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K392N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts benign. Overall, the majority of evidence (9 benign vs 3 pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.136 | Likely Benign | 0.780 | Likely Pathogenic | Likely Benign | 0.24 | Likely Benign | 0.1 | -0.01 | Likely Benign | 0.12 | Likely Benign | 0.20 | Likely Benign | 0.229 | Likely Benign | -2.61 | Deleterious | 0.276 | Benign | 0.083 | Benign | 4.60 | Benign | 0.02 | Affected | 0.4586 | 0.2454 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.1176G>T | K392N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split and is therefore treated as unavailable. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the majority of evidence (nine benign vs. three pathogenic predictions) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.136 | Likely Benign | 0.780 | Likely Pathogenic | Likely Benign | 0.24 | Likely Benign | 0.1 | -0.01 | Likely Benign | 0.12 | Likely Benign | 0.20 | Likely Benign | 0.229 | Likely Benign | -2.61 | Deleterious | 0.276 | Benign | 0.083 | Benign | 4.60 | Benign | 0.02 | Affected | 0.4586 | 0.2454 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.1189T>A | C397S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, FoldX, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign, while Rosetta remains uncertain. When predictions are grouped, the benign category contains all available evidence and the pathogenic category is empty. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports a benign effect. Consequently, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -2.324 | Likely Benign | 0.178 | Likely Benign | Likely Benign | 0.37 | Likely Benign | 0.1 | 0.59 | Ambiguous | 0.48 | Likely Benign | 0.43 | Likely Benign | 0.174 | Likely Benign | -0.01 | Neutral | 0.276 | Benign | 0.066 | Benign | 4.68 | Benign | 0.53 | Tolerated | 0.5221 | 0.2474 | Weaken | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||
| c.1189T>G | C397G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that reach consensus all indicate a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the variant as benign. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies it as “Likely Benign.” No tool predicts pathogenicity. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) is inconclusive. No pathogenic predictions are present. Therefore, based on the available computational evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | 1.244 | Likely Benign | 0.096 | Likely Benign | Likely Benign | 0.68 | Ambiguous | 0.2 | 1.42 | Ambiguous | 1.05 | Ambiguous | 0.04 | Likely Benign | 0.272 | Likely Benign | 1.51 | Neutral | 0.276 | Benign | 0.066 | Benign | 5.93 | Benign | 0.60 | Tolerated | 0.3678 | 0.3729 | -3 | -3 | -2.9 | -46.09 | |||||||||||||||||||||||||||||
| c.1190G>C | C397S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). In silico prediction tools that assess pathogenicity largely agree on a benign outcome: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. No tool predicts pathogenicity; Rosetta’s assessment is uncertain and therefore treated as unavailable. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is benign. Consequently, the variant is most likely benign based on the collective predictions, and this conclusion does not contradict any ClinVar status (none is assigned). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -2.324 | Likely Benign | 0.178 | Likely Benign | Likely Benign | 0.37 | Likely Benign | 0.1 | 0.59 | Ambiguous | 0.48 | Likely Benign | 0.43 | Likely Benign | 0.253 | Likely Benign | -0.01 | Neutral | 0.276 | Benign | 0.066 | Benign | 4.68 | Benign | 0.53 | Tolerated | 0.5221 | 0.2474 | Weaken | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||
| c.472C>A | Q158K 2D AIThe SynGAP1 missense variant Q158K is not reported in ClinVar and has no entry in gnomAD, indicating it is not catalogued in these databases. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, SGM‑Consensus indicates likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the computational evidence strongly suggests the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.527565 | Binding | 0.286 | 0.750 | 0.375 | -6.795 | Likely Benign | 0.481 | Ambiguous | Likely Benign | 0.069 | Likely Benign | -0.86 | Neutral | 0.276 | Benign | 0.121 | Benign | 4.20 | Benign | 0.15 | Tolerated | 0.1851 | 0.3869 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.473A>G | Q158R 2D AIThe SynGAP1 missense variant Q158R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also reports a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. The predictions do not contradict ClinVar status, as ClinVar contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.527565 | Binding | 0.286 | 0.750 | 0.375 | -6.873 | Likely Benign | 0.438 | Ambiguous | Likely Benign | 0.090 | Likely Benign | -0.85 | Neutral | 0.276 | Benign | 0.121 | Benign | 4.14 | Benign | 0.11 | Tolerated | 0.1527 | 0.1498 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.515G>T | R172L 2D  AIThe SynGAP1 missense variant R172L is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. Grouping by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Taken together, the majority of evidence, including the SGM Consensus, points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.525368 | Disordered | 0.491688 | Uncertain | 0.411 | 0.651 | 0.375 | -8.201 | Likely Pathogenic | 0.797 | Likely Pathogenic | Ambiguous | 0.131 | Likely Benign | -3.09 | Deleterious | 0.276 | Benign | 0.103 | Benign | 3.99 | Benign | 0.02 | Affected | 0.1423 | 0.4729 | -3 | -2 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||||
| c.595A>G | N199D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N199D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that reach a consensus classify the variant as benign: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. The only inconclusive results come from ESM1b and Rosetta, which are treated as unavailable. No tool predicts pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts a benign outcome. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.390993 | Structured | 0.431347 | Uncertain | 0.571 | 0.473 | 0.125 | -7.874 | In-Between | 0.338 | Likely Benign | Likely Benign | -0.14 | Likely Benign | 0.1 | -0.67 | Ambiguous | -0.41 | Likely Benign | 0.13 | Likely Benign | 0.028 | Likely Benign | -1.41 | Neutral | 0.276 | Benign | 0.062 | Benign | 4.22 | Benign | 0.35 | Tolerated | 0.1516 | 0.3514 | 2 | 1 | 0.0 | 0.98 | |||||||||||||||||||||||||||||
| c.597C>A | N199K 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant N199K is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized all classify the substitution as benign. Only ESM1b and AlphaMissense‑Default predict pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the preponderance of evidence supports a benign impact, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.390993 | Structured | 0.431347 | Uncertain | 0.571 | 0.473 | 0.125 | Uncertain | 1 | -8.198 | Likely Pathogenic | 0.686 | Likely Pathogenic | Likely Benign | -0.19 | Likely Benign | 0.1 | 0.03 | Likely Benign | -0.08 | Likely Benign | 0.33 | Likely Benign | 0.024 | Likely Benign | -1.48 | Neutral | 0.276 | Benign | 0.083 | Benign | 4.27 | Benign | 0.13 | Tolerated | 3.47 | 9 | 0.1399 | 0.5218 | 1 | 0 | -0.4 | 14.07 | 207.8 | 21.5 | -0.1 | 1.5 | 0.1 | 0.0 | X | Uncertain | Asn199, located in the N-terminal loop before the first anti-parallel β sheet strand (res. Ile205-Pro208), is replaced by a positively charged lysine. On the protein surface, both the carboxamide group of Asn199 and the amino group of Lys199 side chains can form hydrogen bonds with the backbone carbonyl groups of residues (e.g., Ala249) at the end of an α helix (res. Ala236-Lys251). However, since the model ends abruptly at the N-terminus, no definite conclusions can be drawn from the simulations. | |||||||||||||||||
| c.597C>G | N199K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N199K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a benign impact. Overall, the majority of evidence (10 benign vs. 2 pathogenic) supports a benign classification. This consensus does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.390993 | Structured | 0.431347 | Uncertain | 0.571 | 0.473 | 0.125 | -8.198 | Likely Pathogenic | 0.686 | Likely Pathogenic | Likely Benign | -0.19 | Likely Benign | 0.1 | 0.03 | Likely Benign | -0.08 | Likely Benign | 0.33 | Likely Benign | 0.024 | Likely Benign | -1.48 | Neutral | 0.276 | Benign | 0.083 | Benign | 4.27 | Benign | 0.13 | Tolerated | 3.47 | 9 | 0.1399 | 0.5218 | 1 | 0 | -0.4 | 14.07 | 207.8 | 21.5 | -0.1 | 1.5 | 0.1 | 0.0 | X | Uncertain | Asn199, located in the N-terminal loop before the first anti-parallel β sheet strand (res. Ile205-Pro208), is replaced by a positively charged lysine. On the protein surface, both the carboxamide group of Asn199 and the amino group of Lys199 side chains can form hydrogen bonds with the backbone carbonyl groups of residues (e.g., Ala249) at the end of an α helix (res. Ala236-Lys251). However, since the model ends abruptly at the N-terminus, no definite conclusions can be drawn from the simulations. | |||||||||||||||||||
| c.700C>G | R234G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 R234G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that agree on a pathogenic effect are REVEL, PROVEAN, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b, and AlphaMissense‑Optimized) give uncertain or inconclusive results. High‑accuracy methods show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of individual predictors lean toward pathogenicity, and the high‑accuracy consensus also supports a pathogenic classification. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.239899 | Structured | 0.311558 | Uncertain | 0.804 | 0.322 | 0.000 | -7.163 | In-Between | 0.923 | Likely Pathogenic | Ambiguous | 1.50 | Ambiguous | 0.2 | 0.88 | Ambiguous | 1.19 | Ambiguous | 0.61 | Ambiguous | 0.724 | Likely Pathogenic | -4.31 | Deleterious | 0.276 | Benign | 0.103 | Benign | 5.82 | Benign | 0.12 | Tolerated | 0.3194 | 0.3426 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||
| c.1970G>T | W657L 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant W657L is listed in ClinVar with an uncertain significance (ClinVar ID 2767440.0) and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, FoldX, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, ESM1b, and AlphaMissense‑Default. Uncertain predictions come from Foldetta, premPS, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic, and Foldetta predicts a benign folding‑stability change. Overall, the majority of evidence points toward a pathogenic impact, which is consistent with the ClinVar uncertain status but leans toward pathogenic rather than benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.030611 | Structured | 0.208729 | Uncertain | 0.941 | 0.245 | 0.000 | Uncertain | 1 | -14.411 | Likely Pathogenic | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.14 | Likely Benign | 0.1 | 0.73 | Ambiguous | 0.44 | Likely Benign | 0.87 | Ambiguous | 0.213 | Likely Benign | -10.86 | Deleterious | 0.277 | Benign | 0.078 | Benign | 3.52 | Benign | 0.14 | Tolerated | 3.39 | 24 | 0.2302 | 0.2049 | -2 | -2 | 4.7 | -73.05 | |||||||||||||||||||||||||
| c.2711T>A | M904K 2D AIThe SynGAP1 missense variant M904K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.589073 | Binding | 0.350 | 0.920 | 0.250 | -2.333 | Likely Benign | 0.744 | Likely Pathogenic | Likely Benign | 0.033 | Likely Benign | -1.08 | Neutral | 0.277 | Benign | 0.137 | Benign | 2.80 | Benign | 1.00 | Tolerated | 0.1696 | 0.0871 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||||||||||||
| c.2711T>C | M904T 2D AIThe SynGAP1 missense variant M904T is listed in ClinVar (ID 1311496.0) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; a Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.589073 | Binding | 0.350 | 0.920 | 0.250 | Uncertain | 1 | -2.721 | Likely Benign | 0.668 | Likely Pathogenic | Likely Benign | 0.042 | Likely Benign | -1.15 | Neutral | 0.277 | Benign | 0.103 | Benign | 2.78 | Benign | 0.18 | Tolerated | 3.77 | 5 | 0.2394 | 0.2568 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||
| c.1480A>G | I494V 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant I494V is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33438512‑A‑G). Functional prediction tools that agree on benign impact include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Pathogenic predictions come from premPS and FATHMM. Predictions that are inconclusive are FoldX, Rosetta, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign; Foldetta remains uncertain. Overall, the majority of evidence supports a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | Conflicting | 2 | 6-33438512-A-G | 36 | 2.23e-5 | -7.102 | In-Between | 0.112 | Likely Benign | Likely Benign | 1.16 | Ambiguous | 0.0 | 0.71 | Ambiguous | 0.94 | Ambiguous | 1.02 | Destabilizing | 0.439 | Likely Benign | -0.83 | Neutral | 0.278 | Benign | 0.179 | Benign | -1.30 | Pathogenic | 0.07 | Tolerated | 3.37 | 35 | 0.0965 | 0.2491 | 4 | 3 | -0.3 | -14.03 | 248.6 | 29.3 | 0.0 | 0.0 | -1.1 | 0.5 | X | Potentially Benign | The sec-butyl side chain of Ile494, located in an α-helix (res. Leu489-Glu519), packs against hydrophobic residues (e.g., Phe484, Leu465, Trp572, Ala493, Met468) in an inter-helix space (res. Leu489-Glu519 and res. Ala461-Phe476). In the variant simulations, the hydrophobic iso-propyl side chain of Val494, which is of a similar size and has similar physicochemical properties to Ile494 in the WT, resides similarly in the inter-helix hydrophobic space. Thus, no negative effects on the protein structure are observed. | ||||||||||||||
| c.1826G>C | G609A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G609A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX, PROVEAN, and FATHMM. One tool, Foldetta, yields an uncertain result. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.203786 | Uncertain | 0.851 | 0.252 | 0.000 | -6.790 | Likely Benign | 0.153 | Likely Benign | Likely Benign | 2.38 | Destabilizing | 0.3 | 0.01 | Likely Benign | 1.20 | Ambiguous | 0.43 | Likely Benign | 0.494 | Likely Benign | -2.65 | Deleterious | 0.282 | Benign | 0.164 | Benign | -1.43 | Pathogenic | 0.10 | Tolerated | 0.3812 | 0.3896 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||
| c.1940G>A | G647D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G647D is reported in ClinVar as having no entry and is present in gnomAD (6‑33441199‑G‑A). Functional prediction tools largely agree on a benign effect: REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized all predict benign. Only AlphaMissense‑Default predicts pathogenic, while FoldX, Foldetta, premPS, and ESM1b are uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields benign; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is uncertain. Overall, the consensus of most tools and the high‑accuracy predictions indicate that G647D is most likely benign, and this is consistent with the absence of a pathogenic ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.048328 | Structured | 0.325524 | Uncertain | 0.936 | 0.356 | 0.000 | 6-33441199-G-A | 1 | 6.20e-7 | -7.643 | In-Between | 0.582 | Likely Pathogenic | Likely Benign | 0.68 | Ambiguous | 0.1 | 0.33 | Likely Benign | 0.51 | Ambiguous | 0.56 | Ambiguous | 0.098 | Likely Benign | -1.63 | Neutral | 0.282 | Benign | 0.128 | Benign | 3.45 | Benign | 0.24 | Tolerated | 3.37 | 30 | 0.1630 | 0.1372 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||
| c.1321G>A | V441I 2D AIThe SynGAP1 missense variant V441I is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign, while only ESM1b predicts it as pathogenic. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) also indicates benign stability. No contradictory evidence is present. Based on the collective predictions, the variant is most likely benign, and this conclusion does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | -8.773 | Likely Pathogenic | 0.122 | Likely Benign | Likely Benign | -0.24 | Likely Benign | 0.3 | 0.11 | Likely Benign | -0.07 | Likely Benign | 0.25 | Likely Benign | 0.135 | Likely Benign | -0.82 | Neutral | 0.287 | Benign | 0.038 | Benign | 3.41 | Benign | 0.16 | Tolerated | 0.0694 | 0.3412 | 4 | 3 | 0.3 | 14.03 | |||||||||||||||||||||||||||||
| c.1493T>A | M498K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M498K is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: benign calls come only from polyPhen‑2 HumDiv and HumVar, whereas the remaining 15 tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) all predict pathogenicity. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized scores the variant as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) labels it likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts a destabilizing, pathogenic effect. Taken together, the overwhelming majority of evidence indicates that M498K is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.399612 | Uncertain | 0.932 | 0.158 | 0.000 | -11.622 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 2.82 | Destabilizing | 0.1 | 2.86 | Destabilizing | 2.84 | Destabilizing | 1.79 | Destabilizing | 0.867 | Likely Pathogenic | -4.53 | Deleterious | 0.287 | Benign | 0.120 | Benign | -1.37 | Pathogenic | 0.00 | Affected | 0.1281 | 0.0656 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.3682G>C | E1228Q 2D AIThe SynGAP1 missense variant E1228Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized indicates a benign effect, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a likely benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the computational evidence overwhelmingly supports a benign interpretation, and this is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.517562 | Disordered | 0.447051 | Uncertain | 0.892 | 0.546 | 0.500 | -2.675 | Likely Benign | 0.161 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.32 | Neutral | 0.287 | Benign | 0.112 | Benign | 2.69 | Benign | 0.25 | Tolerated | 0.0830 | 0.3797 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||
| c.379C>G | R127G 2D AIThe SynGAP1 R127G missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The high‑accuracy consensus, SGM‑Consensus, is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN; it yields a “Likely Benign” classification (3 benign vs. 1 pathogenic). AlphaMissense‑Optimized also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.711716 | Binding | 0.333 | 0.870 | 0.625 | -1.062 | Likely Benign | 0.700 | Likely Pathogenic | Likely Benign | 0.107 | Likely Benign | -2.17 | Neutral | 0.287 | Benign | 0.038 | Benign | 3.91 | Benign | 0.01 | Affected | 0.3561 | 0.3037 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.2689T>G | S897A 2D AIThe SynGAP1 missense variant S897A is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors—including REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods likewise support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.699094 | Disordered | 0.418474 | Uncertain | 0.292 | 0.928 | 0.500 | -3.959 | Likely Benign | 0.122 | Likely Benign | Likely Benign | 0.060 | Likely Benign | -0.48 | Neutral | 0.288 | Benign | 0.208 | Benign | 2.71 | Benign | 0.81 | Tolerated | 0.4539 | 0.5684 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2725A>G | M909V 2D AIThe SynGAP1 missense variant M909V is not reported in ClinVar and is absent from gnomAD, indicating no documented clinical or population evidence. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.696196 | Binding | 0.314 | 0.914 | 0.250 | -2.906 | Likely Benign | 0.230 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -0.88 | Neutral | 0.288 | Benign | 0.147 | Benign | 2.97 | Benign | 0.45 | Tolerated | 0.3295 | 0.3600 | 2 | 1 | 2.3 | -32.06 | |||||||||||||||||||||||||||||||||||||||
| c.2843G>C | G948A 2D AIThe SynGAP1 missense variant G948A is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988505 | Disordered | 0.862121 | Binding | 0.365 | 0.919 | 0.750 | -5.627 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.189 | Likely Benign | 0.04 | Neutral | 0.288 | Benign | 0.103 | Benign | 4.64 | Benign | 0.39 | Tolerated | 0.3350 | 0.4957 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3161G>C | G1054A 2D AIThe SynGAP1 missense variant G1054A is catalogued in gnomAD (ID 6‑33443713‑G‑C) but has no ClinVar entry. Across a broad panel of in‑silico predictors, every tool reports a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity, so the pathogenic‑prediction group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Based on the unanimous benign predictions and the lack of any ClinVar pathogenic classification, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.983019 | Disordered | 0.878015 | Binding | 0.389 | 0.936 | 0.875 | 6-33443713-G-C | -6.786 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.243 | Likely Benign | -0.05 | Neutral | 0.288 | Benign | 0.071 | Benign | 4.03 | Benign | 1.00 | Tolerated | 3.77 | 5 | 0.3305 | 0.5144 | 0 | 1 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||||||||
| c.3229A>G | T1077A 2D AIThe SynGAP1 missense variant T1077A is catalogued in gnomAD (ID 6‑33443781‑A‑G) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.988141 | Binding | 0.329 | 0.892 | 0.750 | 6-33443781-A-G | -3.303 | Likely Benign | 0.280 | Likely Benign | Likely Benign | 0.153 | Likely Benign | -0.60 | Neutral | 0.288 | Benign | 0.194 | Benign | 4.25 | Benign | 0.10 | Tolerated | 3.77 | 5 | 0.3373 | 0.4393 | 0 | 1 | 2.5 | -30.03 | ||||||||||||||||||||||||||||||||||||
| c.2384C>G | P795R 2D AIThe SynGAP1 missense variant P795R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only SIFT classifies the change as pathogenic, but this is the sole discordant call. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that P795R is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.972450 | Disordered | 0.410339 | Uncertain | 0.457 | 0.903 | 0.875 | -6.536 | Likely Benign | 0.221 | Likely Benign | Likely Benign | 0.059 | Likely Benign | -1.02 | Neutral | 0.290 | Benign | 0.114 | Benign | 4.25 | Benign | 0.05 | Affected | 0.1430 | 0.4065 | 0 | -2 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||||||||||
| c.2915C>G | P972R 2D AIThe SynGAP1 missense variant P972R is reported in gnomAD (ID 6‑33443467‑C‑G) but has no ClinVar entry. All evaluated in silico predictors classify it as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Based on the unanimous benign predictions and lack of ClinVar pathogenic annotation, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.954150 | Binding | 0.472 | 0.904 | 0.625 | 6-33443467-C-G | -4.483 | Likely Benign | 0.139 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -1.44 | Neutral | 0.290 | Benign | 0.114 | Benign | 4.23 | Benign | 0.12 | Tolerated | 4.32 | 2 | 0.1379 | 0.3931 | -2 | 0 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||||||||
| c.3038C>A | S1013Y 2D  AIThe SynGAP1 missense variant S1013Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.899570 | Binding | 0.308 | 0.846 | 0.625 | -4.882 | Likely Benign | 0.338 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.78 | Neutral | 0.290 | Benign | 0.124 | Benign | 2.66 | Benign | 0.03 | Affected | 0.0954 | 0.5629 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.3397A>T | I1133F 2D AIThe SynGAP1 missense variant I1133F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect. The variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.832785 | Binding | 0.316 | 0.892 | 0.750 | -2.941 | Likely Benign | 0.164 | Likely Benign | Likely Benign | 0.272 | Likely Benign | -1.21 | Neutral | 0.290 | Benign | 0.124 | Benign | 5.45 | Benign | 0.05 | Affected | 0.0598 | 0.3522 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2398G>A | G800S 2D AIThe SynGAP1 missense variant G800S is reported as “Likely Benign” by the SGM‑Consensus method and is absent from ClinVar and gnomAD. All evaluated in‑silico predictors agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.852992 | Disordered | 0.588350 | Binding | 0.303 | 0.884 | 0.625 | -3.572 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -0.69 | Neutral | 0.292 | Benign | 0.157 | Benign | 2.78 | Benign | 0.60 | Tolerated | 0.2375 | 0.4923 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.2659C>A | P887T 2D AIThe SynGAP1 missense variant P887T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign,” while Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.716283 | Disordered | 0.602269 | Binding | 0.348 | 0.925 | 0.500 | -4.780 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -1.47 | Neutral | 0.292 | Benign | 0.110 | Benign | 2.79 | Benign | 0.21 | Tolerated | 0.1228 | 0.3822 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2927T>C | F976S 2D  AIThe SynGAP1 missense variant F976S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.975061 | Binding | 0.311 | 0.894 | 0.625 | -2.393 | Likely Benign | 0.432 | Ambiguous | Likely Benign | 0.217 | Likely Benign | -0.73 | Neutral | 0.292 | Benign | 0.102 | Benign | 4.16 | Benign | 0.49 | Tolerated | 0.4432 | 0.1134 | -3 | -2 | -3.6 | -60.10 | |||||||||||||||||||||||||||||||||||||||
| c.3107A>G | Q1036R 2D AIThe SynGAP1 missense variant Q1036R is listed in ClinVar (ID 4746139) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely benign classification (3 benign vs. 1 pathogenic). High‑accuracy assessments further support benignity: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.987955 | Binding | 0.275 | 0.765 | 0.625 | Uncertain | 1 | -3.816 | Likely Benign | 0.596 | Likely Pathogenic | Likely Benign | 0.083 | Likely Benign | -1.32 | Neutral | 0.292 | Benign | 0.071 | Benign | 2.54 | Benign | 0.04 | Affected | 0.1710 | 0.3071 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||
| c.3110T>C | I1037T 2D AIThe SynGAP1 missense variant I1037T is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33443662‑T‑C). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are AlphaMissense‑Default and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains Likely Benign; Foldetta results are unavailable. Overall, the majority of predictions (seven benign vs. two pathogenic) indicate that the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.939629 | Disordered | 0.986140 | Binding | 0.309 | 0.774 | 0.625 | 6-33443662-T-C | 1 | 6.21e-7 | -2.565 | Likely Benign | 0.973 | Likely Pathogenic | Likely Pathogenic | 0.066 | Likely Benign | 0.40 | Neutral | 0.292 | Benign | 0.110 | Benign | 2.79 | Benign | 0.34 | Tolerated | 3.77 | 5 | 0.1071 | 0.2175 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||
| c.3167G>T | G1056V 2D AIThe SynGAP1 missense variant G1056V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence (7 benign vs 2 pathogenic) supports a benign classification. This consensus does not contradict ClinVar status, which has no entry for this variant. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988291 | Disordered | 0.868632 | Binding | 0.402 | 0.935 | 0.875 | -8.130 | Likely Pathogenic | 0.097 | Likely Benign | Likely Benign | 0.448 | Likely Benign | -0.24 | Neutral | 0.292 | Benign | 0.110 | Benign | 1.83 | Pathogenic | 0.08 | Tolerated | 0.1488 | 0.3507 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3221A>G | Q1074R 2D AIThe SynGAP1 missense variant Q1074R is listed in gnomAD (ID 6‑33443773‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none reported). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.987006 | Binding | 0.339 | 0.897 | 0.750 | 6-33443773-A-G | -5.710 | Likely Benign | 0.594 | Likely Pathogenic | Likely Benign | 0.124 | Likely Benign | -0.54 | Neutral | 0.292 | Benign | 0.157 | Benign | 2.70 | Benign | 0.28 | Tolerated | 3.77 | 5 | 0.1523 | 0.2643 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||
| c.3332A>C | K1111T 2D AIThe SynGAP1 missense variant K1111T is listed in ClinVar (ID 4746140) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443884‑A‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. No tool in the dataset predicts pathogenicity. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Overall, the collective predictions point to a benign effect, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.921455 | Binding | 0.300 | 0.902 | 0.875 | Uncertain | 1 | 6-33443884-A-C | 2 | 1.32e-6 | -4.037 | Likely Benign | 0.519 | Ambiguous | Likely Benign | 0.080 | Likely Benign | -0.90 | Neutral | 0.292 | Benign | 0.110 | Benign | 2.64 | Benign | 0.28 | Tolerated | 4.32 | 2 | 0.2250 | 0.4425 | -1 | 0 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||
| c.3359G>T | G1120V 2D AIThe SynGAP1 missense variant G1120V is catalogued in gnomAD (6‑33443911‑G‑T) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the variant as benign, while ESM1b remains uncertain. No tool predicts pathogenicity. The high‑accuracy consensus, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), classifies the variant as “Likely Benign.” AlphaMissense‑Optimized also reports a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the evidence strongly supports a benign classification, and this assessment does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.871313 | Disordered | 0.804931 | Binding | 0.335 | 0.925 | 0.875 | 6-33443911-G-T | -7.659 | In-Between | 0.081 | Likely Benign | Likely Benign | 0.355 | Likely Benign | -1.20 | Neutral | 0.292 | Benign | 0.157 | Benign | 3.61 | Benign | 0.06 | Tolerated | 3.77 | 5 | 0.1360 | 0.3694 | -3 | -1 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||
| c.3367G>A | G1123S 2D AIThe SynGAP1 missense variant G1123S is reported in gnomAD (variant ID 6-33443919-G-A) but has no ClinVar entry. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.837511 | Disordered | 0.827246 | Binding | 0.346 | 0.934 | 0.875 | 6-33443919-G-A | -4.918 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.291 | Likely Benign | -0.32 | Neutral | 0.292 | Benign | 0.157 | Benign | 4.66 | Benign | 0.57 | Tolerated | 3.77 | 5 | 0.2554 | 0.5111 | 0 | 1 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||||||
| c.3368G>T | G1123V 2D AIThe SynGAP1 missense variant G1123V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; ESM1b is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective predictions strongly suggest that G1123V is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.837511 | Disordered | 0.827246 | Binding | 0.346 | 0.934 | 0.875 | -7.129 | In-Between | 0.091 | Likely Benign | Likely Benign | 0.333 | Likely Benign | -1.03 | Neutral | 0.292 | Benign | 0.157 | Benign | 4.35 | Benign | 0.29 | Tolerated | 0.1362 | 0.3694 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3847C>T | P1283S 2D AIThe SynGAP1 missense variant P1283S is catalogued in gnomAD (ID 6‑33447895‑C‑T) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Therefore, the variant is most likely benign, and this assessment does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.819686 | Binding | 0.484 | 0.732 | 0.875 | 6-33447895-C-T | -4.289 | Likely Benign | 0.072 | Likely Benign | Likely Benign | 0.040 | Likely Benign | 0.06 | Neutral | 0.292 | Benign | 0.110 | Benign | 2.86 | Benign | 0.30 | Tolerated | 3.77 | 5 | 0.2837 | 0.3719 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||
| c.2033G>T | S678I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split. Overall, the majority of evidence (10 benign vs. 3 pathogenic) supports a benign classification. This consensus does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.735 | Likely Pathogenic | 0.253 | Likely Benign | Likely Benign | 0.36 | Likely Benign | 0.3 | -0.01 | Likely Benign | 0.18 | Likely Benign | -0.11 | Likely Benign | 0.119 | Likely Benign | -3.99 | Deleterious | 0.294 | Benign | 0.057 | Benign | 3.45 | Benign | 0.01 | Affected | 0.0941 | 0.5954 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||
| c.20C>T | S7F 2D  AIThe SynGAP1 missense variant S7F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. AlphaMissense‑Optimized independently predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.590140 | Disordered | 0.548467 | Binding | 0.386 | 0.922 | 0.750 | -4.346 | Likely Benign | 0.250 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -0.24 | Neutral | 0.296 | Benign | 0.032 | Benign | 4.06 | Benign | 0.00 | Affected | 0.0622 | 0.5177 | -3 | -2 | 3.6 | 60.10 | |||||||||||||||||||||||||||||||||||||||
| c.24C>G | I8M 2D  AIThe SynGAP1 missense variant I8M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign status. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points to a benign effect, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.490133 | Structured | 0.543080 | Binding | 0.341 | 0.916 | 0.625 | -3.957 | Likely Benign | 0.140 | Likely Benign | Likely Benign | 0.085 | Likely Benign | -0.07 | Neutral | 0.296 | Benign | 0.022 | Benign | 4.02 | Benign | 0.00 | Affected | 0.0626 | 0.3034 | 2 | 1 | -2.6 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.39C>G | I13M 2D AIThe SynGAP1 missense variant I13M is reported in gnomAD (ID 6‑33420303‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus itself is likely benign. Foldetta results are not available, so they do not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | 6-33420303-C-G | 1 | 6.49e-7 | -4.097 | Likely Benign | 0.170 | Likely Benign | Likely Benign | 0.093 | Likely Benign | 0.16 | Neutral | 0.296 | Benign | 0.022 | Benign | 4.04 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0883 | 0.3806 | 1 | 2 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||||||
| c.1280A>T | H427L 2D 3DClick to see structure in 3D Viewer AISynGAP1 H427L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: nine tools (REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized) predict a benign effect, while five tools (SGM‑Consensus, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default) predict pathogenicity. High‑accuracy methods provide a more focused view: AlphaMissense‑Optimized indicates a benign outcome; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, remains pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. Taken together, the majority of evidence supports a benign impact for H427L, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.081712 | Structured | 0.394261 | Uncertain | 0.962 | 0.287 | 0.000 | -9.691 | Likely Pathogenic | 0.755 | Likely Pathogenic | Likely Benign | -0.17 | Likely Benign | 0.0 | 0.05 | Likely Benign | -0.06 | Likely Benign | 0.31 | Likely Benign | 0.272 | Likely Benign | -5.38 | Deleterious | 0.299 | Benign | 0.033 | Benign | 3.42 | Benign | 0.01 | Affected | 0.0942 | 0.4953 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||
| c.3835G>C | A1279P 2D  AIThe SynGAP1 missense variant A1279P is not reported in ClinVar and is absent from gnomAD. In silico prediction tools that assess sequence conservation and structural impact (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) all classify the change as benign. No tool in the dataset predicts pathogenicity. High‑accuracy consensus methods reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign effect. Foldetta, a protein‑folding stability predictor, was not available for this variant. Overall, the collective evidence strongly suggests the variant is most likely benign, and this conclusion is consistent with the lack of a ClinVar classification—there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.842060 | Disordered | 0.814139 | Binding | 0.485 | 0.724 | 0.750 | -3.999 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.167 | Likely Benign | -0.68 | Neutral | 0.299 | Benign | 0.087 | Benign | 2.67 | Benign | 0.15 | Tolerated | 0.1589 | 0.3938 | 1 | -1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||||||
| c.1312G>A | A438T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A438T is listed in ClinVar with an “Uncertain” status and is present in the gnomAD database (gnomAD ID 6‑33438217‑G‑A). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool in the dataset predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict the current ClinVar status of “Uncertain.” Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.147574 | Structured | 0.290154 | Uncertain | 0.929 | 0.293 | 0.000 | Conflicting | 3 | 6-33438217-G-A | 16 | 9.91e-6 | -5.339 | Likely Benign | 0.085 | Likely Benign | Likely Benign | 0.21 | Likely Benign | 0.0 | -0.07 | Likely Benign | 0.07 | Likely Benign | 0.36 | Likely Benign | 0.021 | Likely Benign | -0.81 | Neutral | 0.300 | Benign | 0.011 | Benign | 4.18 | Benign | 0.14 | Tolerated | 3.38 | 26 | 0.0999 | 0.5574 | 1 | 0 | -2.5 | 30.03 | 214.2 | -42.7 | -0.3 | 0.1 | -0.4 | 0.1 | X | Potentially Benign | The methyl group of Ala438, located in a four-residue loop connecting two α helices (res. Asn440-Thr458 and Pro413-Glu436), packs against hydrophobic residues from a nearby α helix or loop residues (e.g., Leu703, Val699). In the variant simulations, the methyl group of Thr438 is able to establish similar hydrophobic packing. Moreover, the hydroxyl group also H-bonds with nearby residues, such as the carbonyl group of the neighboring loop residue Pro437. Accordingly, the residue swap does not generate an apparent negative effect on the protein structure based on the simulations. | |||||||||||||
| c.2651G>T | R884L 2D AIThe SynGAP1 missense variant R884L is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification, so there is no contradiction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.720929 | Disordered | 0.641526 | Binding | 0.305 | 0.898 | 0.250 | -3.482 | Likely Benign | 0.280 | Likely Benign | Likely Benign | 0.095 | Likely Benign | -1.08 | Neutral | 0.300 | Benign | 0.191 | Benign | 2.63 | Benign | 0.24 | Tolerated | 0.1809 | 0.4177 | -3 | -2 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||||
| c.652T>G | F218V 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F218V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; pathogenic predictions arise from REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is labeled “Likely Pathogenic.” High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus agrees on a likely pathogenic outcome, and Foldetta (combining FoldX‑MD and Rosetta outputs) also indicates pathogenicity. No predictions or stability results are missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion aligns with the absence of a ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.281712 | Structured | 0.408725 | Uncertain | 0.848 | 0.272 | 0.000 | -10.081 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 4.18 | Destabilizing | 0.2 | 6.35 | Destabilizing | 5.27 | Destabilizing | 1.15 | Destabilizing | 0.691 | Likely Pathogenic | -4.14 | Deleterious | 0.300 | Benign | 0.066 | Benign | 5.81 | Benign | 0.08 | Tolerated | 0.2173 | 0.2258 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.658T>A | F220I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F220I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. All other evaluated algorithms—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic effect. The SGM‑Consensus result is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yielding 3 pathogenic versus 1 benign calls, thus supporting a pathogenic classification. High‑accuracy tools specifically report pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM‑Consensus (majority vote) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is pathogenic. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.219301 | Structured | 0.429422 | Uncertain | 0.898 | 0.295 | 0.000 | -12.041 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 3.11 | Destabilizing | 0.1 | 2.35 | Destabilizing | 2.73 | Destabilizing | 1.03 | Destabilizing | 0.921 | Likely Pathogenic | -4.98 | Deleterious | 0.300 | Benign | 0.098 | Benign | 4.06 | Benign | 0.01 | Affected | 0.2292 | 0.3127 | 1 | 0 | 1.7 | -34.02 | |||||||||||||||||||||||||||||
| c.3758C>A | A1253E 2D AIThe SynGAP1 missense variant A1253E is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset predicts pathogenicity. High‑accuracy assessments corroborate this benign classification: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are not available, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.661982 | Disordered | 0.391377 | Uncertain | 0.881 | 0.550 | 0.750 | -1.744 | Likely Benign | 0.166 | Likely Benign | Likely Benign | 0.059 | Likely Benign | 2.03 | Neutral | 0.301 | Benign | 0.209 | Benign | 3.03 | Benign | 0.98 | Tolerated | 0.0948 | 0.1704 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||||
| c.2974G>A | V992I 2D AIThe SynGAP1 missense variant V992I is catalogued in gnomAD (ID 6‑33443526‑G‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as tolerated or benign. No tool in the set indicates pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a Likely Benign verdict. Foldetta results are not available for this variant. Consequently, the variant is most likely benign, and this prediction does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.921728 | Binding | 0.331 | 0.917 | 0.750 | 6-33443526-G-A | 5 | 3.10e-6 | -3.851 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.052 | Likely Benign | -0.30 | Neutral | 0.302 | Benign | 0.089 | Benign | 4.20 | Benign | 0.29 | Tolerated | 4.32 | 2 | 0.0939 | 0.4541 | 3 | 4 | 0.3 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.2975T>A | V992D 2D  AIThe SynGAP1 missense variant V992D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while only SIFT predicts pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools, including the high‑accuracy methods, indicates that V992D is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.921728 | Binding | 0.331 | 0.917 | 0.750 | -3.976 | Likely Benign | 0.306 | Likely Benign | Likely Benign | 0.029 | Likely Benign | -1.24 | Neutral | 0.302 | Benign | 0.158 | Benign | 4.21 | Benign | 0.05 | Affected | 0.1645 | 0.1056 | -2 | -3 | -7.7 | 15.96 | |||||||||||||||||||||||||||||||||||||||
| c.1259T>A | F420Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F420Y is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show mixed results. Benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions are reported by premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. Predictions from FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized are inconclusive. High‑accuracy assessments indicate that the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Pathogenic, while AlphaMissense‑Optimized and Foldetta remain uncertain. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.104810 | Structured | 0.384475 | Uncertain | 0.974 | 0.255 | 0.000 | -12.138 | Likely Pathogenic | 0.941 | Likely Pathogenic | Ambiguous | 1.18 | Ambiguous | 0.1 | 0.96 | Ambiguous | 1.07 | Ambiguous | 1.31 | Destabilizing | 0.228 | Likely Benign | -2.80 | Deleterious | 0.306 | Benign | 0.100 | Benign | 3.09 | Benign | 0.03 | Affected | 0.1609 | 0.1498 | 7 | 3 | -4.1 | 16.00 | |||||||||||||||||||||||||||||
| c.257T>G | V86G 2D  AIThe SynGAP1 missense variant V86G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. Foldetta, which evaluates protein‑folding stability via FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of evidence (five pathogenic versus four benign predictions) indicates that V86G is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.685117 | Disordered | 0.552911 | Binding | 0.295 | 0.887 | 0.500 | -4.212 | Likely Benign | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.130 | Likely Benign | -2.50 | Deleterious | 0.307 | Benign | 0.824 | Possibly Damaging | 3.74 | Benign | 0.00 | Affected | 0.2321 | 0.2547 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.404G>T | R135L 2D AIThe SynGAP1 missense variant R135L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. The high‑accuracy assessment shows AlphaMissense‑Optimized predicts pathogenicity. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (benign), and PROVEAN (pathogenic), also indicates pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions, including the high‑accuracy tools, support a pathogenic classification. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.465241 | Structured | 0.676514 | Binding | 0.380 | 0.898 | 0.250 | -7.372 | In-Between | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.164 | Likely Benign | -3.61 | Deleterious | 0.308 | Benign | 0.122 | Benign | 3.70 | Benign | 0.01 | Affected | 0.1962 | 0.4727 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.1931A>C | D644A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D644A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, Foldetta, and premPS. Only PROVEAN predicts it as pathogenic, while Rosetta and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign; and Foldetta also predicts benign stability. Based on the aggregate evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.248888 | Uncertain | 0.883 | 0.320 | 0.000 | -3.358 | Likely Benign | 0.502 | Ambiguous | Likely Benign | -0.14 | Likely Benign | 0.1 | -0.83 | Ambiguous | -0.49 | Likely Benign | -0.18 | Likely Benign | 0.274 | Likely Benign | -3.90 | Deleterious | 0.311 | Benign | 0.032 | Benign | 3.51 | Benign | 0.39 | Tolerated | 0.3883 | 0.5481 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||
| c.2252C>T | P751L 2D AIThe SynGAP1 missense variant P751L is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. No pathogenic predictions are present among the evaluated tools. High‑accuracy assessments confirm this trend: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are unavailable. Consequently, the variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.667683 | Binding | 0.386 | 0.866 | 0.625 | -3.558 | Likely Benign | 0.143 | Likely Benign | Likely Benign | 0.207 | Likely Benign | -1.62 | Neutral | 0.316 | Benign | 0.062 | Benign | 2.94 | Benign | 0.24 | Tolerated | 0.2318 | 0.6451 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.2515A>G | K839E 2D AIThe SynGAP1 missense variant K839E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Tools that predict a pathogenic effect are SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, while Foldetta results are unavailable. Overall, the balance of evidence—five pathogenic predictions versus four benign, plus a Likely Pathogenic consensus—suggests the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.642678 | Disordered | 0.611185 | Binding | 0.282 | 0.865 | 0.375 | -12.616 | Likely Pathogenic | 0.931 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -1.90 | Neutral | 0.316 | Benign | 0.139 | Benign | 2.47 | Pathogenic | 0.01 | Affected | 0.3822 | 0.1198 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.2802G>A | M934I 2D AIThe SynGAP1 missense variant M934I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are unavailable. Overall, the majority of evidence (seven benign versus two pathogenic predictions) indicates that M934I is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.984677 | Binding | 0.290 | 0.867 | 0.625 | -4.582 | Likely Benign | 0.757 | Likely Pathogenic | Likely Benign | 0.086 | Likely Benign | -1.78 | Neutral | 0.316 | Benign | 0.101 | Benign | 2.49 | Pathogenic | 0.27 | Tolerated | 0.1686 | 0.3100 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2802G>C | M934I 2D AIThe SynGAP1 missense variant M934I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are unavailable. Overall, the majority of evidence (seven benign versus two pathogenic predictions) indicates that M934I is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.984677 | Binding | 0.290 | 0.867 | 0.625 | -4.582 | Likely Benign | 0.757 | Likely Pathogenic | Likely Benign | 0.085 | Likely Benign | -1.78 | Neutral | 0.316 | Benign | 0.101 | Benign | 2.49 | Pathogenic | 0.27 | Tolerated | 0.1686 | 0.3100 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.2802G>T | M934I 2D AIThe SynGAP1 missense variant M934I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are unavailable. Overall, the majority of evidence (seven benign versus two pathogenic predictions) indicates that M934I is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.762850 | Disordered | 0.984677 | Binding | 0.290 | 0.867 | 0.625 | -4.582 | Likely Benign | 0.757 | Likely Pathogenic | Likely Benign | 0.085 | Likely Benign | -1.78 | Neutral | 0.316 | Benign | 0.101 | Benign | 2.49 | Pathogenic | 0.27 | Tolerated | 0.1686 | 0.3100 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.596A>T | N199I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N199I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields an equal split of benign and pathogenic calls. Overall, the majority of evidence points to a benign impact. The variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.390993 | Structured | 0.431347 | Uncertain | 0.571 | 0.473 | 0.125 | -8.299 | Likely Pathogenic | 0.328 | Likely Benign | Likely Benign | 0.27 | Likely Benign | 0.1 | -0.11 | Likely Benign | 0.08 | Likely Benign | 0.20 | Likely Benign | 0.066 | Likely Benign | -3.27 | Deleterious | 0.316 | Benign | 0.045 | Benign | 4.16 | Benign | 0.01 | Affected | 0.0450 | 0.6009 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||
| c.3917A>G | N1306S 2D AIThe SynGAP1 missense variant N1306S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, while Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -1.344 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.233 | Likely Benign | -3.49 | Deleterious | 0.319 | Benign | 0.242 | Benign | 2.69 | Benign | 0.00 | Affected | 0.4120 | 0.7826 | 1 | 1 | 2.7 | -27.03 | |||||||||||||||||||||||||||||||||||||||
| c.427C>G | R143G 2D AIThe SynGAP1 missense variant R143G is not reported in ClinVar and is absent from gnomAD. In silico predictors that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Predictors that agree on a pathogenic effect include PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the preponderance of evidence from multiple independent tools points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, as no ClinVar annotation exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.575842 | Disordered | 0.538584 | Binding | 0.338 | 0.838 | 0.625 | -12.686 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.151 | Likely Benign | -3.86 | Deleterious | 0.319 | Benign | 0.124 | Benign | 3.51 | Benign | 0.00 | Affected | 0.3502 | 0.3547 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.428G>T | R143L 2D AIThe SynGAP1 missense variant R143L is listed in ClinVar with no submitted interpretation and is present in gnomAD (ID 6‑33432725‑G‑T). Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic; the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.575842 | Disordered | 0.538584 | Binding | 0.338 | 0.838 | 0.625 | 6-33432725-G-T | 1 | 6.77e-7 | -14.250 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.316 | Likely Benign | -4.37 | Deleterious | 0.319 | Benign | 0.124 | Benign | 3.52 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1758 | 0.4886 | -2 | -3 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||
| c.128G>T | G43V 2D AIThe SynGAP1 missense variant G43V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.308712 | Structured | 0.431462 | Uncertain | 0.396 | 0.762 | 0.375 | -3.745 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.115 | Likely Benign | -0.79 | Neutral | 0.320 | Benign | 0.140 | Benign | 4.28 | Benign | 0.00 | Affected | 0.1156 | 0.3120 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.1432G>A | E478K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E478K is not reported in ClinVar (ClinVar ID: None) and has no entries in gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SGM‑Consensus (Likely Pathogenic), PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta as benign. Overall, the majority of evidence (eight benign versus five pathogenic predictions) points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | -12.654 | Likely Pathogenic | 0.899 | Likely Pathogenic | Ambiguous | 0.23 | Likely Benign | 0.0 | 0.12 | Likely Benign | 0.18 | Likely Benign | -0.02 | Likely Benign | 0.309 | Likely Benign | -3.45 | Deleterious | 0.320 | Benign | 0.117 | Benign | 3.49 | Benign | 0.05 | Affected | 0.2066 | 0.6192 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1984C>A | Q662K 2D AIThe SynGAP1 missense variant Q662K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign, while only ESM1b predicts pathogenicity. When high‑accuracy methods are considered separately, AlphaMissense‑Optimized remains benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a Likely Benign verdict, and Foldetta (combining FoldX‑MD and Rosetta outputs) also reports a benign effect. No conflicting evidence is present. Therefore, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.046336 | Structured | 0.103446 | Uncertain | 0.932 | 0.323 | 0.000 | -8.892 | Likely Pathogenic | 0.309 | Likely Benign | Likely Benign | -0.02 | Likely Benign | 0.2 | 0.03 | Likely Benign | 0.01 | Likely Benign | -0.04 | Likely Benign | 0.108 | Likely Benign | -0.80 | Neutral | 0.321 | Benign | 0.030 | Benign | 3.49 | Benign | 0.37 | Tolerated | 0.2391 | 0.3198 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||
| c.1319A>T | N440I 2D AISynGAP1 missense variant N440I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized, while pathogenic calls come from PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. Uncertain results are reported by FoldX, Rosetta, and Foldetta. High‑accuracy assessments indicate that AlphaMissense‑Optimized predicts a benign effect, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as likely pathogenic; Foldetta remains inconclusive. Overall, the balance of evidence favors a pathogenic interpretation, and this conclusion does not contradict the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -10.365 | Likely Pathogenic | 0.778 | Likely Pathogenic | Likely Benign | 0.97 | Ambiguous | 0.9 | 1.10 | Ambiguous | 1.04 | Ambiguous | 0.10 | Likely Benign | 0.100 | Likely Benign | -4.07 | Deleterious | 0.322 | Benign | 0.109 | Benign | 3.47 | Benign | 0.03 | Affected | 0.0554 | 0.3772 | -2 | -3 | 8.0 | -0.94 | |||||||||||||||||||||||||||||
| c.1919C>T | T640I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T640I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only PROVEAN predicts a pathogenic outcome. The remaining tools (FoldX, Rosetta, ESM1b, AlphaMissense‑Default, and Foldetta) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is unavailable due to a tie, and Foldetta also yields an uncertain stability change. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.137043 | Uncertain | 0.893 | 0.284 | 0.000 | -7.256 | In-Between | 0.358 | Ambiguous | Likely Benign | 0.70 | Ambiguous | 0.1 | 0.66 | Ambiguous | 0.68 | Ambiguous | 0.28 | Likely Benign | 0.193 | Likely Benign | -2.75 | Deleterious | 0.322 | Benign | 0.022 | Benign | 3.46 | Benign | 0.12 | Tolerated | 0.1030 | 0.4793 | 0 | -1 | 5.2 | 12.05 | ||||||||||||||||||||||||||||||
| c.3938C>A | P1313Q 2D  AIThe SynGAP1 missense variant P1313Q is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the consensus of all available predictions strongly supports a benign classification, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.970301 | Binding | 0.452 | 0.902 | 0.750 | -4.533 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.063 | Likely Benign | 0.99 | Neutral | 0.324 | Benign | 0.287 | Benign | 4.31 | Benign | 0.15 | Tolerated | 0.1748 | 0.5651 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||||
| c.3938C>G | P1313R 2D AIThe SynGAP1 missense variant P1313R is not reported in ClinVar and has no entry in gnomAD. Prediction tools that assess pathogenicity are unanimous: all listed algorithms (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a benign effect, while none predict pathogenicity. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized scores the variant as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Benign,” and the protein‑folding stability tool Foldetta provides no result, so it does not influence the assessment. Based on the collective predictions, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.970301 | Binding | 0.452 | 0.902 | 0.750 | -4.634 | Likely Benign | 0.161 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.52 | Neutral | 0.324 | Benign | 0.172 | Benign | 4.25 | Benign | 0.22 | Tolerated | 0.1659 | 0.4742 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.2387C>A | P796Q 2D AIThe SynGAP1 missense variant P796Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign, while the only tool predicting a deleterious outcome is SIFT, which labels it pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) indicates likely benign; no Foldetta stability data are available. Overall, the consensus of the majority of predictors and the high‑accuracy tools points to a benign effect for P796Q, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar classification, as no pathogenic claim is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.936162 | Disordered | 0.426363 | Uncertain | 0.427 | 0.900 | 0.875 | -6.575 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.074 | Likely Benign | -0.15 | Neutral | 0.325 | Benign | 0.103 | Benign | 4.27 | Benign | 0.01 | Affected | 0.1562 | 0.4117 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||||
| c.2387C>T | P796L 2D AIThe SynGAP1 P796L missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is also benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.936162 | Disordered | 0.426363 | Uncertain | 0.427 | 0.900 | 0.875 | -5.442 | Likely Benign | 0.069 | Likely Benign | Likely Benign | 0.063 | Likely Benign | -1.06 | Neutral | 0.325 | Benign | 0.182 | Benign | 4.24 | Benign | 0.01 | Affected | 0.2328 | 0.5712 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||||
| c.2885A>G | H962R 2D AIThe SynGAP1 missense variant H962R is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” status. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of any ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.984483 | Binding | 0.369 | 0.886 | 0.750 | -9.166 | Likely Pathogenic | 0.212 | Likely Benign | Likely Benign | 0.117 | Likely Benign | -1.04 | Neutral | 0.325 | Benign | 0.129 | Benign | 4.18 | Benign | 0.07 | Tolerated | 0.2276 | 0.3337 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.2886C>A | H962Q 2D AIThe SynGAP1 missense variant H962Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.984483 | Binding | 0.369 | 0.886 | 0.750 | -8.161 | Likely Pathogenic | 0.130 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -1.04 | Neutral | 0.325 | Benign | 0.045 | Benign | 4.19 | Benign | 0.06 | Tolerated | 0.1943 | 0.3844 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2886C>G | H962Q 2D AIThe SynGAP1 missense variant H962Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. There is no ClinVar entry to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.984483 | Binding | 0.369 | 0.886 | 0.750 | -8.161 | Likely Pathogenic | 0.130 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -1.04 | Neutral | 0.325 | Benign | 0.045 | Benign | 4.19 | Benign | 0.06 | Tolerated | 0.1943 | 0.3844 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.3092T>A | M1031K 2D AIThe SynGAP1 missense variant M1031K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.865454 | Disordered | 0.995959 | Binding | 0.340 | 0.736 | 0.500 | -1.940 | Likely Benign | 0.709 | Likely Pathogenic | Likely Benign | 0.114 | Likely Benign | -0.80 | Neutral | 0.325 | Benign | 0.098 | Benign | 2.66 | Benign | 0.18 | Tolerated | 0.1224 | 0.1412 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||||||||||||
| c.3092T>G | M1031R 2D AIThe SynGAP1 missense variant M1031R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Taken together, the majority of evidence points to a benign impact for M1031R, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.865454 | Disordered | 0.995959 | Binding | 0.340 | 0.736 | 0.500 | -1.365 | Likely Benign | 0.673 | Likely Pathogenic | Likely Benign | 0.117 | Likely Benign | -0.85 | Neutral | 0.325 | Benign | 0.129 | Benign | 2.64 | Benign | 0.12 | Tolerated | 0.1368 | 0.1212 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.1270G>C | V424L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V424L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome, while SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports a likely benign classification. Stability‑based methods are inconclusive: FoldX is uncertain, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.050641 | Structured | 0.411431 | Uncertain | 0.973 | 0.248 | 0.000 | -4.941 | Likely Benign | 0.742 | Likely Pathogenic | Likely Benign | -0.90 | Ambiguous | 0.2 | -0.41 | Likely Benign | -0.66 | Ambiguous | 0.43 | Likely Benign | 0.159 | Likely Benign | -1.68 | Neutral | 0.327 | Benign | 0.026 | Benign | 4.50 | Benign | 0.59 | Tolerated | 0.1018 | 0.2680 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||
| c.2342T>G | M781R 2D AIThe SynGAP1 missense variant M781R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.580690 | Disordered | 0.792850 | Binding | 0.342 | 0.889 | 0.625 | -4.990 | Likely Benign | 0.697 | Likely Pathogenic | Likely Benign | 0.265 | Likely Benign | -1.72 | Neutral | 0.327 | Benign | 0.206 | Benign | 2.71 | Benign | 0.14 | Tolerated | 0.1685 | 0.0837 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.2893C>T | H965Y 2D AIThe SynGAP1 missense variant H965Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the substitution as tolerated. The consensus score from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” No tool predicts pathogenicity, and the single uncertain result from ESM1b does not alter the overall benign consensus. High‑accuracy methods corroborate this: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta data are unavailable. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988505 | Disordered | 0.978700 | Binding | 0.342 | 0.882 | 0.750 | -7.121 | In-Between | 0.133 | Likely Benign | Likely Benign | 0.092 | Likely Benign | -1.02 | Neutral | 0.327 | Benign | 0.147 | Benign | 4.03 | Benign | 0.25 | Tolerated | 0.1582 | 0.4906 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||||||
| c.404G>A | R135Q 2D  AIThe SynGAP1 missense variant R135Q is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33432701‑G‑A). Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; pathogenic predictions come from SIFT, ESM1b, and AlphaMissense‑Default. The remaining high‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑to‑2 tie, and Foldetta stability analysis is not available. Overall, the balance of evidence favors a benign effect, and this conclusion does not conflict with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.465241 | Structured | 0.676514 | Binding | 0.380 | 0.898 | 0.250 | Uncertain | 1 | 6-33432701-G-A | 5 | 3.84e-6 | -8.011 | Likely Pathogenic | 0.853 | Likely Pathogenic | Ambiguous | 0.087 | Likely Benign | -1.94 | Neutral | 0.327 | Benign | 0.100 | Benign | 3.76 | Benign | 0.02 | Affected | 3.61 | 5 | 0.3153 | 0.2741 | 1 | 1 | 1.0 | -28.06 | |||||||||||||||||||||||||||||||||
| c.143T>C | F48S 2D AIThe SynGAP1 missense variant F48S has no ClinVar assertion and is not reported in gnomAD. High‑accuracy predictors: AlphaMissense‑Optimized returned an uncertain classification; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect; Foldetta predictions are unavailable. Among the remaining tools, six predict benign (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM) while two predict pathogenic (SIFT, AlphaMissense‑Default). Based on the aggregate predictions, the variant is most likely benign; this is consistent with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.298791 | Structured | 0.440452 | Uncertain | 0.558 | 0.707 | 0.125 | -6.382 | Likely Benign | 0.916 | Likely Pathogenic | Ambiguous | 0.214 | Likely Benign | -2.25 | Neutral | 0.334 | Benign | 0.099 | Benign | 3.95 | Benign | 0.00 | Affected | 0.4577 | 0.0901 | -3 | -2 | -3.6 | -60.10 | |||||||||||||||||||||||||||||||||||||||
| c.148A>T | I50F 2D AIThe SynGAP1 missense variant I50F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for I50F, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.295083 | Structured | 0.449965 | Uncertain | 0.545 | 0.708 | 0.000 | -5.631 | Likely Benign | 0.681 | Likely Pathogenic | Likely Benign | 0.100 | Likely Benign | -1.27 | Neutral | 0.334 | Benign | 0.074 | Benign | 3.75 | Benign | 0.00 | Affected | 0.0483 | 0.2560 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.149T>G | I50S 2D  AIThe SynGAP1 I50S missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM all classify it as benign. In contrast, SIFT and AlphaMissense‑Default predict a pathogenic impact. AlphaMissense‑Optimized yields an uncertain result, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available prediction for this residue. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments therefore point to a benign outcome: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus is likely benign, and Foldetta data are missing. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.295083 | Structured | 0.449965 | Uncertain | 0.545 | 0.708 | 0.000 | -4.257 | Likely Benign | 0.946 | Likely Pathogenic | Ambiguous | 0.209 | Likely Benign | -2.03 | Neutral | 0.334 | Benign | 0.099 | Benign | 3.74 | Benign | 0.00 | Affected | 0.2626 | 0.0800 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.191T>A | I64K 2D  AIThe SynGAP1 missense variant I64K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains benign; Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, and this conclusion does not contradict the ClinVar status, which contains no report for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.422041 | Structured | 0.475481 | Uncertain | 0.478 | 0.747 | 0.125 | -3.206 | Likely Benign | 0.964 | Likely Pathogenic | Likely Pathogenic | 0.159 | Likely Benign | -0.47 | Neutral | 0.334 | Benign | 0.029 | Benign | 4.07 | Benign | 0.00 | Affected | 0.0878 | 0.0740 | -2 | -3 | -8.4 | 15.01 | |||||||||||||||||||||||||||||||||||||||
| c.212A>T | D71V 2D  AIThe SynGAP1 D71V missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely benign classification (3 benign vs. 1 pathogenic votes). High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.575842 | Disordered | 0.456046 | Uncertain | 0.350 | 0.799 | 0.375 | -3.549 | Likely Benign | 0.780 | Likely Pathogenic | Likely Benign | 0.183 | Likely Benign | -2.28 | Neutral | 0.334 | Benign | 0.060 | Benign | 4.01 | Benign | 0.00 | Affected | 0.0771 | 0.6197 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.287G>T | G96V 2D AIThe SynGAP1 missense variant G96V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. The variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.657645 | Disordered | 0.599491 | Binding | 0.335 | 0.871 | 0.625 | -4.266 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.109 | Likely Benign | -1.43 | Neutral | 0.334 | Benign | 0.029 | Benign | 4.18 | Benign | 0.00 | Affected | 0.1575 | 0.3833 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.4018A>C | T1340P 2D AIThe SynGAP1 missense variant T1340P is catalogued in gnomAD (ID 6‑33451892‑A‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes, while only SIFT predicts a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that T1340P is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.977899 | Binding | 0.444 | 0.697 | 0.750 | 6-33451892-A-C | -2.681 | Likely Benign | 0.102 | Likely Benign | Likely Benign | 0.190 | Likely Benign | -1.81 | Neutral | 0.334 | Benign | 0.099 | Benign | 4.08 | Benign | 0.01 | Affected | 3.77 | 5 | 0.2268 | 0.4983 | -1 | 0 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||
| c.4019C>T | T1340I 2D AIThe SynGAP1 missense variant T1340I is not reported in ClinVar (ClinVar status: “None”) but is present in gnomAD (ID 6‑33451893‑C‑T). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN and SIFT. AlphaMissense‑Default is uncertain, while AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to benign (two benign versus one pathogenic, with the uncertain result treated as unavailable). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the high‑accuracy predictions (AlphaMissense‑Optimized, SGM Consensus) both indicate a benign impact, and no evidence contradicts this assessment with the ClinVar status. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.977899 | Binding | 0.444 | 0.697 | 0.750 | 6-33451893-C-T | -3.476 | Likely Benign | 0.402 | Ambiguous | Likely Benign | 0.089 | Likely Benign | -2.57 | Deleterious | 0.334 | Benign | 0.099 | Benign | 4.08 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1163 | 0.5848 | -1 | 0 | 5.2 | 12.05 | |||||||||||||||||||||||||||||||||||||
| c.1199T>G | V400G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V400G missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect are polyPhen‑2 HumDiv and FATHMM; all other evaluated algorithms (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, is pathogenic. No prediction or stability result is missing or inconclusive. Based on the overwhelming agreement among both general and high‑accuracy tools, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.398279 | Structured | 0.415488 | Uncertain | 0.951 | 0.451 | 0.000 | -11.508 | Likely Pathogenic | 0.969 | Likely Pathogenic | Likely Pathogenic | 4.84 | Destabilizing | 0.1 | 5.40 | Destabilizing | 5.12 | Destabilizing | 2.40 | Destabilizing | 0.819 | Likely Pathogenic | -5.70 | Deleterious | 0.335 | Benign | 0.920 | Probably Damaging | 5.27 | Benign | 0.00 | Affected | 0.2296 | 0.2522 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1852C>A | Q618K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q618K is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33440904‑C‑A). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while ESM1b is uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign result; and Foldetta also predicts benign stability. No predictions or folding stability results are missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.138725 | Uncertain | 0.904 | 0.240 | 0.000 | 6-33440904-C-A | 24 | 1.49e-5 | -7.708 | In-Between | 0.229 | Likely Benign | Likely Benign | 0.02 | Likely Benign | 0.0 | 0.16 | Likely Benign | 0.09 | Likely Benign | -0.46 | Likely Benign | 0.281 | Likely Benign | -0.05 | Neutral | 0.338 | Benign | 0.111 | Benign | -1.21 | Pathogenic | 0.29 | Tolerated | 3.37 | 35 | 0.1413 | 0.2750 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||
| c.1999A>G | I667V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I667V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Only ESM1b predicts a pathogenic outcome; all other tools (FoldX, Rosetta, premPS, AlphaMissense‑Default, Foldetta) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority vote. Foldetta’s stability prediction is uncertain. Overall, the preponderance of evidence points to a benign effect. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.142424 | Structured | 0.083597 | Uncertain | 0.927 | 0.379 | 0.000 | -8.482 | Likely Pathogenic | 0.406 | Ambiguous | Likely Benign | 1.32 | Ambiguous | 0.0 | 1.06 | Ambiguous | 1.19 | Ambiguous | 0.85 | Ambiguous | 0.234 | Likely Benign | -0.86 | Neutral | 0.341 | Benign | 0.052 | Benign | 3.35 | Benign | 0.15 | Tolerated | 0.1178 | 0.3059 | 4 | 3 | -0.3 | -14.03 | ||||||||||||||||||||||||||||||
| c.2200C>T | P734S 2D AIThe SynGAP1 missense variant P734S is listed in ClinVar with an uncertain significance (ClinVar ID 2283225.0) and is present in the gnomAD database (gnomAD ID 6‑33441665‑C‑T). Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign effects. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this benign assessment: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status is unavailable. Overall, the computational evidence strongly supports a benign classification, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.879233 | Disordered | 0.411273 | Uncertain | 0.368 | 0.721 | 0.875 | Uncertain | 2 | 6-33441665-C-T | 2 | 1.24e-6 | -4.291 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.030 | Likely Benign | -2.44 | Neutral | 0.344 | Benign | 0.048 | Benign | 2.77 | Benign | 0.11 | Tolerated | 3.64 | 6 | 0.3775 | 0.3650 | 1 | -1 | 0.8 | -10.04 | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||||||||||||||||
| c.1211C>T | A404V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A404V missense variant is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438116‑C‑T). Prediction tools that agree on a benign effect include REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and ESM1b. Four tools (FoldX, Rosetta, AlphaMissense‑Default, Foldetta) give uncertain or inconclusive results. High‑accuracy methods give mixed evidence: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta remains uncertain. Overall, the majority of predictions lean toward a benign impact, though a key consensus method suggests pathogenicity, leaving the variant’s effect unresolved. **The variant is most likely benign based on the prevailing predictions, and this assessment does not contradict ClinVar status, as no ClinVar classification exists.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.232838 | Structured | 0.415505 | Uncertain | 0.965 | 0.355 | 0.000 | 6-33438116-C-T | 1 | 6.20e-7 | -8.219 | Likely Pathogenic | 0.343 | Ambiguous | Likely Benign | 0.56 | Ambiguous | 0.3 | -1.55 | Ambiguous | -0.50 | Ambiguous | -0.05 | Likely Benign | 0.118 | Likely Benign | -2.96 | Deleterious | 0.345 | Benign | 0.018 | Benign | 4.20 | Benign | 0.47 | Tolerated | 3.38 | 28 | 0.1288 | 0.6842 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||
| c.1328G>A | G443D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G443D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome. Uncertain results come from Rosetta and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely benign, and Foldetta as inconclusive. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, so there is no contradiction with existing clinical data. The variant is most likely benign based on the current predictive landscape. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.250310 | Structured | 0.258623 | Uncertain | 0.935 | 0.206 | 0.000 | -6.818 | Likely Benign | 0.761 | Likely Pathogenic | Likely Benign | -0.19 | Likely Benign | 0.2 | -1.72 | Ambiguous | -0.96 | Ambiguous | 0.32 | Likely Benign | 0.072 | Likely Benign | -0.50 | Neutral | 0.345 | Benign | 0.072 | Benign | 3.63 | Benign | 0.43 | Tolerated | 0.1687 | 0.2090 | 1 | -1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||
| c.2116G>A | E706K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E706K is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM all classify the change as benign. In contrast, ESM1b and AlphaMissense‑Default predict a pathogenic impact. Tools that return uncertain results—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—do not provide decisive evidence. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign calls). High‑accuracy assessments are likewise ambiguous: AlphaMissense‑Optimized is uncertain, Foldetta is uncertain, and the SGM Consensus remains inconclusive. Overall, the preponderance of evidence points to a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.377033 | Uncertain | 0.929 | 0.363 | 0.000 | Uncertain | 1 | -10.519 | Likely Pathogenic | 0.833 | Likely Pathogenic | Ambiguous | 1.17 | Ambiguous | 0.1 | 0.51 | Ambiguous | 0.84 | Ambiguous | 0.08 | Likely Benign | 0.080 | Likely Benign | -1.51 | Neutral | 0.345 | Benign | 0.028 | Benign | 4.15 | Benign | 0.52 | Tolerated | 3.47 | 10 | 0.2065 | 0.4511 | 0 | 1 | -0.4 | -0.94 | 187.1 | 49.2 | 0.0 | 0.0 | 0.4 | 0.1 | X | Uncertain | The carboxylate side chain of Glu706, located at the end and outer surface of an α-helix (res. Thr704-Gly712), forms a salt bridge with Lys710 and a hydrogen bond with its own backbone amino group at the helix end in the WT simulations. Although Lys706 is unable to make these transient interactions in the variant simulations, there is no apparent negative effect on the protein structure due to the residue swap. However, because the model ends abruptly at the C-terminus, no definite conclusions can be drawn based on the simulations. | |||||||||||||||||
| c.224A>C | E75A 2D AIThe SynGAP1 missense variant E75A is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic call comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the consensus of the majority of tools and the high‑accuracy predictions point to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.595080 | Disordered | 0.443881 | Uncertain | 0.303 | 0.822 | 0.500 | -3.111 | Likely Benign | 0.194 | Likely Benign | Likely Benign | 0.055 | Likely Benign | -1.19 | Neutral | 0.345 | Benign | 0.021 | Benign | 4.05 | Benign | 0.00 | Affected | 0.4248 | 0.6413 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.224A>G | E75G 2D  AIThe SynGAP1 missense variant E75G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels it “Likely Benign.” No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the preponderance of evidence indicates that E75G is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.595080 | Disordered | 0.443881 | Uncertain | 0.303 | 0.822 | 0.500 | -2.991 | Likely Benign | 0.175 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -1.74 | Neutral | 0.345 | Benign | 0.023 | Benign | 4.01 | Benign | 0.00 | Affected | 0.3078 | 0.5738 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||||||
| c.4007A>C | E1336A 2D AIThe SynGAP1 missense variant E1336A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of evidence (five benign vs. three pathogenic) supports a benign classification. This prediction does not contradict any ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | -3.545 | Likely Benign | 0.879 | Likely Pathogenic | Ambiguous | 0.191 | Likely Benign | -3.78 | Deleterious | 0.345 | Benign | 0.099 | Benign | 3.22 | Benign | 0.00 | Affected | 0.4000 | 0.7385 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.4007A>G | E1336G 2D AIThe SynGAP1 missense variant E1336G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized result is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2). Foldetta, which would assess protein‑folding stability, has no available result for this variant. Overall, the balance of evidence (five benign vs three pathogenic predictions) suggests the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | -3.574 | Likely Benign | 0.932 | Likely Pathogenic | Ambiguous | 0.211 | Likely Benign | -4.36 | Deleterious | 0.345 | Benign | 0.109 | Benign | 3.20 | Benign | 0.00 | Affected | 0.3092 | 0.6170 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.1108G>T | G370C 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G370C has no ClinVar entry and is not reported in gnomAD. Functional prediction tools fall into two groups: benign predictions come from premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from REVEL, FoldX, Rosetta, Foldetta, and FATHMM. ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. No prediction or stability result is missing. Overall, the majority of tools predict a benign effect, and the high‑accuracy consensus also leans benign, while only one high‑accuracy method (Foldetta) suggests pathogenicity. Thus, the variant is most likely benign based on the available predictions, and this assessment does not contradict any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.461924 | Structured | 0.434325 | Uncertain | 0.359 | 0.720 | 0.500 | -7.071 | In-Between | 0.119 | Likely Benign | Likely Benign | 3.01 | Destabilizing | 2.1 | 2.03 | Destabilizing | 2.52 | Destabilizing | 0.29 | Likely Benign | 0.511 | Likely Pathogenic | -1.00 | Neutral | 0.353 | Benign | 0.010 | Benign | 1.32 | Pathogenic | 0.06 | Tolerated | 0.1245 | 0.4412 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||
| c.1261G>A | A421T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A421T is not reported in ClinVar and is present in gnomAD (ID 6‑33438166‑G‑A). Functional prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; the SGM‑Consensus score is “Likely Pathogenic.” Uncertain results from FoldX and premPS are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as benign. Overall, the majority of consensus predictions lean toward a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.404927 | Uncertain | 0.965 | 0.257 | 0.000 | 6-33438166-G-A | 1 | 6.19e-7 | -9.217 | Likely Pathogenic | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.75 | Ambiguous | 0.2 | 0.18 | Likely Benign | 0.47 | Likely Benign | 0.99 | Ambiguous | 0.179 | Likely Benign | -3.12 | Deleterious | 0.353 | Benign | 0.136 | Benign | 3.43 | Benign | 0.09 | Tolerated | 3.37 | 29 | 0.1346 | 0.4439 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||
| c.1402A>C | M468L 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M468L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumVar and FATHMM. The SGM‑Consensus, which is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments—all available—show benign predictions: AlphaMissense‑Optimized (Benign), SGM‑Consensus (Likely Benign), and Foldetta (Benign). Based on the overall consensus of the majority of tools and the high‑accuracy predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.284882 | Structured | 0.339253 | Uncertain | 0.932 | 0.257 | 0.000 | -6.886 | Likely Benign | 0.290 | Likely Benign | Likely Benign | 0.43 | Likely Benign | 0.1 | -0.38 | Likely Benign | 0.03 | Likely Benign | -0.35 | Likely Benign | 0.412 | Likely Benign | 0.19 | Neutral | 0.359 | Benign | 0.654 | Possibly Damaging | -0.73 | Pathogenic | 1.00 | Tolerated | 0.1564 | 0.4408 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||
| c.1402A>T | M468L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M468L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumVar and FATHMM. The SGM‑Consensus, which is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments—AlphaMissense‑Optimized, the SGM‑Consensus, and Foldetta (combining FoldX‑MD and Rosetta outputs)—all indicate a benign outcome. No prediction or folding‑stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.284882 | Structured | 0.339253 | Uncertain | 0.932 | 0.257 | 0.000 | -6.886 | Likely Benign | 0.290 | Likely Benign | Likely Benign | 0.43 | Likely Benign | 0.1 | -0.38 | Likely Benign | 0.03 | Likely Benign | -0.35 | Likely Benign | 0.412 | Likely Benign | 0.19 | Neutral | 0.359 | Benign | 0.654 | Possibly Damaging | -0.73 | Pathogenic | 1.00 | Tolerated | 0.1564 | 0.4408 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||
| c.220A>C | S74R 2D  AIThe SynGAP1 missense variant S74R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority of the high‑accuracy tools) also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign effect, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.505461 | Disordered | 0.450156 | Uncertain | 0.294 | 0.831 | 0.500 | -3.271 | Likely Benign | 0.418 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -1.34 | Neutral | 0.361 | Benign | 0.019 | Benign | 4.08 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0943 | 0.3562 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.222C>A | S74R 2D AIThe SynGAP1 missense variant S74R is catalogued in gnomAD (ID 6‑33425830‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all report benign or likely benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments confirm the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus, derived from the majority of the high‑confidence predictors, is benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that S74R is most likely benign, and this assessment does not contradict any ClinVar status, as none is reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.505461 | Disordered | 0.450156 | Uncertain | 0.294 | 0.831 | 0.500 | 6-33425830-C-A | 1 | 6.20e-7 | -3.271 | Likely Benign | 0.418 | Ambiguous | Likely Benign | 0.070 | Likely Benign | -1.34 | Neutral | 0.361 | Benign | 0.019 | Benign | 4.08 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0943 | 0.3562 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||
| c.222C>G | S74R 2D AIThe SynGAP1 missense variant S74R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority of the high‑accuracy tools) is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.505461 | Disordered | 0.450156 | Uncertain | 0.294 | 0.831 | 0.500 | -3.271 | Likely Benign | 0.418 | Ambiguous | Likely Benign | 0.070 | Likely Benign | -1.34 | Neutral | 0.361 | Benign | 0.019 | Benign | 4.08 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0943 | 0.3562 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.278G>C | R93P 2D AIThe SynGAP1 missense variant R93P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus indicates Likely Benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.549151 | Binding | 0.290 | 0.874 | 0.625 | -3.164 | Likely Benign | 0.473 | Ambiguous | Likely Benign | 0.121 | Likely Benign | -0.32 | Neutral | 0.361 | Benign | 0.038 | Benign | 3.99 | Benign | 0.00 | Affected | 0.2200 | 0.4844 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3497C>G | A1166G 2D AISynGAP1 missense variant A1166G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools and the high‑accuracy predictions indicate that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.575842 | Disordered | 0.811691 | Binding | 0.381 | 0.803 | 0.375 | -3.679 | Likely Benign | 0.745 | Likely Pathogenic | Likely Benign | 0.355 | Likely Benign | -1.17 | Neutral | 0.361 | Benign | 0.307 | Benign | 5.34 | Benign | 0.31 | Tolerated | 0.2244 | 0.3977 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3931C>G | L1311V 2D AIThe SynGAP1 missense variant L1311V is catalogued in gnomAD (ID 6‑33451805‑C‑G) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status remains unavailable. Overall, the computational evidence overwhelmingly supports a benign impact, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.968153 | Binding | 0.393 | 0.907 | 0.750 | 6-33451805-C-G | 1 | 6.20e-7 | -3.645 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 0.025 | Likely Benign | -0.05 | Neutral | 0.362 | Benign | 0.193 | Benign | 2.89 | Benign | 0.32 | Tolerated | 3.77 | 5 | 0.1666 | 0.3518 | 1 | 2 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||
| c.364C>G | P122A 2D AIThe SynGAP1 missense variant P122A is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors—REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the substitution as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods likewise support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.672358 | Binding | 0.400 | 0.887 | 0.750 | -3.105 | Likely Benign | 0.060 | Likely Benign | Likely Benign | 0.086 | Likely Benign | -1.66 | Neutral | 0.363 | Benign | 0.096 | Benign | 4.27 | Benign | 1.00 | Tolerated | 0.3991 | 0.4286 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.682A>G | T228A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T228A variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and the SGM‑Consensus score (which is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are REVEL, SIFT, and AlphaMissense‑Default. Predictions that are inconclusive or uncertain are Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as benign (majority of its constituent predictors are benign), and Foldetta as uncertain. Overall, the balance of evidence favors a benign interpretation; this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.104810 | Structured | 0.321733 | Uncertain | 0.829 | 0.316 | 0.125 | -5.190 | Likely Benign | 0.793 | Likely Pathogenic | Ambiguous | 0.44 | Likely Benign | 0.0 | 0.82 | Ambiguous | 0.63 | Ambiguous | 0.58 | Ambiguous | 0.520 | Likely Pathogenic | -2.30 | Neutral | 0.363 | Benign | 0.138 | Benign | 5.63 | Benign | 0.04 | Affected | 0.4072 | 0.5014 | 1 | 0 | 2.5 | -30.03 | |||||||||||||||||||||||||||||
| c.286G>A | G96S 2D AIThe SynGAP1 missense variant G96S is listed in ClinVar with an “Uncertain” status and is present in gnomAD (variant ID 6‑33425894‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.657645 | Disordered | 0.599491 | Binding | 0.335 | 0.871 | 0.625 | Uncertain | 1 | 6-33425894-G-A | 5 | 3.10e-6 | -3.049 | Likely Benign | 0.065 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -0.76 | Neutral | 0.364 | Benign | 0.008 | Benign | 4.25 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3001 | 0.5336 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||
| c.1490A>T | Y497F 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y497F is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus, REVEL, PROVEAN, ESM1b, and FATHMM; premPS remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of predictions lean toward a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Thus, the variant is most likely benign based on the current predictive evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.393608 | Uncertain | 0.950 | 0.181 | 0.000 | -8.566 | Likely Pathogenic | 0.199 | Likely Benign | Likely Benign | -0.27 | Likely Benign | 0.1 | 0.47 | Likely Benign | 0.10 | Likely Benign | 0.61 | Ambiguous | 0.578 | Likely Pathogenic | -3.75 | Deleterious | 0.367 | Benign | 0.131 | Benign | -1.15 | Pathogenic | 0.19 | Tolerated | 0.2074 | 0.2242 | 7 | 3 | 4.1 | -16.00 | |||||||||||||||||||||||||||||
| c.1262C>A | A421E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A421E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM, whereas pathogenic calls are made by ESM1b, PROVEAN, AlphaMissense‑Default, AlphaMissense‑Optimized, Rosetta, premPS, and the SGM‑Consensus score (Likely Pathogenic). Stability‑based methods give mixed results: FoldX is uncertain, Foldetta is uncertain, and Rosetta alone predicts pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta remains uncertain. Overall, the majority of evidence, including the high‑accuracy tools, points to a pathogenic impact for A421E, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.404927 | Uncertain | 0.965 | 0.257 | 0.000 | -11.993 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.63 | Ambiguous | 0.2 | 2.07 | Destabilizing | 1.35 | Ambiguous | 1.30 | Destabilizing | 0.233 | Likely Benign | -4.24 | Deleterious | 0.368 | Benign | 0.144 | Benign | 3.44 | Benign | 0.14 | Tolerated | 0.1288 | 0.1830 | 0 | -1 | -5.3 | 58.04 | |||||||||||||||||||||||||||||
| c.2501T>A | M834K 2D AIThe SynGAP1 missense variant M834K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.585406 | Disordered | 0.640801 | Binding | 0.258 | 0.863 | 0.375 | -3.154 | Likely Benign | 0.453 | Ambiguous | Likely Benign | 0.154 | Likely Benign | -2.21 | Neutral | 0.369 | Benign | 0.150 | Benign | 2.43 | Pathogenic | 0.00 | Affected | 0.1131 | 0.0688 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||||||||||||
| c.2653C>A | P885T 2D AIThe SynGAP1 missense variant P885T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.690604 | Disordered | 0.636133 | Binding | 0.344 | 0.917 | 0.250 | -5.152 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -1.44 | Neutral | 0.369 | Benign | 0.171 | Benign | 2.78 | Benign | 0.00 | Affected | 0.1438 | 0.6484 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2653C>T | P885S 2D AIThe SynGAP1 missense variant P885S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.690604 | Disordered | 0.636133 | Binding | 0.344 | 0.917 | 0.250 | -4.089 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.037 | Likely Benign | -1.10 | Neutral | 0.369 | Benign | 0.222 | Benign | 2.87 | Benign | 0.00 | Affected | 0.3363 | 0.5881 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2656G>A | A886T 2D AIThe SynGAP1 missense variant A886T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while only SIFT and FATHMM predict pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.716283 | Disordered | 0.619166 | Binding | 0.359 | 0.922 | 0.500 | -4.319 | Likely Benign | 0.075 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.78 | Neutral | 0.369 | Benign | 0.237 | Benign | 2.21 | Pathogenic | 0.00 | Affected | 0.1581 | 0.6737 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2866T>A | S956T 2D AIThe SynGAP1 missense variant S956T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact for S956T, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984871 | Disordered | 0.957345 | Binding | 0.364 | 0.917 | 0.750 | -5.404 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.079 | Likely Benign | -0.45 | Neutral | 0.369 | Benign | 0.159 | Benign | 2.00 | Pathogenic | 0.08 | Tolerated | 0.2244 | 0.5727 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2887C>A | H963N 2D AIThe SynGAP1 missense variant H963N is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only ESM1b predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.983973 | Binding | 0.325 | 0.886 | 0.750 | -8.274 | Likely Pathogenic | 0.099 | Likely Benign | Likely Benign | 0.089 | Likely Benign | -0.21 | Neutral | 0.369 | Benign | 0.120 | Benign | 4.18 | Benign | 0.16 | Tolerated | 0.2094 | 0.3596 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.2887C>G | H963D 2D  AIThe SynGAP1 missense variant H963D is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic prediction comes from ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” status. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign; Foldetta results are not available. Overall, the consensus of the majority of tools and the high‑accuracy predictions point to a benign impact for H963D, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.983973 | Binding | 0.325 | 0.886 | 0.750 | -12.082 | Likely Pathogenic | 0.204 | Likely Benign | Likely Benign | 0.167 | Likely Benign | -0.81 | Neutral | 0.369 | Benign | 0.159 | Benign | 4.16 | Benign | 0.46 | Tolerated | 0.2482 | 0.3023 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.2990C>T | A997V 2D AIThe SynGAP1 missense variant A997V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. The prediction is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.948624 | Binding | 0.273 | 0.901 | 0.500 | -4.504 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.052 | Likely Benign | -1.01 | Neutral | 0.369 | Benign | 0.120 | Benign | 4.15 | Benign | 0.00 | Affected | 0.1232 | 0.6335 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||||||||||||
| c.3034C>A | P1012T 2D AIThe SynGAP1 missense variant P1012T is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.894674 | Binding | 0.319 | 0.866 | 0.625 | -4.788 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.052 | Likely Benign | -0.56 | Neutral | 0.369 | Benign | 0.171 | Benign | 2.84 | Benign | 0.14 | Tolerated | 0.1399 | 0.5730 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3125A>T | Q1042L 2D AIThe SynGAP1 missense variant Q1042L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.959333 | Binding | 0.310 | 0.846 | 0.625 | -3.796 | Likely Benign | 0.203 | Likely Benign | Likely Benign | 0.338 | Likely Benign | -2.47 | Neutral | 0.369 | Benign | 0.120 | Benign | 5.47 | Benign | 0.05 | Affected | 0.1469 | 0.6276 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3293G>A | S1098N 2D AIThe SynGAP1 missense variant S1098N is listed in ClinVar (ID 864704.0) with an “Uncertain” clinical significance and is present in gnomAD (variant ID 6‑33443845‑G‑A). All evaluated in‑silico predictors agree on a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores. No tool predicts pathogenicity. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status is unavailable. Overall, the computational evidence strongly supports a benign classification, which is consistent with the ClinVar “Uncertain” status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.973030 | Binding | 0.337 | 0.855 | 1.000 | Conflicting | 2 | 6-33443845-G-A | 6 | 3.89e-6 | -5.120 | Likely Benign | 0.156 | Likely Benign | Likely Benign | 0.063 | Likely Benign | -0.58 | Neutral | 0.369 | Benign | 0.120 | Benign | 2.76 | Benign | 0.36 | Tolerated | 3.77 | 5 | 0.1665 | 0.5145 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||
| c.3839T>C | M1280T 2D AIThe SynGAP1 missense variant M1280T is catalogued in gnomAD (6‑33447887‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign or likely benign outcome. In contrast, SIFT and FATHMM predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic designation. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | 6-33447887-T-C | 3 | 1.93e-6 | -2.305 | Likely Benign | 0.112 | Likely Benign | Likely Benign | 0.158 | Likely Benign | -2.26 | Neutral | 0.369 | Benign | 0.120 | Benign | 2.34 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1814 | 0.2031 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||
| c.3845A>T | E1282V 2D  AIThe SynGAP1 missense variant E1282V is reported in gnomAD (ID 6‑33447893‑A‑T) but has no ClinVar entry. In silico prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority‑vote method) is benign; Foldetta results are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.817364 | Binding | 0.465 | 0.725 | 0.875 | 6-33447893-A-T | -2.790 | Likely Benign | 0.190 | Likely Benign | Likely Benign | 0.102 | Likely Benign | -1.71 | Neutral | 0.369 | Benign | 0.078 | Benign | 2.72 | Benign | 0.04 | Affected | 0.0656 | 0.5905 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3866A>C | K1289T 2D AIThe SynGAP1 missense variant K1289T is not reported in ClinVar and is absent from gnomAD. Consensus prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign effect. No Foldetta stability prediction is available for this residue. Overall, the majority of computational evidence points to a benign impact, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.828700 | Binding | 0.548 | 0.787 | 0.625 | -3.618 | Likely Benign | 0.250 | Likely Benign | Likely Benign | 0.094 | Likely Benign | -2.18 | Neutral | 0.369 | Benign | 0.120 | Benign | 2.58 | Benign | 0.02 | Affected | 0.2047 | 0.2447 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.235A>G | N79D 2D AIThe SynGAP1 missense variant N79D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.457064 | Uncertain | 0.290 | 0.876 | 0.375 | -3.746 | Likely Benign | 0.281 | Likely Benign | Likely Benign | 0.021 | Likely Benign | -0.92 | Neutral | 0.371 | Benign | 0.018 | Benign | 4.19 | Benign | 0.00 | Affected | 0.1893 | 0.2535 | 2 | 1 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.236A>C | N79T 2D AIThe SynGAP1 missense variant N79T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.457064 | Uncertain | 0.290 | 0.876 | 0.375 | -3.752 | Likely Benign | 0.102 | Likely Benign | Likely Benign | 0.022 | Likely Benign | -0.61 | Neutral | 0.371 | Benign | 0.018 | Benign | 4.21 | Benign | 0.00 | Affected | 0.1133 | 0.4890 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||||||||||||
| c.244C>G | L82V 2D AIThe SynGAP1 missense variant L82V is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (ID 6‑33425852‑C‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and no Foldetta stability result is available. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments therefore indicate a benign likelihood: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus is likely benign, and Foldetta data are missing. Overall, the majority of predictions support a benign impact, and this is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.517720 | Binding | 0.284 | 0.890 | 0.375 | 6-33425852-C-G | -6.701 | Likely Benign | 0.914 | Likely Pathogenic | Ambiguous | 0.065 | Likely Benign | -1.13 | Neutral | 0.371 | Benign | 0.024 | Benign | 3.72 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1467 | 0.2353 | 1 | 2 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||
| c.2854G>T | G952C 2D AIThe SynGAP1 missense variant G952C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.910621 | Binding | 0.341 | 0.926 | 0.750 | -8.599 | Likely Pathogenic | 0.088 | Likely Benign | Likely Benign | 0.272 | Likely Benign | -0.18 | Neutral | 0.371 | Benign | 0.169 | Benign | 3.20 | Benign | 0.01 | Affected | 0.1458 | 0.4439 | -3 | -3 | 2.9 | 46.09 | |||||||||||||||||||||||||||||||||||||||
| c.4025A>C | D1342A 2D AIThe SynGAP1 missense variant D1342A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports a benign effect, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are unavailable. Overall, the preponderance of evidence supports a benign classification for D1342A, and this conclusion does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.981682 | Binding | 0.316 | 0.678 | 0.875 | -2.719 | Likely Benign | 0.145 | Likely Benign | Likely Benign | 0.049 | Likely Benign | -0.58 | Neutral | 0.371 | Benign | 0.084 | Benign | 4.06 | Benign | 0.02 | Affected | 0.4270 | 0.5676 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||||
| c.55G>A | A19T 2D AIThe SynGAP1 missense variant A19T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. The variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.443393 | Uncertain | 0.378 | 0.906 | 0.500 | -3.422 | Likely Benign | 0.131 | Likely Benign | Likely Benign | 0.030 | Likely Benign | -0.02 | Neutral | 0.371 | Benign | 0.036 | Benign | 4.14 | Benign | 0.00 | Affected | 0.2357 | 0.7248 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.56C>G | A19G 2D AIThe SynGAP1 missense variant A19G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. The prediction is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.443393 | Uncertain | 0.378 | 0.906 | 0.500 | -3.525 | Likely Benign | 0.194 | Likely Benign | Likely Benign | 0.037 | Likely Benign | -0.45 | Neutral | 0.371 | Benign | 0.036 | Benign | 4.10 | Benign | 0.00 | Affected | 0.2324 | 0.5447 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.56C>T | A19V 2D AIThe SynGAP1 missense variant A19V is reported in gnomAD (ID 6‑33420320‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign; Foldetta results are unavailable. Overall, the consensus of available predictions points to a benign impact for A19V, and this conclusion is not contradicted by ClinVar status, which currently lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.443393 | Uncertain | 0.378 | 0.906 | 0.500 | 6-33420320-C-T | -3.157 | Likely Benign | 0.095 | Likely Benign | Likely Benign | 0.063 | Likely Benign | 0.42 | Neutral | 0.371 | Benign | 0.036 | Benign | 4.27 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1804 | 0.6942 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||
| c.1346G>A | S449N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S449N is not reported in ClinVar (ClinVar status: not listed) but is present in the gnomAD database (gnomAD ID: 6‑33438251‑G‑A). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; the only inconclusive results come from premPS (uncertain) and ESM1b (uncertain). High‑accuracy assessments reinforce the benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is benign. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.254060 | Structured | 0.301437 | Uncertain | 0.958 | 0.251 | 0.000 | 6-33438251-G-A | 1 | 6.19e-7 | -7.692 | In-Between | 0.210 | Likely Benign | Likely Benign | 0.38 | Likely Benign | 0.1 | -0.03 | Likely Benign | 0.18 | Likely Benign | 0.81 | Ambiguous | 0.070 | Likely Benign | -2.31 | Neutral | 0.372 | Benign | 0.026 | Benign | 3.37 | Benign | 0.18 | Tolerated | 3.37 | 32 | 0.1085 | 0.3767 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||
| c.329T>G | V110G 2D AIThe SynGAP1 missense variant V110G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.665934 | Binding | 0.347 | 0.860 | 0.750 | -4.012 | Likely Benign | 0.724 | Likely Pathogenic | Likely Benign | 0.162 | Likely Benign | -2.87 | Deleterious | 0.377 | Benign | 0.928 | Probably Damaging | 4.06 | Benign | 0.00 | Affected | 0.2302 | 0.2671 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.325A>G | S109G 2D AIThe SynGAP1 missense variant S109G is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority of the four high‑accuracy tools) also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.622677 | Disordered | 0.669335 | Binding | 0.328 | 0.864 | 0.750 | -3.918 | Likely Benign | 0.549 | Ambiguous | Likely Benign | 0.132 | Likely Benign | -1.95 | Neutral | 0.378 | Benign | 0.067 | Benign | 3.51 | Benign | 0.00 | Affected | 0.2686 | 0.3779 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.400A>C | S134R 2D AIThe SynGAP1 missense variant S134R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -9.053 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.292 | Likely Benign | -2.54 | Deleterious | 0.380 | Benign | 0.147 | Benign | 3.84 | Benign | 0.00 | Affected | 0.0719 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.402C>A | S134R 2D AIThe SynGAP1 missense variant S134R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict pathogenicity are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized also predicts Pathogenic, while the SGM‑Consensus remains Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -9.053 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.146 | Likely Benign | -2.54 | Deleterious | 0.380 | Benign | 0.147 | Benign | 3.84 | Benign | 0.00 | Affected | 0.0719 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.402C>G | S134R 2D AIThe SynGAP1 missense variant S134R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict pathogenicity are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized also predicts Pathogenic, while the SGM‑Consensus remains Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -9.053 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.146 | Likely Benign | -2.54 | Deleterious | 0.380 | Benign | 0.147 | Benign | 3.84 | Benign | 0.00 | Affected | 0.0719 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.415A>C | S139R 2D AIThe SynGAP1 missense variant S139R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that agree on a pathogenic effect are AlphaMissense‑Default and AlphaMissense‑Optimized; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts benign. Foldetta results are unavailable. Overall, the majority of conventional predictors and the SGM Consensus favor a benign interpretation, whereas AlphaMissense‑Optimized suggests pathogenicity. No ClinVar annotation exists to contradict these predictions. Thus, based on the available computational evidence, the variant is most likely benign, though the AlphaMissense‑Optimized prediction introduces some uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.553315 | Disordered | 0.600637 | Binding | 0.353 | 0.900 | 0.250 | -7.547 | In-Between | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.112 | Likely Benign | -2.07 | Neutral | 0.380 | Benign | 0.136 | Benign | 4.16 | Benign | 0.12 | Tolerated | 0.1004 | 0.2971 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.416G>T | S139I 2D AIThe SynGAP1 missense variant S139I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; pathogenic predictions come from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy tools points to a pathogenic effect. This conclusion is consistent with the absence of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.553315 | Disordered | 0.600637 | Binding | 0.353 | 0.900 | 0.250 | -9.228 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 0.203 | Likely Benign | -3.36 | Deleterious | 0.380 | Benign | 0.102 | Benign | 4.09 | Benign | 0.03 | Affected | 0.0948 | 0.4796 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.417C>A | S139R 2D AIThe SynGAP1 missense variant S139R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are AlphaMissense‑Default and AlphaMissense‑Optimized; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence—including the consensus of high‑accuracy tools—suggests a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.553315 | Disordered | 0.600637 | Binding | 0.353 | 0.900 | 0.250 | -7.547 | In-Between | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.110 | Likely Benign | -2.07 | Neutral | 0.380 | Benign | 0.136 | Benign | 4.16 | Benign | 0.12 | Tolerated | 0.1004 | 0.2971 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.417C>G | S139R 2D AIThe SynGAP1 missense variant S139R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are AlphaMissense‑Default and AlphaMissense‑Optimized; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign. Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.553315 | Disordered | 0.600637 | Binding | 0.353 | 0.900 | 0.250 | -7.547 | In-Between | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.110 | Likely Benign | -2.07 | Neutral | 0.380 | Benign | 0.136 | Benign | 4.16 | Benign | 0.12 | Tolerated | 0.1004 | 0.2971 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.441A>C | Q147H 2D  AIThe SynGAP1 missense variant Q147H is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. Grouping by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Overall, the balance of evidence, especially the SGM Consensus and the majority of individual predictors, indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.541878 | Disordered | 0.503877 | Binding | 0.349 | 0.840 | 0.625 | -9.759 | Likely Pathogenic | 0.955 | Likely Pathogenic | Ambiguous | 0.102 | Likely Benign | -2.70 | Deleterious | 0.380 | Benign | 0.265 | Benign | 3.87 | Benign | 0.01 | Affected | 0.1528 | 0.3220 | 3 | 0 | 0.3 | 9.01 | |||||||||||||||||||||||||||||||||||||||
| c.441A>T | Q147H 2D AIThe SynGAP1 missense variant Q147H is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. Grouping by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Taken together, the balance of evidence favors a pathogenic interpretation, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.541878 | Disordered | 0.503877 | Binding | 0.349 | 0.840 | 0.625 | -9.759 | Likely Pathogenic | 0.955 | Likely Pathogenic | Ambiguous | 0.102 | Likely Benign | -2.70 | Deleterious | 0.380 | Benign | 0.265 | Benign | 3.87 | Benign | 0.01 | Affected | 0.1528 | 0.3220 | 3 | 0 | 0.3 | 9.01 | |||||||||||||||||||||||||||||||||||||||
| c.506A>T | D169V 2D AIThe SynGAP1 D169V missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. When predictions are grouped by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized returns an uncertain result, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this residue. Overall, the balance of evidence, particularly the SGM Consensus and the pathogenic calls from multiple independent predictors, indicates that D169V is most likely pathogenic, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.418646 | Structured | 0.497160 | Uncertain | 0.420 | 0.675 | 0.125 | -12.395 | Likely Pathogenic | 0.925 | Likely Pathogenic | Ambiguous | 0.243 | Likely Benign | -3.77 | Deleterious | 0.380 | Benign | 0.193 | Benign | 4.03 | Benign | 0.00 | Affected | 0.0895 | 0.7166 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.1102C>T | P368S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P368S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and FATHMM. The remaining methods (FoldX, Rosetta, Foldetta, premPS) yield uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is uncertain, so these do not alter the overall interpretation. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -4.790 | Likely Benign | 0.247 | Likely Benign | Likely Benign | 1.68 | Ambiguous | 0.4 | 1.60 | Ambiguous | 1.64 | Ambiguous | 0.52 | Ambiguous | 0.090 | Likely Benign | -5.12 | Deleterious | 0.384 | Benign | 0.113 | Benign | 1.80 | Pathogenic | 0.10 | Tolerated | 0.3700 | 0.5635 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||
| c.565C>A | P189T 2D AISynGAP1 missense variant P189T is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority‑vote method) reports it as likely pathogenic; Foldetta stability analysis is unavailable. Overall, the majority of computational evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.428590 | Uncertain | 0.331 | 0.602 | 0.250 | -8.622 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.265 | Likely Benign | -5.26 | Deleterious | 0.384 | Benign | 0.177 | Benign | 4.05 | Benign | 0.05 | Affected | 0.1603 | 0.6392 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.584C>T | A195V 2D AIThe SynGAP1 missense variant A195V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN and AlphaMissense‑Default. High‑accuracy methods give no definitive verdict: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign votes), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.346032 | Structured | 0.430388 | Uncertain | 0.363 | 0.533 | 0.125 | -5.830 | Likely Benign | 0.924 | Likely Pathogenic | Ambiguous | 0.210 | Likely Benign | -2.63 | Deleterious | 0.384 | Benign | 0.070 | Benign | 4.05 | Benign | 0.12 | Tolerated | 0.0782 | 0.5560 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||||||
| c.640C>G | L214V 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant L214V is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SGM‑Consensus, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The remaining methods—FoldX, Rosetta, premPS, and AlphaMissense‑Optimized—yield uncertain results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as uncertain. Overall, the majority of available predictions lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.155435 | Structured | 0.372604 | Uncertain | 0.818 | 0.302 | 0.125 | -9.913 | Likely Pathogenic | 0.950 | Likely Pathogenic | Ambiguous | 1.80 | Ambiguous | 0.4 | 0.54 | Ambiguous | 1.17 | Ambiguous | 0.89 | Ambiguous | 0.495 | Likely Benign | -2.54 | Deleterious | 0.384 | Benign | 0.070 | Benign | 5.78 | Benign | 0.01 | Affected | 0.1469 | 0.3887 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||
| c.748G>A | V250I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V250I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). All evaluated in silico predictors classify the change as benign: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). The high‑accuracy tools likewise predict a benign effect: AlphaMissense‑Optimized is benign, the SGM Consensus is “Likely Benign,” and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No pathogenic predictions are present. Therefore, based on the available computational evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.447574 | Structured | 0.244075 | Uncertain | 0.778 | 0.324 | 0.125 | -6.696 | Likely Benign | 0.085 | Likely Benign | Likely Benign | -0.44 | Likely Benign | 0.1 | -0.01 | Likely Benign | -0.23 | Likely Benign | 0.35 | Likely Benign | 0.362 | Likely Benign | -0.79 | Neutral | 0.384 | Benign | 0.070 | Benign | 6.09 | Benign | 0.13 | Tolerated | 0.0571 | 0.3644 | 4 | 3 | 0.3 | 14.03 | |||||||||||||||||||||||||||||
| c.753G>C | K251N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K251N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM (nine tools). Only AlphaMissense‑Default predicts a pathogenic outcome, while ESM1b and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign stability. Overall, the preponderance of evidence indicates that K251N is most likely benign, and this conclusion does not contradict the current ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.447574 | Structured | 0.226632 | Uncertain | 0.758 | 0.312 | 0.125 | -7.978 | In-Between | 0.930 | Likely Pathogenic | Ambiguous | 0.29 | Likely Benign | 0.1 | 0.21 | Likely Benign | 0.25 | Likely Benign | 0.32 | Likely Benign | 0.298 | Likely Benign | -1.29 | Neutral | 0.384 | Benign | 0.070 | Benign | 5.73 | Benign | 0.39 | Tolerated | 0.3848 | 0.1196 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.753G>T | K251N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K251N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM (nine tools). Only AlphaMissense‑Default predicts a pathogenic outcome, while ESM1b and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign stability. Overall, the preponderance of evidence indicates that K251N is most likely benign, and this conclusion does not contradict the current ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.447574 | Structured | 0.226632 | Uncertain | 0.758 | 0.312 | 0.125 | -7.978 | In-Between | 0.930 | Likely Pathogenic | Ambiguous | 0.29 | Likely Benign | 0.1 | 0.21 | Likely Benign | 0.25 | Likely Benign | 0.32 | Likely Benign | 0.298 | Likely Benign | -1.29 | Neutral | 0.384 | Benign | 0.070 | Benign | 5.73 | Benign | 0.39 | Tolerated | 0.3848 | 0.1196 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||
| c.754C>A | P252T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P252T is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify it as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar and FATHMM. Those that predict a pathogenic effect are REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized and the SGM‑Consensus score (Likely Pathogenic). Four tools (FoldX, Rosetta, Foldetta, premPS) give uncertain results and are treated as unavailable. High‑accuracy methods give the following: AlphaMissense‑Optimized predicts pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic; Foldetta’s stability assessment is uncertain. Overall, the majority of evidence points to a pathogenic effect, and this is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.461924 | Structured | 0.211606 | Uncertain | 0.753 | 0.304 | 0.250 | -11.824 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 1.24 | Ambiguous | 0.1 | 1.89 | Ambiguous | 1.57 | Ambiguous | 0.71 | Ambiguous | 0.828 | Likely Pathogenic | -7.35 | Deleterious | 0.384 | Benign | 0.177 | Benign | 5.78 | Benign | 0.01 | Affected | 0.1514 | 0.4842 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||
| c.762G>C | K254N 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K254N is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that classify the variant as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The majority of other in silico predictors—REVEL, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—indicate a pathogenic effect. Stability‑based methods FoldX, Rosetta, and Foldetta returned uncertain results and are therefore not considered evidence for or against pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as unavailable. Overall, the preponderance of evidence supports a pathogenic classification, which contradicts the current ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.513880 | Disordered | 0.207751 | Uncertain | 0.799 | 0.285 | 0.375 | Uncertain | 1 | -13.306 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.73 | Ambiguous | 0.2 | 1.87 | Ambiguous | 1.30 | Ambiguous | 1.19 | Destabilizing | 0.757 | Likely Pathogenic | -4.23 | Deleterious | 0.384 | Benign | 0.070 | Benign | 5.93 | Benign | 0.01 | Affected | 3.39 | 15 | 0.3200 | 0.1488 | 1 | 0 | 0.4 | -14.07 | 215.3 | -21.0 | -1.0 | 1.7 | 0.2 | 0.3 | X | Potentially Pathogenic | The amino group of Lys254, located in an α-β loop connecting the PH and C2 domains (res. Lys251-Arg258), forms salt bridges with the carboxylate groups of Glu244 and Asp684. Since the neutral carboxamide group of the Asn254 side chain cannot form salt bridges with acidic residues, the residue swap potentially weakens the tertiary structure assembly and/or influences the loop positioning. Regardless, in both the variant and WT simulations, all hydrogen bonds formed by the residue’s side chain were broken, and the residue rotated outwards. The partially α helical conformation of the loop, which extends to a nearby α helix (res. Met414-Asn426), is dynamic, making it unclear if the mutation affects it. | ||||||||||||||||
| c.762G>T | K254N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K254N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are REVEL, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus score, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Pathogenic” (3 pathogenic vs. 1 benign). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. No evidence from FoldX or Rosetta alone is conclusive. Based on the overall pattern of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not currently catalogued in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.513880 | Disordered | 0.207751 | Uncertain | 0.799 | 0.285 | 0.375 | -13.306 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.73 | Ambiguous | 0.2 | 1.87 | Ambiguous | 1.30 | Ambiguous | 1.19 | Destabilizing | 0.757 | Likely Pathogenic | -4.23 | Deleterious | 0.384 | Benign | 0.070 | Benign | 5.93 | Benign | 0.01 | Affected | 3.39 | 15 | 0.3200 | 0.1488 | 1 | 0 | 0.4 | -14.07 | 215.3 | -21.0 | -1.0 | 1.7 | 0.2 | 0.3 | X | Potentially Pathogenic | The amino group of Lys254, located in an α-β loop connecting the PH and C2 domains (res. Lys251-Arg258), forms salt bridges with the carboxylate groups of Glu244 and Asp684. Since the neutral carboxamide group of the Asn254 side chain cannot form salt bridges with acidic residues, the residue swap potentially weakens the tertiary structure assembly and/or influences the loop positioning. Regardless, in both the variant and WT simulations, all hydrogen bonds formed by the residue’s side chain were broken, and the residue rotated outwards. The partially α helical conformation of the loop, which extends to a nearby α helix (res. Met414-Asn426), is dynamic, making it unclear if the mutation affects it. | ||||||||||||||||||
| c.2029A>C | S677R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S677R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Those that predict a pathogenic effect are FoldX, ESM1b, and AlphaMissense‑Default. Uncertain results come from Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, the majority of tools and the high‑accuracy benign prediction suggest the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.194234 | Structured | 0.115685 | Uncertain | 0.555 | 0.338 | 0.125 | -9.388 | Likely Pathogenic | 0.714 | Likely Pathogenic | Likely Benign | 2.59 | Destabilizing | 0.1 | 0.73 | Ambiguous | 1.66 | Ambiguous | 0.71 | Ambiguous | 0.104 | Likely Benign | -2.49 | Neutral | 0.385 | Benign | 0.037 | Benign | 3.31 | Benign | 0.12 | Tolerated | 0.1120 | 0.4195 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2031T>A | S677R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S677R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are FoldX, ESM1b, and AlphaMissense‑Default. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and premPS. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is uncertain. Overall, the majority of consensus tools (six benign vs. three pathogenic) lean toward a benign interpretation. This prediction does not contradict any ClinVar status, as none is available. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.194234 | Structured | 0.115685 | Uncertain | 0.555 | 0.338 | 0.125 | -9.388 | Likely Pathogenic | 0.714 | Likely Pathogenic | Likely Benign | 2.59 | Destabilizing | 0.1 | 0.73 | Ambiguous | 1.66 | Ambiguous | 0.71 | Ambiguous | 0.150 | Likely Benign | -2.49 | Neutral | 0.385 | Benign | 0.037 | Benign | 3.31 | Benign | 0.12 | Tolerated | 0.1120 | 0.4195 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2031T>G | S677R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S677R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are FoldX, ESM1b, and AlphaMissense‑Default. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and premPS. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is uncertain. Overall, the majority of consensus tools (six benign vs. three pathogenic) lean toward a benign interpretation. This prediction does not contradict any ClinVar status, as none is available. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.194234 | Structured | 0.115685 | Uncertain | 0.555 | 0.338 | 0.125 | -9.388 | Likely Pathogenic | 0.714 | Likely Pathogenic | Likely Benign | 2.59 | Destabilizing | 0.1 | 0.73 | Ambiguous | 1.66 | Ambiguous | 0.71 | Ambiguous | 0.150 | Likely Benign | -2.49 | Neutral | 0.385 | Benign | 0.037 | Benign | 3.31 | Benign | 0.12 | Tolerated | 0.1120 | 0.4195 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.232C>A | R78S 2D AIThe SynGAP1 missense variant R78S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized returns an uncertain result. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments are: AlphaMissense‑Optimized (Uncertain), SGM‑Consensus (Likely Benign), and Foldetta (no data available). Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which contains no pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.505461 | Disordered | 0.448183 | Uncertain | 0.304 | 0.866 | 0.500 | -3.680 | Likely Benign | 0.792 | Likely Pathogenic | Ambiguous | 0.063 | Likely Benign | -1.11 | Neutral | 0.385 | Benign | 0.015 | Benign | 3.86 | Benign | 0.00 | Affected | 0.3081 | 0.3274 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||||
| c.232C>G | R78G 2D AIThe SynGAP1 R78G missense variant has no ClinVar entry and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign status: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Taken together, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.505461 | Disordered | 0.448183 | Uncertain | 0.304 | 0.866 | 0.500 | -3.972 | Likely Benign | 0.480 | Ambiguous | Likely Benign | 0.072 | Likely Benign | -1.94 | Neutral | 0.385 | Benign | 0.028 | Benign | 3.83 | Benign | 0.00 | Affected | 0.3285 | 0.2986 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.233G>T | R78L 2D AIThe SynGAP1 missense variant R78L is listed in ClinVar (ID 3390541.0) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score (which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as benign; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.505461 | Disordered | 0.448183 | Uncertain | 0.304 | 0.866 | 0.500 | Uncertain | 1 | -3.389 | Likely Benign | 0.635 | Likely Pathogenic | Likely Benign | 0.062 | Likely Benign | -1.59 | Neutral | 0.385 | Benign | 0.021 | Benign | 3.84 | Benign | 0.00 | Affected | 0.1445 | 0.4276 | -3 | -2 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||
| c.1142G>T | G381V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G381V is listed in ClinVar with an uncertain significance (ClinVar ID 1940172.0) and is present in the gnomAD database (6‑33438047‑G‑T). Functional prediction tools that report a benign effect include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, FoldX, Rosetta, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a majority‑benign vote and is reported as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of predictions lean toward a benign impact, and this is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.724957 | Disordered | 0.431692 | Uncertain | 0.301 | 0.951 | 0.750 | Uncertain | 1 | 6-33438047-G-T | 2 | 1.25e-6 | -5.967 | Likely Benign | 0.146 | Likely Benign | Likely Benign | 7.16 | Destabilizing | 1.0 | 4.10 | Destabilizing | 5.63 | Destabilizing | -0.32 | Likely Benign | 0.618 | Likely Pathogenic | -0.95 | Neutral | 0.386 | Benign | 0.157 | Benign | 1.32 | Pathogenic | 0.10 | Tolerated | 4.32 | 9 | 0.1621 | 0.3902 | -1 | -3 | 4.6 | 42.08 | 214.6 | -68.8 | 0.3 | 0.7 | -0.5 | 0.3 | Uncertain | Gly381 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364, res. Ala399-Ile411). Because the Ω loop is assumed to directly interact with the membrane, it moves arbitrarily throughout the WT solvent simulations. The Ω loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play major roles in protein functions that require flexibility, and thus hydrophobic residues like valine are rarely tolerated. Although no negative structural effects are observed in the variant simulations, Val381 may exert drastic effects on the SynGAP-membrane complex dynamics and stability. However, since the effects on Gly-rich Ω loop dynamics can only be well studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||
| c.3919C>A | P1307T 2D AIThe SynGAP1 missense variant P1307T is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset predicts pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the consensus of all available predictions strongly suggests that P1307T is most likely benign, and this conclusion is consistent with the lack of ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.798249 | Disordered | 0.913511 | Binding | 0.491 | 0.901 | 0.875 | -4.262 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.072 | Likely Benign | 0.20 | Neutral | 0.386 | Benign | 0.266 | Benign | 3.07 | Benign | 0.60 | Tolerated | 0.1732 | 0.5436 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.118G>T | D40Y 2D  AIThe SynGAP1 D40Y missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus indicates Likely Benign, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for D40Y, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.384043 | Structured | 0.432002 | Uncertain | 0.319 | 0.769 | 0.375 | -4.313 | Likely Benign | 0.483 | Ambiguous | Likely Benign | 0.182 | Likely Benign | -1.72 | Neutral | 0.388 | Benign | 0.328 | Benign | 3.98 | Benign | 0.00 | Affected | 0.1173 | 0.7918 | -4 | -3 | 2.2 | 48.09 | |||||||||||||||||||||||||||||||||||||||
| c.1672C>A | H558N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H558N missense variant is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus (Likely Pathogenic), PROVEAN, ESM1b, and FATHMM. Uncertain results come from premPS and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.033407 | Structured | 0.011039 | Uncertain | 0.897 | 0.200 | 0.000 | -9.523 | Likely Pathogenic | 0.257 | Likely Benign | Likely Benign | -0.22 | Likely Benign | 0.1 | 0.82 | Ambiguous | 0.30 | Likely Benign | 0.98 | Ambiguous | 0.433 | Likely Benign | -4.58 | Deleterious | 0.388 | Benign | 0.327 | Benign | -1.25 | Pathogenic | 0.14 | Tolerated | 0.1466 | 0.1281 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||
| c.184G>T | D62Y 2D AIThe SynGAP1 missense variant D62Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.425610 | Structured | 0.476010 | Uncertain | 0.575 | 0.720 | 0.125 | -6.313 | Likely Benign | 0.569 | Likely Pathogenic | Likely Benign | 0.109 | Likely Benign | -2.17 | Neutral | 0.388 | Benign | 0.328 | Benign | 4.03 | Benign | 0.00 | Affected | 0.0657 | 0.5588 | -4 | -3 | 2.2 | 48.09 | |||||||||||||||||||||||||||||||||||||||
| c.3962C>G | P1321R 2D AIThe SynGAP1 missense variant P1321R is reported in gnomAD (ID 6‑33451836‑C‑G) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool examined—REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classifies the substitution as benign. No pathogenic predictions are present. Grouping the results, the benign consensus is unanimous; the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign status. Foldetta stability analysis is not available for this variant. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.933505 | Binding | 0.463 | 0.828 | 0.875 | 6-33451836-C-G | 1 | 6.58e-7 | -5.310 | Likely Benign | 0.154 | Likely Benign | Likely Benign | 0.034 | Likely Benign | -1.10 | Neutral | 0.389 | Benign | 0.024 | Benign | 4.25 | Benign | 0.09 | Tolerated | 3.77 | 5 | 0.1722 | 0.3635 | -2 | 0 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||||||
| c.53A>C | Y18S 2D  AIThe SynGAP1 missense variant Y18S is reported in gnomAD (ID 6‑33420317‑A‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign impact for Y18S. This conclusion is consistent with the absence of a pathogenic ClinVar classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.446314 | Uncertain | 0.345 | 0.908 | 0.375 | 6-33420317-A-C | -1.061 | Likely Benign | 0.280 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -0.50 | Neutral | 0.389 | Benign | 0.036 | Benign | 4.09 | Benign | 0.00 | Affected | 4.32 | 1 | 0.4961 | 0.2640 | -2 | -3 | 0.5 | -76.10 | ||||||||||||||||||||||||||||||||||||
| c.2185A>G | N729D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N729D is predicted to be benign by all available in‑silico tools. Consensus predictors (REVEL, SIFT, polyPhen‑2 HumDiv/HumVar, PROVEAN, premPS, FoldX, Rosetta, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly report a benign effect, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign impact on protein stability. ClinVar contains no entry for this variant, and it is not listed in gnomAD. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.750527 | Disordered | 0.426547 | Uncertain | 0.651 | 0.583 | 0.625 | -5.117 | Likely Benign | 0.270 | Likely Benign | Likely Benign | 0.03 | Likely Benign | 0.2 | 0.10 | Likely Benign | 0.07 | Likely Benign | 0.14 | Likely Benign | 0.054 | Likely Benign | -1.22 | Neutral | 0.390 | Benign | 0.144 | Benign | 3.41 | Benign | 0.55 | Tolerated | 0.1931 | 0.2250 | 2 | 1 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||
| c.349A>G | S117G 2D AIThe SynGAP1 missense variant S117G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. The variant is most likely benign, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.672422 | Binding | 0.357 | 0.877 | 0.625 | -3.280 | Likely Benign | 0.174 | Likely Benign | Likely Benign | 0.122 | Likely Benign | -1.59 | Neutral | 0.390 | Benign | 0.066 | Benign | 3.73 | Benign | 0.01 | Affected | 0.2113 | 0.4627 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.538T>G | S180A 2D AIThe SynGAP1 missense variant S180A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority (2 benign vs. 1 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the S180A variant is most likely benign, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.509769 | Disordered | 0.442877 | Uncertain | 0.320 | 0.616 | 0.500 | -10.413 | Likely Pathogenic | 0.559 | Ambiguous | Likely Benign | 0.095 | Likely Benign | -1.94 | Neutral | 0.390 | Benign | 0.079 | Benign | 3.90 | Benign | 0.04 | Affected | 0.5003 | 0.3788 | Weaken | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.676T>A | S226T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S226T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while FoldX is uncertain and therefore treated as unavailable. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts benign. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.129801 | Structured | 0.334959 | Uncertain | 0.800 | 0.324 | 0.250 | -5.066 | Likely Benign | 0.102 | Likely Benign | Likely Benign | 0.53 | Ambiguous | 0.2 | 0.12 | Likely Benign | 0.33 | Likely Benign | -0.10 | Likely Benign | 0.326 | Likely Benign | -1.47 | Neutral | 0.390 | Benign | 0.079 | Benign | 5.75 | Benign | 0.05 | Affected | 0.1991 | 0.6681 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||
| c.691T>C | F231L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F231L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; the SGM‑Consensus score is “Likely Pathogenic.” Predictions that are uncertain or inconclusive are FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points to a pathogenic impact for F231L, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.366687 | Structured | 0.306467 | Uncertain | 0.895 | 0.300 | 0.000 | -9.482 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 1.07 | Ambiguous | 0.3 | 1.26 | Ambiguous | 1.17 | Ambiguous | 0.85 | Ambiguous | 0.801 | Likely Pathogenic | -5.08 | Deleterious | 0.390 | Benign | 0.142 | Benign | 6.04 | Benign | 0.01 | Affected | 0.2338 | 0.3794 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||
| c.693T>A | F231L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F231L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; the SGM‑Consensus score is “Likely Pathogenic.” Predictions that are uncertain or inconclusive are FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points to a pathogenic impact for F231L, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.366687 | Structured | 0.306467 | Uncertain | 0.895 | 0.300 | 0.000 | -9.482 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 1.07 | Ambiguous | 0.3 | 1.26 | Ambiguous | 1.17 | Ambiguous | 0.85 | Ambiguous | 0.662 | Likely Pathogenic | -5.08 | Deleterious | 0.390 | Benign | 0.142 | Benign | 6.04 | Benign | 0.01 | Affected | 0.2338 | 0.3794 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||
| c.693T>G | F231L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F231L is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify it as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar and FATHMM, whereas REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default and AlphaMissense‑Optimized all predict a pathogenic effect. The SGM‑Consensus score is labeled Likely Pathogenic, and the remaining tools (FoldX, Rosetta, Foldetta, premPS) are inconclusive. High‑accuracy methods give a pathogenic verdict from AlphaMissense‑Optimized, a Likely Pathogenic result from the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and an uncertain outcome from Foldetta. Overall, the majority of evidence points to a pathogenic impact for F231L, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.366687 | Structured | 0.306467 | Uncertain | 0.895 | 0.300 | 0.000 | -9.482 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 1.07 | Ambiguous | 0.3 | 1.26 | Ambiguous | 1.17 | Ambiguous | 0.85 | Ambiguous | 0.661 | Likely Pathogenic | -5.08 | Deleterious | 0.390 | Benign | 0.142 | Benign | 6.04 | Benign | 0.01 | Affected | 0.2338 | 0.3794 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||
| c.3926T>G | V1309G 2D AIThe SynGAP1 missense variant V1309G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact, and this conclusion does not contradict any ClinVar annotation because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.712013 | Disordered | 0.948596 | Binding | 0.402 | 0.907 | 0.750 | -2.618 | Likely Benign | 0.160 | Likely Benign | Likely Benign | 0.326 | Likely Benign | -3.40 | Deleterious | 0.391 | Benign | 0.767 | Possibly Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.1901 | 0.2144 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.436T>C | S146P 2D AIThe SynGAP1 missense variant S146P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions arise from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates likely pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for S146P, and this conclusion is not contradicted by ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.541878 | Disordered | 0.508612 | Binding | 0.349 | 0.837 | 0.625 | -13.292 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.215 | Likely Benign | -3.11 | Deleterious | 0.392 | Benign | 0.230 | Benign | 3.60 | Benign | 0.00 | Affected | 0.1934 | 0.4627 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.2303A>G | D768G 2D AIThe SynGAP1 D768G missense variant is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors shows a predominance of benign calls: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized all predict benign, whereas AlphaMissense‑Default predicts pathogenic. The high‑accuracy AlphaMissense‑Optimized result is benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign classification (2 benign vs. 1 pathogenic, with one uncertain). Foldetta, which would provide a protein‑folding stability assessment, has no available output for this variant. Overall, the preponderance of evidence points to a benign effect, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.332115 | Structured | 0.928237 | Binding | 0.314 | 0.877 | 0.250 | -7.150 | In-Between | 0.697 | Likely Pathogenic | Likely Benign | 0.184 | Likely Benign | 0.03 | Neutral | 0.393 | Benign | 0.131 | Benign | 4.09 | Benign | 0.72 | Tolerated | 0.4130 | 0.7476 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2304C>A | D768E 2D AIThe SynGAP1 missense variant D768E is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all classify the change as benign, and AlphaMissense‑Optimized also predicts a benign outcome. No tool predicts pathogenicity; AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the computational evidence strongly supports a benign effect for D768E, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.332115 | Structured | 0.928237 | Binding | 0.314 | 0.877 | 0.250 | -4.611 | Likely Benign | 0.380 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -1.23 | Neutral | 0.393 | Benign | 0.171 | Benign | 4.16 | Benign | 0.17 | Tolerated | 0.1368 | 0.7729 | 3 | 2 | 0.0 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2304C>G | D768E 2D AIThe SynGAP1 missense variant D768E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the computational evidence strongly suggests that D768E is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.332115 | Structured | 0.928237 | Binding | 0.314 | 0.877 | 0.250 | -4.611 | Likely Benign | 0.380 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -1.23 | Neutral | 0.393 | Benign | 0.171 | Benign | 4.16 | Benign | 0.17 | Tolerated | 0.1368 | 0.7729 | 3 | 2 | 0.0 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2335A>G | S779G 2D AIThe SynGAP1 missense variant S779G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the evidence strongly supports a benign classification, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.509769 | Disordered | 0.834974 | Binding | 0.321 | 0.890 | 0.375 | -4.304 | Likely Benign | 0.111 | Likely Benign | Likely Benign | 0.103 | Likely Benign | 0.38 | Neutral | 0.393 | Benign | 0.324 | Benign | 2.65 | Benign | 0.53 | Tolerated | 0.2894 | 0.5087 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2546A>G | D849G 2D AISynGAP1 missense variant D849G is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) indicates likely benign. Foldetta results are unavailable. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.712013 | Disordered | 0.554191 | Binding | 0.319 | 0.813 | 0.500 | -2.124 | Likely Benign | 0.168 | Likely Benign | Likely Benign | 0.154 | Likely Benign | -0.55 | Neutral | 0.393 | Benign | 0.140 | Benign | 4.17 | Benign | 0.00 | Affected | 0.4622 | 0.7587 | 1 | -1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.2547T>A | D849E 2D AIThe SynGAP1 missense variant D849E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.712013 | Disordered | 0.554191 | Binding | 0.319 | 0.813 | 0.500 | -2.911 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.087 | Likely Benign | -0.55 | Neutral | 0.393 | Benign | 0.187 | Benign | 4.25 | Benign | 0.00 | Affected | 0.1955 | 0.7819 | 3 | 2 | 0.0 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2547T>G | D849E 2D AIThe SynGAP1 missense variant D849E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.712013 | Disordered | 0.554191 | Binding | 0.319 | 0.813 | 0.500 | -2.911 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.087 | Likely Benign | -0.55 | Neutral | 0.393 | Benign | 0.187 | Benign | 4.25 | Benign | 0.00 | Affected | 0.1955 | 0.7819 | 3 | 2 | 0.0 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2554G>A | G852S 2D AIThe SynGAP1 missense variant G852S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.506063 | Binding | 0.276 | 0.816 | 0.625 | -4.786 | Likely Benign | 0.071 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -0.17 | Neutral | 0.393 | Benign | 0.197 | Benign | 4.21 | Benign | 0.07 | Tolerated | 0.2696 | 0.5129 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2555G>C | G852A 2D AIThe SynGAP1 missense variant G852A is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Based on the collective predictions, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.506063 | Binding | 0.276 | 0.816 | 0.625 | -4.493 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.142 | Likely Benign | -0.35 | Neutral | 0.393 | Benign | 0.321 | Benign | 4.25 | Benign | 1.00 | Tolerated | 0.3895 | 0.4934 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2579T>C | V860A 2D AIThe SynGAP1 missense variant V860A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while only SIFT predicts pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence supports a benign impact for V860A, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.545602 | Disordered | 0.518121 | Binding | 0.269 | 0.803 | 0.250 | -3.379 | Likely Benign | 0.161 | Likely Benign | Likely Benign | 0.223 | Likely Benign | -0.91 | Neutral | 0.393 | Benign | 0.185 | Benign | 4.20 | Benign | 0.00 | Affected | 0.3215 | 0.2723 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.2638G>T | A880S 2D AIThe SynGAP1 missense variant A880S is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.703578 | Disordered | 0.621441 | Binding | 0.309 | 0.874 | 0.250 | -3.149 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.092 | Likely Benign | -0.33 | Neutral | 0.393 | Benign | 0.187 | Benign | 2.63 | Benign | 0.10 | Tolerated | 0.2504 | 0.5108 | 1 | 1 | -2.6 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2906G>C | G969A 2D AIThe SynGAP1 missense variant G969A is not reported in ClinVar and is absent from gnomAD, indicating no documented clinical or population evidence. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity, so the pathogenic‑prediction group is empty. High‑accuracy assessments corroborate this benign consensus: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are not available, so they do not influence the interpretation. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.950334 | Disordered | 0.956572 | Binding | 0.405 | 0.898 | 0.750 | -4.693 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.120 | Likely Benign | -0.36 | Neutral | 0.393 | Benign | 0.096 | Benign | 4.25 | Benign | 0.51 | Tolerated | 0.3414 | 0.5096 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3242C>G | A1081G 2D AIThe missense variant A1081G in SynGAP1 has no entry in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as likely benign. Foldetta results are unavailable. Overall, the computational evidence strongly supports a benign classification, with no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.874069 | Disordered | 0.979759 | Binding | 0.288 | 0.895 | 0.750 | -3.174 | Likely Benign | 0.191 | Likely Benign | Likely Benign | 0.033 | Likely Benign | -1.43 | Neutral | 0.393 | Benign | 0.184 | Benign | 3.99 | Benign | 0.23 | Tolerated | 0.1751 | 0.4348 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3323G>C | S1108T 2D AIThe SynGAP1 missense variant S1108T is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Overall, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.949221 | Binding | 0.324 | 0.886 | 0.875 | -5.710 | Likely Benign | 0.105 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -1.49 | Neutral | 0.393 | Benign | 0.239 | Benign | 2.56 | Benign | 0.25 | Tolerated | 0.1304 | 0.5365 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3335A>C | E1112A 2D AIThe SynGAP1 missense variant E1112A is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.909381 | Binding | 0.335 | 0.902 | 0.875 | -3.227 | Likely Benign | 0.373 | Ambiguous | Likely Benign | 0.096 | Likely Benign | -1.86 | Neutral | 0.393 | Benign | 0.131 | Benign | 2.71 | Benign | 0.02 | Affected | 0.3929 | 0.7528 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.3356G>C | G1119A 2D AIThe SynGAP1 missense variant G1119A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.818538 | Binding | 0.339 | 0.928 | 0.875 | -6.703 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.201 | Likely Benign | -0.33 | Neutral | 0.393 | Benign | 0.187 | Benign | 3.93 | Benign | 1.00 | Tolerated | 0.3591 | 0.4957 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3385C>G | L1129V 2D AIThe SynGAP1 missense variant L1129V is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.876543 | Binding | 0.339 | 0.909 | 0.875 | -3.595 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.262 | Likely Benign | -0.87 | Neutral | 0.393 | Benign | 0.187 | Benign | 5.46 | Benign | 0.00 | Affected | 0.1814 | 0.3804 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.1105A>C | T369P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T369P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome, while FoldX and Foldetta are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as Likely Benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact for T369P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | -2.743 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 1.20 | Ambiguous | 2.1 | 0.18 | Likely Benign | 0.69 | Ambiguous | 0.17 | Likely Benign | 0.138 | Likely Benign | -2.09 | Neutral | 0.396 | Benign | 0.142 | Benign | 1.83 | Pathogenic | 0.16 | Tolerated | 0.2589 | 0.6046 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1106C>T | T369I 2D AIThe SynGAP1 missense variant T369I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. Stability‑based methods (FoldX, Rosetta, Foldetta) are inconclusive, providing no definitive evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as likely benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact; there is no conflict with ClinVar status, which contains no entry for this variant. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | -6.759 | Likely Benign | 0.289 | Likely Benign | Likely Benign | 0.60 | Ambiguous | 0.8 | 1.41 | Ambiguous | 1.01 | Ambiguous | -0.08 | Likely Benign | 0.078 | Likely Benign | -2.37 | Neutral | 0.396 | Benign | 0.142 | Benign | 1.72 | Pathogenic | 0.13 | Tolerated | 0.1106 | 0.7207 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.1111A>C | S371R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense change S371R is catalogued in gnomAD (ID 6‑33438016‑A‑C) but has no ClinVar entry. Functional prediction programs largely agree on a benign effect: REVEL, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, Foldetta, and the SGM‑Consensus score (Likely Benign) all report a non‑pathogenic outcome. Only AlphaMissense‑Default predicts a pathogenic effect, while FoldX and premPS are inconclusive. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta) is benign. Taken together, the majority of evidence supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic report. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.444081 | Structured | 0.432086 | Uncertain | 0.294 | 0.746 | 0.375 | 6-33438016-A-C | -6.415 | Likely Benign | 0.762 | Likely Pathogenic | Likely Benign | 0.51 | Ambiguous | 1.2 | -0.25 | Likely Benign | 0.13 | Likely Benign | 0.57 | Ambiguous | 0.295 | Likely Benign | -1.17 | Neutral | 0.396 | Benign | 0.099 | Benign | 5.35 | Benign | 0.26 | Tolerated | 3.52 | 18 | 0.1362 | 0.4131 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||
| c.1113T>A | S371R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S371R is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools largely agree on a benign effect: SGM‑Consensus, REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all classify the substitution as benign, while AlphaMissense‑Optimized also predicts benign. Only AlphaMissense‑Default indicates a pathogenic outcome; FoldX and premPS are inconclusive. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. Overall, the majority of evidence supports a benign impact, and this is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.444081 | Structured | 0.432086 | Uncertain | 0.294 | 0.746 | 0.375 | -6.415 | Likely Benign | 0.762 | Likely Pathogenic | Likely Benign | 0.51 | Ambiguous | 1.2 | -0.25 | Likely Benign | 0.13 | Likely Benign | 0.57 | Ambiguous | 0.344 | Likely Benign | -1.17 | Neutral | 0.396 | Benign | 0.099 | Benign | 5.35 | Benign | 0.26 | Tolerated | 3.52 | 18 | 0.1362 | 0.4131 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||
| c.1113T>G | S371R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S371R is reported in gnomAD (variant ID 6‑33438018‑T‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves as “Likely Benign” (3 benign vs. 1 pathogenic). High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM‑Consensus is benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. No prediction or stability result is missing or inconclusive. Overall, the evidence strongly favors a benign classification, and this is consistent with the absence of a ClinVar pathogenic report. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.444081 | Structured | 0.432086 | Uncertain | 0.294 | 0.746 | 0.375 | 6-33438018-T-G | 1 | 1.18e-6 | -6.415 | Likely Benign | 0.762 | Likely Pathogenic | Likely Benign | 0.51 | Ambiguous | 1.2 | -0.25 | Likely Benign | 0.13 | Likely Benign | 0.57 | Ambiguous | 0.340 | Likely Benign | -1.17 | Neutral | 0.396 | Benign | 0.099 | Benign | 5.35 | Benign | 0.26 | Tolerated | 3.52 | 18 | 0.1362 | 0.4131 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||
| c.1120T>C | S374P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S374P is reported in gnomAD (6‑33438025‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and Rosetta; FoldX and Foldetta are inconclusive. The high‑accuracy consensus (SGM‑Consensus) is “Likely Benign,” derived from the unanimous benign calls of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN. AlphaMissense‑Optimized also predicts benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign impact. There is no ClinVar status to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.642678 | Disordered | 0.428948 | Uncertain | 0.333 | 0.812 | 0.625 | 6-33438025-T-C | 1 | 7.85e-7 | -4.849 | Likely Benign | 0.125 | Likely Benign | Likely Benign | 0.66 | Ambiguous | 0.6 | 2.22 | Destabilizing | 1.44 | Ambiguous | 0.34 | Likely Benign | 0.388 | Likely Benign | -0.89 | Neutral | 0.396 | Benign | 0.099 | Benign | 5.30 | Benign | 0.02 | Affected | 4.32 | 13 | 0.3012 | 0.6813 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||
| c.353T>A | M118K 2D AIThe SynGAP1 missense variant M118K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation, as none exists for M118K. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.676867 | Binding | 0.330 | 0.883 | 0.500 | -3.979 | Likely Benign | 0.767 | Likely Pathogenic | Likely Benign | 0.276 | Likely Benign | -2.98 | Deleterious | 0.396 | Benign | 0.099 | Benign | 3.84 | Benign | 0.01 | Affected | 0.2010 | 0.1131 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||||||||||||
| c.353T>C | M118T 2D AIThe SynGAP1 missense variant M118T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The high‑accuracy consensus (SGM‑Consensus) also indicates a likely benign classification, and AlphaMissense‑Optimized independently predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the evidence from multiple independent predictors and the high‑accuracy consensus strongly supports a benign impact. This conclusion is consistent with the lack of any ClinVar pathogenic annotation, so there is no contradiction with existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.694846 | Disordered | 0.676867 | Binding | 0.330 | 0.883 | 0.500 | -2.468 | Likely Benign | 0.454 | Ambiguous | Likely Benign | 0.217 | Likely Benign | -2.41 | Neutral | 0.396 | Benign | 0.067 | Benign | 3.86 | Benign | 0.08 | Tolerated | 0.2207 | 0.2237 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.511G>C | A171P 2D AIThe SynGAP1 missense variant A171P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates likely benign. Foldetta results are unavailable for this variant. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.492272 | Uncertain | 0.358 | 0.652 | 0.375 | -5.071 | Likely Benign | 0.569 | Likely Pathogenic | Likely Benign | 0.160 | Likely Benign | -1.42 | Neutral | 0.396 | Benign | 0.099 | Benign | 4.14 | Benign | 0.02 | Affected | 0.1630 | 0.3938 | 1 | -1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||||||
| c.524A>C | Q175P 2D AIThe SynGAP1 missense variant Q175P is reported in gnomAD (ID 6‑33435166‑A‑C) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the substitution as benign. The consensus from the SGM framework, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely benign. High‑accuracy predictors corroborate this assessment: AlphaMissense‑Optimized predicts a benign outcome, and the SGM‑Consensus (majority vote) confirms a likely benign status. No high‑accuracy folding‑stability analysis (Foldetta) is available for this residue, so stability effects remain unassessed. Overall, the collective evidence points to a benign impact for Q175P, and this conclusion is consistent with the absence of a pathogenic ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.653063 | Disordered | 0.474689 | Uncertain | 0.367 | 0.618 | 0.375 | 6-33435166-A-C | 1 | 6.20e-7 | -6.995 | Likely Benign | 0.432 | Ambiguous | Likely Benign | 0.211 | Likely Benign | 1.55 | Neutral | 0.396 | Benign | 0.188 | Benign | 4.20 | Benign | 1.00 | Tolerated | 3.61 | 5 | 0.2075 | 0.4584 | -1 | 0 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||
| c.589G>C | E197Q 2D AIThe SynGAP1 E197Q variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The high‑accuracy consensus, SGM‑Consensus, is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN and therefore reports a likely benign outcome. AlphaMissense‑Optimized also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.349426 | Structured | 0.431896 | Uncertain | 0.452 | 0.492 | 0.125 | -6.347 | Likely Benign | 0.723 | Likely Pathogenic | Likely Benign | 0.145 | Likely Benign | -1.75 | Neutral | 0.396 | Benign | 0.067 | Benign | 4.05 | Benign | 0.02 | Affected | 0.0867 | 0.5228 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.590A>T | E197V 2D AIThe SynGAP1 missense variant E197V is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split opinion: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. When predictions are grouped by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Taken together, the preponderance of evidence from both general and high‑accuracy predictors indicates that E197V is most likely pathogenic, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.349426 | Structured | 0.431896 | Uncertain | 0.452 | 0.492 | 0.125 | -9.023 | Likely Pathogenic | 0.932 | Likely Pathogenic | Ambiguous | 0.247 | Likely Benign | -4.51 | Deleterious | 0.396 | Benign | 0.099 | Benign | 4.01 | Benign | 0.00 | Affected | 0.0490 | 0.6024 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.673T>C | S225P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S225P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign calls come from premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic calls arise from REVEL, FoldX, PROVEAN, and ESM1b. Two tools—Rosetta and Foldetta—return uncertain results and are treated as unavailable. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts benign, whereas the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta remains unavailable. Overall, the majority of evidence leans toward a benign effect, and this conclusion does not conflict with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.150080 | Structured | 0.344191 | Uncertain | 0.817 | 0.319 | 0.250 | -8.084 | Likely Pathogenic | 0.156 | Likely Benign | Likely Benign | 2.22 | Destabilizing | 1.8 | 1.14 | Ambiguous | 1.68 | Ambiguous | 0.15 | Likely Benign | 0.503 | Likely Pathogenic | -3.31 | Deleterious | 0.396 | Benign | 0.133 | Benign | 5.91 | Benign | 0.17 | Tolerated | 0.2128 | 0.6946 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||
| c.2020A>G | T674A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T674A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly favor a benign effect: AlphaMissense‑Default, AlphaMissense‑Optimized, ESM1b, FATHMM, PROVEAN, SIFT, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, REVEL, FoldX, premPS, Foldetta, and the SGM‑Consensus score all indicate benign or likely benign. No tool predicts pathogenicity; the only inconclusive result comes from Rosetta, which is listed as uncertain. High‑accuracy methods corroborate the benign assessment: AlphaMissense‑Optimized is benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. Based on the aggregate predictions, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.129801 | Structured | 0.109297 | Uncertain | 0.521 | 0.349 | 0.000 | -5.915 | Likely Benign | 0.099 | Likely Benign | Likely Benign | -0.13 | Likely Benign | 0.0 | 0.89 | Ambiguous | 0.38 | Likely Benign | 0.29 | Likely Benign | 0.052 | Likely Benign | -1.48 | Neutral | 0.398 | Benign | 0.045 | Benign | 3.45 | Benign | 0.28 | Tolerated | 0.4042 | 0.4845 | 1 | 0 | 2.5 | -30.03 | |||||||||||||||||||||||||||||
| c.425A>G | K142R 2D AIThe SynGAP1 missense variant K142R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors benign (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -9.327 | Likely Pathogenic | 0.361 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -1.87 | Neutral | 0.399 | Benign | 0.212 | Benign | 3.65 | Benign | 0.00 | Affected | 0.4459 | 0.0858 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||||
| c.434A>G | K145R 2D AIThe SynGAP1 missense variant K145R is listed in gnomAD (ID 6‑33432731‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign or likely benign. Only SIFT predicts a pathogenic outcome, while ESM1b remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status, as none is currently assigned to the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | 6-33432731-A-G | 1 | 6.20e-7 | -7.685 | In-Between | 0.214 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -1.58 | Neutral | 0.399 | Benign | 0.212 | Benign | 3.71 | Benign | 0.00 | Affected | 3.61 | 5 | 0.4661 | 0.1242 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||
| c.496G>A | A166T 2D AIThe SynGAP1 missense variant A166T is not reported in ClinVar and is absent from gnomAD. In silico prediction tools that assess sequence conservation and structural impact (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) all classify the change as benign. No tool in the dataset predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports “Likely Benign.” Foldetta results are not available. Overall, the consensus of all available predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.425610 | Structured | 0.505037 | Binding | 0.384 | 0.658 | 0.125 | -6.700 | Likely Benign | 0.169 | Likely Benign | Likely Benign | 0.115 | Likely Benign | -0.78 | Neutral | 0.399 | Benign | 0.273 | Benign | 4.10 | Benign | 0.31 | Tolerated | 0.1420 | 0.5134 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.496G>T | A166S 2D AIThe SynGAP1 missense variant A166 S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.425610 | Structured | 0.505037 | Binding | 0.384 | 0.658 | 0.125 | -6.008 | Likely Benign | 0.120 | Likely Benign | Likely Benign | 0.080 | Likely Benign | -0.78 | Neutral | 0.399 | Benign | 0.212 | Benign | 4.07 | Benign | 0.05 | Affected | 0.2286 | 0.4146 | 1 | 1 | -2.6 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.497C>G | A166G 2D AIThe SynGAP1 missense variant A166G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT and ESM1b predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a majority‑benign vote and is reported as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.425610 | Structured | 0.505037 | Binding | 0.384 | 0.658 | 0.125 | -8.188 | Likely Pathogenic | 0.215 | Likely Benign | Likely Benign | 0.101 | Likely Benign | -1.16 | Neutral | 0.399 | Benign | 0.212 | Benign | 4.02 | Benign | 0.03 | Affected | 0.1665 | 0.3201 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2204G>A | S735N 2D AIThe SynGAP1 missense variant S735N is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. No pathogenic predictions are present among the evaluated tools. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.916840 | Disordered | 0.412174 | Uncertain | 0.290 | 0.752 | 0.875 | -6.697 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -0.68 | Neutral | 0.400 | Benign | 0.138 | Benign | 2.65 | Benign | 0.18 | Tolerated | 0.1375 | 0.3827 | 1 | 1 | -2.7 | 27.03 | |||||||||||||||||||||||||||||||||||||||
| c.2410G>A | D804N 2D  AIThe SynGAP1 D804N variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of conventional tools predict a benign impact, and this assessment does not contradict the lack of ClinVar annotation. Thus, based on the current predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | SH3-binding motif | 0.801317 | Disordered | 0.786762 | Binding | 0.294 | 0.900 | 0.625 | -4.681 | Likely Benign | 0.522 | Ambiguous | Likely Benign | 0.104 | Likely Benign | -2.62 | Deleterious | 0.400 | Benign | 0.212 | Benign | 1.22 | Pathogenic | 0.06 | Tolerated | 0.1528 | 0.7393 | 2 | 1 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.2412C>A | D804E 2D AIThe SynGAP1 D804E missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign” (3 benign vs. 1 pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote) as benign, and Foldetta results are unavailable. Based on the overall consensus of the available predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.801317 | Disordered | 0.786762 | Binding | 0.294 | 0.900 | 0.625 | -4.155 | Likely Benign | 0.243 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -2.03 | Neutral | 0.400 | Benign | 0.138 | Benign | 1.26 | Pathogenic | 0.04 | Affected | 0.1765 | 0.6850 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.2412C>G | D804E 2D AIThe SynGAP1 D804E missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign” (3 benign vs. 1 pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote) as benign, and Foldetta results are unavailable. Based on the overall consensus of the available predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.801317 | Disordered | 0.786762 | Binding | 0.294 | 0.900 | 0.625 | -4.155 | Likely Benign | 0.243 | Likely Benign | Likely Benign | 0.038 | Likely Benign | -2.03 | Neutral | 0.400 | Benign | 0.138 | Benign | 1.26 | Pathogenic | 0.04 | Affected | 0.1765 | 0.6850 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.14G>A | R5Q 2D AIThe SynGAP1 missense variant R5Q is reported in gnomAD (variant ID 6‑33420278‑G‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools, including the high‑accuracy methods, points to a benign impact. This prediction does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.595080 | Disordered | 0.547847 | Binding | 0.363 | 0.920 | 0.750 | 6-33420278-G-A | 2 | 1.30e-6 | -4.261 | Likely Benign | 0.223 | Likely Benign | Likely Benign | 0.094 | Likely Benign | -0.06 | Neutral | 0.403 | Benign | 0.007 | Benign | 4.15 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3740 | 0.3122 | 1 | 1 | 1.0 | -28.06 | ||||||||||||||||||||||||||||||||||
| c.44C>A | A15E 2D AIThe SynGAP1 missense variant A15E is reported in gnomAD (ID 6‑33420308‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | 6-33420308-C-A | -3.423 | Likely Benign | 0.277 | Likely Benign | Likely Benign | 0.169 | Likely Benign | 0.46 | Neutral | 0.406 | Benign | 0.040 | Benign | 4.13 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1563 | 0.1908 | -1 | 0 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||
| c.2530C>G | L844V 2D AIThe SynGAP1 missense variant L844V is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.595080 | Disordered | 0.611301 | Binding | 0.304 | 0.835 | 0.375 | 0.094 | Likely Benign | 0.240 | Likely Benign | Likely Benign | 0.043 | Likely Benign | 0.20 | Neutral | 0.409 | Benign | 0.253 | Benign | 2.87 | Benign | 0.62 | Tolerated | 0.1692 | 0.3744 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2581T>G | S861A 2D AIThe SynGAP1 missense variant S861A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.557691 | Disordered | 0.540903 | Binding | 0.285 | 0.797 | 0.250 | -5.059 | Likely Benign | 0.092 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -1.02 | Neutral | 0.409 | Benign | 0.172 | Benign | 4.08 | Benign | 0.48 | Tolerated | 0.4541 | 0.5401 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2818G>A | G940S 2D AIThe SynGAP1 missense variant G940S is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443370‑G‑A). All available in silico predictors agree on a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” High‑accuracy tools reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no reported result for this variant, so its status is unavailable. Overall, the computational evidence strongly supports a benign classification, which does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.889439 | Disordered | 0.920635 | Binding | 0.383 | 0.902 | 0.625 | Uncertain | 1 | 6-33443370-G-A | 1 | 6.20e-7 | -5.451 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.135 | Likely Benign | 0.45 | Neutral | 0.409 | Benign | 0.253 | Benign | 2.77 | Benign | 0.44 | Tolerated | 3.77 | 5 | 0.2590 | 0.4907 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||
| c.3310C>G | P1104A 2D AIThe SynGAP1 missense variant P1104A is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.936162 | Disordered | 0.954801 | Binding | 0.440 | 0.863 | 0.875 | -3.677 | Likely Benign | 0.059 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -0.54 | Neutral | 0.409 | Benign | 0.184 | Benign | 2.74 | Benign | 0.22 | Tolerated | 0.3289 | 0.5315 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.1099C>G | L367V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L367V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only FATHMM predicts a pathogenic outcome. Stability‑based methods (FoldX, Rosetta, Foldetta) are inconclusive, so they provide no evidence for or against pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as likely benign, and Foldetta as uncertain. Overall, the preponderance of evidence points to a benign effect for L367V, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.370445 | Structured | 0.441805 | Uncertain | 0.790 | 0.657 | 0.250 | -2.383 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 1.48 | Ambiguous | 0.2 | 1.72 | Ambiguous | 1.60 | Ambiguous | 0.15 | Likely Benign | 0.040 | Likely Benign | 0.19 | Neutral | 0.410 | Benign | 0.104 | Benign | 1.67 | Pathogenic | 0.13 | Tolerated | 0.2321 | 0.3285 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||
| c.358G>T | G120C 2D AIThe SynGAP1 missense variant G120C is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.659993 | Binding | 0.359 | 0.887 | 0.750 | -5.979 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -0.73 | Neutral | 0.410 | Benign | 0.146 | Benign | 4.20 | Benign | 0.10 | Tolerated | 0.1376 | 0.4225 | -3 | -3 | 2.9 | 46.09 | |||||||||||||||||||||||||||||||||||||||
| c.1879G>T | A627S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A627S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are Rosetta, PROVEAN, and ESM1b. The remaining tools—FoldX, Foldetta, and premPS—return uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta is uncertain. Overall, the majority of available predictions lean toward a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.100716 | Structured | 0.037862 | Uncertain | 0.970 | 0.210 | 0.000 | -10.782 | Likely Pathogenic | 0.329 | Likely Benign | Likely Benign | 1.11 | Ambiguous | 0.2 | 2.05 | Destabilizing | 1.58 | Ambiguous | 0.71 | Ambiguous | 0.316 | Likely Benign | -2.94 | Deleterious | 0.411 | Benign | 0.387 | Benign | 2.78 | Benign | 0.11 | Tolerated | 0.2266 | 0.4224 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||
| c.2351C>A | A784D 2D AIThe SynGAP1 missense variant A784D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.801317 | Disordered | 0.708872 | Binding | 0.314 | 0.896 | 0.625 | -3.784 | Likely Benign | 0.766 | Likely Pathogenic | Likely Benign | 0.206 | Likely Benign | -1.21 | Neutral | 0.411 | Benign | 0.237 | Benign | 2.68 | Benign | 0.16 | Tolerated | 0.1824 | 0.2193 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.2405G>T | G802V 2D AIThe SynGAP1 missense variant G802V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign. No Foldetta stability analysis is available for this variant. Overall, the consensus of the available predictions indicates that G802V is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.894241 | Disordered | 0.681966 | Binding | 0.294 | 0.898 | 0.625 | -3.871 | Likely Benign | 0.126 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -1.49 | Neutral | 0.411 | Benign | 0.321 | Benign | 2.67 | Benign | 0.00 | Affected | 0.1345 | 0.3533 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||
| c.2563C>T | L855F 2D  AIThe SynGAP1 missense variant L855F is predicted to be benign by all evaluated in‑silico tools. Consensus predictions from **SGM‑Consensus** (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classify the variant as *Likely Benign*. High‑accuracy predictors **AlphaMissense‑Optimized** also report a benign effect. Other pathogenicity predictors—REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default—uniformly predict benign. No tools predict pathogenicity. **Foldetta**, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its stability impact remains unknown. The variant is not listed in ClinVar and has no entry in gnomAD, so no population frequency or clinical annotation is available. Based on the unanimous benign predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.485558 | Uncertain | 0.285 | 0.823 | 0.625 | -4.985 | Likely Benign | 0.106 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -1.86 | Neutral | 0.411 | Benign | 0.187 | Benign | 4.00 | Benign | 0.12 | Tolerated | 0.0700 | 0.3529 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.2825C>G | P942R 2D AIThe SynGAP1 missense variant P942R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.878102 | Binding | 0.365 | 0.915 | 0.625 | -5.405 | Likely Benign | 0.159 | Likely Benign | Likely Benign | 0.045 | Likely Benign | -0.78 | Neutral | 0.411 | Benign | 0.139 | Benign | 2.36 | Pathogenic | 0.00 | Affected | 0.1307 | 0.3693 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.2825C>T | P942L 2D AIThe SynGAP1 missense variant P942L is listed in ClinVar (ID 2851884.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443377‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.878102 | Binding | 0.365 | 0.915 | 0.625 | Uncertain | 1 | 6-33443377-C-T | 4 | 2.48e-6 | -5.063 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.048 | Likely Benign | -2.00 | Neutral | 0.411 | Benign | 0.239 | Benign | 2.37 | Pathogenic | 0.00 | Affected | 4.32 | 4 | 0.2094 | 0.5507 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||
| c.2827G>C | G943R 2D AIThe SynGAP1 missense variant G943R is reported in gnomAD (variant ID 6-33443379‑G‑C) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity; only ESM1b is uncertain. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions strongly supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.973328 | Disordered | 0.860437 | Binding | 0.372 | 0.910 | 0.750 | 6-33443379-G-C | 1 | 6.20e-7 | -7.159 | In-Between | 0.335 | Likely Benign | Likely Benign | 0.308 | Likely Benign | 1.00 | Neutral | 0.411 | Benign | 0.062 | Benign | 2.86 | Benign | 0.94 | Tolerated | 4.32 | 4 | 0.0976 | 0.4933 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||
| c.2839G>A | G947R 2D AIThe SynGAP1 missense variant G947R is listed in gnomAD (6-33443391-G-A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Optimized) and pathogenic (SIFT). Two tools remain uncertain (ESM1b, AlphaMissense‑Default). High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—does not reach a definitive conclusion (two benign, two uncertain). Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no reported result for this variant. Overall, the preponderance of evidence points to a benign impact, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988695 | Disordered | 0.850554 | Binding | 0.358 | 0.919 | 0.750 | 6-33443391-G-A | 6 | 3.72e-6 | -7.481 | In-Between | 0.343 | Ambiguous | Likely Benign | 0.273 | Likely Benign | -0.60 | Neutral | 0.411 | Benign | 0.140 | Benign | 4.94 | Benign | 0.04 | Affected | 4.32 | 4 | 0.1034 | 0.4733 | -2 | -3 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||
| c.2839G>C | G947R 2D AIThe SynGAP1 missense variant G947R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988695 | Disordered | 0.850554 | Binding | 0.358 | 0.919 | 0.750 | -7.481 | In-Between | 0.343 | Ambiguous | Likely Benign | 0.273 | Likely Benign | -0.60 | Neutral | 0.411 | Benign | 0.140 | Benign | 4.94 | Benign | 0.04 | Affected | 4.32 | 4 | 0.1034 | 0.4733 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||
| c.2849G>A | G950D 2D AIThe SynGAP1 missense variant G950D is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM, while ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.987317 | Disordered | 0.888649 | Binding | 0.368 | 0.923 | 0.750 | -7.515 | In-Between | 0.245 | Likely Benign | Likely Benign | 0.402 | Likely Benign | -0.61 | Neutral | 0.411 | Benign | 0.131 | Benign | 2.26 | Pathogenic | 0.02 | Affected | 0.1874 | 0.2826 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2849G>T | G950V 2D AIThe SynGAP1 missense variant G950V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM, while ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a benign prediction. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that G950V is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.987317 | Disordered | 0.888649 | Binding | 0.368 | 0.923 | 0.750 | -7.796 | In-Between | 0.093 | Likely Benign | Likely Benign | 0.428 | Likely Benign | -0.91 | Neutral | 0.411 | Benign | 0.239 | Benign | 2.26 | Pathogenic | 0.01 | Affected | 0.1392 | 0.3507 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2867C>A | S956Y 2D AIThe SynGAP1 missense variant S956Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Thus, the variant is most likely benign, with no contradiction from ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984871 | Disordered | 0.957345 | Binding | 0.364 | 0.917 | 0.750 | -4.920 | Likely Benign | 0.233 | Likely Benign | Likely Benign | 0.085 | Likely Benign | -0.90 | Neutral | 0.411 | Benign | 0.097 | Benign | 1.93 | Pathogenic | 0.22 | Tolerated | 0.1449 | 0.4932 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.2889T>A | H963Q 2D AIThe SynGAP1 missense variant H963Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; the only uncertain result comes from ESM1b. The high‑accuracy consensus methods also support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability predictor, has no available result for this variant. Overall, the evidence overwhelmingly indicates that H963Q is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.983973 | Binding | 0.325 | 0.886 | 0.750 | -7.798 | In-Between | 0.116 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -0.75 | Neutral | 0.411 | Benign | 0.132 | Benign | 4.17 | Benign | 0.29 | Tolerated | 0.2112 | 0.3979 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2889T>G | H963Q 2D AIThe SynGAP1 missense variant H963Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity; the only uncertain result comes from ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” consensus. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the collective evidence strongly supports a benign classification, and there is no ClinVar annotation to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.983973 | Binding | 0.325 | 0.886 | 0.750 | -7.798 | In-Between | 0.116 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -0.75 | Neutral | 0.411 | Benign | 0.132 | Benign | 4.17 | Benign | 0.29 | Tolerated | 0.2112 | 0.3979 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.2924C>T | T975I 2D AIThe SynGAP1 missense variant T975I is listed in ClinVar with an “Uncertain” status and is present in the gnomAD database (ID 6‑33443476‑C‑T). Prediction tools that agree on benign impact include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts pathogenicity. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, which does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.871313 | Disordered | 0.969331 | Binding | 0.332 | 0.890 | 0.625 | Uncertain | 1 | 6-33443476-C-T | 6 | 3.72e-6 | -3.912 | Likely Benign | 0.164 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -1.66 | Neutral | 0.411 | Benign | 0.239 | Benign | 4.11 | Benign | 0.66 | Tolerated | 4.32 | 2 | 0.1107 | 0.5198 | 0 | -1 | 5.2 | 12.05 | ||||||||||||||||||||||||||||||||
| c.2984C>T | P995L 2D AIThe SynGAP1 missense variant P995L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.862302 | Disordered | 0.935305 | Binding | 0.338 | 0.902 | 0.750 | -4.948 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.063 | Likely Benign | -1.03 | Neutral | 0.411 | Benign | 0.096 | Benign | 4.16 | Benign | 0.00 | Affected | 0.2294 | 0.6324 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.2990C>A | A997D 2D AIThe SynGAP1 missense variant A997D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of any ClinVar pathogenic annotation. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.948624 | Binding | 0.273 | 0.901 | 0.500 | -5.251 | Likely Benign | 0.319 | Likely Benign | Likely Benign | 0.131 | Likely Benign | -1.09 | Neutral | 0.411 | Benign | 0.120 | Benign | 4.15 | Benign | 0.00 | Affected | 0.1815 | 0.2143 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.3053C>A | T1018N 2D AIThe SynGAP1 missense variant T1018N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a benign impact. The predictions do not contradict any ClinVar status, as no ClinVar claim exists for this variant. Thus, based on current computational predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.849326 | Disordered | 0.959985 | Binding | 0.348 | 0.801 | 0.500 | -3.888 | Likely Benign | 0.165 | Likely Benign | Likely Benign | 0.045 | Likely Benign | -1.74 | Neutral | 0.411 | Benign | 0.139 | Benign | 2.25 | Pathogenic | 0.01 | Affected | 0.1537 | 0.3979 | 0 | 0 | -2.8 | 13.00 | |||||||||||||||||||||||||||||||||||||||
| c.3076G>A | D1026N 2D AIThe SynGAP1 D1026N variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The high‑accuracy consensus, SGM‑Consensus, is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN and therefore reports a likely benign outcome. AlphaMissense‑Optimized also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.993931 | Binding | 0.324 | 0.739 | 0.500 | -4.166 | Likely Benign | 0.608 | Likely Pathogenic | Likely Benign | 0.117 | Likely Benign | -2.07 | Neutral | 0.411 | Benign | 0.239 | Benign | 2.55 | Benign | 0.01 | Affected | 0.1240 | 0.5060 | 2 | 1 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3131C>T | P1044L 2D AIThe SynGAP1 missense variant P1044L is not represented in ClinVar (no ClinVar ID) and is absent from gnomAD. All evaluated in‑silico predictors classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool reports a pathogenic or likely pathogenic outcome. High‑accuracy assessments reinforce this benign prediction: AlphaMissense‑Optimized indicates benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.979741 | Disordered | 0.952126 | Binding | 0.331 | 0.855 | 0.750 | -4.327 | Likely Benign | 0.144 | Likely Benign | Likely Benign | 0.418 | Likely Benign | -1.64 | Neutral | 0.411 | Benign | 0.187 | Benign | 5.43 | Benign | 0.15 | Tolerated | 0.2264 | 0.6586 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.3134C>A | A1045D 2D AIThe SynGAP1 missense variant A1045D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that reach consensus all indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the variant as benign. AlphaMissense‑Optimized also predicts a benign outcome, whereas AlphaMissense‑Default remains uncertain. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as “Likely Benign.” No tools predict pathogenicity. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the collective predictions strongly suggest that A1045D is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.975609 | Disordered | 0.948874 | Binding | 0.352 | 0.882 | 0.750 | -5.734 | Likely Benign | 0.418 | Ambiguous | Likely Benign | 0.066 | Likely Benign | -1.00 | Neutral | 0.411 | Benign | 0.172 | Benign | 2.64 | Benign | 0.16 | Tolerated | 0.1954 | 0.2102 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.3136C>A | P1046T 2D AIThe SynGAP1 missense variant P1046T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.970265 | Disordered | 0.942366 | Binding | 0.364 | 0.898 | 0.750 | -5.249 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.052 | Likely Benign | -1.18 | Neutral | 0.411 | Benign | 0.131 | Benign | 2.37 | Pathogenic | 0.21 | Tolerated | 0.1430 | 0.5934 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3149G>A | G1050E 2D AIThe SynGAP1 missense variant G1050E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. There is no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987317 | Disordered | 0.906802 | Binding | 0.370 | 0.928 | 0.875 | -8.175 | Likely Pathogenic | 0.266 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -0.28 | Neutral | 0.411 | Benign | 0.171 | Benign | 2.50 | Benign | 0.08 | Tolerated | 0.1418 | 0.4258 | 0 | -2 | -3.1 | 72.06 | |||||||||||||||||||||||||||||||||||||||
| c.3181G>C | G1061R 2D AIThe SynGAP1 missense variant G1061R is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it contains two benign and two uncertain calls, and Foldetta results are unavailable. Overall, the balance of evidence favors a benign classification. This conclusion does not contradict ClinVar status, as the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.978672 | Disordered | 0.926729 | Binding | 0.394 | 0.923 | 0.875 | -7.721 | In-Between | 0.343 | Ambiguous | Likely Benign | 0.315 | Likely Benign | -0.17 | Neutral | 0.411 | Benign | 0.132 | Benign | 3.99 | Benign | 0.00 | Affected | 0.1037 | 0.4332 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.3187G>C | G1063R 2D AIThe SynGAP1 missense variant G1063R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the collective evidence strongly supports a benign classification, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.975134 | Disordered | 0.945784 | Binding | 0.394 | 0.913 | 0.875 | -5.711 | Likely Benign | 0.391 | Ambiguous | Likely Benign | 0.078 | Likely Benign | 0.55 | Neutral | 0.411 | Benign | 0.114 | Benign | 4.28 | Benign | 0.09 | Tolerated | 0.1037 | 0.5133 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||||||
| c.3188G>A | G1063D 2D AIThe SynGAP1 missense variant G1063D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the computational evidence overwhelmingly supports a benign impact for G1063D, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.975134 | Disordered | 0.945784 | Binding | 0.394 | 0.913 | 0.875 | -6.950 | Likely Benign | 0.367 | Ambiguous | Likely Benign | 0.057 | Likely Benign | -0.30 | Neutral | 0.411 | Benign | 0.058 | Benign | 4.24 | Benign | 0.08 | Tolerated | 0.1878 | 0.3026 | 1 | -1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||||||
| c.3365G>A | G1122D 2D AIThe SynGAP1 missense variant G1122D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors benign (2 benign vs. 1 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that G1122D is most likely benign, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.896620 | Disordered | 0.814918 | Binding | 0.357 | 0.932 | 0.875 | -9.991 | Likely Pathogenic | 0.430 | Ambiguous | Likely Benign | 0.332 | Likely Benign | -0.49 | Neutral | 0.411 | Benign | 0.091 | Benign | 4.49 | Benign | 0.10 | Tolerated | 0.1746 | 0.2035 | 1 | -1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3404A>C | K1135T 2D AIThe SynGAP1 missense variant K1135T is listed in ClinVar (ID 1166087.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443956‑A‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.790969 | Binding | 0.303 | 0.889 | 0.875 | Conflicting | 2 | 6-33443956-A-C | 1 | 6.75e-7 | -4.778 | Likely Benign | 0.779 | Likely Pathogenic | Likely Benign | 0.210 | Likely Benign | -0.90 | Neutral | 0.411 | Benign | 0.321 | Benign | 5.46 | Benign | 0.10 | Tolerated | 4.32 | 2 | 0.2544 | 0.3521 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||
| c.3405G>C | K1135N 2D AIThe SynGAP1 missense variant K1135N is listed in ClinVar (ID 633521.0) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. In contrast, AlphaMissense‑Default and AlphaMissense‑Optimized both predict a pathogenic outcome. High‑accuracy assessments further show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) remains Likely Benign. No Foldetta (FoldX‑MD/Rosetta stability) result is available for this variant. Overall, the majority of predictions support a benign classification, which does not contradict the current ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.790969 | Binding | 0.303 | 0.889 | 0.875 | Uncertain | 1 | -5.715 | Likely Benign | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.166 | Likely Benign | -0.97 | Neutral | 0.411 | Benign | 0.321 | Benign | 5.43 | Benign | 0.07 | Tolerated | 4.32 | 2 | 0.4152 | 0.1793 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||
| c.3405G>T | K1135N 2D AIThe SynGAP1 missense variant K1135N is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33443957‑G‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are AlphaMissense‑Default and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains likely benign; Foldetta results are unavailable. Overall, the majority of predictions (seven benign vs. two pathogenic) support a benign interpretation. This consensus does not contradict ClinVar, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.790969 | Binding | 0.303 | 0.889 | 0.875 | 6-33443957-G-T | -5.715 | Likely Benign | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.166 | Likely Benign | -0.97 | Neutral | 0.411 | Benign | 0.321 | Benign | 5.43 | Benign | 0.07 | Tolerated | 4.32 | 2 | 0.4152 | 0.1793 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||
| c.3440C>G | T1147R 2D AIThe SynGAP1 missense variant T1147R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta results are unavailable. Taken together, the preponderance of evidence points to a benign impact for T1147R, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.831250 | Disordered | 0.746520 | Binding | 0.345 | 0.839 | 0.875 | -3.234 | Likely Benign | 0.534 | Ambiguous | Likely Benign | 0.442 | Likely Benign | -2.44 | Neutral | 0.411 | Benign | 0.139 | Benign | 5.46 | Benign | 0.01 | Affected | 0.1014 | 0.2742 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3829C>G | H1277D 2D AIThe SynGAP1 missense variant H1277D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447877‑C‑G). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), and ESM1b, while those that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. AlphaMissense‑Default is uncertain, whereas AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a pathogenic outcome (two pathogenic, one benign, one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy and consensus predictions indicate a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | 6-33447877-C-G | -4.632 | Likely Benign | 0.537 | Ambiguous | Likely Benign | 0.172 | Likely Benign | -6.38 | Deleterious | 0.411 | Benign | 0.091 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2389 | 0.1266 | -1 | 1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||
| c.238A>C | K80Q 2D  AIThe SynGAP1 missense variant K80Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence points to a benign impact, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.477530 | Uncertain | 0.331 | 0.873 | 0.500 | -4.475 | Likely Benign | 0.662 | Likely Pathogenic | Likely Benign | 0.064 | Likely Benign | -0.81 | Neutral | 0.414 | Benign | 0.040 | Benign | 3.93 | Benign | 0.00 | Affected | 0.4373 | 0.1132 | 1 | 1 | 0.4 | -0.04 | |||||||||||||||||||||||||||||||||||||||
| c.3948T>A | N1316K 2D AIThe SynGAP1 missense variant N1316K is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451822‑T‑A). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none). Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451822-T-A | -3.815 | Likely Benign | 0.699 | Likely Pathogenic | Likely Benign | 0.044 | Likely Benign | -3.27 | Deleterious | 0.414 | Benign | 0.063 | Benign | 4.00 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2065 | 0.5500 | 0 | 1 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||
| c.3948T>G | N1316K 2D AIThe SynGAP1 missense variant N1316K is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451822‑T‑G). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign + 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451822-T-G | -3.815 | Likely Benign | 0.699 | Likely Pathogenic | Likely Benign | 0.044 | Likely Benign | -3.27 | Deleterious | 0.414 | Benign | 0.063 | Benign | 4.00 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2065 | 0.5500 | 0 | 1 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||
| c.3968C>T | P1323L 2D AIThe SynGAP1 missense variant P1323L is reported in gnomAD (variant ID 6‑33451842‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority‑vote) is benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451842-C-T | 3 | 1.95e-6 | -6.005 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.045 | Likely Benign | -1.04 | Neutral | 0.414 | Benign | 0.175 | Benign | 3.82 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2650 | 0.6052 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||
| c.3971C>G | P1324R 2D AIThe SynGAP1 missense variant P1324R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | -5.718 | Likely Benign | 0.154 | Likely Benign | Likely Benign | 0.053 | Likely Benign | -0.80 | Neutral | 0.414 | Benign | 0.133 | Benign | 4.28 | Benign | 0.00 | Affected | 0.1723 | 0.3613 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3971C>T | P1324L 2D AIThe SynGAP1 missense variant P1324L is reported in gnomAD (ID 6‑33451845‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | 6-33451845-C-T | -5.549 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.072 | Likely Benign | -1.17 | Neutral | 0.414 | Benign | 0.175 | Benign | 4.26 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2507 | 0.5864 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||
| c.1187G>A | G396D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G396D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the substitution as benign, whereas only AlphaMissense‑Default predicts a pathogenic outcome. Tools that assess protein stability (FoldX, Rosetta, Foldetta) yield uncertain or inconclusive results. High‑accuracy consensus methods reinforce the benign assessment: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports “Likely Benign”; AlphaMissense‑Optimized also predicts benign; Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains uncertain. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.394626 | Uncertain | 0.640 | 0.584 | 0.500 | -4.148 | Likely Benign | 0.678 | Likely Pathogenic | Likely Benign | 1.92 | Ambiguous | 0.8 | 1.33 | Ambiguous | 1.63 | Ambiguous | 0.17 | Likely Benign | 0.272 | Likely Benign | -1.49 | Neutral | 0.421 | Benign | 0.080 | Benign | 3.91 | Benign | 0.35 | Tolerated | 0.1702 | 0.1122 | 1 | -1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||
| c.1196C>A | A399D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A399D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that agree on a pathogenic effect are REVEL, FoldX, SIFT, ESM1b, and AlphaMissense‑Default. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized—return uncertain or inconclusive results. High‑accuracy methods give no definitive verdict: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable, and Foldetta is uncertain. Overall, the majority of available predictions (five pathogenic vs. four benign) lean toward pathogenicity. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.394753 | Structured | 0.407674 | Uncertain | 0.939 | 0.490 | 0.125 | -10.441 | Likely Pathogenic | 0.943 | Likely Pathogenic | Ambiguous | 2.45 | Destabilizing | 0.2 | 1.17 | Ambiguous | 1.81 | Ambiguous | 0.91 | Ambiguous | 0.525 | Likely Pathogenic | -1.84 | Neutral | 0.421 | Benign | 0.054 | Benign | 5.38 | Benign | 0.05 | Affected | 0.1533 | 0.1760 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1196C>T | A399V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A399V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). No tool predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Overall, the evidence overwhelmingly supports a benign classification, and this is consistent with the lack of ClinVar reporting. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.394753 | Structured | 0.407674 | Uncertain | 0.939 | 0.490 | 0.125 | -4.943 | Likely Benign | 0.170 | Likely Benign | Likely Benign | 0.49 | Likely Benign | 0.2 | 0.56 | Ambiguous | 0.53 | Ambiguous | 0.22 | Likely Benign | 0.305 | Likely Benign | -1.18 | Neutral | 0.421 | Benign | 0.073 | Benign | 5.37 | Benign | 0.24 | Tolerated | 0.1145 | 0.6807 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1848T>A | D616E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D616E missense variant is catalogued in gnomAD (ID 6‑33440900‑T‑A) but has no ClinVar submission. Functional prediction tools show a split assessment: benign calls come from REVEL, both polyPhen‑2 HumDiv and HumVar, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls arise from PROVEAN, SIFT, and AlphaMissense‑Default. The remaining predictors (FoldX, Rosetta, Foldetta, premPS, ESM1b) are inconclusive. A high‑accuracy consensus (SGM) that aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a pathogenic verdict. AlphaMissense‑Optimized remains benign, and Foldetta, which evaluates protein‑folding stability, is uncertain. Overall, the majority of evidence leans toward pathogenicity, and this conclusion does not conflict with ClinVar because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.166689 | Uncertain | 0.867 | 0.252 | 0.000 | 6-33440900-T-A | 1 | 6.20e-7 | -7.250 | In-Between | 0.695 | Likely Pathogenic | Likely Benign | 0.96 | Ambiguous | 0.1 | 1.52 | Ambiguous | 1.24 | Ambiguous | 0.58 | Ambiguous | 0.092 | Likely Benign | -2.85 | Deleterious | 0.421 | Benign | 0.232 | Benign | 3.32 | Benign | 0.03 | Affected | 3.37 | 35 | 0.1225 | 0.4128 | 2 | 3 | 0.0 | 14.03 | |||||||||||||||||||||||||
| c.1848T>G | D616E 2D 3DClick to see structure in 3D Viewer AISynGAP1 D616E is not reported in ClinVar but is present in gnomAD (ID 6‑33440900‑T‑G). Functional prediction tools that agree on benign impact include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, SIFT, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b) are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of conventional predictors lean toward a benign effect, but the high‑accuracy consensus is split, leaving the variant’s clinical significance unresolved. Thus, the variant is most likely benign based on the bulk of predictions, and this does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.129801 | Structured | 0.166689 | Uncertain | 0.867 | 0.252 | 0.000 | 6-33440900-T-G | 3 | 1.86e-6 | -7.250 | In-Between | 0.695 | Likely Pathogenic | Likely Benign | 0.96 | Ambiguous | 0.1 | 1.52 | Ambiguous | 1.24 | Ambiguous | 0.58 | Ambiguous | 0.092 | Likely Benign | -2.85 | Deleterious | 0.421 | Benign | 0.232 | Benign | 3.32 | Benign | 0.03 | Affected | 3.37 | 35 | 0.1225 | 0.4128 | 2 | 3 | 0.0 | 14.03 | |||||||||||||||||||||||||
| c.2677C>G | Q893E 2D AIThe SynGAP1 missense variant Q893E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that assess sequence conservation and structural impact all converge on a benign interpretation: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. No tool in the dataset indicates pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence strongly supports a benign effect, and this conclusion is consistent with the absence of any ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.447267 | Uncertain | 0.310 | 0.925 | 0.750 | -3.888 | Likely Benign | 0.296 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.67 | Neutral | 0.421 | Benign | 0.181 | Benign | 2.76 | Benign | 0.06 | Tolerated | 0.1535 | 0.2555 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3109A>C | I1037L 2D AIThe SynGAP1 missense variant I1037L is not reported in ClinVar and is absent from gnomAD, indicating no known clinical or population evidence. Functional prediction tools uniformly favor a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity, and the only uncertain call comes from AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as “Likely Benign.” High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that I1037L is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.939629 | Disordered | 0.986140 | Binding | 0.309 | 0.774 | 0.625 | -2.563 | Likely Benign | 0.448 | Ambiguous | Likely Benign | 0.074 | Likely Benign | -0.46 | Neutral | 0.421 | Benign | 0.128 | Benign | 2.82 | Benign | 0.81 | Tolerated | 0.0939 | 0.4302 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3109A>G | I1037V 2D AIThe SynGAP1 missense variant I1037V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that I1037V is most likely benign, and this conclusion is not contradicted by ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.939629 | Disordered | 0.986140 | Binding | 0.309 | 0.774 | 0.625 | -2.326 | Likely Benign | 0.461 | Ambiguous | Likely Benign | 0.085 | Likely Benign | -0.09 | Neutral | 0.421 | Benign | 0.128 | Benign | 2.76 | Benign | 0.93 | Tolerated | 0.1229 | 0.3985 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3169A>G | S1057G 2D AIThe SynGAP1 missense variant S1057G is reported in gnomAD (ID 6‑33443721‑A‑G) but has no ClinVar entry. All evaluated in‑silico predictors uniformly classify the change as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. The high‑accuracy consensus methods likewise support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Consequently, the variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.869507 | Binding | 0.413 | 0.927 | 0.875 | 6-33443721-A-G | 1 | 7.20e-7 | -1.005 | Likely Benign | 0.049 | Likely Benign | Likely Benign | 0.211 | Likely Benign | -0.47 | Neutral | 0.421 | Benign | 0.111 | Benign | 5.76 | Benign | 0.52 | Tolerated | 3.77 | 5 | 0.2654 | 0.4846 | 0 | 1 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||||||
| c.316A>G | R106G 2D AIThe SynGAP1 missense variant R106G is listed in gnomAD (ID 6‑33432181‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Those that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. There is no ClinVar classification to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.663409 | Binding | 0.345 | 0.862 | 0.875 | 6-33432181-A-G | 1 | 6.20e-7 | -2.617 | Likely Benign | 0.835 | Likely Pathogenic | Ambiguous | 0.161 | Likely Benign | -2.21 | Neutral | 0.421 | Benign | 0.050 | Benign | 3.65 | Benign | 0.00 | Affected | 4.05 | 3 | 0.3680 | 0.3980 | -2 | -3 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||
| c.319A>G | R107G 2D AIThe SynGAP1 missense variant R107G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM, while those that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a tie and thus unavailable, and Foldetta stability analysis is missing. Overall, the majority of tools (five benign vs. three pathogenic) suggest a benign impact, but the lack of consensus from the most accurate predictors means the variant’s effect remains uncertain. This assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.663448 | Binding | 0.331 | 0.863 | 0.875 | -3.486 | Likely Benign | 0.948 | Likely Pathogenic | Ambiguous | 0.180 | Likely Benign | -3.15 | Deleterious | 0.421 | Benign | 0.050 | Benign | 2.98 | Benign | 0.00 | Affected | 0.3280 | 0.4000 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.320G>C | R107T 2D AIThe SynGAP1 missense variant R107T is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign) and Foldetta results are unavailable. Overall, the majority of predictions (five benign vs. four pathogenic) lean toward a benign interpretation. Thus, the variant is most likely benign based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.663448 | Binding | 0.331 | 0.863 | 0.875 | -2.902 | Likely Benign | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.191 | Likely Benign | -2.63 | Deleterious | 0.421 | Benign | 0.050 | Benign | 2.98 | Benign | 0.00 | Affected | 0.1549 | 0.4761 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.331C>A | P111T 2D AIThe SynGAP1 missense variant P111T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.650020 | Binding | 0.438 | 0.858 | 0.750 | -2.800 | Likely Benign | 0.305 | Likely Benign | Likely Benign | 0.061 | Likely Benign | -1.44 | Neutral | 0.421 | Benign | 0.050 | Benign | 4.11 | Benign | 0.02 | Affected | 0.1650 | 0.5402 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.332C>A | P111Q 2D AIThe SynGAP1 missense variant P111Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign status. High‑accuracy assessments further support this view: AlphaMissense‑Optimized indicates benign, SGM‑Consensus confirms likely benign, and Foldetta data are unavailable. Taken together, the preponderance of evidence points to a benign effect for P111Q, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.650020 | Binding | 0.438 | 0.858 | 0.750 | -4.726 | Likely Benign | 0.543 | Ambiguous | Likely Benign | 0.079 | Likely Benign | -2.10 | Neutral | 0.421 | Benign | 0.054 | Benign | 4.06 | Benign | 0.00 | Affected | 0.1593 | 0.4232 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||||
| c.332C>G | P111R 2D AIThe SynGAP1 missense variant P111R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta results are unavailable for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.650020 | Binding | 0.438 | 0.858 | 0.750 | -4.811 | Likely Benign | 0.782 | Likely Pathogenic | Likely Benign | 0.100 | Likely Benign | -2.34 | Neutral | 0.421 | Benign | 0.075 | Benign | 4.06 | Benign | 0.00 | Affected | 0.1578 | 0.3015 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.332C>T | P111L 2D AIThe SynGAP1 missense variant P111L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also yields a benign prediction (2 benign vs. 1 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that P111L is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.707965 | Disordered | 0.650020 | Binding | 0.438 | 0.858 | 0.750 | -4.430 | Likely Benign | 0.486 | Ambiguous | Likely Benign | 0.089 | Likely Benign | -2.81 | Deleterious | 0.421 | Benign | 0.055 | Benign | 4.06 | Benign | 0.00 | Affected | 0.2355 | 0.7085 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.335G>A | G112E 2D AIThe SynGAP1 missense variant G112E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized result is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta predictions are not available. Overall, the majority of evidence (five benign vs. three pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.728858 | Disordered | 0.640153 | Binding | 0.332 | 0.867 | 0.750 | -3.470 | Likely Benign | 0.818 | Likely Pathogenic | Ambiguous | 0.134 | Likely Benign | -3.30 | Deleterious | 0.421 | Benign | 0.146 | Benign | 3.96 | Benign | 0.00 | Affected | 0.1330 | 0.3814 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.335G>T | G112V 2D AIThe SynGAP1 missense variant G112V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.728858 | Disordered | 0.640153 | Binding | 0.332 | 0.867 | 0.750 | -2.411 | Likely Benign | 0.230 | Likely Benign | Likely Benign | 0.159 | Likely Benign | -3.39 | Deleterious | 0.421 | Benign | 0.108 | Benign | 3.96 | Benign | 0.00 | Affected | 0.1225 | 0.4218 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.356A>G | E119G 2D AIThe SynGAP1 missense variant E119G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status because no ClinVar claim exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.690604 | Disordered | 0.661946 | Binding | 0.346 | 0.881 | 0.750 | -4.349 | Likely Benign | 0.584 | Likely Pathogenic | Likely Benign | 0.143 | Likely Benign | -2.40 | Neutral | 0.421 | Benign | 0.055 | Benign | 3.82 | Benign | 0.01 | Affected | 0.3246 | 0.6405 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||||||
| c.368C>G | A123G 2D AIThe SynGAP1 missense variant A123G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.521092 | Disordered | 0.689505 | Binding | 0.324 | 0.886 | 0.750 | -2.799 | Likely Benign | 0.121 | Likely Benign | Likely Benign | 0.065 | Likely Benign | -1.34 | Neutral | 0.421 | Benign | 0.074 | Benign | 4.13 | Benign | 0.03 | Affected | 0.2070 | 0.5039 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3941C>T | P1314L 2D AIThe SynGAP1 missense variant P1314L is listed in ClinVar as a benign alteration (ClinVar ID 646689.0) and is present in the gnomAD database (gnomAD ID 6‑33451815‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, which is consistent with the ClinVar classification. Thus, the variant is most likely benign and does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.971592 | Binding | 0.467 | 0.903 | 0.750 | Likely Benign | 1 | 6-33451815-C-T | 2 | 1.24e-6 | -4.040 | Likely Benign | 0.118 | Likely Benign | Likely Benign | 0.049 | Likely Benign | -0.20 | Neutral | 0.421 | Benign | 0.066 | Benign | 4.19 | Benign | 0.05 | Affected | 3.77 | 5 | 0.2310 | 0.5967 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||
| c.527G>A | S176N 2D AIThe SynGAP1 missense variant S176N is catalogued in gnomAD (ID 6‑33435169‑G‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none reported). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.466016 | Uncertain | 0.380 | 0.597 | 0.375 | 6-33435169-G-A | 1 | 6.20e-7 | -6.286 | Likely Benign | 0.594 | Likely Pathogenic | Likely Benign | 0.070 | Likely Benign | -0.56 | Neutral | 0.421 | Benign | 0.080 | Benign | 4.10 | Benign | 0.22 | Tolerated | 3.54 | 6 | 0.0996 | 0.3930 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||
| c.527G>C | S176T 2D AIThe SynGAP1 missense variant S176T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.562014 | Disordered | 0.466016 | Uncertain | 0.380 | 0.597 | 0.375 | -3.682 | Likely Benign | 0.351 | Ambiguous | Likely Benign | 0.045 | Likely Benign | -0.73 | Neutral | 0.421 | Benign | 0.054 | Benign | 4.09 | Benign | 0.19 | Tolerated | 0.1103 | 0.4979 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.541C>A | H181N 2D AIThe SynGAP1 missense variant H181N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign prediction (2 benign vs. 1 pathogenic). Foldetta results are unavailable. Overall, the preponderance of evidence indicates that H181N is most likely benign, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.439530 | Uncertain | 0.294 | 0.616 | 0.500 | -10.315 | Likely Pathogenic | 0.526 | Ambiguous | Likely Benign | 0.090 | Likely Benign | -1.50 | Neutral | 0.421 | Benign | 0.107 | Benign | 4.18 | Benign | 0.05 | Affected | 0.1335 | 0.1839 | 2 | 1 | -0.3 | -23.04 | ||||||||||||||||||||||||||||||||||||||||
| c.547C>G | H183D 2D AIThe SynGAP1 missense variant H183D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Pathogenic, and AlphaMissense‑Optimized independently predicts Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant. Overall, the majority of predictions (seven pathogenic vs. four benign) indicate a pathogenic impact. This conclusion is not contradicted by ClinVar status, as no ClinVar entry exists for H183D. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.432952 | Uncertain | 0.421 | 0.622 | 0.500 | -18.626 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.311 | Likely Benign | -6.55 | Deleterious | 0.421 | Benign | 0.107 | Benign | 3.81 | Benign | 0.01 | Affected | 0.2620 | 0.1817 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.548A>T | H183L 2D AIThe SynGAP1 missense variant H183L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Pathogenic” verdict (3 pathogenic vs. 1 benign votes). AlphaMissense‑Optimized alone also predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.432952 | Uncertain | 0.421 | 0.622 | 0.500 | -12.898 | Likely Pathogenic | 0.968 | Likely Pathogenic | Likely Pathogenic | 0.325 | Likely Benign | -8.12 | Deleterious | 0.421 | Benign | 0.058 | Benign | 3.79 | Benign | 0.01 | Affected | 0.0911 | 0.5304 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||||||||||||
| c.572G>T | S191I 2D AIThe SynGAP1 missense variant S191I is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split opinion: benign predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), and FATHMM, while pathogenic predictions arise from PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. When the predictions are grouped by consensus, four tools favor benign and four favor pathogenic. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized returns an uncertain result, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Taken together, the majority of evidence, including the SGM Consensus, points to a pathogenic effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.428475 | Uncertain | 0.322 | 0.615 | 0.125 | -11.271 | Likely Pathogenic | 0.927 | Likely Pathogenic | Ambiguous | 0.283 | Likely Benign | -4.51 | Deleterious | 0.421 | Benign | 0.086 | Benign | 3.76 | Benign | 0.00 | Affected | 0.0954 | 0.6842 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.614T>C | I205T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I205T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; the remaining predictions (FoldX, premPS, AlphaMissense‑Default) are uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, classifies the variant as benign. Taken together, the consensus of both general and high‑accuracy predictors indicates that I205T is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.264545 | Structured | 0.409933 | Uncertain | 0.821 | 0.414 | 0.125 | -6.869 | Likely Benign | 0.450 | Ambiguous | Likely Benign | 0.65 | Ambiguous | 0.0 | 0.28 | Likely Benign | 0.47 | Likely Benign | 0.81 | Ambiguous | 0.118 | Likely Benign | -2.19 | Neutral | 0.421 | Benign | 0.156 | Benign | 4.16 | Benign | 0.15 | Tolerated | 0.0933 | 0.0541 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.617T>C | I206T 2D AIThe SynGAP1 I206T missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, the majority of tools predict a pathogenic impact: SGM‑Consensus (Likely Pathogenic), FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods give consistent results: AlphaMissense‑Optimized is pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No predictions or stability results are missing or inconclusive. Based on the overall evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not currently listed. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.298791 | Structured | 0.405123 | Uncertain | 0.863 | 0.391 | 0.125 | -10.812 | Likely Pathogenic | 0.973 | Likely Pathogenic | Likely Pathogenic | 2.43 | Destabilizing | 0.5 | 2.68 | Destabilizing | 2.56 | Destabilizing | 1.94 | Destabilizing | 0.252 | Likely Benign | -3.78 | Deleterious | 0.421 | Benign | 0.156 | Benign | 3.65 | Benign | 0.00 | Affected | 0.0830 | 0.0669 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.663G>C | E221D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E221D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that agree on a pathogenic effect are REVEL, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Four tools (FoldX, Rosetta, Foldetta, premPS) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs 2 benign), and Foldetta remains uncertain. Overall, the majority of available predictions lean toward pathogenicity, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.127496 | Structured | 0.413334 | Uncertain | 0.891 | 0.283 | 0.000 | -12.237 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 1.59 | Ambiguous | 0.1 | 1.07 | Ambiguous | 1.33 | Ambiguous | 0.84 | Ambiguous | 0.750 | Likely Pathogenic | -2.43 | Neutral | 0.421 | Benign | 0.107 | Benign | 5.80 | Benign | 0.02 | Affected | 0.1755 | 0.5228 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.663G>T | E221D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E221D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that agree on a pathogenic effect are REVEL, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Four tools (FoldX, Rosetta, Foldetta, premPS) returned uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta is uncertain. Overall, the majority of available predictions lean toward pathogenicity, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.127496 | Structured | 0.413334 | Uncertain | 0.891 | 0.283 | 0.000 | -12.237 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 1.59 | Ambiguous | 0.1 | 1.07 | Ambiguous | 1.33 | Ambiguous | 0.84 | Ambiguous | 0.750 | Likely Pathogenic | -2.43 | Neutral | 0.421 | Benign | 0.107 | Benign | 5.80 | Benign | 0.02 | Affected | 0.1755 | 0.5228 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||
| c.667A>T | T223S 2D 3DClick to see structure in 3D Viewer AISynGAP1 T223S is listed in ClinVar as a variant of uncertain significance and is present in the gnomAD database (ID 6‑33435518‑A‑T). Functional prediction tools that reach consensus classify the variant as benign: FoldX, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict pathogenicity include REVEL, PROVEAN, and SIFT. Predictions that are inconclusive or uncertain are Rosetta, premPS, AlphaMissense‑Default, and ESM1b. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, Foldetta is benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 1‑to‑1 split between benign and pathogenic calls. Overall, the majority of evidence points to a benign effect, which does not contradict the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.070400 | Structured | 0.382605 | Uncertain | 0.867 | 0.316 | 0.125 | Conflicting | 2 | 6-33435518-A-T | 3 | 1.86e-6 | -7.714 | In-Between | 0.410 | Ambiguous | Likely Benign | 0.26 | Likely Benign | 0.1 | 0.50 | Ambiguous | 0.38 | Likely Benign | 0.62 | Ambiguous | 0.535 | Likely Pathogenic | -2.86 | Deleterious | 0.421 | Benign | 0.058 | Benign | 5.80 | Benign | 0.02 | Affected | 3.41 | 13 | 0.2388 | 0.2972 | 1 | 1 | -0.1 | -14.03 | 200.7 | 17.3 | -0.2 | 0.2 | 0.0 | 0.0 | X | Uncertain | The introduced residue Ser223 is located on the outer surface of an anti-parallel β sheet strand (res. Cys219-Thr224). Its hydroxyl group forms hydrogen bonds with nearby residues Thr228 and Lys207 in the variant simulations, similar to the hydroxyl group of Thr223 in the WT simulations. These hydrogen-bonding interactions at the β sheet surface contribute to the stability of the secondary structure element and may prevent it from unfolding. However, since the model ends abruptly at the N-terminus, no definite conclusions can be drawn from the simulations. | ||||||||||||||
| c.688T>A | C230S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C230S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are REVEL, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as benign. With an equal split of 7 benign versus 7 pathogenic predictions overall, the higher‑confidence tools lean toward pathogenicity. Therefore, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.268042 | Structured | 0.308076 | Uncertain | 0.870 | 0.308 | 0.000 | -9.994 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.42 | Likely Benign | 0.1 | 0.05 | Likely Benign | 0.24 | Likely Benign | 1.08 | Destabilizing | 0.859 | Likely Pathogenic | -8.24 | Deleterious | 0.421 | Benign | 0.115 | Benign | 5.91 | Benign | 0.07 | Tolerated | 0.4926 | 0.1822 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.689G>C | C230S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant C230S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools cluster into two groups: benign calls come from FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM; pathogenic calls come from REVEL, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic, and Foldetta as benign. No prediction or stability result is missing. Overall, the predictions are evenly split between benign and pathogenic, leaving the variant’s functional impact uncertain. This uncertainty does not contradict ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.268042 | Structured | 0.308076 | Uncertain | 0.870 | 0.308 | 0.000 | -9.994 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.42 | Likely Benign | 0.1 | 0.05 | Likely Benign | 0.24 | Likely Benign | 1.08 | Destabilizing | 0.844 | Likely Pathogenic | -8.24 | Deleterious | 0.421 | Benign | 0.115 | Benign | 5.91 | Benign | 0.07 | Tolerated | 0.4926 | 0.1822 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.697T>A | C233S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C233S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are REVEL, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. FoldX, Rosetta, and Foldetta give uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors pathogenicity (3 pathogenic vs 1 benign). Foldetta remains uncertain. Overall, the majority of evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.239899 | Structured | 0.306787 | Uncertain | 0.868 | 0.322 | 0.000 | -10.862 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.61 | Ambiguous | 0.1 | 1.25 | Ambiguous | 0.93 | Ambiguous | 1.50 | Destabilizing | 0.764 | Likely Pathogenic | -8.89 | Deleterious | 0.421 | Benign | 0.080 | Benign | 5.79 | Benign | 0.03 | Affected | 0.4528 | 0.2833 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.698G>C | C233S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C233S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, while pathogenic predictions arise from SGM‑Consensus, REVEL, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Pathogenic.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, yields an uncertain result and is treated as unavailable evidence. Overall, the preponderance of evidence points to a pathogenic impact for C233S, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.239899 | Structured | 0.306787 | Uncertain | 0.868 | 0.322 | 0.000 | -10.862 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.61 | Ambiguous | 0.1 | 1.25 | Ambiguous | 0.93 | Ambiguous | 1.50 | Destabilizing | 0.830 | Likely Pathogenic | -8.89 | Deleterious | 0.421 | Benign | 0.080 | Benign | 5.79 | Benign | 0.03 | Affected | 0.4528 | 0.2833 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.2657C>A | A886E 2D AIThe SynGAP1 missense variant A886E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome. Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.716283 | Disordered | 0.619166 | Binding | 0.359 | 0.922 | 0.500 | -3.747 | Likely Benign | 0.422 | Ambiguous | Likely Benign | 0.096 | Likely Benign | -1.31 | Neutral | 0.423 | Benign | 0.230 | Benign | 2.17 | Pathogenic | 0.00 | Affected | 0.1483 | 0.2008 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.2213G>A | S738N 2D AIThe SynGAP1 missense variant S738N is reported in ClinVar as having no entry and is not present in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a benign majority vote, matching the SGM‑Consensus label of “Likely Benign.” Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this assessment is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.441162 | Uncertain | 0.284 | 0.827 | 0.875 | -5.005 | Likely Benign | 0.075 | Likely Benign | Likely Benign | 0.059 | Likely Benign | -0.53 | Neutral | 0.425 | Benign | 0.233 | Benign | 2.69 | Benign | 0.02 | Affected | 0.1284 | 0.3251 | 1 | 1 | -2.7 | 27.03 | |||||||||||||||||||||||||||||||||||||||
| c.2221C>G | P741A 2D AIThe SynGAP1 missense variant P741A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign,” while Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.885302 | Disordered | 0.493550 | Uncertain | 0.354 | 0.859 | 0.875 | -3.995 | Likely Benign | 0.054 | Likely Benign | Likely Benign | 0.094 | Likely Benign | -0.33 | Neutral | 0.425 | Benign | 0.136 | Benign | 3.01 | Benign | 0.98 | Tolerated | 0.2942 | 0.3883 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.2231A>T | Q744L 2D AIThe SynGAP1 missense variant Q744L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta results are not available for this variant. Overall, the preponderance of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.540428 | Binding | 0.316 | 0.866 | 0.875 | -3.724 | Likely Benign | 0.100 | Likely Benign | Likely Benign | 0.052 | Likely Benign | -1.96 | Neutral | 0.425 | Benign | 0.158 | Benign | 2.70 | Benign | 0.02 | Affected | 0.0813 | 0.4731 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.2240T>C | V747A 2D AIThe SynGAP1 missense variant V747A is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) indicates likely benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the majority of computational evidence supports a benign classification for V747A, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.594069 | Binding | 0.343 | 0.873 | 0.750 | -3.118 | Likely Benign | 0.171 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -0.69 | Neutral | 0.425 | Benign | 0.252 | Benign | 2.73 | Benign | 0.00 | Affected | 0.2585 | 0.1714 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.2279T>C | M760T 2D AIThe SynGAP1 missense variant M760T is reported in gnomAD (ID 6‑33441744‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the consensus of available predictions indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.541878 | Disordered | 0.893402 | Binding | 0.346 | 0.865 | 0.375 | 6-33441744-T-C | 1 | 6.20e-7 | -2.365 | Likely Benign | 0.519 | Ambiguous | Likely Benign | 0.096 | Likely Benign | -1.56 | Neutral | 0.425 | Benign | 0.252 | Benign | 2.71 | Benign | 0.41 | Tolerated | 3.99 | 5 | 0.2515 | 0.2875 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||
| c.2750C>T | P917L 2D AIThe SynGAP1 missense variant P917L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.599170 | Disordered | 0.863949 | Binding | 0.314 | 0.862 | 0.375 | -3.369 | Likely Benign | 0.172 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -2.35 | Neutral | 0.425 | Benign | 0.233 | Benign | 2.75 | Benign | 0.00 | Affected | 0.2001 | 0.6351 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.3704T>C | M1235T 2D AIThe SynGAP1 missense variant M1235T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of tools, including the high‑accuracy methods, points to a benign impact. This prediction does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.690604 | Disordered | 0.577958 | Binding | 0.872 | 0.532 | 0.125 | -5.381 | Likely Benign | 0.149 | Likely Benign | Likely Benign | 0.060 | Likely Benign | -2.39 | Neutral | 0.425 | Benign | 0.158 | Benign | 2.66 | Benign | 0.01 | Affected | 0.2079 | 0.1814 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||||||
| c.3716C>T | A1239V 2D AIThe A1239V missense change occurs in a coiled‑coil region of SynGAP1. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. ClinVar has no entry for this variant, and it is not present in gnomAD. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -5.914 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.095 | Likely Benign | -2.28 | Neutral | 0.425 | Benign | 0.233 | Benign | 2.35 | Pathogenic | 0.00 | Affected | 0.0771 | 0.4518 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||||
| c.1985A>G | Q662R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q662R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that assess sequence conservation, structural impact, or functional effect all converge on a benign outcome: SIFT, PolyPhen‑2 (HumDiv and HumVar), REVEL, PROVEAN, FATHMM, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, premPS, FoldX, and Rosetta all predict benign. No tool in the dataset indicates pathogenicity. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts benign. Thus, based on the available predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.046336 | Structured | 0.103446 | Uncertain | 0.932 | 0.323 | 0.000 | -6.678 | Likely Benign | 0.300 | Likely Benign | Likely Benign | -0.41 | Likely Benign | 0.0 | 0.10 | Likely Benign | -0.16 | Likely Benign | -0.42 | Likely Benign | 0.159 | Likely Benign | 0.49 | Neutral | 0.428 | Benign | 0.095 | Benign | 3.48 | Benign | 1.00 | Tolerated | 0.2023 | 0.1391 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||
| c.3535A>C | K1179Q 2D AIThe SynGAP1 missense variant K1179Q is reported in gnomAD (variant ID 6‑33444570‑A‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions, including the high‑accuracy tools, point to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | 6-33444570-A-C | 1 | 6.20e-7 | -4.237 | Likely Benign | 0.679 | Likely Pathogenic | Likely Benign | 0.078 | Likely Benign | -1.20 | Neutral | 0.430 | Benign | 0.211 | Benign | 2.67 | Benign | 0.00 | Affected | 4.32 | 2 | 0.4037 | 0.0807 | 1 | 1 | 0.4 | -0.04 | |||||||||||||||||||||||||||||||||
| c.1210G>C | A404P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A404P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, Rosetta, both polyPhen‑2 versions, and FATHMM, while pathogenic calls arise from FoldX, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic verdict. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus confirms likely pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, remains uncertain. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion does not conflict with the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.232838 | Structured | 0.415505 | Uncertain | 0.965 | 0.355 | 0.000 | -11.819 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 3.03 | Destabilizing | 0.1 | 0.24 | Likely Benign | 1.64 | Ambiguous | 1.17 | Destabilizing | 0.227 | Likely Benign | -3.16 | Deleterious | 0.433 | Benign | 0.080 | Benign | 4.02 | Benign | 0.03 | Affected | 0.2271 | 0.6366 | 1 | -1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||
| c.2116G>C | E706Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E706Q missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are AlphaMissense‑Default and ESM1b. Rosetta and Foldetta provide uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta also yields an uncertain outcome. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.200174 | Structured | 0.377033 | Uncertain | 0.929 | 0.363 | 0.000 | -8.171 | Likely Pathogenic | 0.588 | Likely Pathogenic | Likely Benign | 0.44 | Likely Benign | 0.0 | 0.61 | Ambiguous | 0.53 | Ambiguous | -0.14 | Likely Benign | 0.059 | Likely Benign | -1.00 | Neutral | 0.433 | Benign | 0.051 | Benign | 4.13 | Benign | 0.31 | Tolerated | 0.0941 | 0.3831 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||
| c.1433A>T | E478V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E478V missense change is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL, Rosetta, polyPhen‑2 (HumDiv and HumVar), FATHMM, and premPS; pathogenic predictions from SGM‑Consensus, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. Two tools give inconclusive results: FoldX (uncertain) and AlphaMissense‑Optimized (uncertain). High‑accuracy assessments further split the signal: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, predicts pathogenic; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts benign. With six benign versus five pathogenic calls and no ClinVar evidence, the overall evidence slightly favors a benign interpretation, and there is no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | -10.322 | Likely Pathogenic | 0.831 | Likely Pathogenic | Ambiguous | 0.55 | Ambiguous | 0.0 | 0.33 | Likely Benign | 0.44 | Likely Benign | 0.04 | Likely Benign | 0.385 | Likely Benign | -5.84 | Deleterious | 0.434 | Benign | 0.199 | Benign | 3.35 | Benign | 0.01 | Affected | 0.0586 | 0.6604 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||
| c.1049T>C | V350A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant V350A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), premPS, PROVEAN, ESM1b, and FATHMM. Stability‑based methods FoldX, Rosetta, and Foldetta give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as pathogenic, and Foldetta as unavailable. Overall, the predictions are split, with a slight edge toward pathogenicity from the consensus and high‑accuracy tools. Therefore, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.179055 | Structured | 0.353227 | Uncertain | 0.931 | 0.371 | 0.000 | -8.323 | Likely Pathogenic | 0.280 | Likely Benign | Likely Benign | 1.42 | Ambiguous | 0.0 | 1.97 | Ambiguous | 1.70 | Ambiguous | 2.07 | Destabilizing | 0.096 | Likely Benign | -2.73 | Deleterious | 0.435 | Benign | 0.115 | Benign | 1.64 | Pathogenic | 0.55 | Tolerated | 0.3207 | 0.2967 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1049T>G | V350G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V350G lies in the C2 domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that classify it as benign include REVEL, polyPhen‑2 HumDiv, and AlphaMissense‑Optimized. The remaining tools—FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—predict it to be pathogenic. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. All predictions are available and consistent. Based on the preponderance of pathogenic calls, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.179055 | Structured | 0.353227 | Uncertain | 0.931 | 0.371 | 0.000 | -12.267 | Likely Pathogenic | 0.597 | Likely Pathogenic | Likely Benign | 2.57 | Destabilizing | 0.0 | 4.25 | Destabilizing | 3.41 | Destabilizing | 1.93 | Destabilizing | 0.330 | Likely Benign | -5.46 | Deleterious | 0.435 | Benign | 0.920 | Probably Damaging | 1.61 | Pathogenic | 0.02 | Affected | 0.2316 | 0.2522 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.1199T>C | V400A 2D 3DClick to see structure in 3D Viewer AISynGAP1 V400A is not reported in ClinVar and is absent from gnomAD. Consensus from standard predictors shows a split: benign calls from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls come from FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT and AlphaMissense‑Default. Two high‑accuracy tools give a clear signal: AlphaMissense‑Optimized is uncertain, but the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta also predicts pathogenic. With most evidence pointing to deleterious effects, the variant is most likely pathogenic, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.398279 | Structured | 0.415488 | Uncertain | 0.951 | 0.451 | 0.000 | -7.564 | In-Between | 0.871 | Likely Pathogenic | Ambiguous | 3.14 | Destabilizing | 0.1 | 3.12 | Destabilizing | 3.13 | Destabilizing | 2.29 | Destabilizing | 0.479 | Likely Benign | -3.12 | Deleterious | 0.435 | Benign | 0.049 | Benign | 5.32 | Benign | 0.01 | Affected | 0.3291 | 0.2767 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||||||||
| c.1175A>G | K392R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). All available in silico predictors classify the change as benign: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign effect. The high‑accuracy folding‑stability tool Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts a benign impact. No tool predicts pathogenicity. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.006 | Likely Benign | 0.091 | Likely Benign | Likely Benign | -0.03 | Likely Benign | 0.0 | 0.44 | Likely Benign | 0.21 | Likely Benign | 0.23 | Likely Benign | 0.131 | Likely Benign | -1.44 | Neutral | 0.436 | Benign | 0.112 | Benign | 7.12 | Benign | 0.08 | Tolerated | 0.5686 | 0.1875 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||
| c.2998A>G | I1000V 2D AIThe SynGAP1 missense variant I1000V is listed in ClinVar (ID 2572013.0) with an “Uncertain” clinical significance and is not reported in gnomAD. Functional prediction tools that assess evolutionary conservation and structural impact (REVEL, SIFT, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, PROVEAN, AlphaMissense‑Default) all converge on a benign outcome. No tool in the dataset predicts pathogenicity. High‑accuracy predictors reinforce this consensus: AlphaMissense‑Optimized reports a benign effect, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict the ClinVar status, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.957020 | Binding | 0.293 | 0.904 | 0.625 | Uncertain | 2 | -4.102 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.086 | Likely Benign | -0.20 | Neutral | 0.437 | Benign | 0.170 | Benign | 2.76 | Benign | 0.81 | Tolerated | 4.32 | 4 | 0.1220 | 0.3404 | 3 | 4 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||
| c.3000C>G | I1000M 2D AIThe SynGAP1 missense variant I1000M is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. Foldetta results are not available, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.957020 | Binding | 0.293 | 0.904 | 0.625 | -3.541 | Likely Benign | 0.161 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -0.32 | Neutral | 0.437 | Benign | 0.108 | Benign | 2.70 | Benign | 0.17 | Tolerated | 0.0769 | 0.3104 | 2 | 1 | -2.6 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3697A>G | I1233V 2D AIThe SynGAP1 missense change I1233V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign effect for I1233V, and this conclusion is consistent with the lack of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -2.826 | Likely Benign | 0.615 | Likely Pathogenic | Likely Benign | 0.036 | Likely Benign | -0.59 | Neutral | 0.437 | Benign | 0.170 | Benign | 2.79 | Benign | 0.06 | Tolerated | 0.0950 | 0.3270 | 4 | 3 | -0.3 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3699C>G | I1233M 2D AIThe SynGAP1 missense variant I1233M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic effect, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments further support this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also indicates likely benign; Foldetta results are not available. Overall, the consensus of the available predictions points to a benign impact for I1233M, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Thus, the variant is most likely benign, and this assessment does not contradict ClinVar, which contains no pathogenic claim. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -4.360 | Likely Benign | 0.449 | Ambiguous | Likely Benign | 0.069 | Likely Benign | -0.70 | Neutral | 0.437 | Benign | 0.108 | Benign | 2.70 | Benign | 0.04 | Affected | 0.0562 | 0.2726 | 2 | 1 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3787A>G | I1263V 2D AIThe SynGAP1 missense variant I1263V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two benign vs. two pathogenic votes), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -4.230 | Likely Benign | 0.729 | Likely Pathogenic | Likely Benign | 0.221 | Likely Benign | -0.83 | Neutral | 0.437 | Benign | 0.170 | Benign | 1.99 | Pathogenic | 0.00 | Affected | 0.1136 | 0.3102 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.559C>G | L187V 2D AIThe SynGAP1 missense variant L187V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign). High‑accuracy AlphaMissense‑Optimized predicts pathogenic, while Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the majority of standard predictors lean toward a benign classification, but the single high‑accuracy tool indicates pathogenicity. Thus, the variant is most likely benign based on the collective evidence, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.465241 | Structured | 0.428046 | Uncertain | 0.367 | 0.625 | 0.375 | -10.543 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.136 | Likely Benign | -2.24 | Neutral | 0.437 | Benign | 0.079 | Benign | 3.81 | Benign | 0.02 | Affected | 0.1525 | 0.3509 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.587T>C | L196S 2D  AIThe SynGAP1 missense variant L196S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic, SGM‑Consensus is Likely Pathogenic, while Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.284882 | Structured | 0.429452 | Uncertain | 0.489 | 0.521 | 0.125 | -9.987 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.353 | Likely Benign | -4.75 | Deleterious | 0.437 | Benign | 0.186 | Benign | 3.65 | Benign | 0.00 | Affected | 0.3473 | 0.0736 | -3 | -2 | -4.6 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.677C>T | S226L 2D AIThe SynGAP1 missense variant S226L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, PROVEAN, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign; the SGM‑Consensus (majority vote) also indicates benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts benign. No prediction or folding result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.129801 | Structured | 0.334959 | Uncertain | 0.800 | 0.324 | 0.250 | -6.098 | Likely Benign | 0.239 | Likely Benign | Likely Benign | -0.12 | Likely Benign | 0.5 | -0.14 | Likely Benign | -0.13 | Likely Benign | 0.36 | Likely Benign | 0.578 | Likely Pathogenic | -3.87 | Deleterious | 0.437 | Benign | 0.048 | Benign | 5.73 | Benign | 0.03 | Affected | 0.1525 | 0.5907 | -3 | -2 | 4.6 | 26.08 | |||||||||||||||||||||||||||||
| c.692T>A | F231Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F231Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on benign effects include Rosetta, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict pathogenicity are REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; FoldX and premPS are inconclusive. High‑accuracy methods give AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of standard tools lean benign, but the two most accurate predictors and the consensus vote favor pathogenicity. Thus, the variant is most likely pathogenic, and this assessment does not contradict ClinVar status, which currently has no classification for F231Y. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.366687 | Structured | 0.306467 | Uncertain | 0.895 | 0.300 | 0.000 | -9.831 | Likely Pathogenic | 0.980 | Likely Pathogenic | Likely Pathogenic | 0.69 | Ambiguous | 0.1 | 0.00 | Likely Benign | 0.35 | Likely Benign | 0.85 | Ambiguous | 0.687 | Likely Pathogenic | -2.60 | Deleterious | 0.437 | Benign | 0.079 | Benign | 5.50 | Benign | 0.12 | Tolerated | 0.1355 | 0.2732 | 7 | 3 | -4.1 | 16.00 | |||||||||||||||||||||||||||||
| c.2027G>A | S676N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S676N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote). Only SIFT predicts a pathogenic outcome. Predictions that are inconclusive (FoldX, premPS, ESM1b) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates benign. Foldetta, which integrates FoldX‑MD and Rosetta outputs, is unavailable because one component (FoldX) is uncertain. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.209395 | Structured | 0.113632 | Uncertain | 0.551 | 0.338 | 0.125 | -7.486 | In-Between | 0.282 | Likely Benign | Likely Benign | 0.60 | Ambiguous | 0.3 | -0.21 | Likely Benign | 0.20 | Likely Benign | 0.55 | Ambiguous | 0.099 | Likely Benign | -1.63 | Neutral | 0.438 | Benign | 0.036 | Benign | 3.42 | Benign | 0.05 | Affected | 0.1454 | 0.4599 | 1 | 1 | -2.7 | 27.03 | |||||||||||||||||||||||||||||
| c.2593G>A | A865T 2D AIThe SynGAP1 missense variant A865T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of all available predictions is that the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.521092 | Disordered | 0.626222 | Binding | 0.271 | 0.788 | 0.250 | -4.435 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.026 | Likely Benign | -0.84 | Neutral | 0.440 | Benign | 0.197 | Benign | 2.72 | Benign | 0.29 | Tolerated | 0.1263 | 0.6083 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.2903G>A | G968D 2D AIThe SynGAP1 missense variant G968D is catalogued in gnomAD (ID 6‑33443455‑G‑A) but has no ClinVar entry. Across a broad panel of in‑silico predictors, every tool reports a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity, so the pathogenic‑prediction group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available for this variant. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.961360 | Binding | 0.327 | 0.896 | 0.750 | 6-33443455-G-A | 1 | 6.20e-7 | -5.134 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.157 | Likely Benign | -0.48 | Neutral | 0.440 | Benign | 0.198 | Benign | 4.19 | Benign | 0.21 | Tolerated | 4.32 | 2 | 0.1965 | 0.3243 | -1 | 1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||
| c.2972G>T | G991V 2D AIThe SynGAP1 missense variant G991V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.911393 | Binding | 0.286 | 0.920 | 0.750 | -4.129 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -1.61 | Neutral | 0.440 | Benign | 0.253 | Benign | 4.16 | Benign | 0.01 | Affected | 0.1148 | 0.3961 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3098C>T | S1033F 2D AIThe SynGAP1 missense variant S1033F is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus is Likely Benign, and Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.993473 | Binding | 0.294 | 0.737 | 0.625 | -5.013 | Likely Benign | 0.555 | Ambiguous | Likely Benign | 0.022 | Likely Benign | -1.02 | Neutral | 0.440 | Benign | 0.185 | Benign | 2.70 | Benign | 0.05 | Affected | 0.0699 | 0.5546 | -3 | -2 | 3.6 | 60.10 | |||||||||||||||||||||||||||||||||||||||
| c.3241G>A | A1081T 2D AIThe SynGAP1 missense variant A1081T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a benign effect. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the evidence strongly supports a benign classification, and there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.874069 | Disordered | 0.979759 | Binding | 0.288 | 0.895 | 0.750 | -3.887 | Likely Benign | 0.129 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -1.07 | Neutral | 0.440 | Benign | 0.184 | Benign | 4.01 | Benign | 0.15 | Tolerated | 0.1567 | 0.6112 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3335A>T | E1112V 2D AIThe SynGAP1 missense variant E1112V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote) also as benign; Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a benign impact for E1112V, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.909381 | Binding | 0.335 | 0.902 | 0.875 | -3.971 | Likely Benign | 0.579 | Likely Pathogenic | Likely Benign | 0.139 | Likely Benign | -2.28 | Neutral | 0.440 | Benign | 0.140 | Benign | 2.70 | Benign | 0.00 | Affected | 0.1481 | 0.7567 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.292C>T | H98Y 2D AIThe SynGAP1 missense variant H98Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.631713 | Binding | 0.348 | 0.872 | 0.625 | -4.050 | Likely Benign | 0.135 | Likely Benign | Likely Benign | 0.112 | Likely Benign | -0.86 | Neutral | 0.444 | Benign | 0.024 | Benign | 4.19 | Benign | 0.00 | Affected | 0.0950 | 0.5024 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||||||
| c.4027C>A | H1343N 2D AIThe SynGAP1 missense variant H1343N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -3.077 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -1.09 | Neutral | 0.444 | Benign | 0.071 | Benign | 4.06 | Benign | 0.00 | Affected | 0.2398 | 0.3299 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.4027C>G | H1343D 2D AIThe SynGAP1 missense variant H1343D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only SIFT predicts a pathogenic impact. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. AlphaMissense‑Optimized also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -3.136 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.051 | Likely Benign | -1.29 | Neutral | 0.444 | Benign | 0.071 | Benign | 4.07 | Benign | 0.00 | Affected | 0.3011 | 0.2403 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.4027C>T | H1343Y 2D AIThe SynGAP1 missense variant H1343Y is reported in gnomAD (ID 6‑33451901‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact for H1343Y, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | 6-33451901-C-T | -3.479 | Likely Benign | 0.139 | Likely Benign | Likely Benign | 0.040 | Likely Benign | -0.85 | Neutral | 0.444 | Benign | 0.071 | Benign | 4.06 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1030 | 0.4792 | 2 | 0 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||||||||
| c.1927G>C | E643Q 2D AIThe SynGAP1 missense variant E643Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SGM‑Consensus (Likely Pathogenic), PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized predicts a benign outcome, while premPS is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions (8 benign vs. 5 pathogenic) and the two of three high‑accuracy tools favor a benign classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.033407 | Structured | 0.215915 | Uncertain | 0.871 | 0.315 | 0.000 | -12.404 | Likely Pathogenic | 0.688 | Likely Pathogenic | Likely Benign | 0.49 | Likely Benign | 0.6 | 0.15 | Likely Benign | 0.32 | Likely Benign | 0.83 | Ambiguous | 0.341 | Likely Benign | -2.86 | Deleterious | 0.446 | Benign | 0.038 | Benign | 2.94 | Benign | 0.01 | Affected | 0.1603 | 0.6308 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||
| c.2296T>G | S766A 2D AIThe SynGAP1 missense variant S766A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. The variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.405110 | Structured | 0.923125 | Binding | 0.338 | 0.874 | 0.250 | -6.115 | Likely Benign | 0.186 | Likely Benign | Likely Benign | 0.055 | Likely Benign | -0.98 | Neutral | 0.447 | Benign | 0.198 | Benign | 4.17 | Benign | 0.02 | Affected | 0.5059 | 0.5705 | Strenghten | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||
| c.3217T>G | S1073A 2D AIThe SynGAP1 missense variant S1073A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) confirms a benign status. Foldetta’s protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and high‑accuracy tools suggests that the variant is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.916840 | Disordered | 0.985818 | Binding | 0.313 | 0.905 | 0.750 | -5.333 | Likely Benign | 0.220 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -0.96 | Neutral | 0.447 | Benign | 0.103 | Benign | 3.95 | Benign | 0.02 | Affected | 0.4571 | 0.5883 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.3259T>G | S1087A 2D AIThe SynGAP1 missense variant S1087A is not reported in ClinVar and is absent from gnomAD, indicating no documented clinical or population data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Based on the consensus of all available predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.974805 | Binding | 0.357 | 0.891 | 1.000 | -3.649 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.057 | Likely Benign | -1.03 | Neutral | 0.447 | Benign | 0.139 | Benign | 2.64 | Benign | 0.34 | Tolerated | 0.4464 | 0.5405 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.3280T>G | S1094A 2D AIThe SynGAP1 missense variant S1094A is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity, so the pathogenic‑prediction group is empty. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta results are not available, so they do not influence the assessment. Overall, the variant is most likely benign, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.938133 | Disordered | 0.981352 | Binding | 0.358 | 0.877 | 1.000 | -3.595 | Likely Benign | 0.188 | Likely Benign | Likely Benign | 0.117 | Likely Benign | -0.78 | Neutral | 0.447 | Benign | 0.252 | Benign | 2.66 | Benign | 1.00 | Tolerated | 0.4692 | 0.5403 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.637A>C | I213L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I213L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM all classify the variant as benign, while AlphaMissense‑Default predicts it as pathogenic and ESM1b remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores the variant benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign outcome; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, also reports a benign effect. No prediction or stability result is missing or inconclusive. Overall, the variant is most likely benign based on the collective evidence, and this benign prediction does not contradict the ClinVar status, which simply lacks an entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.158265 | Structured | 0.372201 | Uncertain | 0.850 | 0.295 | 0.125 | -7.673 | In-Between | 0.577 | Likely Pathogenic | Likely Benign | 0.29 | Likely Benign | 0.4 | 0.20 | Likely Benign | 0.25 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -1.45 | Neutral | 0.447 | Benign | 0.177 | Benign | 5.89 | Benign | 0.18 | Tolerated | 0.0771 | 0.2857 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.2095G>C | V699L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V699L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while FoldX, premPS, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority; and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the preponderance of evidence, and this assessment does not contradict any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.432975 | Uncertain | 0.935 | 0.315 | 0.000 | -8.301 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | -0.58 | Ambiguous | 0.1 | -0.36 | Likely Benign | -0.47 | Likely Benign | 0.69 | Ambiguous | 0.150 | Likely Benign | -2.14 | Neutral | 0.448 | Benign | 0.153 | Benign | 3.48 | Benign | 0.10 | Tolerated | 0.0900 | 0.3289 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.2095G>T | V699L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V699L has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Only ESM1b predicts a pathogenic outcome, while FoldX, premPS, and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign majority; and Foldetta also predicts benign stability. No prediction or folding result is missing or inconclusive. Overall, the variant is most likely benign based on the consensus of the available tools, and this benign prediction does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.069024 | Structured | 0.432975 | Uncertain | 0.935 | 0.315 | 0.000 | -8.301 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | -0.58 | Ambiguous | 0.1 | -0.36 | Likely Benign | -0.47 | Likely Benign | 0.69 | Ambiguous | 0.150 | Likely Benign | -2.14 | Neutral | 0.448 | Benign | 0.153 | Benign | 3.48 | Benign | 0.10 | Tolerated | 0.0900 | 0.3289 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||
| c.2573G>A | S858N 2D AIThe SynGAP1 missense variant S858N is listed in ClinVar with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443125‑G‑A). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate benign. In contrast, polyPhen‑2 HumVar and SIFT predict pathogenicity, but these two tools are in the minority. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.724957 | Disordered | 0.482724 | Uncertain | 0.305 | 0.833 | 0.375 | Uncertain | 1 | 6-33443125-G-A | 2 | 1.24e-6 | -4.311 | Likely Benign | 0.121 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -0.67 | Neutral | 0.448 | Benign | 0.846 | Possibly Damaging | 4.13 | Benign | 0.02 | Affected | 3.77 | 5 | 0.1527 | 0.4772 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||
| c.1093G>A | V365I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V365I is listed in ClinVar with an “Uncertain” status and is present in gnomAD (variant ID 6‑33437998‑G‑A). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of evidence points to a benign effect, and this consensus does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.441505 | Uncertain | 0.923 | 0.608 | 0.250 | Uncertain | 1 | 6-33437998-G-A | 4 | 2.65e-6 | -5.943 | Likely Benign | 0.155 | Likely Benign | Likely Benign | -0.65 | Ambiguous | 0.2 | -0.11 | Likely Benign | -0.38 | Likely Benign | 0.13 | Likely Benign | 0.036 | Likely Benign | -0.47 | Neutral | 0.451 | Benign | 0.137 | Benign | 1.76 | Pathogenic | 0.10 | Tolerated | 3.38 | 21 | 0.0657 | 0.4447 | 3 | 4 | 0.3 | 14.03 | ||||||||||||||||||||||
| c.2371A>G | K791E 2D AIThe SynGAP1 missense variant K791E is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools uniformly favor a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) indicates likely benign; Foldetta results are unavailable. Taken together, the preponderance of evidence supports a benign impact for K791E, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.966441 | Disordered | 0.478670 | Uncertain | 0.356 | 0.896 | 0.875 | -3.823 | Likely Benign | 0.465 | Ambiguous | Likely Benign | 0.053 | Likely Benign | -0.58 | Neutral | 0.451 | Benign | 0.193 | Benign | 4.22 | Benign | 0.65 | Tolerated | 0.4829 | 0.0876 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.2399G>A | G800D 2D AIThe SynGAP1 missense variant G800D is catalogued in gnomAD (6-33442951‑G‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign or likely benign. No tool predicts pathogenicity. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized reports a benign outcome, and the SGM‑Consensus (derived from the majority of the four high‑accuracy predictors) also yields a benign verdict. Foldetta results are unavailable, so they do not influence the assessment. Overall, the evidence strongly supports a benign classification, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.852992 | Disordered | 0.588350 | Binding | 0.303 | 0.884 | 0.625 | 6-33442951-G-A | -5.929 | Likely Benign | 0.400 | Ambiguous | Likely Benign | 0.088 | Likely Benign | -0.84 | Neutral | 0.451 | Benign | 0.265 | Benign | 2.76 | Benign | 0.34 | Tolerated | 4.32 | 2 | 0.1664 | 0.1877 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||
| c.2399G>T | G800V 2D AIThe SynGAP1 missense variant G800V is predicted to be benign by all evaluated in‑silico tools. ClinVar contains no entry for this variant, and it is not reported in gnomAD. Consensus predictions from SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a benign effect. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a likely benign outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the collective evidence, the variant is most likely benign, and this conclusion is consistent with the absence of a pathogenic ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.852992 | Disordered | 0.588350 | Binding | 0.303 | 0.884 | 0.625 | -4.890 | Likely Benign | 0.113 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -1.24 | Neutral | 0.451 | Benign | 0.157 | Benign | 2.73 | Benign | 0.25 | Tolerated | 0.1130 | 0.4198 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||
| c.2647C>G | L883V 2D AIThe SynGAP1 missense variant L883V is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is consistent with the lack of a ClinVar classification—there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.716283 | Disordered | 0.641952 | Binding | 0.334 | 0.886 | 0.250 | -4.075 | Likely Benign | 0.079 | Likely Benign | Likely Benign | 0.059 | Likely Benign | -0.28 | Neutral | 0.451 | Benign | 0.157 | Benign | 2.67 | Benign | 0.28 | Tolerated | 0.1611 | 0.3744 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2677C>A | Q893K 2D AIThe SynGAP1 missense variant Q893K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the substitution as benign, and AlphaMissense‑Optimized also predicts a benign outcome. No tool predicts pathogenicity; AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the computational evidence strongly supports a benign classification, and this conclusion does not contradict ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.447267 | Uncertain | 0.310 | 0.925 | 0.750 | -5.622 | Likely Benign | 0.496 | Ambiguous | Likely Benign | 0.053 | Likely Benign | -1.45 | Neutral | 0.451 | Benign | 0.265 | Benign | 2.78 | Benign | 0.10 | Tolerated | 0.1901 | 0.4381 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.2678A>T | Q893L 2D AIThe SynGAP1 missense variant Q893L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of all available predictions is benign, and this conclusion does not contradict any ClinVar annotation (none present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.447267 | Uncertain | 0.310 | 0.925 | 0.750 | -1.964 | Likely Benign | 0.204 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -1.92 | Neutral | 0.451 | Benign | 0.209 | Benign | 2.82 | Benign | 1.00 | Tolerated | 0.0904 | 0.5643 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.2731G>A | V911I 2D AIThe SynGAP1 missense variant V911I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign,” while Foldetta results are unavailable. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.724137 | Binding | 0.327 | 0.914 | 0.375 | -3.983 | Likely Benign | 0.085 | Likely Benign | Likely Benign | 0.024 | Likely Benign | -0.32 | Neutral | 0.451 | Benign | 0.209 | Benign | 2.70 | Benign | 0.22 | Tolerated | 0.0881 | 0.4331 | 4 | 3 | 0.3 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2731G>C | V911L 2D AIThe SynGAP1 missense variant V911L is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity all converge on a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. No tool in the dataset indicates a pathogenic effect. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports a benign effect, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is classified as Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence strongly supports a benign classification, and this is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.724137 | Binding | 0.327 | 0.914 | 0.375 | -2.722 | Likely Benign | 0.166 | Likely Benign | Likely Benign | 0.100 | Likely Benign | -0.39 | Neutral | 0.451 | Benign | 0.157 | Benign | 2.73 | Benign | 0.16 | Tolerated | 0.1067 | 0.5310 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2863T>A | S955T 2D AIThe SynGAP1 missense variant S955T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for S955T, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984871 | Disordered | 0.945325 | Binding | 0.350 | 0.924 | 0.750 | -4.717 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.070 | Likely Benign | -0.96 | Neutral | 0.451 | Benign | 0.265 | Benign | 2.38 | Pathogenic | 0.00 | Affected | 0.2184 | 0.5927 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2926T>G | F976V 2D AIThe SynGAP1 missense variant F976V is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the collective evidence strongly supports a benign classification, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.975061 | Binding | 0.311 | 0.894 | 0.625 | -3.328 | Likely Benign | 0.268 | Likely Benign | Likely Benign | 0.190 | Likely Benign | -1.32 | Neutral | 0.451 | Benign | 0.157 | Benign | 4.18 | Benign | 0.23 | Tolerated | 0.2696 | 0.2902 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.2936T>A | F979Y 2D AIThe SynGAP1 missense variant F979Y is not reported in ClinVar and is absent from gnomAD. All available in‑silico predictors classify the change as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence strongly supports a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.977500 | Binding | 0.274 | 0.889 | 0.625 | -3.420 | Likely Benign | 0.277 | Likely Benign | Likely Benign | 0.123 | Likely Benign | -0.26 | Neutral | 0.451 | Benign | 0.285 | Benign | 4.18 | Benign | 0.06 | Tolerated | 0.1451 | 0.2843 | 7 | 3 | -4.1 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.2936T>C | F979S 2D AIThe SynGAP1 missense variant F979S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and the AlphaMissense‑Optimized score also indicates a benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists for F979S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.977500 | Binding | 0.274 | 0.889 | 0.625 | -2.350 | Likely Benign | 0.718 | Likely Pathogenic | Likely Benign | 0.211 | Likely Benign | -0.05 | Neutral | 0.451 | Benign | 0.220 | Benign | 4.23 | Benign | 0.01 | Affected | 0.4397 | 0.0384 | -3 | -2 | -3.6 | -60.10 | |||||||||||||||||||||||||||||||||||||||
| c.2938C>A | H980N 2D AIThe SynGAP1 missense variant H980N is not reported in ClinVar or gnomAD. Functional prediction tools largely agree on a benign effect. Benign calls come from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus confirms a benign result; Foldetta data are unavailable, so no additional stability evidence is considered. Overall, the computational evidence indicates that H980N is most likely benign, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.974598 | Binding | 0.309 | 0.892 | 0.625 | -4.728 | Likely Benign | 0.291 | Likely Benign | Likely Benign | 0.070 | Likely Benign | -1.07 | Neutral | 0.451 | Benign | 0.209 | Benign | 4.17 | Benign | 0.00 | Affected | 0.2346 | 0.3638 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.2938C>G | H980D 2D AIThe SynGAP1 missense variant H980D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.974598 | Binding | 0.309 | 0.892 | 0.625 | -5.489 | Likely Benign | 0.729 | Likely Pathogenic | Likely Benign | 0.117 | Likely Benign | -1.38 | Neutral | 0.451 | Benign | 0.265 | Benign | 4.18 | Benign | 0.00 | Affected | 0.2608 | 0.3066 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.3166G>A | G1056S 2D AIThe SynGAP1 missense variant G1056S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.868632 | Binding | 0.402 | 0.935 | 0.875 | -5.252 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.339 | Likely Benign | -0.28 | Neutral | 0.451 | Benign | 0.149 | Benign | 1.87 | Pathogenic | 0.55 | Tolerated | 0.2497 | 0.5702 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3170G>A | S1057N 2D AIThe SynGAP1 missense variant S1057N is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. All available in‑silico predictors classify the change as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” No tool predicts pathogenicity. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.869507 | Binding | 0.413 | 0.927 | 0.875 | Uncertain | 1 | -6.386 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.218 | Likely Benign | -0.41 | Neutral | 0.451 | Benign | 0.129 | Benign | 5.25 | Benign | 0.28 | Tolerated | 0.2232 | 0.4605 | 1 | 1 | -2.7 | 27.03 | |||||||||||||||||||||||||||||||||||||
| c.3202T>G | L1068V 2D AIThe SynGAP1 missense variant L1068V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.947281 | Disordered | 0.981041 | Binding | 0.362 | 0.907 | 0.875 | -5.325 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.051 | Likely Benign | -0.58 | Neutral | 0.451 | Benign | 0.110 | Benign | 2.54 | Benign | 0.00 | Affected | 0.1619 | 0.4404 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3205C>G | Q1069E 2D AIThe SynGAP1 missense variant Q1069E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of all available predictions is that the variant is most likely benign, and this conclusion is consistent with the lack of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.943310 | Disordered | 0.981477 | Binding | 0.333 | 0.906 | 0.875 | -5.165 | Likely Benign | 0.322 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -0.68 | Neutral | 0.451 | Benign | 0.266 | Benign | 2.76 | Benign | 0.33 | Tolerated | 0.1460 | 0.3127 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3232G>C | V1078L 2D AIThe SynGAP1 missense variant V1078L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective evidence strongly suggests that V1078L is most likely benign, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.986989 | Binding | 0.294 | 0.898 | 0.750 | -2.547 | Likely Benign | 0.523 | Ambiguous | Likely Benign | 0.091 | Likely Benign | -0.16 | Neutral | 0.451 | Benign | 0.209 | Benign | 4.13 | Benign | 0.56 | Tolerated | 0.0889 | 0.5349 | 2 | 1 | -0.4 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3331A>G | K1111E 2D AIThe SynGAP1 missense variant K1111E is not reported in ClinVar and has no entry in gnomAD. Consensus from multiple in‑silico predictors indicates a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score it as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also classifies it as likely benign. Only AlphaMissense‑Default predicts a pathogenic outcome. High‑accuracy tools further support a benign interpretation: AlphaMissense‑Optimized returns a benign prediction, while the SGM‑Consensus (majority vote) remains benign; a Foldetta stability assessment is unavailable. Taken together, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.921455 | Binding | 0.300 | 0.902 | 0.875 | -3.666 | Likely Benign | 0.565 | Likely Pathogenic | Likely Benign | 0.089 | Likely Benign | -0.86 | Neutral | 0.451 | Benign | 0.193 | Benign | 2.69 | Benign | 0.23 | Tolerated | 0.3846 | 0.1833 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3358G>A | G1120S 2D AIThe SynGAP1 missense variant G1120S is not reported in ClinVar and is absent from gnomAD, indicating no documented allele frequency data. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this benign prediction: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of all available predictions strongly suggests that G1120S is most likely benign, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.871313 | Disordered | 0.804931 | Binding | 0.335 | 0.925 | 0.875 | -4.959 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.286 | Likely Benign | -0.27 | Neutral | 0.451 | Benign | 0.209 | Benign | 3.69 | Benign | 0.77 | Tolerated | 0.2515 | 0.5311 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3515A>T | H1172L 2D AIThe SynGAP1 missense variant H1172L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for H1172L, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.673805 | Binding | 0.465 | 0.758 | 0.625 | -0.545 | Likely Benign | 0.446 | Ambiguous | Likely Benign | 0.426 | Likely Benign | -2.30 | Neutral | 0.451 | Benign | 0.265 | Benign | 5.47 | Benign | 0.01 | Affected | 0.0815 | 0.5285 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||
| c.3516C>A | H1172Q 2D AIThe SynGAP1 missense variant H1172Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus confirms Likely Benign, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.673805 | Binding | 0.465 | 0.758 | 0.625 | -2.169 | Likely Benign | 0.414 | Ambiguous | Likely Benign | 0.227 | Likely Benign | -0.51 | Neutral | 0.451 | Benign | 0.265 | Benign | 5.47 | Benign | 0.39 | Tolerated | 0.1249 | 0.3557 | 3 | 0 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3516C>G | H1172Q 2D AIThe SynGAP1 missense variant H1172Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus confirms Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Based on the collective predictions, H1172Q is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.673805 | Binding | 0.465 | 0.758 | 0.625 | -2.169 | Likely Benign | 0.414 | Ambiguous | Likely Benign | 0.227 | Likely Benign | -0.51 | Neutral | 0.451 | Benign | 0.265 | Benign | 5.47 | Benign | 0.39 | Tolerated | 0.1249 | 0.3557 | 3 | 0 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3518T>C | I1173T 2D AIThe SynGAP1 missense variant I1173T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for I1173T, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.501700 | Disordered | 0.653145 | Binding | 0.521 | 0.756 | 0.375 | -3.162 | Likely Benign | 0.441 | Ambiguous | Likely Benign | 0.405 | Likely Benign | -1.18 | Neutral | 0.451 | Benign | 0.265 | Benign | 5.46 | Benign | 0.05 | Affected | 0.1016 | 0.0821 | 0 | -1 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||||||
| c.3847C>A | P1283T 2D AIThe SynGAP1 missense variant P1283T is catalogued in gnomAD (ID 6‑33447895‑C‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Therefore, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.819686 | Binding | 0.484 | 0.732 | 0.875 | 6-33447895-C-A | -4.781 | Likely Benign | 0.064 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -0.45 | Neutral | 0.451 | Benign | 0.193 | Benign | 2.76 | Benign | 0.11 | Tolerated | 3.77 | 5 | 0.1269 | 0.3784 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3853C>A | P1285T 2D AIThe SynGAP1 missense variant P1285T is catalogued in gnomAD (ID 6‑33447901‑C‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as tolerated or benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a Likely Benign verdict. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Overall, the computational evidence overwhelmingly supports a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.821643 | Binding | 0.557 | 0.759 | 0.750 | 6-33447901-C-A | -4.230 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.047 | Likely Benign | -0.35 | Neutral | 0.451 | Benign | 0.193 | Benign | 4.33 | Benign | 0.40 | Tolerated | 4.32 | 2 | 0.1496 | 0.3556 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3870G>C | R1290S 2D AIThe SynGAP1 missense variant R1290S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM, while pathogenic predictions come from PROVEAN and SIFT. The high‑accuracy consensus (SGM Consensus) – derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN – favors a benign outcome (2 benign vs. 1 pathogenic, with one uncertain). AlphaMissense‑Optimized also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence indicates a benign effect, and this assessment does not contradict any ClinVar annotation, as none exists for R1290S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.844138 | Binding | 0.567 | 0.795 | 0.625 | -3.507 | Likely Benign | 0.467 | Ambiguous | Likely Benign | 0.120 | Likely Benign | -2.99 | Deleterious | 0.451 | Benign | 0.209 | Benign | 2.65 | Benign | 0.01 | Affected | 0.2655 | 0.2733 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.3870G>T | R1290S 2D AIThe SynGAP1 missense variant R1290S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is benign (2 benign vs. 1 pathogenic, 1 uncertain). High‑accuracy predictions show AlphaMissense‑Optimized as benign and the SGM Consensus as benign; Foldetta results are unavailable. Overall, the consensus of available evidence indicates that R1290S is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.844138 | Binding | 0.567 | 0.795 | 0.625 | -3.507 | Likely Benign | 0.467 | Ambiguous | Likely Benign | 0.120 | Likely Benign | -2.99 | Deleterious | 0.451 | Benign | 0.209 | Benign | 2.65 | Benign | 0.01 | Affected | 0.2655 | 0.2733 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.2870A>C | H957P 2D  AIThe SynGAP1 H957P missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. The high‑accuracy consensus (SGM‑Consensus) is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yielding a Likely Benign classification (3 benign vs. 1 pathogenic). AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.968874 | Binding | 0.362 | 0.915 | 0.750 | -6.347 | Likely Benign | 0.061 | Likely Benign | Likely Benign | 0.244 | Likely Benign | -0.61 | Neutral | 0.453 | Possibly Damaging | 0.105 | Benign | 2.42 | Pathogenic | 0.13 | Tolerated | 0.2300 | 0.4701 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2876A>C | H959P 2D AIThe SynGAP1 missense variant H959P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.980566 | Binding | 0.333 | 0.905 | 0.750 | -8.902 | Likely Pathogenic | 0.070 | Likely Benign | Likely Benign | 0.232 | Likely Benign | -0.61 | Neutral | 0.453 | Possibly Damaging | 0.105 | Benign | 4.14 | Benign | 0.38 | Tolerated | 0.2284 | 0.4901 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2912C>G | P971R 2D AIThe SynGAP1 missense variant P971R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.950334 | Disordered | 0.951523 | Binding | 0.545 | 0.905 | 0.625 | -4.407 | Likely Benign | 0.149 | Likely Benign | Likely Benign | 0.042 | Likely Benign | -1.01 | Neutral | 0.453 | Possibly Damaging | 0.078 | Benign | 3.91 | Benign | 0.00 | Affected | 0.1306 | 0.3818 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3037T>C | S1013P 2D AIThe SynGAP1 missense variant S1013P is reported in gnomAD (ID 6‑33443589‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the consensus score from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.899570 | Binding | 0.308 | 0.846 | 0.625 | 6-33443589-T-C | 2 | 1.24e-6 | -2.563 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 0.103 | Likely Benign | -1.27 | Neutral | 0.453 | Possibly Damaging | 0.150 | Benign | 2.66 | Benign | 0.15 | Tolerated | 3.77 | 5 | 0.2338 | 0.5430 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.3038C>T | S1013F 2D AIThe SynGAP1 missense variant S1013F is catalogued in gnomAD (ID 6‑33443590‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen2_HumVar, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen2_HumDiv, and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign; the SGM Consensus, derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive due to mixed signals, and Foldetta results are unavailable. Overall, the balance of evidence, including the benign call from AlphaMissense‑Optimized, points to a likely benign effect. This conclusion does not conflict with ClinVar, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.899570 | Binding | 0.308 | 0.846 | 0.625 | 6-33443590-C-T | 1 | 6.20e-7 | -5.370 | Likely Benign | 0.353 | Ambiguous | Likely Benign | 0.057 | Likely Benign | -2.54 | Deleterious | 0.453 | Possibly Damaging | 0.272 | Benign | 2.65 | Benign | 0.03 | Affected | 3.77 | 5 | 0.0922 | 0.5803 | -2 | -3 | 3.6 | 60.10 | |||||||||||||||||||||||||||||||||||
| c.3398T>A | I1133N 2D  AIThe SynGAP1 missense variant I1133N is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Pathogenicity is suggested only by polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a benign effect for I1133N, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.832785 | Binding | 0.316 | 0.892 | 0.750 | -5.887 | Likely Benign | 0.520 | Ambiguous | Likely Benign | 0.290 | Likely Benign | -1.07 | Neutral | 0.453 | Possibly Damaging | 0.162 | Benign | 5.51 | Benign | 0.02 | Affected | 0.0961 | 0.1140 | -2 | -3 | -8.0 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3401C>T | T1134I 2D AIThe SynGAP1 missense variant T1134I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also reports a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.885302 | Disordered | 0.813034 | Binding | 0.335 | 0.885 | 0.875 | -4.072 | Likely Benign | 0.416 | Ambiguous | Likely Benign | 0.242 | Likely Benign | -1.93 | Neutral | 0.453 | Possibly Damaging | 0.162 | Benign | 5.52 | Benign | 0.44 | Tolerated | 0.1071 | 0.5447 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||||||||||||
| c.1018G>A | A340T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A340T is reported in gnomAD (ID 6‑33437923‑G‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus all classify the change as benign or likely benign. Only two tools predict pathogenicity—polyPhen‑2 HumDiv and FATHMM—while stability‑based methods (FoldX, Rosetta, premPS, Foldetta) return uncertain or inconclusive results. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely benign, and Foldetta provides no definitive stability change. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.390993 | Structured | 0.410781 | Uncertain | 0.558 | 0.485 | 0.250 | 6-33437923-G-A | -3.286 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.84 | Ambiguous | 0.2 | 0.96 | Ambiguous | 0.90 | Ambiguous | -0.54 | Ambiguous | 0.105 | Likely Benign | 0.62 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 1.93 | Pathogenic | 0.47 | Tolerated | 3.42 | 13 | 0.1740 | 0.7297 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||||
| c.1021G>A | G341S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G341S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, polyPhen‑2 HumVar, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. Predictions that are inconclusive or uncertain are Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM‑Consensus as Likely Benign, and Foldetta as Uncertain. Overall, the majority of evidence points to a benign impact for G341S, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.356642 | Structured | 0.431164 | Uncertain | 0.745 | 0.479 | 0.250 | -3.206 | Likely Benign | 0.090 | Likely Benign | Likely Benign | 0.41 | Likely Benign | 0.3 | -1.46 | Ambiguous | -0.53 | Ambiguous | -0.67 | Ambiguous | 0.343 | Likely Benign | 0.73 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 0.37 | Pathogenic | 0.72 | Tolerated | 0.2416 | 0.4215 | 1 | 0 | -0.4 | 30.03 | |||||||||||||||||||||||||||||
| c.1022G>A | G341D 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G341D is not reported in ClinVar (ClinVar ID: None) but is present in gnomAD (ID: 6-33437927‑G‑A). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, FATHMM, and AlphaMissense‑Default. The remaining tools—Foldetta, AlphaMissense‑Optimized, ESM1b, and Rosetta—return uncertain or inconclusive results and are treated as unavailable for pathogenicity inference. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Considering the majority of standard tools lean benign but the high‑accuracy consensus indicates pathogenicity, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.356642 | Structured | 0.431164 | Uncertain | 0.745 | 0.479 | 0.250 | 6-33437927-G-A | 6 | 3.72e-6 | -7.402 | In-Between | 0.871 | Likely Pathogenic | Ambiguous | 0.28 | Likely Benign | 0.1 | -1.32 | Ambiguous | -0.52 | Ambiguous | -0.04 | Likely Benign | 0.295 | Likely Benign | -0.11 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 0.34 | Pathogenic | 0.25 | Tolerated | 3.42 | 13 | 0.1727 | 0.2241 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||
| c.1097C>A | T366N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense change T366N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) all indicate a benign or likely benign outcome. Only two tools—polyPhen‑2 HumDiv and FATHMM—suggest a pathogenic effect, while premPS remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) classifies the variant as benign. Taken together, the majority of evidence supports a benign impact and is consistent with the absence of any ClinVar pathogenic annotation. Therefore, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.441902 | Uncertain | 0.897 | 0.642 | 0.250 | -5.694 | Likely Benign | 0.176 | Likely Benign | Likely Benign | 0.06 | Likely Benign | 0.0 | 0.36 | Likely Benign | 0.21 | Likely Benign | 0.92 | Ambiguous | 0.038 | Likely Benign | -1.72 | Neutral | 0.454 | Possibly Damaging | 0.038 | Benign | 1.72 | Pathogenic | 0.34 | Tolerated | 0.1595 | 0.4934 | 0 | 0 | -2.8 | 13.00 | |||||||||||||||||||||||||||||
| c.2032A>C | S678R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta predicts a benign effect. Overall, the majority of tools (8 benign vs. 4 pathogenic) support a benign classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.708 | Likely Pathogenic | 0.878 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.2 | 0.48 | Likely Benign | 0.06 | Likely Benign | 0.32 | Likely Benign | 0.106 | Likely Benign | -2.07 | Neutral | 0.454 | Possibly Damaging | 0.057 | Benign | 3.44 | Benign | 0.02 | Affected | 0.0950 | 0.3519 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2034C>A | S678R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is classified as uncertain. High‑accuracy assessments show AlphaMissense‑Optimized uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta predicts a benign effect. Overall, the majority of evidence (8 benign vs. 4 pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.708 | Likely Pathogenic | 0.878 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.2 | 0.48 | Likely Benign | 0.06 | Likely Benign | 0.32 | Likely Benign | 0.158 | Likely Benign | -2.07 | Neutral | 0.454 | Possibly Damaging | 0.057 | Benign | 3.44 | Benign | 0.02 | Affected | 0.0950 | 0.3519 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2034C>G | S678R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S678R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is classified as uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a benign stability change. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.301917 | Structured | 0.123585 | Uncertain | 0.660 | 0.321 | 0.000 | -9.708 | Likely Pathogenic | 0.878 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.2 | 0.48 | Likely Benign | 0.06 | Likely Benign | 0.32 | Likely Benign | 0.157 | Likely Benign | -2.07 | Neutral | 0.454 | Possibly Damaging | 0.057 | Benign | 3.44 | Benign | 0.02 | Affected | 0.0950 | 0.3519 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||
| c.2215G>A | E739K 2D AIThe SynGAP1 missense variant E739K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.456400 | Uncertain | 0.313 | 0.834 | 0.875 | -5.420 | Likely Benign | 0.343 | Ambiguous | Likely Benign | 0.107 | Likely Benign | -1.49 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.55 | Benign | 0.00 | Affected | 0.2697 | 0.7044 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.2227C>A | P743T 2D AIThe SynGAP1 missense variant P743T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags the variant as pathogenic, creating a single discordant prediction. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence supports a benign classification, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.526809 | Binding | 0.317 | 0.862 | 0.875 | -4.892 | Likely Benign | 0.061 | Likely Benign | Likely Benign | 0.050 | Likely Benign | -1.11 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.75 | Benign | 0.07 | Tolerated | 0.1461 | 0.4750 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.2270G>A | G757D 2D AIThe SynGAP1 missense variant G757D is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Only polyPhen‑2 HumDiv flags it as pathogenic, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Taken together, the preponderance of evidence supports a benign interpretation, and this assessment does not contradict any ClinVar annotation (none is present). Therefore, the variant is most likely benign, with no conflict with ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.830995 | Binding | 0.310 | 0.869 | 0.375 | -4.613 | Likely Benign | 0.387 | Ambiguous | Likely Benign | 0.150 | Likely Benign | -0.90 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.71 | Benign | 0.11 | Tolerated | 0.1826 | 0.1611 | 1 | -1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||||||
| c.2277G>A | M759I 2D AIThe SynGAP1 missense variant M759I is listed in ClinVar (ID 3686687.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33441742‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.525368 | Disordered | 0.879389 | Binding | 0.299 | 0.864 | 0.375 | Uncertain | 1 | 6-33441742-G-A | 1 | 6.20e-7 | -4.058 | Likely Benign | 0.393 | Ambiguous | Likely Benign | 0.075 | Likely Benign | -0.88 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.83 | Benign | 0.34 | Tolerated | 3.99 | 5 | 0.1235 | 0.3129 | 1 | 2 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||
| c.2277G>C | M759I 2D AIThe SynGAP1 missense variant M759I is catalogued in gnomAD (6‑33441742‑G‑C) and has no ClinVar entry. Consensus from multiple in‑silico predictors points to a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all score the variant as benign, while only polyPhen‑2 HumDiv flags it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as “Likely Benign.” High‑accuracy tools reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta results are not available, so they do not influence the assessment. Overall, the computational evidence overwhelmingly supports a benign classification, and this is consistent with the absence of a pathogenic ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.525368 | Disordered | 0.879389 | Binding | 0.299 | 0.864 | 0.375 | 6-33441742-G-C | -4.058 | Likely Benign | 0.393 | Ambiguous | Likely Benign | 0.075 | Likely Benign | -0.88 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.83 | Benign | 0.34 | Tolerated | 3.99 | 5 | 0.1235 | 0.3129 | 1 | 2 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||
| c.2277G>T | M759I 2D AIThe SynGAP1 missense variant M759I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status is unavailable. Overall, the preponderance of evidence from multiple prediction algorithms and high‑accuracy tools points to a benign impact for M759I, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.525368 | Disordered | 0.879389 | Binding | 0.299 | 0.864 | 0.375 | -4.058 | Likely Benign | 0.393 | Ambiguous | Likely Benign | 0.075 | Likely Benign | -0.88 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.83 | Benign | 0.34 | Tolerated | 3.99 | 5 | 0.1235 | 0.3129 | 1 | 2 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||
| c.2807C>G | A936G 2D AIThe SynGAP1 missense variant A936G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.973218 | Binding | 0.319 | 0.874 | 0.625 | -2.720 | Likely Benign | 0.095 | Likely Benign | Likely Benign | 0.065 | Likely Benign | -1.34 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.48 | Pathogenic | 0.12 | Tolerated | 0.2380 | 0.4529 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3769T>G | S1257A 2D AIThe SynGAP1 missense variant S1257A is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also reports a benign likelihood. Foldetta results are unavailable, so they do not influence the conclusion. Overall, the preponderance of evidence points to a benign impact for S1257A, and this assessment is consistent with the lack of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.482380 | Uncertain | 0.889 | 0.572 | 0.375 | -3.937 | Likely Benign | 0.096 | Likely Benign | Likely Benign | 0.118 | Likely Benign | -1.02 | Neutral | 0.454 | Possibly Damaging | 0.266 | Benign | 2.64 | Benign | 0.26 | Tolerated | 0.4083 | 0.3800 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||
| c.3791G>C | G1264A 2D AIThe SynGAP1 missense variant G1264A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while only polyPhen‑2 HumDiv indicates a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign likelihood. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is consistent with the lack of any ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.429200 | Structured | 0.762385 | Binding | 0.897 | 0.579 | 0.000 | -3.460 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -0.32 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 2.79 | Benign | 0.91 | Tolerated | 0.3504 | 0.4220 | 1 | 0 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.385T>C | S129P 2D AIThe SynGAP1 missense variant S129P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of tools, including the high‑accuracy predictors, indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.517562 | Disordered | 0.713635 | Binding | 0.311 | 0.880 | 0.625 | -2.469 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.220 | Likely Benign | -0.09 | Neutral | 0.454 | Possibly Damaging | 0.133 | Benign | 4.11 | Benign | 0.32 | Tolerated | 0.1998 | 0.4989 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.1931A>G | D644G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D644G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and AlphaMissense‑Default (polyPhen‑2 HumVar is benign). High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.066181 | Structured | 0.248888 | Uncertain | 0.883 | 0.320 | 0.000 | -4.496 | Likely Benign | 0.602 | Likely Pathogenic | Likely Benign | 0.13 | Likely Benign | 0.0 | -0.18 | Likely Benign | -0.03 | Likely Benign | 0.04 | Likely Benign | 0.271 | Likely Benign | -4.38 | Deleterious | 0.456 | Possibly Damaging | 0.069 | Benign | 3.46 | Benign | 0.14 | Tolerated | 0.4035 | 0.5610 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||
| c.1999A>C | I667L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I667L is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and gnomAD presence, indicating no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.142424 | Structured | 0.083597 | Uncertain | 0.927 | 0.379 | 0.000 | -10.452 | Likely Pathogenic | 0.459 | Ambiguous | Likely Benign | 0.93 | Ambiguous | 0.2 | 0.72 | Ambiguous | 0.83 | Ambiguous | 0.75 | Ambiguous | 0.300 | Likely Benign | -1.97 | Neutral | 0.457 | Possibly Damaging | 0.392 | Benign | 3.36 | Benign | 0.13 | Tolerated | 0.0980 | 0.3145 | 2 | 2 | -0.7 | 0.00 | ||||||||||||||||||||||||||||||
| c.101A>T | Y34F 2D AIThe SynGAP1 missense variant Y34F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; no Foldetta stability result is available. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.435847 | Uncertain | 0.303 | 0.855 | 0.375 | -3.275 | Likely Benign | 0.127 | Likely Benign | Likely Benign | 0.094 | Likely Benign | -0.50 | Neutral | 0.458 | Possibly Damaging | 0.481 | Possibly Damaging | 4.20 | Benign | 0.00 | Affected | 0.2604 | 0.3591 | 7 | 3 | 4.1 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.116A>T | Y39F 2D AIThe SynGAP1 missense variant Y39F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.425610 | Structured | 0.432876 | Uncertain | 0.343 | 0.787 | 0.375 | -3.410 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.073 | Likely Benign | -0.78 | Neutral | 0.458 | Possibly Damaging | 0.481 | Possibly Damaging | 4.05 | Benign | 0.00 | Affected | 0.2641 | 0.3363 | 7 | 3 | 4.1 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.133A>G | N45D 2D AIThe SynGAP1 missense variant N45D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as likely benign; no Foldetta stability result is available. Overall, the majority of evidence—including the consensus and high‑accuracy predictions—supports a benign classification for this variant, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.401658 | Structured | 0.431853 | Uncertain | 0.498 | 0.741 | 0.375 | -3.340 | Likely Benign | 0.278 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -0.37 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.17 | Benign | 0.00 | Affected | 0.2234 | 0.4999 | 2 | 1 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.134A>G | N45S 2D AIThe SynGAP1 missense variant N45S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for the variant, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.401658 | Structured | 0.431853 | Uncertain | 0.498 | 0.741 | 0.375 | -2.740 | Likely Benign | 0.217 | Likely Benign | Likely Benign | 0.050 | Likely Benign | -0.38 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.14 | Benign | 0.00 | Affected | 0.3949 | 0.7617 | 1 | 1 | 2.7 | -27.03 | |||||||||||||||||||||||||||||||||||||||
| c.160A>G | N54D 2D AIThe SynGAP1 missense variant N54D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is labeled “Likely Benign”). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for the variant, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -6.980 | Likely Benign | 0.325 | Likely Benign | Likely Benign | 0.074 | Likely Benign | -0.75 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.22 | Benign | 0.00 | Affected | 0.1826 | 0.4496 | 2 | 1 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.161A>G | N54S 2D AIThe SynGAP1 missense variant N54S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for the variant, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -5.358 | Likely Benign | 0.125 | Likely Benign | Likely Benign | 0.121 | Likely Benign | -0.17 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.31 | Benign | 0.00 | Affected | 0.3740 | 0.7048 | 1 | 1 | 2.7 | -27.03 | |||||||||||||||||||||||||||||||||||||||
| c.166C>G | L56V 2D AIThe SynGAP1 missense variant L56V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also leans benign (2 benign vs 1 pathogenic, 1 uncertain). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for the L56V variant, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.342579 | Structured | 0.476218 | Uncertain | 0.495 | 0.657 | 0.000 | -8.104 | Likely Pathogenic | 0.471 | Ambiguous | Likely Benign | 0.123 | Likely Benign | -0.99 | Neutral | 0.458 | Possibly Damaging | 0.745 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.1635 | 0.3655 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.169C>A | L57I 2D  AIThe SynGAP1 missense variant L57I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for the L57I variant, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.254060 | Structured | 0.481044 | Uncertain | 0.554 | 0.642 | 0.000 | -3.681 | Likely Benign | 0.275 | Likely Benign | Likely Benign | 0.024 | Likely Benign | -0.07 | Neutral | 0.458 | Possibly Damaging | 0.745 | Possibly Damaging | 4.10 | Benign | 0.00 | Affected | 0.0973 | 0.4106 | 2 | 2 | 0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.169C>G | L57V 2D AIThe SynGAP1 missense variant L57V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for the variant, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.254060 | Structured | 0.481044 | Uncertain | 0.554 | 0.642 | 0.000 | -3.598 | Likely Benign | 0.256 | Likely Benign | Likely Benign | 0.051 | Likely Benign | -0.14 | Neutral | 0.458 | Possibly Damaging | 0.745 | Possibly Damaging | 4.17 | Benign | 0.00 | Affected | 0.1533 | 0.4006 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.175C>G | L59V 2D AIThe SynGAP1 missense variant L59V is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for this variant. This conclusion is consistent with the absence of a ClinVar classification, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.212910 | Structured | 0.484882 | Uncertain | 0.510 | 0.668 | 0.000 | -4.465 | Likely Benign | 0.588 | Likely Pathogenic | Likely Benign | 0.049 | Likely Benign | -1.15 | Neutral | 0.458 | Possibly Damaging | 0.745 | Possibly Damaging | 3.36 | Benign | 0.00 | Affected | 0.1508 | 0.3217 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.181G>A | E61K 2D AIThe SynGAP1 E61K missense variant has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two consensus groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the aggregate evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | -4.953 | Likely Benign | 0.425 | Ambiguous | Likely Benign | 0.120 | Likely Benign | -0.22 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.34 | Benign | 0.00 | Affected | 0.2736 | 0.5691 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.182A>C | E61A 2D AIThe SynGAP1 missense variant E61A is listed in ClinVar (ID 3767543.0) with an *Uncertain* clinical significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the current ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | Uncertain | 1 | -5.235 | Likely Benign | 0.453 | Ambiguous | Likely Benign | 0.074 | Likely Benign | -1.52 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.12 | Benign | 0.00 | Affected | 0.4499 | 0.5878 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||
| c.187G>A | E63K 2D AIThe SynGAP1 E63K missense variant (ClinVar ID 2830630.0) is listed as “Uncertain” and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default all predict a pathogenic outcome. AlphaMissense‑Optimized is inconclusive, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. Overall, the high‑accuracy consensus leans toward a benign effect, and this assessment does not contradict the ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.474807 | Uncertain | 0.494 | 0.739 | 0.125 | Uncertain | 1 | -4.976 | Likely Benign | 0.894 | Likely Pathogenic | Ambiguous | 0.103 | Likely Benign | -0.70 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 3.98 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1995 | 0.7261 | 1 | 0 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||
| c.188A>C | E63A 2D AIThe SynGAP1 missense variant E63A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on benign impact include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign effect, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.474807 | Uncertain | 0.494 | 0.739 | 0.125 | -3.426 | Likely Benign | 0.850 | Likely Pathogenic | Ambiguous | 0.120 | Likely Benign | -1.84 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.3281 | 0.6649 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.199C>A | L67I 2D AIThe SynGAP1 missense variant L67I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for the variant, and this is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.458154 | Structured | 0.473668 | Uncertain | 0.428 | 0.761 | 0.125 | -4.387 | Likely Benign | 0.307 | Likely Benign | Likely Benign | 0.084 | Likely Benign | -0.29 | Neutral | 0.458 | Possibly Damaging | 0.364 | Benign | 4.10 | Benign | 0.00 | Affected | 0.0680 | 0.2919 | 2 | 2 | 0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.199C>G | L67V 2D AIThe SynGAP1 missense variant L67V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for the L67V substitution, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.458154 | Structured | 0.473668 | Uncertain | 0.428 | 0.761 | 0.125 | -3.617 | Likely Benign | 0.285 | Likely Benign | Likely Benign | 0.122 | Likely Benign | -0.31 | Neutral | 0.458 | Possibly Damaging | 0.364 | Benign | 4.15 | Benign | 0.00 | Affected | 0.1138 | 0.2817 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.202C>G | L68V 2D AIThe SynGAP1 missense variant L68V has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized independently predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.534167 | Disordered | 0.470567 | Uncertain | 0.405 | 0.768 | 0.250 | -4.079 | Likely Benign | 0.470 | Ambiguous | Likely Benign | 0.028 | Likely Benign | -0.43 | Neutral | 0.458 | Possibly Damaging | 0.364 | Benign | 4.13 | Benign | 0.00 | Affected | 0.1077 | 0.2817 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.1135T>A | S379T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S379T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, polyPhen‑2 HumVar, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are Rosetta, polyPhen‑2 HumDiv, and the Foldetta stability method. FoldX is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as likely benign, and Foldetta as pathogenic. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.728858 | Disordered | 0.433206 | Uncertain | 0.327 | 0.931 | 0.625 | -5.646 | Likely Benign | 0.092 | Likely Benign | Likely Benign | 1.42 | Ambiguous | 0.6 | 3.96 | Destabilizing | 2.69 | Destabilizing | 0.06 | Likely Benign | 0.230 | Likely Benign | -0.50 | Neutral | 0.462 | Possibly Damaging | 0.084 | Benign | 3.87 | Benign | 0.16 | Tolerated | 0.2293 | 0.6248 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||
| c.1166C>T | S389L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S389L is catalogued in gnomAD (6‑33438071‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, Foldetta, and the SGM‑Consensus score (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments are consistent with the benign consensus: AlphaMissense‑Optimized reports Benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts Benign. No prediction or folding stability result is missing or inconclusive. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.703578 | Disordered | 0.417444 | Uncertain | 0.306 | 0.803 | 0.875 | 6-33438071-C-T | 7 | 4.40e-6 | -6.040 | Likely Benign | 0.165 | Likely Benign | Likely Benign | 0.04 | Likely Benign | 0.1 | 0.69 | Ambiguous | 0.37 | Likely Benign | -0.23 | Likely Benign | 0.456 | Likely Benign | -0.75 | Neutral | 0.462 | Possibly Damaging | 0.693 | Possibly Damaging | 5.05 | Benign | 0.01 | Affected | 4.32 | 8 | 0.1844 | 0.5871 | -2 | -3 | 4.6 | 26.08 | ||||||||||||||||||||||||
| c.145T>A | C49S 2D AIThe SynGAP1 missense variant C49S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of standard predictors lean toward pathogenicity, but the high‑accuracy tools do not provide definitive support. Thus, the variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.575 | Likely Benign | 0.704 | Likely Pathogenic | Likely Benign | 0.260 | Likely Benign | -3.07 | Deleterious | 0.462 | Possibly Damaging | 0.478 | Possibly Damaging | 3.91 | Benign | 0.00 | Affected | 0.3855 | 0.1686 | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.145T>G | C49G 2D AIThe SynGAP1 missense variant C49G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic versus four benign) lean toward a pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.464 | Likely Benign | 0.674 | Likely Pathogenic | Likely Benign | 0.351 | Likely Benign | -3.64 | Deleterious | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.87 | Benign | 0.00 | Affected | 0.2419 | 0.2528 | -3 | -3 | -2.9 | -46.09 | ||||||||||||||||||||||||||||||||||||||||
| c.146G>C | C49S 2D AIThe SynGAP1 missense variant C49S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the majority of conventional tools (five pathogenic vs four benign) lean toward a pathogenic interpretation, but the single high‑accuracy benign prediction and the inconclusive SGM Consensus leave the assessment uncertain. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.575 | Likely Benign | 0.704 | Likely Pathogenic | Likely Benign | 0.224 | Likely Benign | -3.07 | Deleterious | 0.462 | Possibly Damaging | 0.478 | Possibly Damaging | 3.91 | Benign | 0.00 | Affected | 0.3855 | 0.1686 | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.164A>C | Q55P 2D AIThe SynGAP1 missense variant Q55P is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33423573‑A‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs two benign), and Foldetta results are unavailable. Overall, more tools predict pathogenicity than benignity, and no ClinVar entry contradicts this assessment. Therefore, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.332115 | Structured | 0.470108 | Uncertain | 0.461 | 0.657 | 0.000 | 6-33423573-A-C | 1 | 6.20e-7 | -13.163 | Likely Pathogenic | 0.897 | Likely Pathogenic | Ambiguous | 0.260 | Likely Benign | -2.06 | Neutral | 0.462 | Possibly Damaging | 0.480 | Possibly Damaging | 3.83 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2557 | 0.5508 | -1 | 0 | 1.9 | -31.01 | |||||||||||||||||||||||||||||||||||
| c.193C>G | H65D 2D AIThe SynGAP1 H65D missense variant has no ClinVar record and is not listed in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Benign, and AlphaMissense‑Optimized is classified as Uncertain. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | -2.240 | Likely Benign | 0.937 | Likely Pathogenic | Ambiguous | 0.126 | Likely Benign | -1.78 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.17 | Benign | 0.00 | Affected | 0.2142 | 0.1828 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.194A>G | H65R 2D AIThe SynGAP1 missense variant H65R is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33425802‑A‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, whereas the SGM‑Consensus remains benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | Uncertain | 1 | 6-33425802-A-G | 1 | 6.20e-7 | -1.980 | Likely Benign | 0.967 | Likely Pathogenic | Likely Pathogenic | 0.073 | Likely Benign | -1.60 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.19 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1483 | 0.1671 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||
| c.194A>T | H65L 2D AIThe SynGAP1 H65L missense variant has no ClinVar record and is not listed in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” AlphaMissense‑Optimized returns an uncertain result, and Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | -1.889 | Likely Benign | 0.836 | Likely Pathogenic | Ambiguous | 0.159 | Likely Benign | -1.65 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.22 | Benign | 0.00 | Affected | 0.0718 | 0.4760 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||||||||||||
| c.195C>A | H65Q 2D AIThe SynGAP1 missense variant H65Q is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33425803‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain,” SGM‑Consensus as “Likely Benign,” and Foldetta results are unavailable. Based on the overall distribution of predictions, the variant is most likely benign, and this assessment does not contradict the ClinVar status (which has no entry). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | 6-33425803-C-A | 1 | 6.20e-7 | -2.966 | Likely Benign | 0.953 | Likely Pathogenic | Ambiguous | 0.035 | Likely Benign | -1.46 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.19 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1059 | 0.2900 | 0 | 3 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||
| c.195C>G | H65Q 2D AIThe SynGAP1 H65Q missense variant has no ClinVar record and is not listed in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign, while AlphaMissense‑Optimized is uncertain. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for H65Q. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.411940 | Structured | 0.476188 | Uncertain | 0.458 | 0.758 | 0.125 | -2.966 | Likely Benign | 0.953 | Likely Pathogenic | Ambiguous | 0.035 | Likely Benign | -1.46 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.19 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1059 | 0.2900 | 0 | 3 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||
| c.329T>C | V110A 2D AIThe SynGAP1 missense variant V110A is reported as “Likely Benign” in ClinVar and is not listed in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the consensus of the available predictions points to a benign impact for V110A, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.622677 | Disordered | 0.665934 | Binding | 0.347 | 0.860 | 0.750 | -2.971 | Likely Benign | 0.717 | Likely Pathogenic | Likely Benign | 0.075 | Likely Benign | -1.35 | Neutral | 0.462 | Possibly Damaging | 0.122 | Benign | 4.14 | Benign | 0.13 | Tolerated | 0.3196 | 0.2872 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.610T>A | S204T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S204T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate benign or likely benign. Only polyPhen‑2 HumDiv predicts a pathogenic effect. Stability‑based methods (FoldX, Rosetta, Foldetta, premPS) are inconclusive, providing no definitive evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta as uncertain. Taken together, the majority of reliable predictors classify S204T as benign, and this conclusion does not contradict the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | PH | 0.268042 | Structured | 0.420667 | Uncertain | 0.816 | 0.405 | 0.125 | -4.164 | Likely Benign | 0.167 | Likely Benign | Likely Benign | 0.75 | Ambiguous | 0.1 | 1.19 | Ambiguous | 0.97 | Ambiguous | -0.52 | Ambiguous | 0.104 | Likely Benign | 0.55 | Neutral | 0.462 | Possibly Damaging | 0.173 | Benign | 4.19 | Benign | 1.00 | Tolerated | 0.0857 | 0.5063 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||
| c.61T>A | F21I 2D AIThe SynGAP1 missense variant F21I is listed in gnomAD (ID 6‑33420325‑T‑A) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also benign; Foldetta results are unavailable. Taken together, the majority of evidence points to a benign effect. Thus, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.534167 | Disordered | 0.443284 | Uncertain | 0.369 | 0.897 | 0.500 | 6-33420325-T-A | -3.678 | Likely Benign | 0.678 | Likely Pathogenic | Likely Benign | 0.141 | Likely Benign | 0.56 | Neutral | 0.462 | Possibly Damaging | 0.307 | Benign | 4.29 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2703 | 0.2469 | 0 | 1 | 1.7 | -34.02 | ||||||||||||||||||||||||||||||||||||
| c.61T>G | F21V 2D AIThe SynGAP1 missense variant F21V is listed in gnomAD (ID 6‑33420325‑T‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.534167 | Disordered | 0.443284 | Uncertain | 0.369 | 0.897 | 0.500 | 6-33420325-T-G | -2.823 | Likely Benign | 0.563 | Ambiguous | Likely Benign | 0.224 | Likely Benign | 0.64 | Neutral | 0.462 | Possibly Damaging | 0.307 | Benign | 4.44 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2536 | 0.2497 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||
| c.62T>C | F21S 2D AIThe SynGAP1 missense variant F21S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.534167 | Disordered | 0.443284 | Uncertain | 0.369 | 0.897 | 0.500 | -2.396 | Likely Benign | 0.745 | Likely Pathogenic | Likely Benign | 0.182 | Likely Benign | -0.22 | Neutral | 0.462 | Possibly Damaging | 0.307 | Benign | 4.15 | Benign | 0.00 | Affected | 0.4793 | 0.1133 | -3 | -2 | -3.6 | -60.10 | |||||||||||||||||||||||||||||||||||||||
| c.64A>G | R22G 2D AIThe SynGAP1 missense variant R22G is reported in ClinVar as “Not submitted” and is present in gnomAD (ID 6‑33420328‑A‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta results are unavailable. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.441505 | Uncertain | 0.377 | 0.891 | 0.500 | 6-33420328-A-G | -3.628 | Likely Benign | 0.322 | Likely Benign | Likely Benign | 0.185 | Likely Benign | -0.42 | Neutral | 0.462 | Possibly Damaging | 0.152 | Benign | 4.19 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3590 | 0.4098 | -2 | -3 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||
| c.65G>C | R22T 2D AIThe SynGAP1 missense variant R22T is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools show a split: benign calls from REVEL, PROVEAN, polyPhen2_HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic calls come from polyPhen2_HumDiv and SIFT. The majority‑vote SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy predictors further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign; Foldetta predictions are not available. Given the preponderance of benign predictions and the lack of pathogenic evidence, the variant is most likely benign. This assessment aligns with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.441505 | Uncertain | 0.377 | 0.891 | 0.500 | -4.079 | Likely Benign | 0.510 | Ambiguous | Likely Benign | 0.146 | Likely Benign | -0.21 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.23 | Benign | 0.00 | Affected | 0.2140 | 0.5599 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||||
| c.66A>C | R22S 2D AIThe SynGAP1 missense variant R22S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; a Foldetta stability prediction is unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.441505 | Uncertain | 0.377 | 0.891 | 0.500 | -3.419 | Likely Benign | 0.602 | Likely Pathogenic | Likely Benign | 0.190 | Likely Benign | 0.01 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3252 | 0.5187 | -1 | 0 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||
| c.66A>T | R22S 2D AIThe SynGAP1 missense variant R22S is listed in gnomAD (ID 6‑33420330‑A‑T) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its stability impact is unavailable. Overall, the majority of evidence points to a benign effect, and this is not in conflict with ClinVar, which has no classification for R22S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.441505 | Uncertain | 0.377 | 0.891 | 0.500 | 6-33420330-A-T | -3.419 | Likely Benign | 0.602 | Likely Pathogenic | Likely Benign | 0.190 | Likely Benign | 0.01 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3252 | 0.5187 | -1 | 0 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||
| c.88C>G | H30D 2D AIThe SynGAP1 missense variant H30D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for H30D, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | -2.838 | Likely Benign | 0.318 | Likely Benign | Likely Benign | 0.150 | Likely Benign | -2.25 | Neutral | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.93 | Benign | 0.00 | Affected | 0.3048 | 0.3296 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.89A>G | H30R 2D AIThe SynGAP1 missense variant H30R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | -2.156 | Likely Benign | 0.186 | Likely Benign | Likely Benign | 0.138 | Likely Benign | -2.17 | Neutral | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.94 | Benign | 0.00 | Affected | 0.2762 | 0.3590 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.89A>T | H30L 2D AIThe SynGAP1 H30L missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | -2.073 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.163 | Likely Benign | -2.88 | Deleterious | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.94 | Benign | 0.00 | Affected | 0.1523 | 0.5793 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||||||||||||
| c.90C>A | H30Q 2D AIThe SynGAP1 H30Q missense change is catalogued in gnomAD (ID 6‑33423499‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: six methods (REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) and the SGM‑Consensus score all indicate a benign effect, while three tools (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT) predict pathogenicity. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports benign, and the SGM‑Consensus (derived from the four benign‑predicting tools) also yields a benign verdict. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not conflict with ClinVar, which currently contains no classification for H30Q. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | 6-33423499-C-A | 1 | 6.20e-7 | -3.016 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -2.21 | Neutral | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.93 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2409 | 0.4422 | 0 | 3 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||
| c.90C>G | H30Q 2D AIThe SynGAP1 missense variant H30Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is assigned). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.570702 | Disordered | 0.438063 | Uncertain | 0.373 | 0.883 | 0.250 | -3.016 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -2.21 | Neutral | 0.462 | Possibly Damaging | 0.599 | Possibly Damaging | 3.93 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2409 | 0.4422 | 0 | 3 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||
| c.3118G>C | G1040R 2D AIThe SynGAP1 missense variant G1040R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely converge on a deleterious effect: pathogenic calls are made by REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Benign predictions are limited to polyPhen‑2 HumVar and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus remains pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Taken together, the majority of evidence supports a pathogenic classification, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.964893 | Disordered | 0.973805 | Binding | 0.332 | 0.816 | 0.625 | -2.901 | Likely Benign | 0.949 | Likely Pathogenic | Ambiguous | 0.704 | Likely Pathogenic | -3.00 | Deleterious | 0.463 | Possibly Damaging | 0.194 | Benign | -0.74 | Pathogenic | 0.00 | Affected | 0.0924 | 0.4415 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||||||
| c.3200C>A | P1067Q 2D AIThe SynGAP1 missense variant P1067Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 HumDiv and SIFT predict a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign status. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for P1067Q, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.966441 | Disordered | 0.975099 | Binding | 0.459 | 0.907 | 0.875 | -4.767 | Likely Benign | 0.114 | Likely Benign | Likely Benign | 0.136 | Likely Benign | -2.39 | Neutral | 0.463 | Possibly Damaging | 0.087 | Benign | 2.84 | Benign | 0.01 | Affected | 0.1369 | 0.5415 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||||
| c.1493T>G | M498R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M498R is listed in ClinVar as Pathogenic (ClinVar ID 3907767.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include only polyPhen‑2 HumVar; all other evaluated algorithms (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. No predictions or stability results are missing or inconclusive. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, and this conclusion aligns with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.399612 | Uncertain | 0.932 | 0.158 | 0.000 | Likely Pathogenic | 1 | -8.812 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 3.85 | Destabilizing | 0.2 | 2.35 | Destabilizing | 3.10 | Destabilizing | 1.76 | Destabilizing | 0.869 | Likely Pathogenic | -4.53 | Deleterious | 0.464 | Possibly Damaging | 0.120 | Benign | -1.36 | Pathogenic | 0.00 | Affected | 0.1482 | 0.0757 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||
| c.2264T>A | M755K 2D AIThe SynGAP1 missense variant M755K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, SGM‑Consensus indicates Likely Benign, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which contains no conflicting report. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.490133 | Structured | 0.783855 | Binding | 0.336 | 0.873 | 0.375 | -5.642 | Likely Benign | 0.531 | Ambiguous | Likely Benign | 0.101 | Likely Benign | -1.88 | Neutral | 0.468 | Possibly Damaging | 0.206 | Benign | 2.63 | Benign | 0.28 | Tolerated | 0.1183 | 0.0688 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||||||||||||
| c.2264T>G | M755R 2D AIThe SynGAP1 missense variant M755R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and high‑accuracy tools indicates that the M755R variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.490133 | Structured | 0.783855 | Binding | 0.336 | 0.873 | 0.375 | -4.247 | Likely Benign | 0.453 | Ambiguous | Likely Benign | 0.104 | Likely Benign | -2.06 | Neutral | 0.468 | Possibly Damaging | 0.206 | Benign | 2.63 | Benign | 0.18 | Tolerated | 0.1407 | 0.0837 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.2711T>G | M904R 2D AIThe SynGAP1 missense variant M904R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for M904R, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.589073 | Binding | 0.350 | 0.920 | 0.250 | -1.238 | Likely Benign | 0.693 | Likely Pathogenic | Likely Benign | 0.078 | Likely Benign | -0.81 | Neutral | 0.468 | Possibly Damaging | 0.206 | Benign | 2.76 | Benign | 0.82 | Tolerated | 0.1742 | 0.1318 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.3628C>A | H1210N 2D AIThe SynGAP1 missense variant H1210N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta results are not available, so they do not influence the assessment. Overall, the consensus of available predictions indicates that H1210N is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.587579 | Binding | 0.900 | 0.567 | 0.375 | -5.022 | Likely Benign | 0.175 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.48 | Neutral | 0.468 | Possibly Damaging | 0.206 | Benign | 2.71 | Benign | 0.05 | Affected | 0.1335 | 0.2030 | 2 | 1 | -0.3 | -23.04 | ||||||||||||||||||||||||||||||||||||||
| c.2212A>C | S738R 2D AIThe SynGAP1 missense variant S738R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, SGM‑Consensus as Benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for S738R, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.441162 | Uncertain | 0.284 | 0.827 | 0.875 | -4.241 | Likely Benign | 0.570 | Likely Pathogenic | Likely Benign | 0.066 | Likely Benign | -1.55 | Neutral | 0.473 | Possibly Damaging | 0.193 | Benign | 2.69 | Benign | 0.01 | Affected | 4.32 | 2 | 0.0887 | 0.2891 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.2214T>A | S738R 2D AIThe SynGAP1 missense variant S738R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus (majority vote) as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.441162 | Uncertain | 0.284 | 0.827 | 0.875 | -4.241 | Likely Benign | 0.570 | Likely Pathogenic | Likely Benign | 0.068 | Likely Benign | -1.55 | Neutral | 0.473 | Possibly Damaging | 0.193 | Benign | 2.69 | Benign | 0.01 | Affected | 4.32 | 2 | 0.0887 | 0.2891 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.2214T>G | S738R 2D AIThe SynGAP1 missense variant S738R is listed in ClinVar (ID 1592652.0) as Benign and is present in gnomAD (variant ID 6‑33441679‑T‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to Likely Benign (three benign votes versus one pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Benign; Foldetta’s protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, aligning with the ClinVar designation and not contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.441162 | Uncertain | 0.284 | 0.827 | 0.875 | Benign | 1 | 6-33441679-T-G | 1 | 6.20e-7 | -4.241 | Likely Benign | 0.570 | Likely Pathogenic | Likely Benign | 0.068 | Likely Benign | -1.55 | Neutral | 0.473 | Possibly Damaging | 0.193 | Benign | 2.69 | Benign | 0.01 | Affected | 4.32 | 2 | 0.0887 | 0.2891 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||
| c.3032G>T | G1011V 2D AIThe SynGAP1 missense variant G1011V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.898380 | Binding | 0.332 | 0.869 | 0.625 | -4.883 | Likely Benign | 0.129 | Likely Benign | Likely Benign | 0.133 | Likely Benign | -1.21 | Neutral | 0.473 | Possibly Damaging | 0.192 | Benign | 2.71 | Benign | 0.01 | Affected | 0.1352 | 0.3434 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3607C>G | H1203D 2D AIThe SynGAP1 missense variant H1203D is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of predictions points to a benign impact, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.618285 | Disordered | 0.527023 | Binding | 0.892 | 0.589 | 0.250 | Uncertain | 2 | -6.729 | Likely Benign | 0.525 | Ambiguous | Likely Benign | 0.403 | Likely Benign | -1.89 | Neutral | 0.473 | Possibly Damaging | 0.265 | Benign | 5.51 | Benign | 0.24 | Tolerated | 3.77 | 5 | 0.1926 | 0.0530 | 1 | -1 | -0.3 | -22.05 | ||||||||||||||||||||||||||||||||||
| c.3608A>G | H1203R 2D AIThe SynGAP1 missense variant H1203R is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.618285 | Disordered | 0.527023 | Binding | 0.892 | 0.589 | 0.250 | -3.355 | Likely Benign | 0.204 | Likely Benign | Likely Benign | 0.287 | Likely Benign | -1.61 | Neutral | 0.473 | Possibly Damaging | 0.265 | Benign | 5.51 | Benign | 0.20 | Tolerated | 0.1263 | 0.1178 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||||
| c.3608A>T | H1203L 2D AIThe SynGAP1 missense variant H1203L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags the variant as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Benign. High‑accuracy assessments confirm this trend: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.618285 | Disordered | 0.527023 | Binding | 0.892 | 0.589 | 0.250 | -4.179 | Likely Benign | 0.153 | Likely Benign | Likely Benign | 0.361 | Likely Benign | -2.23 | Neutral | 0.473 | Possibly Damaging | 0.265 | Benign | 5.53 | Benign | 0.21 | Tolerated | 0.0682 | 0.2951 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||
| c.1192C>T | P398S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P398S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include polyPhen‑2 (HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv), and SIFT. The remaining tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of definitive predictions lean toward a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | -6.757 | Likely Benign | 0.490 | Ambiguous | Likely Benign | 1.86 | Ambiguous | 0.3 | 1.78 | Ambiguous | 1.82 | Ambiguous | 0.85 | Ambiguous | 0.544 | Likely Pathogenic | -5.58 | Deleterious | 0.478 | Possibly Damaging | 0.130 | Benign | 5.68 | Benign | 0.03 | Affected | 0.3619 | 0.5737 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||
| c.1168G>A | G390R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G390R is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict pathogenicity are SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and Foldetta (combining FoldX‑MD and Rosetta) both predict pathogenicity. Overall, the majority of tools and the high‑accuracy methods indicate a pathogenic effect. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | -9.242 | Likely Pathogenic | 0.686 | Likely Pathogenic | Likely Benign | 2.43 | Destabilizing | 0.9 | 4.85 | Destabilizing | 3.64 | Destabilizing | 0.16 | Likely Benign | 0.605 | Likely Pathogenic | -0.92 | Neutral | 0.480 | Possibly Damaging | 0.163 | Benign | 1.32 | Pathogenic | 0.08 | Tolerated | 0.1190 | 0.4524 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||
| c.1168G>C | G390R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G390R is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict pathogenicity are SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and Foldetta (combining FoldX‑MD and Rosetta) both predict pathogenicity. Overall, the majority of tools and the high‑accuracy methods indicate a pathogenic effect. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | -9.242 | Likely Pathogenic | 0.686 | Likely Pathogenic | Likely Benign | 2.43 | Destabilizing | 0.9 | 4.85 | Destabilizing | 3.64 | Destabilizing | 0.16 | Likely Benign | 0.605 | Likely Pathogenic | -0.92 | Neutral | 0.480 | Possibly Damaging | 0.163 | Benign | 1.32 | Pathogenic | 0.08 | Tolerated | 0.1190 | 0.4524 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||
| c.1189T>C | C397R 2D 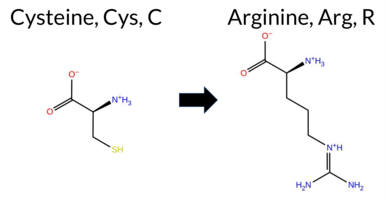 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, and AlphaMissense‑Default. The remaining tools (FoldX, premPS, ESM1b) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign (2 benign vs. 1 pathogenic, 1 uncertain), and Foldetta predicts a benign impact on protein folding stability. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing clinical classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -7.094 | In-Between | 0.708 | Likely Pathogenic | Likely Benign | 0.55 | Ambiguous | 0.8 | 0.40 | Likely Benign | 0.48 | Likely Benign | 0.96 | Ambiguous | 0.514 | Likely Pathogenic | -1.46 | Neutral | 0.480 | Possibly Damaging | 0.226 | Benign | 4.65 | Benign | 0.11 | Tolerated | 0.1780 | 0.1853 | -4 | -3 | -7.0 | 53.05 | ||||||||||||||||||||||||||||||
| c.103G>C | V35L 2D AIThe SynGAP1 missense variant V35L is reported in gnomAD (variant ID 6-33423512‑G‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence, including the high‑accuracy tools, points to a benign effect. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.613573 | Disordered | 0.434838 | Uncertain | 0.360 | 0.851 | 0.375 | 6-33423512-G-C | 4 | 2.48e-6 | -2.893 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.039 | Likely Benign | -0.58 | Neutral | 0.481 | Possibly Damaging | 0.506 | Possibly Damaging | 4.18 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1026 | 0.4025 | 1 | 2 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.2539C>A | Q847K 2D AIThe SynGAP1 missense variant Q847K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign calls come from REVEL, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic calls come from PROVEAN, polyPhen‑2 HumDiv, SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a Likely Pathogenic verdict. High‑accuracy assessments further indicate that AlphaMissense‑Optimized predicts a benign effect, while the SGM Consensus remains Likely Pathogenic; no Foldetta stability data are available. Overall, the majority of predictions lean toward pathogenicity, and this is consistent with the SGM Consensus result. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar annotation exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.653063 | Disordered | 0.577677 | Binding | 0.282 | 0.818 | 0.500 | -5.507 | Likely Benign | 0.736 | Likely Pathogenic | Likely Benign | 0.214 | Likely Benign | -2.82 | Deleterious | 0.481 | Possibly Damaging | 0.373 | Benign | 2.32 | Pathogenic | 0.00 | Affected | 0.1694 | 0.4129 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.2605C>G | L869V 2D AIThe SynGAP1 missense variant L869V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.501700 | Disordered | 0.688653 | Binding | 0.272 | 0.839 | 0.250 | -3.241 | Likely Benign | 0.161 | Likely Benign | Likely Benign | 0.093 | Likely Benign | -0.33 | Neutral | 0.481 | Possibly Damaging | 0.300 | Benign | 2.86 | Benign | 0.11 | Tolerated | 0.1501 | 0.3616 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.2671C>A | L891I 2D AIThe SynGAP1 missense variant L891I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the majority of computational evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.712013 | Disordered | 0.505861 | Binding | 0.305 | 0.923 | 0.750 | -5.803 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.048 | Likely Benign | -0.71 | Neutral | 0.481 | Possibly Damaging | 0.202 | Benign | 2.74 | Benign | 0.14 | Tolerated | 0.0957 | 0.3494 | 2 | 2 | 0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.2727G>A | M909I 2D AIThe SynGAP1 missense variant M909I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.696196 | Binding | 0.314 | 0.914 | 0.250 | -3.636 | Likely Benign | 0.703 | Likely Pathogenic | Likely Benign | 0.097 | Likely Benign | -1.15 | Neutral | 0.481 | Possibly Damaging | 0.220 | Benign | 2.82 | Benign | 0.25 | Tolerated | 0.1392 | 0.3298 | 2 | 1 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.2727G>C | M909I 2D AIThe SynGAP1 missense variant M909I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.696196 | Binding | 0.314 | 0.914 | 0.250 | -3.636 | Likely Benign | 0.703 | Likely Pathogenic | Likely Benign | 0.097 | Likely Benign | -1.15 | Neutral | 0.481 | Possibly Damaging | 0.220 | Benign | 2.82 | Benign | 0.25 | Tolerated | 0.1392 | 0.3298 | 2 | 1 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.2727G>T | M909I 2D AIThe SynGAP1 missense variant M909I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.696196 | Binding | 0.314 | 0.914 | 0.250 | -3.636 | Likely Benign | 0.703 | Likely Pathogenic | Likely Benign | 0.097 | Likely Benign | -1.15 | Neutral | 0.481 | Possibly Damaging | 0.220 | Benign | 2.82 | Benign | 0.25 | Tolerated | 0.1392 | 0.3298 | 2 | 1 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.2932C>T | P978S 2D AIThe SynGAP1 missense variant P978S is listed in ClinVar (ID 3379672.0) with an “Uncertain” clinical significance and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of predictions points to a benign effect, which is consistent with the ClinVar “Uncertain” status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.819762 | Disordered | 0.975775 | Binding | 0.425 | 0.892 | 0.625 | Uncertain | 1 | -3.913 | Likely Benign | 0.151 | Likely Benign | Likely Benign | 0.085 | Likely Benign | -1.07 | Neutral | 0.481 | Possibly Damaging | 0.220 | Benign | 4.22 | Benign | 0.48 | Tolerated | 0.3428 | 0.6001 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||
| c.3080A>C | N1027T 2D AIThe SynGAP1 missense variant N1027T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while only polyPhen‑2 HumDiv indicates a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign and the SGM‑Consensus also reports Likely Benign; Foldetta results are not available. Overall, the consensus of the available predictions points to a benign effect, and this conclusion is consistent with the absence of any ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.831250 | Disordered | 0.994357 | Binding | 0.347 | 0.745 | 0.500 | -3.604 | Likely Benign | 0.199 | Likely Benign | Likely Benign | 0.046 | Likely Benign | -0.71 | Neutral | 0.481 | Possibly Damaging | 0.220 | Benign | 2.75 | Benign | 0.13 | Tolerated | 0.1239 | 0.7188 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||||||||||||
| c.3081C>A | N1027K 2D AIThe SynGAP1 missense variant N1027K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and the SGM‑Consensus result is benign; Foldetta predictions are unavailable. Overall, the balance of evidence favors a benign interpretation, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.831250 | Disordered | 0.994357 | Binding | 0.347 | 0.745 | 0.500 | -3.177 | Likely Benign | 0.841 | Likely Pathogenic | Ambiguous | 0.063 | Likely Benign | -0.64 | Neutral | 0.481 | Possibly Damaging | 0.220 | Benign | 2.81 | Benign | 0.65 | Tolerated | 0.1808 | 0.6079 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3081C>G | N1027K 2D AIThe SynGAP1 missense variant N1027K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign, and AlphaMissense‑Optimized is uncertain. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.831250 | Disordered | 0.994357 | Binding | 0.347 | 0.745 | 0.500 | -3.177 | Likely Benign | 0.841 | Likely Pathogenic | Ambiguous | 0.063 | Likely Benign | -0.64 | Neutral | 0.481 | Possibly Damaging | 0.220 | Benign | 2.81 | Benign | 0.65 | Tolerated | 0.1808 | 0.6079 | 1 | 0 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3103C>G | P1035A 2D AIThe SynGAP1 missense variant P1035A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic outcome, creating a single discordant signal. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also reports Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.945666 | Disordered | 0.989572 | Binding | 0.300 | 0.756 | 0.625 | -4.293 | Likely Benign | 0.241 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.02 | Neutral | 0.481 | Possibly Damaging | 0.222 | Benign | 2.74 | Benign | 0.41 | Tolerated | 0.3188 | 0.6022 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3239C>T | A1080V 2D AIThe SynGAP1 missense variant A1080V is listed in gnomAD (ID 6‑33443791‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; no Foldetta stability data are available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.981457 | Binding | 0.303 | 0.900 | 0.750 | 6-33443791-C-T | -4.087 | Likely Benign | 0.229 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -1.06 | Neutral | 0.481 | Possibly Damaging | 0.144 | Benign | 3.99 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1298 | 0.6056 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||
| c.3250C>T | P1084S 2D AIThe SynGAP1 missense variant P1084S is reported in gnomAD (variant ID 6‑33443802‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 HumDiv and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.842060 | Disordered | 0.979020 | Binding | 0.348 | 0.889 | 1.000 | 6-33443802-C-T | 1 | 6.31e-7 | -3.987 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.086 | Likely Benign | -2.24 | Neutral | 0.481 | Possibly Damaging | 0.157 | Benign | 4.03 | Benign | 0.03 | Affected | 3.77 | 5 | 0.3102 | 0.6215 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3389A>C | K1130T 2D AIThe SynGAP1 missense variant K1130T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) which classifies it as Likely Benign. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized result is Uncertain, and the Foldetta protein‑folding stability assessment is unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.863782 | Binding | 0.350 | 0.904 | 0.750 | -3.550 | Likely Benign | 0.829 | Likely Pathogenic | Ambiguous | 0.397 | Likely Benign | -1.38 | Neutral | 0.481 | Possibly Damaging | 0.157 | Benign | 5.47 | Benign | 0.00 | Affected | 0.2572 | 0.4349 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.3898C>T | P1300S 2D AIThe SynGAP1 missense variant P1300S is reported in gnomAD (variant ID 6‑33451772‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant call. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that P1300S is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.885826 | Binding | 0.400 | 0.834 | 0.875 | 6-33451772-C-T | 1 | 6.20e-7 | -3.476 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.036 | Likely Benign | -0.29 | Neutral | 0.481 | Possibly Damaging | 0.157 | Benign | 2.89 | Benign | 0.27 | Tolerated | 3.77 | 5 | 0.3242 | 0.4139 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.1408A>C | M470L 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant M470L is listed in ClinVar as benign (ClinVar ID 536996.0) and is present in gnomAD (variant ID 6‑33438440‑A‑C). Functional prediction tools cluster into two groups: benign predictions come from SIFT and AlphaMissense‑Optimized, while pathogenic predictions are made by REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as likely pathogenic. High‑accuracy assessments further show AlphaMissense‑Optimized as benign, SGM Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. No definitive folding‑stability change is reported by FoldX or Rosetta individually. Overall, the majority of predictive algorithms favor a pathogenic effect, directly contradicting the benign classification in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.298791 | Structured | 0.351497 | Uncertain | 0.908 | 0.272 | 0.000 | Likely Benign | 1 | 6-33438440-A-C | 1 | 6.20e-7 | -8.993 | Likely Pathogenic | 0.406 | Ambiguous | Likely Benign | 0.73 | Ambiguous | 0.1 | 0.84 | Ambiguous | 0.79 | Ambiguous | 1.04 | Destabilizing | 0.678 | Likely Pathogenic | -2.72 | Deleterious | 0.484 | Possibly Damaging | 0.654 | Possibly Damaging | -1.22 | Pathogenic | 0.16 | Tolerated | 3.37 | 34 | 0.1192 | 0.4071 | 4 | 2 | 1.9 | -18.03 | 225.3 | 17.9 | 0.0 | 0.0 | -0.8 | 0.5 | X | Potentially Benign | The thioether group of Met470, located in the middle of an α helix (res. Ala461–Phe476), interacts with hydrophobic residues in the inter-helix space (e.g., Val473, Leu558) formed by two other α helices (res. Ser604–Arg581, res. Pro562–Arg579). In the WT simulations, Met470 also packs against the positively charged guanidinium groups of Arg575, Arg429, and Arg579, which form salt bridges with the negatively charged carboxylate groups of the Asp474 and Asp467 side chains at the protein surface. In the variant simulations, the iso-butyl side chain of Leu470 packs similarly with the hydrophobic residues as methionine, resulting in no negative effects on the protein structure during the simulation. | |||||||||||||
| c.1408A>T | M470L 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant M470L is not reported in ClinVar and has no gnomAD entry. Prediction tools that agree on benign effect include SIFT and AlphaMissense‑Optimized. Tools that agree on pathogenic effect include SGM Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. AlphaMissense‑Default, FoldX, Rosetta, and Foldetta are inconclusive and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, whereas the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts likely pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is uncertain. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.298791 | Structured | 0.351497 | Uncertain | 0.908 | 0.272 | 0.000 | -8.993 | Likely Pathogenic | 0.406 | Ambiguous | Likely Benign | 0.73 | Ambiguous | 0.1 | 0.84 | Ambiguous | 0.79 | Ambiguous | 1.04 | Destabilizing | 0.678 | Likely Pathogenic | -2.72 | Deleterious | 0.484 | Possibly Damaging | 0.654 | Possibly Damaging | -1.22 | Pathogenic | 0.16 | Tolerated | 3.37 | 34 | 0.1192 | 0.4071 | 4 | 2 | 1.9 | -18.03 | 225.3 | 17.9 | 0.0 | 0.0 | -0.8 | 0.5 | X | Potentially Benign | The thioether group of Met470, located in the middle of an α helix (res. Ala461–Phe476), interacts with hydrophobic residues in the inter-helix space (e.g., Val473, Leu558) formed by two other α helices (res. Ser604–Arg581, res. Pro562–Arg579). In the WT simulations, Met470 also packs against the positively charged guanidinium groups of Arg575, Arg429, and Arg579, which form salt bridges with the negatively charged carboxylate groups of the Asp474 and Asp467 side chains at the protein surface. In the variant simulations, the iso-butyl side chain of Leu470 packs similarly with the hydrophobic residues as methionine, resulting in no negative effects on the protein structure during the simulation. | ||||||||||||||||||
| c.1807A>C | M603L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 M603L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that classify the variant as benign include FoldX, premPS, PROVEAN, SIFT, AlphaMissense‑Optimized, and the protein‑folding stability method Foldetta. Those that predict pathogenicity are REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. Two tools give uncertain results: Rosetta and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, more evidence supports a benign effect (six benign versus five pathogenic predictions, with two uncertain). Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.011342 | Structured | 0.197847 | Uncertain | 0.942 | 0.176 | 0.000 | -7.134 | In-Between | 0.590 | Likely Pathogenic | Likely Benign | 0.04 | Likely Benign | 0.1 | 0.86 | Ambiguous | 0.45 | Likely Benign | -0.18 | Likely Benign | 0.548 | Likely Pathogenic | -2.16 | Neutral | 0.484 | Possibly Damaging | 0.654 | Possibly Damaging | -0.98 | Pathogenic | 0.52 | Tolerated | 0.1345 | 0.3227 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.1807A>T | M603L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 M603L missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. Rosetta and ESM1b give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools lean toward a benign interpretation, but the SGM Consensus indicates pathogenicity, leaving the variant’s clinical significance uncertain. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for M603L. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.011342 | Structured | 0.197847 | Uncertain | 0.942 | 0.176 | 0.000 | -7.134 | In-Between | 0.590 | Likely Pathogenic | Likely Benign | 0.04 | Likely Benign | 0.1 | 0.86 | Ambiguous | 0.45 | Likely Benign | -0.18 | Likely Benign | 0.548 | Likely Pathogenic | -2.16 | Neutral | 0.484 | Possibly Damaging | 0.654 | Possibly Damaging | -0.98 | Pathogenic | 0.52 | Tolerated | 0.1345 | 0.3227 | 4 | 2 | 1.9 | -18.03 | ||||||||||||||||||||||||||||||
| c.41C>G | P14R 2D AIThe SynGAP1 missense variant P14R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The high‑accuracy consensus methods give a benign verdict: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” No result is available from Foldetta, so its folding‑stability assessment is not considered. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of any ClinVar classification. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | -2.794 | Likely Benign | 0.253 | Likely Benign | Likely Benign | 0.183 | Likely Benign | 0.13 | Neutral | 0.486 | Possibly Damaging | 0.032 | Benign | 4.20 | Benign | 0.00 | Affected | 0.1597 | 0.3536 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.50C>G | S17C 2D 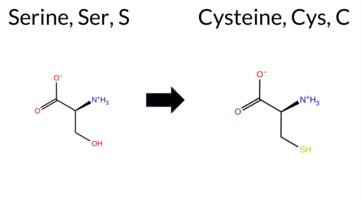 AIThe SynGAP1 missense variant S17C is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools (polyPhen‑2 HumDiv and SIFT) predict pathogenicity, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | -4.364 | Likely Benign | 0.279 | Likely Benign | Likely Benign | 0.044 | Likely Benign | 0.17 | Neutral | 0.486 | Possibly Damaging | 0.048 | Benign | 3.99 | Benign | 0.00 | Affected | 0.1226 | 0.6721 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.50C>T | S17F 2D AIThe SynGAP1 missense variant S17F is listed in ClinVar (ID 3451958) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33420314‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | Conflicting | 2 | 6-33420314-C-T | 10 | 6.49e-6 | -3.888 | Likely Benign | 0.637 | Likely Pathogenic | Likely Benign | 0.048 | Likely Benign | -0.99 | Neutral | 0.486 | Possibly Damaging | 0.032 | Benign | 3.99 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0729 | 0.6468 | -2 | -3 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||
| c.406C>G | R136G 2D AIThe SynGAP1 missense variant R136G is not reported in ClinVar and is absent from gnomAD. Consensus from standard in‑silico predictors shows a split: benign calls come from REVEL, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic calls arise from PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.433034 | Structured | 0.657394 | Binding | 0.351 | 0.894 | 0.250 | -10.641 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.246 | Likely Benign | -3.46 | Deleterious | 0.487 | Possibly Damaging | 0.211 | Benign | 3.47 | Benign | 0.00 | Affected | 0.3581 | 0.3585 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.484C>A | R162S 2D AIThe SynGAP1 missense variant R162S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign (three benign votes versus one pathogenic). AlphaMissense‑Optimized is currently Uncertain, and no Foldetta stability result is available. Overall, the majority of high‑accuracy and consensus predictions indicate a benign impact. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.501700 | Disordered | 0.516348 | Binding | 0.315 | 0.692 | 0.250 | -1.395 | Likely Benign | 0.894 | Likely Pathogenic | Ambiguous | 0.191 | Likely Benign | -0.24 | Neutral | 0.487 | Possibly Damaging | 0.272 | Benign | 4.14 | Benign | 0.75 | Tolerated | 0.3080 | 0.4974 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||||
| c.484C>G | R162G 2D AIThe SynGAP1 missense variant R162G is listed in ClinVar (ID 2703066.0) with an uncertain significance status and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign (three benign votes versus one pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta stability analysis is unavailable. Overall, the majority of predictions support a benign impact, and this is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.501700 | Disordered | 0.516348 | Binding | 0.315 | 0.692 | 0.250 | Uncertain | 1 | -6.985 | Likely Benign | 0.664 | Likely Pathogenic | Likely Benign | 0.190 | Likely Benign | -0.73 | Neutral | 0.487 | Possibly Damaging | 0.272 | Benign | 4.09 | Benign | 0.78 | Tolerated | 3.74 | 4 | 0.3562 | 0.4308 | -2 | -3 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||
| c.490C>G | R164G 2D AIThe SynGAP1 missense variant R164G has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a split opinion: benign calls come from REVEL, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: the SGM‑Consensus remains Likely Pathogenic, AlphaMissense‑Optimized is inconclusive, and the Foldetta stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic impact. Because there is no ClinVar classification to oppose this, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.414856 | Structured | 0.512396 | Binding | 0.317 | 0.666 | 0.250 | -12.416 | Likely Pathogenic | 0.879 | Likely Pathogenic | Ambiguous | 0.190 | Likely Benign | -3.01 | Deleterious | 0.487 | Possibly Damaging | 0.272 | Benign | 3.77 | Benign | 0.00 | Affected | 0.3657 | 0.3631 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.2761C>A | L921M 2D 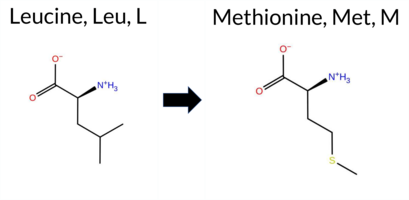 AIThe SynGAP1 missense variant L921M is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and FATHMM. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign impact for L921M, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.608892 | Disordered | 0.943282 | Binding | 0.311 | 0.845 | 0.375 | -4.485 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -0.38 | Neutral | 0.489 | Possibly Damaging | 0.137 | Benign | 2.40 | Pathogenic | 0.02 | Affected | 0.0762 | 0.3732 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.2191C>A | Q731K 2D AIThe SynGAP1 missense variant Q731K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy tools points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.750527 | Disordered | 0.415202 | Uncertain | 0.507 | 0.654 | 0.750 | -6.686 | Likely Benign | 0.349 | Ambiguous | Likely Benign | 0.066 | Likely Benign | -1.58 | Neutral | 0.490 | Possibly Damaging | 0.149 | Benign | 2.67 | Benign | 0.20 | Tolerated | 0.1998 | 0.3932 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.2386C>A | P796T 2D AIThe SynGAP1 P796T missense variant is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | SH3-binding motif | 0.936162 | Disordered | 0.426363 | Uncertain | 0.427 | 0.900 | 0.875 | -5.399 | Likely Benign | 0.062 | Likely Benign | Likely Benign | 0.082 | Likely Benign | -0.01 | Neutral | 0.494 | Possibly Damaging | 0.236 | Benign | 4.26 | Benign | 0.04 | Affected | 0.1705 | 0.4961 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||||
| c.2878C>A | H960N 2D AIThe SynGAP1 missense variant H960N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987911 | Disordered | 0.983385 | Binding | 0.380 | 0.901 | 0.750 | -8.822 | Likely Pathogenic | 0.101 | Likely Benign | Likely Benign | 0.130 | Likely Benign | -0.57 | Neutral | 0.494 | Possibly Damaging | 0.129 | Benign | 4.19 | Benign | 0.40 | Tolerated | 0.2137 | 0.3695 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.2878C>G | H960D 2D AIThe SynGAP1 missense variant H960D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. The predictions do not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987911 | Disordered | 0.983385 | Binding | 0.380 | 0.901 | 0.750 | -12.235 | Likely Pathogenic | 0.243 | Likely Benign | Likely Benign | 0.147 | Likely Benign | -1.09 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.19 | Benign | 0.31 | Tolerated | 0.2504 | 0.2923 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.2879A>C | H960P 2D AIThe SynGAP1 missense variant H960P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987911 | Disordered | 0.983385 | Binding | 0.380 | 0.901 | 0.750 | -8.672 | Likely Pathogenic | 0.064 | Likely Benign | Likely Benign | 0.119 | Likely Benign | -0.95 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.18 | Benign | 0.28 | Tolerated | 0.2157 | 0.4901 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2879A>G | H960R 2D AIThe SynGAP1 missense variant H960R is reported in gnomAD (ID 6‑33443431‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only two tools predict pathogenicity—polyPhen‑2 HumDiv and ESM1b—while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also reports likely benign. No Foldetta stability data are available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987911 | Disordered | 0.983385 | Binding | 0.380 | 0.901 | 0.750 | 6-33443431-A-G | 1 | 6.20e-7 | -9.238 | Likely Pathogenic | 0.192 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -1.10 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.19 | Benign | 0.25 | Tolerated | 3.77 | 5 | 0.2237 | 0.3667 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.2885A>T | H962L 2D AIThe SynGAP1 missense variant H962L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and ESM1b. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for H962L, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.991070 | Disordered | 0.984483 | Binding | 0.369 | 0.886 | 0.750 | -8.478 | Likely Pathogenic | 0.108 | Likely Benign | Likely Benign | 0.151 | Likely Benign | -1.49 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.15 | Benign | 0.03 | Affected | 0.1738 | 0.5487 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||||||||||||
| c.2896C>G | H966D 2D AIThe SynGAP1 missense variant H966D is listed in gnomAD (ID 6‑33443448‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign or likely benign. Only two tools predict a damaging outcome—polyPhen‑2 HumDiv and ESM1b—which are outliers relative to the consensus. High‑accuracy assessments confirm the benign trend: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus also indicates a likely benign status. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions support a benign impact, and this is consistent with the absence of a pathogenic ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.976962 | Disordered | 0.974672 | Binding | 0.378 | 0.879 | 0.750 | 6-33443448-C-G | 1 | 6.20e-7 | -8.426 | Likely Pathogenic | 0.201 | Likely Benign | Likely Benign | 0.182 | Likely Benign | -1.09 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.05 | Benign | 0.93 | Tolerated | 4.32 | 2 | 0.2504 | 0.3066 | -1 | 1 | -0.3 | -22.05 | ||||||||||||||||||||||||||||||||||
| c.2897A>G | H966R 2D AIThe SynGAP1 missense variant H966R is reported in gnomAD (ID 6‑33443449‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant call. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that H966R is most likely benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.976962 | Disordered | 0.974672 | Binding | 0.378 | 0.879 | 0.750 | 6-33443449-A-G | -5.474 | Likely Benign | 0.157 | Likely Benign | Likely Benign | 0.172 | Likely Benign | -0.71 | Neutral | 0.494 | Possibly Damaging | 0.170 | Benign | 4.06 | Benign | 0.69 | Tolerated | 4.32 | 2 | 0.2198 | 0.3410 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||
| c.1069C>A | H357N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H357N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Taken together, the overwhelming majority of evidence points to a benign impact. There is no ClinVar annotation to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -7.617 | In-Between | 0.103 | Likely Benign | Likely Benign | 0.14 | Likely Benign | 0.2 | 0.31 | Likely Benign | 0.23 | Likely Benign | 0.31 | Likely Benign | 0.126 | Likely Benign | -1.47 | Neutral | 0.495 | Possibly Damaging | 0.169 | Benign | 4.25 | Benign | 0.15 | Tolerated | 0.2052 | 0.3266 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||
| c.1069C>G | H357D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H357D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta also predicts a benign outcome. No prediction or folding‑stability result is missing or inconclusive. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -12.013 | Likely Pathogenic | 0.559 | Ambiguous | Likely Benign | -0.23 | Likely Benign | 0.4 | -0.27 | Likely Benign | -0.25 | Likely Benign | 0.48 | Likely Benign | 0.208 | Likely Benign | -1.90 | Neutral | 0.495 | Possibly Damaging | 0.169 | Benign | 4.22 | Benign | 0.08 | Tolerated | 0.2782 | 0.1976 | 1 | -1 | -0.3 | -22.05 | ||||||||||||||||||||||||||||||
| c.1070A>G | H357R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H357R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. Two tools remain uncertain: Rosetta and AlphaMissense‑Default. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority; and Foldetta also predicts benign. No prediction or stability result is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -9.212 | Likely Pathogenic | 0.348 | Ambiguous | Likely Benign | -0.10 | Likely Benign | 0.3 | 1.07 | Ambiguous | 0.49 | Likely Benign | 0.32 | Likely Benign | 0.107 | Likely Benign | -1.03 | Neutral | 0.495 | Possibly Damaging | 0.095 | Benign | 4.22 | Benign | 0.59 | Tolerated | 0.2147 | 0.2818 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||
| c.1258T>G | F420V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F420V is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM. In contrast, the majority of tools predict a pathogenic outcome: FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts Pathogenic; the SGM‑Consensus itself is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts Pathogenic. Based on the preponderance of pathogenic predictions and the high‑accuracy tool consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.104810 | Structured | 0.384475 | Uncertain | 0.974 | 0.255 | 0.000 | -11.849 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 3.42 | Destabilizing | 0.1 | 3.35 | Destabilizing | 3.39 | Destabilizing | 1.58 | Destabilizing | 0.402 | Likely Benign | -6.41 | Deleterious | 0.495 | Possibly Damaging | 0.191 | Benign | 3.16 | Benign | 0.01 | Affected | 0.2038 | 0.2022 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.1147G>A | G383R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G383R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that agree on a pathogenic effect are FoldX, Rosetta, Foldetta, polyPhen2_HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, Foldetta predicts a pathogenic outcome, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs. two benign). Overall, the majority of predictions lean toward pathogenicity, and this does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | -9.067 | Likely Pathogenic | 0.660 | Likely Pathogenic | Likely Benign | 4.03 | Destabilizing | 2.3 | 3.36 | Destabilizing | 3.70 | Destabilizing | 0.29 | Likely Benign | 0.449 | Likely Benign | -0.84 | Neutral | 0.498 | Possibly Damaging | 0.119 | Benign | 4.10 | Benign | 0.00 | Affected | 0.1295 | 0.3741 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.1147G>C | G383R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G383R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that agree on a pathogenic effect are FoldX, Rosetta, Foldetta, polyPhen2_HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, Foldetta predicts a pathogenic outcome, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs. two benign). Overall, the majority of predictions lean toward pathogenicity, and this does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | -9.067 | Likely Pathogenic | 0.660 | Likely Pathogenic | Likely Benign | 4.03 | Destabilizing | 2.3 | 3.36 | Destabilizing | 3.70 | Destabilizing | 0.29 | Likely Benign | 0.440 | Likely Benign | -0.84 | Neutral | 0.498 | Possibly Damaging | 0.119 | Benign | 4.10 | Benign | 0.00 | Affected | 0.1295 | 0.3741 | -3 | -2 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||
| c.31G>A | G11R 2D AIThe SynGAP1 missense variant G11R is catalogued in gnomAD (ID 6‑33420295‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Pathogenic predictions come from polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.444081 | Structured | 0.501027 | Binding | 0.348 | 0.915 | 0.375 | 6-33420295-G-A | -3.418 | Likely Benign | 0.428 | Ambiguous | Likely Benign | 0.102 | Likely Benign | -0.47 | Neutral | 0.498 | Possibly Damaging | 0.026 | Benign | 3.92 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1022 | 0.4596 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||
| c.31G>C | G11R 2D AIThe SynGAP1 missense variant G11R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.444081 | Structured | 0.501027 | Binding | 0.348 | 0.915 | 0.375 | -3.418 | Likely Benign | 0.428 | Ambiguous | Likely Benign | 0.109 | Likely Benign | -0.47 | Neutral | 0.498 | Possibly Damaging | 0.026 | Benign | 3.92 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1022 | 0.4596 | -2 | -3 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||||
| c.2732T>A | V911D 2D AIThe SynGAP1 missense variant V911D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of high‑confidence predictors indicate a benign impact, and this is consistent with the lack of ClinVar evidence. Therefore, the variant is most likely benign and does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.685117 | Disordered | 0.724137 | Binding | 0.327 | 0.914 | 0.375 | -4.422 | Likely Benign | 0.624 | Likely Pathogenic | Likely Benign | 0.114 | Likely Benign | -0.41 | Neutral | 0.500 | Possibly Damaging | 0.157 | Benign | 2.71 | Benign | 0.05 | Affected | 0.1416 | 0.1289 | -2 | -3 | -7.7 | 15.96 | |||||||||||||||||||||||||||||||||||||||
| c.3368G>A | G1123D 2D AISynGAP1 missense variant G1123D is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33443920‑G‑A). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv and ESM1b. AlphaMissense‑Default remains uncertain, and Foldetta stability analysis is unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM Consensus also benign, while Foldetta provides no result. Overall, the preponderance of evidence points to a benign effect, which does not conflict with the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.837511 | Disordered | 0.827246 | Binding | 0.346 | 0.934 | 0.875 | Uncertain | 2 | 6-33443920-G-A | 2 | 1.33e-6 | -10.321 | Likely Pathogenic | 0.405 | Ambiguous | Likely Benign | 0.360 | Likely Benign | -0.78 | Neutral | 0.500 | Possibly Damaging | 0.157 | Benign | 4.34 | Benign | 0.19 | Tolerated | 3.77 | 5 | 0.1792 | 0.2035 | 1 | -1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||
| c.3395C>A | S1132Y 2D AIThe SynGAP1 missense variant S1132Y is listed in ClinVar as a benign alteration (ClinVar ID 845357.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence supports a benign classification, which aligns with the ClinVar status and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.845506 | Binding | 0.289 | 0.894 | 0.750 | Likely Benign | 1 | -5.894 | Likely Benign | 0.392 | Ambiguous | Likely Benign | 0.401 | Likely Benign | -1.76 | Neutral | 0.500 | Possibly Damaging | 0.208 | Benign | 5.40 | Benign | 0.09 | Tolerated | 4.32 | 4 | 0.0889 | 0.5233 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||
| c.2186A>T | N729I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N729I is listed in gnomAD (ID 6‑33441651‑A‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, premPS, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Tools that predict a pathogenic effect are PROVEAN and polyPhen‑2 HumDiv. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicting benign, while Foldetta’s stability analysis is inconclusive. Overall, the majority of evidence points to a benign impact. There is no ClinVar status to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.750527 | Disordered | 0.426547 | Uncertain | 0.651 | 0.583 | 0.625 | 6-33441651-A-T | 1 | 6.20e-7 | -3.308 | Likely Benign | 0.234 | Likely Benign | Likely Benign | 0.54 | Ambiguous | 0.6 | 0.79 | Ambiguous | 0.67 | Ambiguous | 0.29 | Likely Benign | 0.043 | Likely Benign | -2.96 | Deleterious | 0.506 | Possibly Damaging | 0.243 | Benign | 3.26 | Benign | 0.13 | Tolerated | 3.59 | 7 | 0.0625 | 0.4698 | -3 | -2 | 8.0 | -0.94 | |||||||||||||||||||||||||
| c.1381G>A | A461T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A461T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and ESM1b. Four tools (FoldX, Rosetta, Foldetta, premPS) returned uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta is also inconclusive. Overall, the balance of evidence favors a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.292531 | Uncertain | 0.936 | 0.151 | 0.125 | -10.885 | Likely Pathogenic | 0.323 | Likely Benign | Likely Benign | 1.21 | Ambiguous | 0.2 | 0.51 | Ambiguous | 0.86 | Ambiguous | 0.65 | Ambiguous | 0.214 | Likely Benign | -3.27 | Deleterious | 0.508 | Possibly Damaging | 0.042 | Benign | 3.40 | Benign | 0.13 | Tolerated | 0.1083 | 0.5930 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||
| c.1678G>C | V560L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V560L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Optimized, and Foldetta; pathogenic predictions come from polyPhen‑2 HumDiv, ESM1b, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized predicts benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, also predicts benign. Overall, the bulk of evidence points to a benign effect, but the SGM Consensus and the presence of pathogenic signals from several high‑confidence tools introduce uncertainty. Thus, the variant is most likely benign based on the prevailing predictions, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | -10.191 | Likely Pathogenic | 0.533 | Ambiguous | Likely Benign | -0.44 | Likely Benign | 0.0 | 0.48 | Likely Benign | 0.02 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -2.45 | Neutral | 0.508 | Possibly Damaging | 0.209 | Benign | -1.24 | Pathogenic | 0.40 | Tolerated | 3.37 | 35 | 0.1430 | 0.4162 | 1 | 2 | -0.4 | 14.03 | ||||||||||||||||||||||||||||
| c.1678G>T | V560L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V560L variant has no ClinVar entry (ClinVar status: None) but is catalogued in gnomAD (ID 6‑33440730‑G‑T). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are polyPhen‑2 HumDiv, ESM1b, and FATHMM (polyPhen‑2 HumVar is benign, AlphaMissense‑Default is uncertain). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning pathogenic (2 pathogenic vs 1 benign), and Foldetta indicating a benign stability change. Overall, the majority of conventional tools favor a benign classification, yet the high‑accuracy consensus and Foldetta suggest a pathogenic signal. Based on the most reliable predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | 6-33440730-G-T | 1 | 6.34e-7 | -10.191 | Likely Pathogenic | 0.533 | Ambiguous | Likely Benign | -0.44 | Likely Benign | 0.0 | 0.48 | Likely Benign | 0.02 | Likely Benign | 0.45 | Likely Benign | 0.489 | Likely Benign | -2.45 | Neutral | 0.508 | Possibly Damaging | 0.209 | Benign | -1.24 | Pathogenic | 0.40 | Tolerated | 3.37 | 35 | 0.1430 | 0.4162 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||
| c.2314T>C | F772L 2D AIThe SynGAP1 missense variant F772L is listed in gnomAD (6‑33442472‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are PolyPhen‑2 HumDiv, PolyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.450668 | Structured | 0.922662 | Binding | 0.329 | 0.884 | 0.250 | 6-33442472-T-C | 1 | 1.28e-6 | -1.751 | Likely Benign | 0.762 | Likely Pathogenic | Likely Benign | 0.161 | Likely Benign | -0.47 | Neutral | 0.508 | Possibly Damaging | 0.786 | Possibly Damaging | 4.31 | Benign | 1.00 | Tolerated | 3.64 | 6 | 0.1661 | 0.3033 | 0 | 2 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||
| c.2316C>A | F772L 2D AIThe SynGAP1 missense variant F772L is catalogued in gnomAD (ID 6‑33442474‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is Likely Benign, while Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the preponderance of evidence indicates that F772L is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.450668 | Structured | 0.922662 | Binding | 0.329 | 0.884 | 0.250 | 6-33442474-C-A | -1.751 | Likely Benign | 0.762 | Likely Pathogenic | Likely Benign | 0.109 | Likely Benign | -0.47 | Neutral | 0.508 | Possibly Damaging | 0.786 | Possibly Damaging | 4.31 | Benign | 1.00 | Tolerated | 3.64 | 6 | 0.1661 | 0.3033 | 0 | 2 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||
| c.2316C>G | F772L 2D AIThe SynGAP1 missense variant F772L has no ClinVar record and is not listed in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; no Foldetta stability data are available, so it is treated as unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the absence of a ClinVar classification, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.450668 | Structured | 0.922662 | Binding | 0.329 | 0.884 | 0.250 | -1.751 | Likely Benign | 0.762 | Likely Pathogenic | Likely Benign | 0.109 | Likely Benign | -0.47 | Neutral | 0.508 | Possibly Damaging | 0.786 | Possibly Damaging | 4.31 | Benign | 1.00 | Tolerated | 3.64 | 6 | 0.1661 | 0.3033 | 0 | 2 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||||
| c.2611C>T | H871Y 2D AIThe SynGAP1 missense variant H871Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.679301 | Binding | 0.279 | 0.858 | 0.250 | -4.070 | Likely Benign | 0.202 | Likely Benign | Likely Benign | 0.089 | Likely Benign | -1.44 | Neutral | 0.510 | Possibly Damaging | 0.206 | Benign | 2.63 | Benign | 0.08 | Tolerated | 0.0872 | 0.3945 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||||||
| c.2612A>C | H871P 2D AIThe SynGAP1 missense variant H871P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign and the SGM‑Consensus also reports likely benign; Foldetta results are unavailable. Overall, the consensus of the available predictions points to a benign impact, and this is consistent with the lack of ClinVar evidence; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.679301 | Binding | 0.279 | 0.858 | 0.250 | -3.420 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.207 | Likely Benign | -0.91 | Neutral | 0.510 | Possibly Damaging | 0.206 | Benign | 2.63 | Benign | 0.18 | Tolerated | 0.2163 | 0.3757 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||||||||||||
| c.2869C>T | H957Y 2D AIThe SynGAP1 missense variant H957Y is listed in gnomAD (ID 6‑33443421‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which reports “Likely Benign.” Pathogenic predictions come from polyPhen‑2 HumDiv and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.968874 | Binding | 0.362 | 0.915 | 0.750 | 6-33443421-C-T | 1 | 6.20e-7 | -6.631 | Likely Benign | 0.129 | Likely Benign | Likely Benign | 0.099 | Likely Benign | -1.00 | Neutral | 0.510 | Possibly Damaging | 0.147 | Benign | 2.42 | Pathogenic | 0.10 | Tolerated | 3.77 | 5 | 0.1560 | 0.4906 | 2 | 0 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||||||
| c.2875C>T | H959Y 2D AIThe SynGAP1 missense variant H959Y is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a tolerated change, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also classifies it as Likely Benign. In contrast, polyPhen‑2 HumDiv, SIFT, and ESM1b predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect for H959Y, and this conclusion does not conflict with ClinVar, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.985964 | Disordered | 0.980566 | Binding | 0.333 | 0.905 | 0.750 | -8.367 | Likely Pathogenic | 0.146 | Likely Benign | Likely Benign | 0.140 | Likely Benign | -1.09 | Neutral | 0.510 | Possibly Damaging | 0.147 | Benign | 4.09 | Benign | 0.05 | Affected | 0.1672 | 0.4906 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||||||
| c.652T>A | F218I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F218I is reported in gnomAD (variant ID 6‑33435294‑T‑A) but has no ClinVar entry. Functional prediction tools largely agree on a deleterious effect: pathogenic predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, while benign predictions are limited to polyPhen‑2 (HumVar), SIFT, and FATHMM. High‑accuracy assessments reinforce the pathogenic view: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No prediction or folding stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.281712 | Structured | 0.408725 | Uncertain | 0.848 | 0.272 | 0.000 | 6-33435294-T-A | 1 | 6.20e-7 | -12.211 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 4.69 | Destabilizing | 0.2 | 5.93 | Destabilizing | 5.31 | Destabilizing | 1.18 | Destabilizing | 0.612 | Likely Pathogenic | -3.81 | Deleterious | 0.510 | Possibly Damaging | 0.066 | Benign | 5.85 | Benign | 0.08 | Tolerated | 3.41 | 13 | 0.2106 | 0.2459 | 0 | 1 | 1.7 | -34.02 | ||||||||||||||||||||||||
| c.1702G>C | V568L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V568L variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include FoldX, Foldetta, and polyPhen‑2 HumVar, while those that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (which itself is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Uncertain predictions come from Rosetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools predict a pathogenic impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.024826 | Structured | 0.053503 | Uncertain | 0.937 | 0.257 | 0.000 | -9.503 | Likely Pathogenic | 0.921 | Likely Pathogenic | Ambiguous | -0.30 | Likely Benign | 0.3 | 0.57 | Ambiguous | 0.14 | Likely Benign | 0.56 | Ambiguous | 0.651 | Likely Pathogenic | -2.69 | Deleterious | 0.511 | Possibly Damaging | 0.147 | Benign | -1.23 | Pathogenic | 0.04 | Affected | 3.37 | 35 | 0.0944 | 0.3312 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||||
| c.1702G>T | V568L 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V568L is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Among the available in‑silico predictors, eight tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus) predict a pathogenic effect, whereas three tools (FoldX, Foldetta, and polyPhen‑2 HumVar) predict a benign outcome; the remaining three (Rosetta, premPS, AlphaMissense‑Optimized) are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta stability outputs) as benign. Overall, the preponderance of evidence points to a pathogenic impact, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.024826 | Structured | 0.053503 | Uncertain | 0.937 | 0.257 | 0.000 | Uncertain | 1 | -9.503 | Likely Pathogenic | 0.921 | Likely Pathogenic | Ambiguous | -0.30 | Likely Benign | 0.3 | 0.57 | Ambiguous | 0.14 | Likely Benign | 0.56 | Ambiguous | 0.651 | Likely Pathogenic | -2.69 | Deleterious | 0.511 | Possibly Damaging | 0.147 | Benign | -1.23 | Pathogenic | 0.04 | Affected | 3.37 | 35 | 0.0944 | 0.3312 | 1 | 2 | -0.4 | 14.03 | |||||||||||||||||||||||||
| c.3307C>A | R1103S 2D AIThe SynGAP1 missense variant R1103S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta results are unavailable. Overall, the majority of evidence (six benign versus two pathogenic predictions) indicates that R1103S is most likely benign, and this conclusion does not contradict any ClinVar annotation because no ClinVar claim exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.945666 | Disordered | 0.957363 | Binding | 0.328 | 0.862 | 0.875 | -3.611 | Likely Benign | 0.464 | Ambiguous | Likely Benign | 0.111 | Likely Benign | -1.94 | Neutral | 0.511 | Possibly Damaging | 0.187 | Benign | 2.48 | Pathogenic | 0.28 | Tolerated | 0.3025 | 0.4224 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.1505G>C | G502A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G502A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from Foldetta, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized; pathogenic predictions arise from SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain results are reported by FoldX, Rosetta, and premPS. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized indicates a benign effect, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—concludes pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, also predicts a benign outcome. Overall, the majority of evidence points to a pathogenic impact for G502A, and this assessment does not conflict with the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.083462 | Structured | 0.340113 | Uncertain | 0.882 | 0.152 | 0.000 | -10.191 | Likely Pathogenic | 0.607 | Likely Pathogenic | Likely Benign | 0.82 | Ambiguous | 0.4 | -0.53 | Ambiguous | 0.15 | Likely Benign | 0.54 | Ambiguous | 0.725 | Likely Pathogenic | -5.64 | Deleterious | 0.512 | Possibly Damaging | 0.157 | Benign | -1.59 | Pathogenic | 0.17 | Tolerated | 0.3548 | 0.3393 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||
| c.3345T>G | I1115M 2D AIThe SynGAP1 missense variant I1115M is reported in gnomAD (variant ID 6‑33443897‑T‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant call. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.889439 | Disordered | 0.892339 | Binding | 0.308 | 0.912 | 0.750 | 6-33443897-T-G | 4 | 2.77e-6 | -3.708 | Likely Benign | 0.067 | Likely Benign | Likely Benign | 0.106 | Likely Benign | -0.38 | Neutral | 0.512 | Possibly Damaging | 0.200 | Benign | 2.71 | Benign | 0.23 | Tolerated | 4.32 | 2 | 0.1030 | 0.4162 | 1 | 2 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||||||
| c.3630C>A | H1210Q 2D AIThe SynGAP1 missense variant H1210Q is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as “Likely Benign.” Only polyPhen‑2 HumDiv predicts a pathogenic outcome. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also indicates a benign likelihood. Foldetta results are unavailable, so they do not influence the assessment. Overall, the consensus of the available predictions indicates that H1210Q is most likely benign, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.587579 | Binding | 0.900 | 0.567 | 0.375 | -1.917 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.060 | Likely Benign | -0.83 | Neutral | 0.512 | Possibly Damaging | 0.223 | Benign | 2.74 | Benign | 0.09 | Tolerated | 0.1133 | 0.3092 | 3 | 0 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3630C>G | H1210Q 2D AIThe SynGAP1 missense variant H1210Q is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.587579 | Binding | 0.900 | 0.567 | 0.375 | -1.917 | Likely Benign | 0.142 | Likely Benign | Likely Benign | 0.060 | Likely Benign | -0.83 | Neutral | 0.512 | Possibly Damaging | 0.223 | Benign | 2.74 | Benign | 0.09 | Tolerated | 0.1133 | 0.3092 | 3 | 0 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||||
| c.1322T>C | V441A 2D 3DClick to see structure in 3D Viewer AISynGAP1 variant V441A is listed in ClinVar as uncertain and is present in gnomAD (ID 6‑33438227‑T‑C). Consensus from most in silico predictors favors a benign effect: REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized all report benign. Pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, and ESM1b, while premPS and AlphaMissense‑Default remain uncertain. High‑accuracy assessments give a mixed picture: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports benign. Overall, the preponderance of evidence points to a benign impact, aligning with the ClinVar uncertain designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | Conflicting | 2 | 6-33438227-T-C | 3 | 1.86e-6 | -9.439 | Likely Pathogenic | 0.359 | Ambiguous | Likely Benign | -0.14 | Likely Benign | 0.0 | 0.33 | Likely Benign | 0.10 | Likely Benign | 0.95 | Ambiguous | 0.053 | Likely Benign | -2.92 | Deleterious | 0.513 | Possibly Damaging | 0.214 | Benign | 3.44 | Benign | 0.93 | Tolerated | 3.37 | 29 | 0.2390 | 0.1800 | 0 | 0 | -2.4 | -28.05 | 195.0 | 44.6 | 0.0 | 0.1 | 0.5 | 0.0 | X | X | Uncertain | The iso-propyl side chain of Val441, located on the outer surface of an α helix (res. Asn440-Thr458), does not interact with other residues in the WT simulations. In the variant simulations, the methyl side chain of Ala441 is similarly hydrophobic and does not form any interactions on the outer helix surface. Although the residue swap does not negatively affect the protein structure based on the simulations, it is noteworthy that the residue faces the RasGTPase interface. Thus, the effect of the residue swap on the SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | |||||||||||||
| c.2251C>T | P751S 2D AIThe SynGAP1 missense variant P751S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also predicts Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of tools and the high‑accuracy predictions indicate that P751S is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.667683 | Binding | 0.386 | 0.866 | 0.625 | -4.157 | Likely Benign | 0.109 | Likely Benign | Likely Benign | 0.109 | Likely Benign | -0.80 | Neutral | 0.514 | Possibly Damaging | 0.216 | Benign | 2.70 | Benign | 0.33 | Tolerated | 0.3446 | 0.5837 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3145C>A | P1049T 2D AIThe SynGAP1 missense variant P1049T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta results are unavailable, so no additional stability evidence is present. Overall, the consensus of available predictions indicates that P1049T is most likely benign, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.917915 | Binding | 0.428 | 0.920 | 0.750 | -5.100 | Likely Benign | 0.056 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -1.18 | Neutral | 0.519 | Possibly Damaging | 0.222 | Benign | 2.74 | Benign | 0.02 | Affected | 0.1791 | 0.5514 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3145C>T | P1049S 2D AIThe SynGAP1 missense variant P1049S is reported in gnomAD (variant ID 6‑33443697‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. No Foldetta stability data are available, so it does not influence the assessment. Overall, the majority of predictions, including the high‑accuracy tools, indicate that P1049S is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.917915 | Binding | 0.428 | 0.920 | 0.750 | 6-33443697-C-T | 2 | 1.24e-6 | -2.351 | Likely Benign | 0.053 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -1.53 | Neutral | 0.519 | Possibly Damaging | 0.303 | Benign | 2.76 | Benign | 0.04 | Affected | 3.77 | 5 | 0.3144 | 0.5083 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.1051G>A | A351T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A351T missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that uniformly indicate a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. Uncertain predictions come from Rosetta and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely benign, and Foldetta as inconclusive. Overall, the majority of evidence supports a benign impact for A351T, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.216401 | Structured | 0.362025 | Uncertain | 0.925 | 0.342 | 0.000 | -6.863 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.22 | Likely Benign | 0.0 | 0.80 | Ambiguous | 0.51 | Ambiguous | 0.49 | Likely Benign | 0.040 | Likely Benign | -2.42 | Neutral | 0.524 | Possibly Damaging | 0.138 | Benign | 1.71 | Pathogenic | 0.06 | Tolerated | 0.1337 | 0.6713 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||
| c.2224C>G | R742G 2D AIThe SynGAP1 missense variant R742G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.871313 | Disordered | 0.509587 | Binding | 0.309 | 0.856 | 0.875 | -4.065 | Likely Benign | 0.101 | Likely Benign | Likely Benign | 0.067 | Likely Benign | -1.25 | Neutral | 0.524 | Possibly Damaging | 0.259 | Benign | 2.70 | Benign | 0.02 | Affected | 0.3974 | 0.2805 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.2111G>C | S704T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S704T is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Consensus from multiple in‑silico predictors shows a predominance of benign calls: REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus. Only polyPhen‑2 HumDiv predicts a pathogenic effect, while FoldX remains inconclusive. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts benign. Overall, the aggregate evidence indicates that S704T is most likely benign, which is consistent with its ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.096677 | Structured | 0.383620 | Uncertain | 0.928 | 0.363 | 0.000 | Uncertain | 1 | -4.930 | Likely Benign | 0.265 | Likely Benign | Likely Benign | 0.80 | Ambiguous | 0.0 | 0.15 | Likely Benign | 0.48 | Likely Benign | 0.29 | Likely Benign | 0.071 | Likely Benign | -1.72 | Neutral | 0.525 | Possibly Damaging | 0.107 | Benign | 3.45 | Benign | 0.07 | Tolerated | 3.47 | 10 | 0.1036 | 0.4674 | 1 | 1 | 0.1 | 14.03 | 201.7 | -18.0 | 0.0 | 0.0 | -0.2 | 0.7 | X | Potentially Benign | Ser704 is located at the end and outer surface of an α-helix (res. Thr704-Gly712), which is connected via a tight turn or loop to another α-helix (res. Asp684-Gln702). The hydroxyl side chain of Ser704 occasionally forms a hydrogen bond with the amide group of Ala707. Similarly, in the variant simulations, the hydroxyl side chain of Thr704 forms hydrogen bonds with the amide groups of Ala707 and Leu708. Thus, the residue swap does not cause any apparent structural change. | ||||||||||||||||
| c.1609G>T | A537S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A537S is reported in gnomAD (6-33438852-G-T) and has no ClinVar entry. Consensus from multiple in silico predictors indicates a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all score benign, while polyPhen‑2 (HumDiv and HumVar) and FATHMM predict pathogenicity. When predictions are grouped, the majority of tools (ten) support benign, whereas three tools support pathogenic. High‑accuracy assessments further reinforce the benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts benign. Consequently, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.116183 | Structured | 0.037313 | Uncertain | 0.928 | 0.362 | 0.000 | 6-33438852-G-T | 1 | 6.20e-7 | -3.602 | Likely Benign | 0.095 | Likely Benign | Likely Benign | 0.28 | Likely Benign | 0.0 | 0.46 | Likely Benign | 0.37 | Likely Benign | 0.28 | Likely Benign | 0.206 | Likely Benign | -0.66 | Neutral | 0.528 | Possibly Damaging | 0.592 | Possibly Damaging | -1.25 | Pathogenic | 0.60 | Tolerated | 3.37 | 35 | 0.2729 | 0.4431 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||
| c.1804A>G | I602V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I602V missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the change as benign include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS, polyPhen‑2 HumDiv, SIFT, and FATHMM. Uncertain or inconclusive results come from FoldX, Rosetta, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign, while Foldetta’s stability analysis remains uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.010221 | Structured | 0.186541 | Uncertain | 0.963 | 0.171 | 0.000 | -6.317 | Likely Benign | 0.075 | Likely Benign | Likely Benign | 1.24 | Ambiguous | 0.0 | 0.78 | Ambiguous | 1.01 | Ambiguous | 1.01 | Destabilizing | 0.309 | Likely Benign | -0.84 | Neutral | 0.528 | Possibly Damaging | 0.179 | Benign | -1.89 | Pathogenic | 0.03 | Affected | 0.1173 | 0.3326 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||
| c.3918C>A | N1306K 2D AIThe SynGAP1 missense variant N1306K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -3.021 | Likely Benign | 0.585 | Likely Pathogenic | Likely Benign | 0.140 | Likely Benign | -4.00 | Deleterious | 0.532 | Possibly Damaging | 0.327 | Benign | 2.61 | Benign | 0.00 | Affected | 0.2375 | 0.6234 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.3918C>G | N1306K 2D AIThe SynGAP1 missense variant N1306K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -3.021 | Likely Benign | 0.585 | Likely Pathogenic | Likely Benign | 0.177 | Likely Benign | -4.00 | Deleterious | 0.532 | Possibly Damaging | 0.327 | Benign | 2.61 | Benign | 0.00 | Affected | 0.2375 | 0.6234 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.494G>A | S165N 2D AIThe SynGAP1 missense variant S165N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and ESM1b. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for S165N. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -9.041 | Likely Pathogenic | 0.509 | Ambiguous | Likely Benign | 0.074 | Likely Benign | -0.49 | Neutral | 0.532 | Possibly Damaging | 0.229 | Benign | 4.03 | Benign | 0.00 | Affected | 0.1589 | 0.4862 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||||||||
Found 8751 rows. Show 800 rows per page. Page 3/11 « Previous | Next »