
Table of SynGAP1 Isoform α2 (UniProt Q96PV0-1) Missense Variants.
| c.dna | Variant | SGM Consensus | Domain and Structure information: based on WT protein | Annotated databases | Deep learning-based pathogenicity predictions | Folding stability-based pathogenicity predictions | Sequence/structure-based pathogenicity predictions | Phase Separation | Evolutionary/physical properties | Molecular Dynamics-based analysis | DOI | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Domain | IUPred2 | ANCHOR2 | AlphaFold | MobiDB | PhosphoSitePlus | ClinVar | gnomAD | ESM1b | AlphaMissense | FoldX | Rosetta | Foldetta | PremPS | REVEL | PROVEAN | PolyPhen-2 HumDiv | PolyPhen-2 HumVar | FATHMM | SIFT | PSMutPred | PAM | Physical | SASA | Normalized B-factor backbone | Normalized B-factor sidechain | SynGAP Structural Annotation | |||||||||||||||||||||||||||||||||||||||||||||
| Score | Prediction | Score | Prediction | pLDDT | disorder | disorder | LTP | HTP | KL | PTM | Clinical Status | Review | Subm. | ID | Allele count | Allele freq. | LLR score | Prediction | Pathogenicity | Class | Optimized | Average ΔΔG | Prediction | StdDev | ΔΔG | Prediction | ΔΔG | Prediction | ΔΔG | Prediction | Score | Prediction | Score | Prediction | pph2_prob | Prediction | pph2_prob | Prediction | Nervous System Score | Prediction | Prediction | Status | Conservation | Sequences | IP RF | SP RF | Prediction | PAM250 | PAM120 | Hydropathy Δ | MW Δ | Average | Δ | Δ | StdDev | Δ | StdDev | Secondary | Tertiary bonds | Inside out | GAP-Ras interface | At membrane | No effect | MD Alert | Verdict | Description | |||||
| c.3391C>G | P1131A 2D  AIThe SynGAP1 missense variant P1131A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | -4.785 | Likely Benign | 0.171 | Likely Benign | Likely Benign | 0.207 | Likely Benign | -2.88 | Deleterious | 0.009 | Benign | 0.008 | Benign | 5.36 | Benign | 0.00 | Affected | 0.2977 | 0.5501 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3391C>T | P1131S 2D  AIThe SynGAP1 missense variant P1131S is reported in gnomAD (variant ID 6‑33443943‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign, and this is not contradictory to ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | 6-33443943-C-T | 1 | 6.72e-7 | -4.089 | Likely Benign | 0.246 | Likely Benign | Likely Benign | 0.209 | Likely Benign | -2.62 | Deleterious | 0.025 | Benign | 0.015 | Benign | 5.54 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2936 | 0.5936 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3392C>A | P1131H 2D  AIThe SynGAP1 missense variant P1131H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the balance of evidence leans toward a benign classification, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | -5.207 | Likely Benign | 0.454 | Ambiguous | Likely Benign | 0.357 | Likely Benign | -3.50 | Deleterious | 0.971 | Probably Damaging | 0.750 | Possibly Damaging | 5.24 | Benign | 0.00 | Affected | 0.1519 | 0.5546 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3392C>G | P1131R 2D  AIThe SynGAP1 P1131R missense variant has no ClinVar record and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta data are unavailable. With five benign versus four pathogenic calls and a benign result from the most accurate tool, the variant is most likely benign. This assessment does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | -3.792 | Likely Benign | 0.702 | Likely Pathogenic | Likely Benign | 0.416 | Likely Benign | -2.58 | Deleterious | 0.918 | Possibly Damaging | 0.420 | Benign | 5.26 | Benign | 0.00 | Affected | 0.1252 | 0.4010 | 0 | -2 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||||||||||||
| c.3392C>T | P1131L 2D  AIThe SynGAP1 missense variant P1131L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN and SIFT, while AlphaMissense‑Default remains uncertain. The high‑accuracy AlphaMissense‑Optimized tool classifies the variant as benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also yields a benign prediction (2 benign vs. 1 pathogenic, with one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | -5.267 | Likely Benign | 0.420 | Ambiguous | Likely Benign | 0.293 | Likely Benign | -3.62 | Deleterious | 0.002 | Benign | 0.005 | Benign | 5.26 | Benign | 0.00 | Affected | 0.2043 | 0.6998 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3409C>A | H1137N 2D  AIThe SynGAP1 missense variant H1137N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | -3.105 | Likely Benign | 0.063 | Likely Benign | Likely Benign | 0.208 | Likely Benign | -1.53 | Neutral | 0.625 | Possibly Damaging | 0.353 | Benign | 5.34 | Benign | 0.00 | Affected | 0.1983 | 0.3638 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.3409C>G | H1137D 2D  AIThe SynGAP1 missense variant H1137D is not reported in ClinVar and has no allele in gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Pathogenicity is suggested only by polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for H1137D, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | -4.934 | Likely Benign | 0.343 | Ambiguous | Likely Benign | 0.418 | Likely Benign | -2.26 | Neutral | 0.802 | Possibly Damaging | 0.430 | Benign | 5.56 | Benign | 0.00 | Affected | 0.2519 | 0.2916 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.3409C>T | H1137Y 2D  AIThe SynGAP1 missense variant H1137Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for H1137Y, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | -4.506 | Likely Benign | 0.124 | Likely Benign | Likely Benign | 0.310 | Likely Benign | -1.93 | Neutral | 0.925 | Possibly Damaging | 0.629 | Possibly Damaging | 5.28 | Benign | 0.00 | Affected | 0.1073 | 0.5123 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||||||||||||
| c.340A>C | K114Q 2D  AIThe SynGAP1 missense variant K114Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized all predict benign. Pathogenic predictions come from polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is Likely Benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and there is no ClinVar annotation to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -3.221 | Likely Benign | 0.467 | Ambiguous | Likely Benign | 0.058 | Likely Benign | -1.33 | Neutral | 0.608 | Possibly Damaging | 0.108 | Benign | 3.98 | Benign | 0.00 | Affected | 0.5476 | 0.1530 | Weaken | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||||||||||
| c.340A>G | K114E 2D  AISynGAP1 missense variant K114E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default, while AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation, as none exists for K114E. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -2.648 | Likely Benign | 0.885 | Likely Pathogenic | Ambiguous | 0.093 | Likely Benign | -1.27 | Neutral | 0.005 | Benign | 0.003 | Benign | 4.01 | Benign | 0.00 | Affected | 0.4815 | 0.1340 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3410A>C | H1137P 2D  AIThe SynGAP1 missense variant H1137P is listed in ClinVar as a benign alteration (ClinVar ID 3685596.0) and is present in the gnomAD database (gnomAD ID 6‑33444445‑A‑C). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (derived from the same four high‑accuracy predictors) also indicates benign. No Foldetta (FoldX‑MD/Rosetta stability) result is available for this variant. Overall, the majority of predictions, including the most reliable tools, support a benign classification, which is consistent with the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | Benign | 1 | 6-33444445-A-C | 12 | 7.44e-6 | -2.098 | Likely Benign | 0.054 | Likely Benign | Likely Benign | 0.419 | Likely Benign | -1.93 | Neutral | 0.925 | Possibly Damaging | 0.703 | Possibly Damaging | 5.29 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2045 | 0.4501 | -2 | 0 | 1.6 | -40.02 | ||||||||||||||||||||||||||||||||
| c.3410A>G | H1137R 2D  AIThe SynGAP1 missense variant H1137R is reported in gnomAD (ID 6‑33444445‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | 6-33444445-A-G | 1 | 6.20e-7 | -3.468 | Likely Benign | 0.228 | Likely Benign | Likely Benign | 0.296 | Likely Benign | -1.19 | Neutral | 0.925 | Possibly Damaging | 0.629 | Possibly Damaging | 5.32 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2131 | 0.3449 | 0 | 2 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||
| c.3410A>T | H1137L 2D  AIThe SynGAP1 missense variant H1137L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Based on the preponderance of benign predictions and the consensus from high‑accuracy methods, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | -2.215 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.359 | Likely Benign | -2.77 | Deleterious | 0.802 | Possibly Damaging | 0.534 | Possibly Damaging | 5.30 | Benign | 0.00 | Affected | 0.1199 | 0.6082 | -2 | -3 | 7.0 | -23.98 | |||||||||||||||||||||||||||||||||||||||
| c.3411T>A | H1137Q 2D  AIThe SynGAP1 missense variant H1137Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, while polyPhen‑2 (HumDiv and HumVar) and SIFT predict pathogenic. Grouping by consensus, the benign‑predicting tools outnumber the pathogenic ones. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized scores benign, and the SGM Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | -2.958 | Likely Benign | 0.110 | Likely Benign | Likely Benign | 0.239 | Likely Benign | -1.19 | Neutral | 0.925 | Possibly Damaging | 0.703 | Possibly Damaging | 5.34 | Benign | 0.00 | Affected | 0.1873 | 0.4090 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.3411T>G | H1137Q 2D AIThe SynGAP1 missense variant H1137Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for H1137Q, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.756488 | Binding | 0.314 | 0.879 | 0.875 | -2.958 | Likely Benign | 0.110 | Likely Benign | Likely Benign | 0.239 | Likely Benign | -1.19 | Neutral | 0.925 | Possibly Damaging | 0.703 | Possibly Damaging | 5.34 | Benign | 0.00 | Affected | 0.1873 | 0.4090 | 3 | 0 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||||
| c.3415C>A | Q1139K 2D  AIThe SynGAP1 missense variant Q1139K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -4.768 | Likely Benign | 0.222 | Likely Benign | Likely Benign | 0.253 | Likely Benign | -1.70 | Neutral | 0.004 | Benign | 0.006 | Benign | 5.44 | Benign | 0.00 | Affected | 0.1667 | 0.4139 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||||
| c.3415C>G | Q1139E 2D  AIThe SynGAP1 missense variant Q1139E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign or likely benign outcome, whereas only SIFT classifies it as pathogenic. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the consensus of the available predictions points to a benign effect, and this is consistent with the lack of ClinVar evidence for pathogenicity. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -3.065 | Likely Benign | 0.204 | Likely Benign | Likely Benign | 0.285 | Likely Benign | -1.34 | Neutral | 0.112 | Benign | 0.089 | Benign | 5.36 | Benign | 0.00 | Affected | 0.1360 | 0.2269 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3416A>C | Q1139P 2D  AIThe SynGAP1 missense variant Q1139P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -3.753 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.437 | Likely Benign | -1.74 | Neutral | 0.812 | Possibly Damaging | 0.396 | Benign | 5.28 | Benign | 0.00 | Affected | 0.2199 | 0.5349 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||||||||||||||
| c.3416A>G | Q1139R 2D  AIThe SynGAP1 missense variant Q1139R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of ClinVar annotation—there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -4.249 | Likely Benign | 0.267 | Likely Benign | Likely Benign | 0.315 | Likely Benign | -1.91 | Neutral | 0.126 | Benign | 0.138 | Benign | 5.47 | Benign | 0.00 | Affected | 0.1342 | 0.2405 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3416A>T | Q1139L 2D  AIThe SynGAP1 missense variant Q1139L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -1.689 | Likely Benign | 0.153 | Likely Benign | Likely Benign | 0.472 | Likely Benign | -3.75 | Deleterious | 0.224 | Benign | 0.237 | Benign | 5.30 | Benign | 0.00 | Affected | 0.0745 | 0.5599 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3417G>C | Q1139H 2D  AIThe SynGAP1 missense variant Q1139H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -3.962 | Likely Benign | 0.125 | Likely Benign | Likely Benign | 0.265 | Likely Benign | -2.40 | Neutral | 0.002 | Benign | 0.007 | Benign | 5.41 | Benign | 0.00 | Affected | 0.1313 | 0.3801 | 3 | 0 | 0.3 | 9.01 | |||||||||||||||||||||||||||||||||||||||
| c.3417G>T | Q1139H 2D AIThe SynGAP1 missense variant Q1139H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of benign predictions and the lack of contradictory evidence, the variant is most likely benign. This assessment does not conflict with ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.721191 | Binding | 0.313 | 0.866 | 1.000 | -3.962 | Likely Benign | 0.125 | Likely Benign | Likely Benign | 0.265 | Likely Benign | -2.40 | Neutral | 0.002 | Benign | 0.007 | Benign | 5.41 | Benign | 0.00 | Affected | 0.1313 | 0.3801 | 3 | 0 | 0.3 | 9.01 | |||||||||||||||||||||||||||||||||||||||
| c.341A>C | K114T 2D 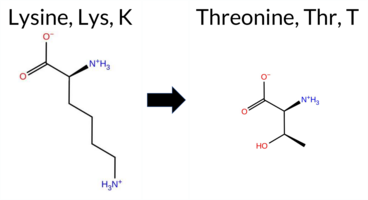 AIThe SynGAP1 missense variant K114T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), which collectively indicate a likely benign outcome. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, the SGM‑Consensus score is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the overall distribution of predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -3.366 | Likely Benign | 0.804 | Likely Pathogenic | Ambiguous | 0.091 | Likely Benign | -1.48 | Neutral | 0.759 | Possibly Damaging | 0.190 | Benign | 3.95 | Benign | 0.00 | Affected | 0.2796 | 0.3391 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.341A>G | K114R 2D 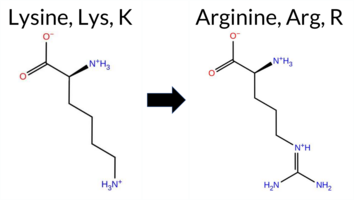 AIThe SynGAP1 missense variant K114R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect; there is no conflict with ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -1.861 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.02 | Neutral | 0.005 | Benign | 0.003 | Benign | 4.09 | Benign | 0.00 | Affected | 0.5668 | 0.1498 | Weaken | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.341A>T | K114M 2D 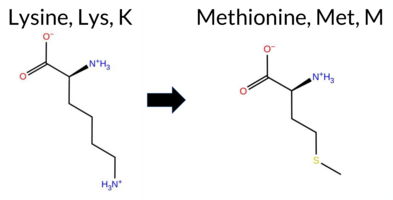 AIThe SynGAP1 missense variant K114M is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is unavailable for this variant. Overall, the majority of evidence points toward a benign effect, and there is no ClinVar entry to contradict this conclusion. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -3.953 | Likely Benign | 0.877 | Likely Pathogenic | Ambiguous | 0.120 | Likely Benign | -1.89 | Neutral | 0.992 | Probably Damaging | 0.615 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.1809 | 0.4075 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||||||||||||
| c.3421C>A | P1141T 2D 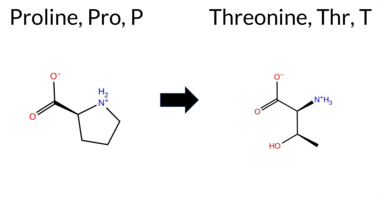 AIThe SynGAP1 missense variant P1141T is not reported in ClinVar (ClinVar status: not reported) and is absent from gnomAD (gnomAD: not present). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic). High‑accuracy methods give AlphaMissense‑Optimized a benign prediction; the SGM Consensus remains inconclusive, and Foldetta (combining FoldX‑MD and Rosetta) has no available result. Overall, the balance of evidence, particularly the benign call from the high‑accuracy AlphaMissense‑Optimized model, suggests that the variant is most likely benign, and this assessment does not contradict the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.716087 | Binding | 0.364 | 0.852 | 1.000 | -4.090 | Likely Benign | 0.176 | Likely Benign | Likely Benign | 0.106 | Likely Benign | -3.85 | Deleterious | 0.856 | Possibly Damaging | 0.723 | Possibly Damaging | 0.98 | Pathogenic | 0.00 | Affected | 0.1594 | 0.6145 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.3421C>G | P1141A 2D AIThe SynGAP1 missense variant P1141A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding‑stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools therefore provide mixed evidence: AlphaMissense‑Optimized indicates benign, while the consensus and stability analyses are unavailable. Overall, the majority of individual predictors (five versus four) favor a pathogenic interpretation, and this does not contradict any ClinVar annotation because none exists. Thus, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.716087 | Binding | 0.364 | 0.852 | 1.000 | -3.885 | Likely Benign | 0.120 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -3.67 | Deleterious | 0.856 | Possibly Damaging | 0.492 | Possibly Damaging | 1.00 | Pathogenic | 0.00 | Affected | 0.3525 | 0.5135 | 1 | -1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3421C>T | P1141S 2D AIThe SynGAP1 missense variant P1141S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, five of the nine evaluated tools predict pathogenicity while four predict benignity, giving a slight majority toward a deleterious effect. Thus, the variant is most likely pathogenic based on current computational predictions, and this assessment does not contradict any ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.716087 | Binding | 0.364 | 0.852 | 1.000 | -2.574 | Likely Benign | 0.187 | Likely Benign | Likely Benign | 0.094 | Likely Benign | -3.58 | Deleterious | 0.856 | Possibly Damaging | 0.492 | Possibly Damaging | 0.99 | Pathogenic | 0.00 | Affected | 0.3379 | 0.5566 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3422C>A | P1141Q 2D  AIThe SynGAP1 missense variant P1141Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.716087 | Binding | 0.364 | 0.852 | 1.000 | -4.331 | Likely Benign | 0.274 | Likely Benign | Likely Benign | 0.136 | Likely Benign | -2.72 | Deleterious | 0.074 | Benign | 0.047 | Benign | 0.97 | Pathogenic | 0.00 | Affected | 0.1449 | 0.5186 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||||||
| c.3422C>G | P1141R 2D AIThe SynGAP1 missense variant P1141R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus indicates a likely pathogenic outcome; a Foldetta stability analysis is unavailable. Overall, the majority of computational predictions (seven pathogenic vs. three benign) support a pathogenic classification. This consensus does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.930790 | Disordered | 0.716087 | Binding | 0.364 | 0.852 | 1.000 | -4.768 | Likely Benign | 0.626 | Likely Pathogenic | Likely Benign | 0.120 | Likely Benign | -3.90 | Deleterious | 0.913 | Possibly Damaging | 0.690 | Possibly Damaging | 0.97 | Pathogenic | 0.00 | Affected | 0.1329 | 0.3614 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3422C>T | P1141L 2D AIThe SynGAP1 missense variant P1141L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (two pathogenic versus one benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy tools indicates that P1141L is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.716087 | Binding | 0.364 | 0.852 | 1.000 | -3.817 | Likely Benign | 0.352 | Ambiguous | Likely Benign | 0.110 | Likely Benign | -4.64 | Deleterious | 0.954 | Possibly Damaging | 0.759 | Possibly Damaging | 0.98 | Pathogenic | 0.00 | Affected | 0.2147 | 0.6099 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3424T>A | S1142T 2D  AIThe SynGAP1 missense variant S1142T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | -3.712 | Likely Benign | 0.100 | Likely Benign | Likely Benign | 0.106 | Likely Benign | -1.63 | Neutral | 0.611 | Possibly Damaging | 0.324 | Benign | 2.69 | Benign | 0.00 | Affected | 0.1548 | 0.6427 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3424T>C | S1142P 2D  AIThe SynGAP1 missense variant S1142P is listed in ClinVar (ID 2747352.0) as Benign and is present in gnomAD (variant ID 6‑33444459‑T‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, aligning with the ClinVar classification and not contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | Likely Benign | 1 | 6-33444459-T-C | 1 | 6.20e-7 | -2.713 | Likely Benign | 0.222 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -2.19 | Neutral | 0.918 | Possibly Damaging | 0.761 | Possibly Damaging | 2.64 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2080 | 0.5618 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||
| c.3424T>G | S1142A 2D  AIThe SynGAP1 missense variant S1142A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) confirms a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that S1142A is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | -3.694 | Likely Benign | 0.085 | Likely Benign | Likely Benign | 0.074 | Likely Benign | -1.31 | Neutral | 0.002 | Benign | 0.015 | Benign | 2.73 | Benign | 0.00 | Affected | 0.4689 | 0.5319 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.3425C>A | S1142Y 2D 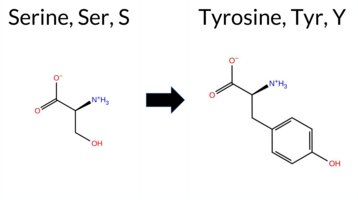 AIThe SynGAP1 missense variant S1142Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of conventional predictors lean toward pathogenicity, but the single high‑accuracy benign prediction and the inconclusive consensus temper this view. Thus, the variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | -6.146 | Likely Benign | 0.569 | Likely Pathogenic | Likely Benign | 0.200 | Likely Benign | -3.39 | Deleterious | 0.971 | Probably Damaging | 0.876 | Possibly Damaging | 2.64 | Benign | 0.00 | Affected | 0.0849 | 0.5429 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||||
| c.3425C>G | S1142C 2D 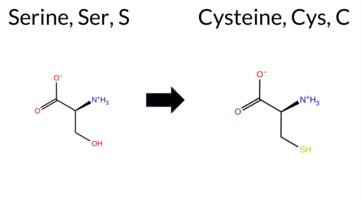 AIThe SynGAP1 missense variant S1142C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields benign, and Foldetta data are unavailable. Consequently, the variant is most likely benign based on the collective evidence, and this conclusion does not conflict with any ClinVar annotation because no ClinVar entry exists for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | -7.355 | In-Between | 0.182 | Likely Benign | Likely Benign | 0.146 | Likely Benign | -2.89 | Deleterious | 0.992 | Probably Damaging | 0.866 | Possibly Damaging | 2.70 | Benign | 0.00 | Affected | 0.1170 | 0.6016 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.3425C>T | S1142F 2D 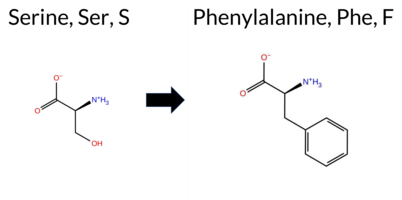 AIThe SynGAP1 missense variant S1142F has no ClinVar record and is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign majority, and Foldetta data are unavailable. Overall, the balance of evidence—especially from the high‑accuracy tools—suggests that the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | -5.074 | Likely Benign | 0.508 | Ambiguous | Likely Benign | 0.206 | Likely Benign | -3.70 | Deleterious | 0.918 | Possibly Damaging | 0.827 | Possibly Damaging | 2.65 | Benign | 0.00 | Affected | 0.0861 | 0.5603 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.342G>C | K114N 2D 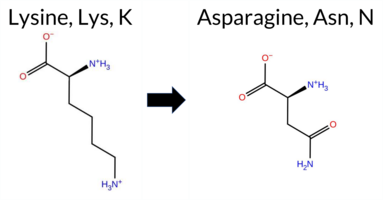 AIThe SynGAP1 missense variant K114N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus remains likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -3.851 | Likely Benign | 0.937 | Likely Pathogenic | Ambiguous | 0.045 | Likely Benign | -1.41 | Neutral | 0.608 | Possibly Damaging | 0.190 | Benign | 3.95 | Benign | 0.00 | Affected | 0.4590 | 0.1877 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.342G>T | K114N 2D AIThe SynGAP1 missense variant K114N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is not contradicted by ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.649749 | Binding | 0.381 | 0.879 | 0.750 | -3.851 | Likely Benign | 0.937 | Likely Pathogenic | Ambiguous | 0.045 | Likely Benign | -1.41 | Neutral | 0.608 | Possibly Damaging | 0.190 | Benign | 3.95 | Benign | 0.00 | Affected | 0.4590 | 0.1877 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3434A>T | N1145I 2D  AIThe SynGAP1 missense variant N1145I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools cluster into two groups: pathogenic predictions come from REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; benign predictions come from ESM1b, FATHMM, and AlphaMissense‑Optimized. AlphaMissense‑Default is uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, while Foldetta results are unavailable. Overall, the majority of conventional predictors indicate pathogenicity, whereas the high‑accuracy subset leans benign. Based on the aggregate evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.922952 | Disordered | 0.722723 | Binding | 0.284 | 0.850 | 1.000 | -3.172 | Likely Benign | 0.378 | Ambiguous | Likely Benign | 0.504 | Likely Pathogenic | -4.19 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 5.41 | Benign | 0.00 | Affected | 0.0720 | 0.6145 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.3436C>A | P1146T 2D AIThe SynGAP1 missense variant P1146T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, as well as the SGM‑Consensus call of “Likely Benign.” In contrast, PROVEAN, polyPhen‑2 HumDiv, and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. Foldetta results are not available. Overall, the majority of evidence points to a benign effect for P1146T, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | -3.494 | Likely Benign | 0.272 | Likely Benign | Likely Benign | 0.454 | Likely Benign | -4.11 | Deleterious | 0.573 | Possibly Damaging | 0.334 | Benign | 5.51 | Benign | 0.00 | Affected | 0.1478 | 0.5789 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3436C>G | P1146A 2D AIThe SynGAP1 missense variant P1146A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, tools that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P1146A, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | -3.896 | Likely Benign | 0.203 | Likely Benign | Likely Benign | 0.300 | Likely Benign | -3.56 | Deleterious | 0.972 | Probably Damaging | 0.760 | Possibly Damaging | 5.53 | Benign | 0.00 | Affected | 0.3516 | 0.4791 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3436C>T | P1146S 2D AIThe SynGAP1 missense variant P1146S is reported in ClinVar as “Not submitted” and is present in gnomAD (variant ID 6-33444471-C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of a ClinVar pathogenic classification. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | 6-33444471-C-T | 2 | 1.24e-6 | -3.045 | Likely Benign | 0.318 | Likely Benign | Likely Benign | 0.470 | Likely Benign | -3.66 | Deleterious | 0.945 | Possibly Damaging | 0.760 | Possibly Damaging | 5.52 | Benign | 0.00 | Affected | 4.32 | 4 | 0.3408 | 0.5186 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3437C>A | P1146H 2D AIThe SynGAP1 missense variant P1146H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT) predict a pathogenic impact; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign outcome (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | -4.428 | Likely Benign | 0.518 | Ambiguous | Likely Benign | 0.529 | Likely Pathogenic | -4.93 | Deleterious | 1.000 | Probably Damaging | 0.983 | Probably Damaging | 5.46 | Benign | 0.00 | Affected | 0.1625 | 0.4512 | 0 | -2 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3437C>G | P1146R 2D AIThe SynGAP1 missense variant P1146R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a pathogenic effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. Tools that agree on a benign effect are ESM1b, FATHMM, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of available predictions (seven pathogenic versus three benign) indicate that the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | -3.826 | Likely Benign | 0.729 | Likely Pathogenic | Likely Benign | 0.603 | Likely Pathogenic | -4.81 | Deleterious | 0.996 | Probably Damaging | 0.967 | Probably Damaging | 5.50 | Benign | 0.00 | Affected | 0.1315 | 0.3193 | 0 | -2 | -2.9 | 59.07 | ||||||||||||||||||||||||||||||||||||||||
| c.3437C>T | P1146L 2D AISynGAP1 missense variant P1146L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas a separate group predicts pathogenicity: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also favors a benign outcome, while Foldetta results are unavailable. Overall, the majority of conventional predictors indicate pathogenicity, but the most accurate tools lean benign. Thus, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | -2.182 | Likely Benign | 0.483 | Ambiguous | Likely Benign | 0.564 | Likely Pathogenic | -5.25 | Deleterious | 0.992 | Probably Damaging | 0.912 | Probably Damaging | 5.51 | Benign | 0.00 | Affected | 0.2151 | 0.6446 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3442A>C | M1148L 2D  AIThe SynGAP1 missense variant M1148L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and there is no conflict with ClinVar status because no ClinVar classification exists. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -1.777 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -1.13 | Neutral | 0.016 | Benign | 0.016 | Benign | 2.62 | Benign | 0.00 | Affected | 4.32 | 2 | 0.1641 | 0.4135 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||||
| c.3442A>G | M1148V 2D  AIThe SynGAP1 missense variant M1148V is catalogued in ClinVar with no submitted interpretation and is present in gnomAD (ID 6‑33444477‑A‑G). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of available predictions indicates that M1148V is most likely benign, and this conclusion does not contradict any ClinVar status, as no pathogenic claim has been reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | 6-33444477-A-G | 3 | 1.86e-6 | -2.358 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.057 | Likely Benign | -1.47 | Neutral | 0.016 | Benign | 0.026 | Benign | 2.59 | Benign | 0.00 | Affected | 4.32 | 2 | 0.3154 | 0.3371 | 1 | 2 | 2.3 | -32.06 | ||||||||||||||||||||||||||||||||||
| c.3442A>T | M1148L 2D AIThe SynGAP1 missense variant M1148L is listed in ClinVar (ID 1010061.0) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this is consistent with the ClinVar “Uncertain” designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | Uncertain | 1 | -1.777 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -1.13 | Neutral | 0.016 | Benign | 0.016 | Benign | 2.62 | Benign | 0.00 | Affected | 4.32 | 2 | 0.1641 | 0.4135 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||
| c.3443T>A | M1148K 2D  AIThe SynGAP1 missense variant M1148K is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -2.527 | Likely Benign | 0.624 | Likely Pathogenic | Likely Benign | 0.105 | Likely Benign | -1.58 | Neutral | 0.144 | Benign | 0.085 | Benign | 2.55 | Benign | 0.00 | Affected | 0.1539 | 0.1056 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||||||||||||
| c.3443T>C | M1148T 2D  AIThe SynGAP1 missense variant M1148T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -0.972 | Likely Benign | 0.159 | Likely Benign | Likely Benign | 0.151 | Likely Benign | -1.50 | Neutral | 0.001 | Benign | 0.002 | Benign | 2.56 | Benign | 0.00 | Affected | 0.2063 | 0.2214 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.3443T>G | M1148R 2D  AIThe SynGAP1 missense variant M1148R is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. High‑accuracy assessments reinforce the benign prediction: AlphaMissense‑Optimized scores the variant as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. No Foldetta stability analysis is available for this variant. Overall, the majority of computational evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -2.303 | Likely Benign | 0.669 | Likely Pathogenic | Likely Benign | 0.129 | Likely Benign | -1.77 | Neutral | 0.255 | Benign | 0.113 | Benign | 2.54 | Benign | 0.00 | Affected | 0.1624 | 0.0888 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.3444G>A | M1148I 2D  AIThe SynGAP1 missense variant M1148I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. There is no ClinVar annotation to contradict this conclusion, so the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -3.017 | Likely Benign | 0.348 | Ambiguous | Likely Benign | 0.021 | Likely Benign | -1.55 | Neutral | 0.144 | Benign | 0.062 | Benign | 2.57 | Benign | 0.00 | Affected | 0.1453 | 0.3041 | 2 | 1 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3444G>C | M1148I 2D AIThe SynGAP1 missense variant M1148I has no ClinVar entry and is not reported in gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and AlphaMissense‑Optimized all indicate a benign or likely benign outcome. Only SIFT predicts a pathogenic effect, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, the SGM‑Consensus also reports likely benign, and Foldetta data are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -3.017 | Likely Benign | 0.348 | Ambiguous | Likely Benign | 0.021 | Likely Benign | -1.55 | Neutral | 0.144 | Benign | 0.062 | Benign | 2.57 | Benign | 0.00 | Affected | 0.1453 | 0.3041 | 2 | 1 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3444G>T | M1148I 2D AIThe SynGAP1 missense variant M1148I has no ClinVar entry and is not reported in gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and AlphaMissense‑Optimized all indicate a benign or likely benign outcome. Only SIFT predicts a pathogenic effect, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, the SGM‑Consensus also reports likely benign, and Foldetta data are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -3.017 | Likely Benign | 0.348 | Ambiguous | Likely Benign | 0.021 | Likely Benign | -1.55 | Neutral | 0.144 | Benign | 0.062 | Benign | 2.57 | Benign | 0.00 | Affected | 0.1453 | 0.3041 | 2 | 1 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3455A>T | E1152V 2D  AIThe SynGAP1 missense variant E1152V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.741537 | Disordered | 0.811118 | Binding | 0.395 | 0.846 | 0.500 | -3.304 | Likely Benign | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.408 | Likely Benign | -4.65 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.33 | Pathogenic | 0.00 | Affected | 0.1247 | 0.6384 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.3457C>G | R1153G 2D  AIThe SynGAP1 missense variant R1153G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that R1153G is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.762850 | Disordered | 0.820118 | Binding | 0.361 | 0.848 | 0.625 | -3.010 | Likely Benign | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.309 | Likely Benign | -5.05 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 1.48 | Pathogenic | 0.00 | Affected | 0.3408 | 0.3475 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.3457C>T | R1153W 2D  AIThe SynGAP1 missense variant R1153W is listed in ClinVar (ID 521099.0) with an uncertain significance designation and is present in the gnomAD database (variant ID 6‑33444492‑C‑T). Functional prediction tools cluster into two groups: benign predictions from REVEL and ESM1b, and pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus method SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as likely pathogenic. AlphaMissense‑Optimized independently predicts pathogenicity. No Foldetta stability assessment is available for this residue. Taken together, the majority of evidence points to a pathogenic effect, which is in contrast to the ClinVar uncertain classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.762850 | Disordered | 0.820118 | Binding | 0.361 | 0.848 | 0.625 | Uncertain | 2 | 6-33444492-C-T | 2 | 1.24e-6 | -5.812 | Likely Benign | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.317 | Likely Benign | -5.88 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.46 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1205 | 0.3340 | 2 | -3 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||
| c.3458G>A | R1153Q 2D  AIThe SynGAP1 missense variant R1153Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized model rates the variant as uncertain, and the SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies it as Likely Pathogenic. No Foldetta stability assessment is available for this residue. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status. Thus, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.762850 | Disordered | 0.820118 | Binding | 0.361 | 0.848 | 0.625 | -3.349 | Likely Benign | 0.938 | Likely Pathogenic | Ambiguous | 0.285 | Likely Benign | -2.86 | Deleterious | 0.998 | Probably Damaging | 0.992 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.3115 | 0.2175 | 1 | 1 | 1.0 | -28.06 | |||||||||||||||||||||||||||||||||||||||
| c.3458G>C | R1153P 2D  AIThe SynGAP1 missense variant R1153P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that R1153P is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar assertion exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.762850 | Disordered | 0.820118 | Binding | 0.361 | 0.848 | 0.625 | -2.431 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.384 | Likely Benign | -5.01 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.47 | Pathogenic | 0.00 | Affected | 0.2043 | 0.4315 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3458G>T | R1153L 2D  AIThe SynGAP1 missense variant R1153L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. When predictions are grouped by consensus, the majority of algorithms (seven of nine) favor a deleterious effect, while only two suggest a benign outcome. High‑accuracy assessments further support a damaging interpretation: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Pathogenic. Foldetta results are not available. Overall, the evidence points to a pathogenic impact for R1153L, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.762850 | Disordered | 0.820118 | Binding | 0.361 | 0.848 | 0.625 | -3.595 | Likely Benign | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.470 | Likely Benign | -5.05 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 1.48 | Pathogenic | 0.00 | Affected | 0.1913 | 0.4400 | -3 | -2 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||||
| c.3460A>C | T1154P 2D  AIThe SynGAP1 missense variant T1154P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that T1154P is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -2.513 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.368 | Likely Benign | -4.42 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.1667 | 0.3732 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.3460A>G | T1154A 2D  AIThe SynGAP1 missense variant T1154A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that T1154A is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar status is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -3.312 | Likely Benign | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.290 | Likely Benign | -3.55 | Deleterious | 0.992 | Probably Damaging | 0.989 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.3509 | 0.2965 | 1 | 0 | 2.5 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3460A>T | T1154S 2D  AIThe SynGAP1 missense variant T1154S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also reports it as likely pathogenic. Foldetta results are unavailable. Overall, the consensus of the available predictions indicates that T1154S is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -3.253 | Likely Benign | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.192 | Likely Benign | -2.92 | Deleterious | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 1.78 | Pathogenic | 0.00 | Affected | 0.2817 | 0.3206 | 1 | 1 | -0.1 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3461C>A | T1154K 2D  AIThe SynGAP1 missense variant T1154K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Benign predictions are limited to REVEL and ESM1b. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus classifies the variant as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that T1154K is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -3.641 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.364 | Likely Benign | -4.25 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.74 | Pathogenic | 0.00 | Affected | 0.1053 | 0.2378 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||||||||||||
| c.3461C>G | T1154R 2D  AIThe SynGAP1 missense variant T1154R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that T1154R is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -3.464 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.300 | Likely Benign | -4.05 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.73 | Pathogenic | 0.00 | Affected | 0.0903 | 0.2101 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3461C>T | T1154I 2D  AIThe SynGAP1 missense variant T1154I is not reported in ClinVar and has no gnomAD entry. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -4.489 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.351 | Likely Benign | -4.38 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.74 | Pathogenic | 0.00 | Affected | 0.0927 | 0.4956 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||||||||||||
| c.3466G>A | A1156T 2D  AIThe SynGAP1 missense variant A1156T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence from multiple in silico tools points to a pathogenic effect for A1156T, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.720929 | Disordered | 0.871395 | Binding | 0.294 | 0.861 | 0.500 | -3.732 | Likely Benign | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.285 | Likely Benign | -2.99 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 1.63 | Pathogenic | 0.00 | Affected | 0.1366 | 0.7442 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3466G>C | A1156P 2D  AIThe SynGAP1 missense variant A1156P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the remaining tools—SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic.” No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the majority of evidence indicates that A1156P is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.720929 | Disordered | 0.871395 | Binding | 0.294 | 0.861 | 0.500 | -2.847 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.405 | Likely Benign | -3.49 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.59 | Pathogenic | 0.00 | Affected | 0.1718 | 0.5104 | 1 | -1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3466G>T | A1156S 2D  AIThe SynGAP1 missense variant A1156S is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33444501‑G‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.720929 | Disordered | 0.871395 | Binding | 0.294 | 0.861 | 0.500 | 6-33444501-G-T | 1 | 6.20e-7 | -2.052 | Likely Benign | 0.798 | Likely Pathogenic | Ambiguous | 0.197 | Likely Benign | -2.23 | Neutral | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.65 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2594 | 0.6154 | 1 | 1 | -2.6 | 16.00 | |||||||||||||||||||||||||||||||||||
| c.3467C>A | A1156D 2D  AIThe SynGAP1 missense variant A1156D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that A1156D is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.720929 | Disordered | 0.871395 | Binding | 0.294 | 0.861 | 0.500 | -3.497 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.350 | Likely Benign | -4.45 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.59 | Pathogenic | 0.00 | Affected | 0.1662 | 0.2058 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.3467C>G | A1156G 2D  AIThe SynGAP1 missense variant A1156G is not reported in ClinVar and has no allele in gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the change as pathogenic, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also reports a likely pathogenic outcome. Foldetta results are unavailable for this variant. Overall, the consensus of the available predictions indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.720929 | Disordered | 0.871395 | Binding | 0.294 | 0.861 | 0.500 | -3.728 | Likely Benign | 0.964 | Likely Pathogenic | Likely Pathogenic | 0.230 | Likely Benign | -3.02 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.62 | Pathogenic | 0.00 | Affected | 0.2214 | 0.4461 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3467C>T | A1156V 2D  AIThe SynGAP1 missense variant A1156V has no ClinVar entry and is not reported in gnomAD. Prediction tools that indicate a benign effect are REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labeling it “Likely Pathogenic”; Foldetta results are unavailable. Based on the aggregate predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.720929 | Disordered | 0.871395 | Binding | 0.294 | 0.861 | 0.500 | -5.087 | Likely Benign | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.270 | Likely Benign | -2.99 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.62 | Pathogenic | 0.00 | Affected | 0.1062 | 0.6202 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||||||||||||
| c.3469T>A | W1157R 2D  AIThe SynGAP1 missense variant W1157R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only ESM1b, whereas all other evaluated algorithms—REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. No Foldetta stability analysis is available, so it does not influence the conclusion. Overall, the preponderance of evidence indicates that W1157R is most likely pathogenic, and this assessment is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.694846 | Disordered | 0.877471 | Binding | 0.364 | 0.861 | 0.375 | -2.440 | Likely Benign | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.549 | Likely Pathogenic | -10.19 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.06 | Pathogenic | 0.00 | Affected | 0.4129 | 0.0612 | 2 | -3 | -3.6 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3469T>C | W1157R 2D AIThe SynGAP1 missense variant W1157R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only ESM1b, whereas all other evaluated algorithms—REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. No Foldetta stability analysis is available, so it does not influence the conclusion. Overall, the preponderance of evidence indicates that W1157R is most likely pathogenic, and this assessment is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.694846 | Disordered | 0.877471 | Binding | 0.364 | 0.861 | 0.375 | -2.440 | Likely Benign | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.549 | Likely Pathogenic | -10.19 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.06 | Pathogenic | 0.00 | Affected | 0.4129 | 0.0612 | 2 | -3 | -3.6 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3469T>G | W1157G 2D  AIThe SynGAP1 missense variant W1157G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely converge on a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate pathogenicity. Only ESM1b predicts a benign outcome, representing the sole disagreement. High‑accuracy assessments further support a harmful impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus labels it as Likely Pathogenic. No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the consensus of the majority of tools and the high‑accuracy predictions point to a pathogenic effect, and this conclusion is consistent with the absence of ClinVar annotation, so there is no contradiction with ClinVar status. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.694846 | Disordered | 0.877471 | Binding | 0.364 | 0.861 | 0.375 | -2.328 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.558 | Likely Pathogenic | -9.50 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 1.06 | Pathogenic | 0.00 | Affected | 0.4631 | 0.2065 | -7 | -2 | 0.5 | -129.16 | |||||||||||||||||||||||||||||||||||||||
| c.3470G>C | W1157S 2D  AIThe SynGAP1 missense variant W1157S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that W1157S is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.694846 | Disordered | 0.877471 | Binding | 0.364 | 0.861 | 0.375 | -1.158 | Likely Benign | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.394 | Likely Benign | -9.96 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.06 | Pathogenic | 0.00 | Affected | 0.4772 | 0.1227 | -2 | -3 | 0.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.3470G>T | W1157L 2D  AIThe SynGAP1 missense variant W1157L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—classify the change as pathogenic or likely pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic. Foldetta results are not available. Overall, the majority of evidence points to a pathogenic impact for W1157L, and this conclusion is consistent with the absence of a ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.694846 | Disordered | 0.877471 | Binding | 0.364 | 0.861 | 0.375 | -1.336 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.306 | Likely Benign | -9.46 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 1.08 | Pathogenic | 0.00 | Affected | 0.2400 | 0.2719 | -2 | -2 | 4.7 | -73.05 | |||||||||||||||||||||||||||||||||||||||
| c.3471G>C | W1157C 2D  AIThe SynGAP1 missense variant W1157C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that W1157C is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.694846 | Disordered | 0.877471 | Binding | 0.364 | 0.861 | 0.375 | -4.730 | Likely Benign | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.478 | Likely Benign | -9.46 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.04 | Pathogenic | 0.00 | Affected | 0.3927 | 0.1406 | -8 | -2 | 3.4 | -83.07 | |||||||||||||||||||||||||||||||||||||||
| c.3471G>T | W1157C 2D AIThe SynGAP1 missense variant W1157C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that W1157C is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.694846 | Disordered | 0.877471 | Binding | 0.364 | 0.861 | 0.375 | -4.730 | Likely Benign | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.478 | Likely Benign | -9.46 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.04 | Pathogenic | 0.00 | Affected | 0.3927 | 0.1406 | -8 | -2 | 3.4 | -83.07 | |||||||||||||||||||||||||||||||||||||||
| c.3473T>A | V1158D 2D  AIThe SynGAP1 missense variant V1158D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that V1158D is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.599170 | Disordered | 0.877504 | Binding | 0.369 | 0.847 | 0.250 | -4.076 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.486 | Likely Benign | -4.56 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.29 | Pathogenic | 0.00 | Affected | 0.1502 | 0.1820 | -2 | -3 | -7.7 | 15.96 | |||||||||||||||||||||||||||||||||||||||
| c.3473T>G | V1158G 2D  AIThe SynGAP1 missense variant V1158G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic outcome. High‑accuracy assessments reinforce this pattern: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a pathogenic verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that V1158G is most likely pathogenic, and this conclusion does not contradict the current ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.877504 | Binding | 0.369 | 0.847 | 0.250 | -3.058 | Likely Benign | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.433 | Likely Benign | -4.62 | Deleterious | 0.997 | Probably Damaging | 0.999 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1870 | 0.3289 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3499G>C | D1167H 2D  AIThe SynGAP1 missense variant D1167H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b. Tools that agree on a pathogenic effect include polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic, two benign). Foldetta predictions are unavailable. Overall, the majority of evaluated tools (six pathogenic vs. three benign) indicate a pathogenic effect. This conclusion is consistent with the lack of ClinVar reporting; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.783999 | Binding | 0.336 | 0.798 | 0.500 | -3.774 | Likely Benign | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.257 | Likely Benign | -2.36 | Neutral | 1.000 | Probably Damaging | 0.995 | Probably Damaging | 2.27 | Pathogenic | 0.00 | Affected | 0.1514 | 0.8433 | 1 | -1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||||||||||
| c.3499G>T | D1167Y 2D  AIThe SynGAP1 missense variant D1167Y is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a 2‑to‑2 split and is therefore inconclusive. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictors (six pathogenic vs. three benign) indicate that D1167Y is most likely pathogenic. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.783999 | Binding | 0.336 | 0.798 | 0.500 | -4.050 | Likely Benign | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.281 | Likely Benign | -2.46 | Neutral | 1.000 | Probably Damaging | 0.995 | Probably Damaging | 2.25 | Pathogenic | 0.00 | Affected | 0.0640 | 0.7307 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||||
| c.349A>T | S117C 2D AIThe SynGAP1 missense variant S117C is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.672422 | Binding | 0.357 | 0.877 | 0.625 | -5.980 | Likely Benign | 0.201 | Likely Benign | Likely Benign | 0.246 | Likely Benign | -1.81 | Neutral | 0.992 | Probably Damaging | 0.667 | Possibly Damaging | 3.68 | Benign | 0.00 | Affected | 0.1222 | 0.5057 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.34A>C | S12R 2D  AIThe SynGAP1 missense variant S12R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority of the four high‑accuracy tools) also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | -4.033 | Likely Benign | 0.500 | Ambiguous | Likely Benign | 0.116 | Likely Benign | -0.30 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.09 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0944 | 0.3678 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.34A>G | S12G 2D  AIThe SynGAP1 missense variant S12G is reported in gnomAD (ID 6‑33420298‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign effect for S12G, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | 6-33420298-A-G | -4.229 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.079 | Likely Benign | 0.22 | Neutral | 0.103 | Benign | 0.015 | Benign | 4.11 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2876 | 0.5080 | 0 | 1 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||||||||
| c.34A>T | S12C 2D AIThe SynGAP1 missense variant S12C is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools largely support a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, two tools—polyPhen‑2 HumDiv and SIFT—predict a pathogenic impact. High‑accuracy methods give a consistent benign signal: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this is not in conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | -5.413 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.101 | Likely Benign | 0.00 | Neutral | 0.872 | Possibly Damaging | 0.206 | Benign | 4.05 | Benign | 0.00 | Affected | 0.1025 | 0.6092 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.3500A>T | D1167V 2D  AIThe SynGAP1 missense variant D1167V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Based on the preponderance of pathogenic predictions and the high‑accuracy tool results, the variant is most likely pathogenic; this conclusion is not contradicted by any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.599170 | Disordered | 0.783999 | Binding | 0.336 | 0.798 | 0.500 | -2.750 | Likely Benign | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.288 | Likely Benign | -3.34 | Deleterious | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.26 | Pathogenic | 0.00 | Affected | 0.0946 | 0.7515 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.3505G>A | E1169K 2D  AISynGAP1 missense variant E1169K is listed in gnomAD (ID 6‑33444540‑G‑A) but has no ClinVar record. Functional prediction tools fall into two groups: benign predictions come from SGM‑Consensus, REVEL, PROVEAN, ESM1b, and FATHMM; pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely benign. Foldetta stability analysis is unavailable. Overall, the evidence is split evenly, with one high‑accuracy tool supporting pathogenicity and the consensus tool supporting benignity. Therefore, the variant’s impact remains uncertain; it is not contradicted by ClinVar status, which has no entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.699094 | Disordered | 0.732455 | Binding | 0.400 | 0.781 | 0.625 | 6-33444540-G-A | 1 | 6.20e-7 | -3.335 | Likely Benign | 0.973 | Likely Pathogenic | Likely Pathogenic | 0.185 | Likely Benign | -1.81 | Neutral | 0.997 | Probably Damaging | 0.898 | Possibly Damaging | 2.51 | Benign | 0.00 | Affected | 3.88 | 3 | 0.1969 | 0.6422 | 1 | 0 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||
| c.3505G>C | E1169Q 2D  AIThe SynGAP1 missense variant E1169Q is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33444540‑G‑C). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and ESM1b, while pathogenic predictions are made by polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, more tools (five) predict pathogenicity than benign (three), and no evidence from ClinVar contradicts this assessment. Thus, the variant is most likely pathogenic based on the current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.699094 | Disordered | 0.732455 | Binding | 0.400 | 0.781 | 0.625 | 6-33444540-G-C | 1 | 6.20e-7 | -3.096 | Likely Benign | 0.852 | Likely Pathogenic | Ambiguous | 0.119 | Likely Benign | -1.27 | Neutral | 0.872 | Possibly Damaging | 0.701 | Possibly Damaging | 2.48 | Pathogenic | 0.00 | Affected | 3.88 | 3 | 0.0940 | 0.6392 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||
| c.3506A>C | E1169A 2D  AIThe SynGAP1 missense variant E1169A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on benign impact include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, a majority‑vote method from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and Foldetta results are unavailable. Considering the consensus of benign‑predicting tools and the SGM‑Consensus outcome, the variant is most likely benign. This assessment does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.699094 | Disordered | 0.732455 | Binding | 0.400 | 0.781 | 0.625 | -2.132 | Likely Benign | 0.901 | Likely Pathogenic | Ambiguous | 0.217 | Likely Benign | -2.46 | Neutral | 0.995 | Probably Damaging | 0.949 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.3714 | 0.6293 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.3506A>G | E1169G 2D 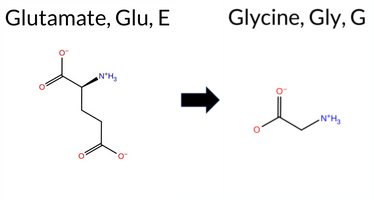 AIThe SynGAP1 missense variant E1169G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated predictors (five pathogenic vs. three benign) lean toward a pathogenic classification. This prediction is not contradicted by ClinVar status, as the variant is not yet catalogued there. Thus, based on current computational evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.699094 | Disordered | 0.732455 | Binding | 0.400 | 0.781 | 0.625 | -3.172 | Likely Benign | 0.940 | Likely Pathogenic | Ambiguous | 0.203 | Likely Benign | -2.33 | Neutral | 0.995 | Probably Damaging | 0.963 | Probably Damaging | 2.45 | Pathogenic | 0.00 | Affected | 0.3005 | 0.6019 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.3506A>T | E1169V 2D AIThe SynGAP1 E1169V missense variant is not reported in ClinVar and is absent from gnomAD. Consensus from standard prediction algorithms shows a split: benign predictions come from REVEL, ESM1b, and FATHMM, whereas pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a pathogenic signal: AlphaMissense‑Optimized is pathogenic, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—remains inconclusive (2 pathogenic vs 2 benign), and Foldetta stability analysis is unavailable. Overall, the preponderance of evidence (seven pathogenic vs three benign predictions) indicates that E1169V is most likely pathogenic. This conclusion is consistent with the absence of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.699094 | Disordered | 0.732455 | Binding | 0.400 | 0.781 | 0.625 | -2.482 | Likely Benign | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.227 | Likely Benign | -2.94 | Deleterious | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.51 | Benign | 0.00 | Affected | 0.0581 | 0.7028 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3507G>C | E1169D 2D 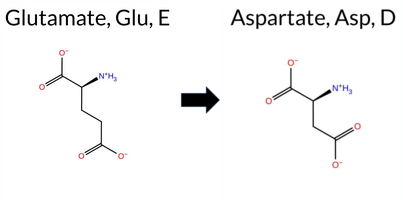 AIThe SynGAP1 missense variant E1169D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of consensus tools predict a pathogenic impact, and no ClinVar entry contradicts this assessment. Thus, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.699094 | Disordered | 0.732455 | Binding | 0.400 | 0.781 | 0.625 | -3.457 | Likely Benign | 0.913 | Likely Pathogenic | Ambiguous | 0.104 | Likely Benign | -1.12 | Neutral | 0.989 | Probably Damaging | 0.924 | Probably Damaging | 2.49 | Pathogenic | 0.00 | Affected | 0.1597 | 0.3968 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3507G>T | E1169D 2D AIThe SynGAP1 missense variant E1169D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of consensus tools predict a pathogenic impact, and no ClinVar entry contradicts this assessment. Thus, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.699094 | Disordered | 0.732455 | Binding | 0.400 | 0.781 | 0.625 | -3.457 | Likely Benign | 0.913 | Likely Pathogenic | Ambiguous | 0.103 | Likely Benign | -1.12 | Neutral | 0.989 | Probably Damaging | 0.924 | Probably Damaging | 2.49 | Pathogenic | 0.00 | Affected | 0.1597 | 0.3968 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.350G>T | S117I 2D 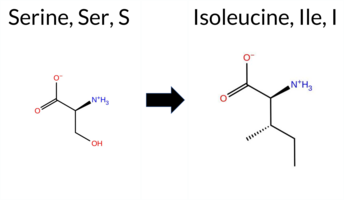 AIThe SynGAP1 missense variant S117I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus (SGM‑Consensus) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.672422 | Binding | 0.357 | 0.877 | 0.625 | -5.483 | Likely Benign | 0.559 | Ambiguous | Likely Benign | 0.137 | Likely Benign | -2.33 | Neutral | 0.971 | Probably Damaging | 0.598 | Possibly Damaging | 3.70 | Benign | 0.00 | Affected | 0.1060 | 0.5048 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3523C>G | R1175G 2D AIThe SynGAP1 missense variant R1175G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.538167 | Disordered | 0.589347 | Binding | 0.545 | 0.732 | 0.375 | -4.888 | Likely Benign | 0.865 | Likely Pathogenic | Ambiguous | 0.477 | Likely Benign | -1.64 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 5.35 | Benign | 0.00 | Affected | 0.3006 | 0.2350 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||
| c.3523C>T | R1175W 2D AIThe SynGAP1 missense variant R1175W is listed in gnomAD (ID 6‑33444558‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from ESM1b and FATHMM, while pathogenic predictions are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta data are unavailable. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no classification for this variant. Thus, based on current predictions, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.538167 | Disordered | 0.589347 | Binding | 0.545 | 0.732 | 0.375 | 6-33444558-C-T | 3 | 1.86e-6 | -5.807 | Likely Benign | 0.907 | Likely Pathogenic | Ambiguous | 0.514 | Likely Pathogenic | -2.83 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 5.32 | Benign | 0.00 | Affected | 4.32 | 2 | 0.1065 | 0.2148 | -3 | 2 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||||
| c.3524G>A | R1175Q 2D AIThe SynGAP1 missense variant R1175Q is reported in gnomAD (ID 6‑33444559‑G‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status because none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.538167 | Disordered | 0.589347 | Binding | 0.545 | 0.732 | 0.375 | 6-33444559-G-A | 1 | 6.20e-7 | -3.968 | Likely Benign | 0.529 | Ambiguous | Likely Benign | 0.328 | Likely Benign | -0.76 | Neutral | 0.998 | Probably Damaging | 0.992 | Probably Damaging | 5.39 | Benign | 0.00 | Affected | 4.32 | 2 | 0.1938 | 0.1835 | 1 | 1 | 1.0 | -28.06 | |||||||||||||||||||||||||||||||||
| c.3524G>C | R1175P 2D AIThe SynGAP1 missense variant R1175P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign predictions come from PROVEAN, ESM1b, and FATHMM, while pathogenic predictions are returned by REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign outcome. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenic, whereas the SGM‑Consensus (majority vote) predicts benign. No Foldetta stability analysis is available. Overall, the majority of high‑confidence tools lean toward a benign interpretation, and this is consistent with the absence of ClinVar evidence. Therefore, the variant is most likely benign, and there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.538167 | Disordered | 0.589347 | Binding | 0.545 | 0.732 | 0.375 | -4.746 | Likely Benign | 0.970 | Likely Pathogenic | Likely Pathogenic | 0.518 | Likely Pathogenic | -0.80 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 5.37 | Benign | 0.00 | Affected | 0.1715 | 0.3223 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||||||||
| c.3524G>T | R1175L 2D AISynGAP1 missense variant R1175L is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show a split: benign predictions come from PROVEAN, ESM1b, and FATHMM, while pathogenic predictions arise from REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized is Uncertain, SGM‑Consensus remains Likely Benign, and Foldetta data are unavailable. Overall, the majority of individual predictors lean toward pathogenicity, yet the consensus of the most reliable tools suggests a benign outcome, leaving the variant’s clinical significance ambiguous. Consequently, the variant is most likely pathogenic based on the bulk of predictions, and this assessment does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.538167 | Disordered | 0.589347 | Binding | 0.545 | 0.732 | 0.375 | -2.560 | Likely Benign | 0.876 | Likely Pathogenic | Ambiguous | 0.535 | Likely Pathogenic | -2.37 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 5.38 | Benign | 0.00 | Affected | 0.1240 | 0.2849 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3535A>C | K1179Q 2D AIThe SynGAP1 missense variant K1179Q is reported in gnomAD (variant ID 6‑33444570‑A‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions, including the high‑accuracy tools, point to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | 6-33444570-A-C | 1 | 6.20e-7 | -4.237 | Likely Benign | 0.679 | Likely Pathogenic | Likely Benign | 0.078 | Likely Benign | -1.20 | Neutral | 0.430 | Benign | 0.211 | Benign | 2.67 | Benign | 0.00 | Affected | 4.32 | 2 | 0.4037 | 0.0807 | 1 | 1 | 0.4 | -0.04 | |||||||||||||||||||||||||||||||||
| c.3535A>G | K1179E 2D AIThe SynGAP1 missense variant K1179E is reported in gnomAD (ID 6‑33444570‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic impact are polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic, SGM‑Consensus as Likely Benign, and Foldetta results are unavailable. Overall, the majority of individual predictors (five versus four) lean toward pathogenicity, and the high‑accuracy AlphaMissense‑Optimized result supports this. No ClinVar status is available to contradict these findings. Thus, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | 6-33444570-A-G | 1 | 6.20e-7 | -4.040 | Likely Benign | 0.961 | Likely Pathogenic | Likely Pathogenic | 0.143 | Likely Benign | -1.02 | Neutral | 0.800 | Possibly Damaging | 0.525 | Possibly Damaging | 2.96 | Benign | 0.00 | Affected | 4.32 | 2 | 0.3476 | 0.0876 | 1 | 0 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||
| c.3536A>C | K1179T 2D AIThe SynGAP1 missense variant K1179T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and Foldetta results are not available. Taken together, the majority of evidence points toward a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | -4.447 | Likely Benign | 0.927 | Likely Pathogenic | Ambiguous | 0.156 | Likely Benign | -1.80 | Neutral | 0.975 | Probably Damaging | 0.819 | Possibly Damaging | 2.65 | Benign | 0.00 | Affected | 0.2107 | 0.2027 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||||||||
| c.3536A>G | K1179R 2D AIThe SynGAP1 missense variant K1179R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available. Overall, the majority of evidence points to a benign effect for K1179R, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | -2.677 | Likely Benign | 0.178 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -0.92 | Neutral | 0.951 | Possibly Damaging | 0.628 | Possibly Damaging | 2.66 | Benign | 0.00 | Affected | 0.4010 | 0.0782 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3536A>T | K1179M 2D AIThe SynGAP1 missense variant K1179M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and Foldetta results are unavailable. Taken together, the majority of evidence points toward a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | -4.429 | Likely Benign | 0.929 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -1.98 | Neutral | 0.998 | Probably Damaging | 0.969 | Probably Damaging | 2.61 | Benign | 0.00 | Affected | 0.1057 | 0.2715 | 0 | -1 | 5.8 | 3.02 | ||||||||||||||||||||||||||||||||||||||
| c.3537G>C | K1179N 2D AIThe SynGAP1 missense variant K1179N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Among general in‑silico predictors, benign calls come from REVEL, PROVEAN, ESM1b, and FATHMM, whereas pathogenic calls come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy tools give a mixed picture: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of high‑confidence predictions lean toward pathogenicity, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely pathogenic based on the current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | -4.764 | Likely Benign | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.113 | Likely Benign | -1.57 | Neutral | 0.975 | Probably Damaging | 0.766 | Possibly Damaging | 2.68 | Benign | 0.00 | Affected | 0.3490 | 0.0901 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3537G>T | K1179N 2D AIThe SynGAP1 K1179N missense variant is not reported in ClinVar and has no entries in gnomAD. General in silico predictors cluster into two groups: benign predictions from REVEL, PROVEAN, ESM1b, and FATHMM; pathogenic predictions from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy tools provide a mixed signal: AlphaMissense‑Optimized classifies the change as pathogenic, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect; Foldetta results are unavailable. Overall, the evidence is split, with no single consensus. Thus, the variant is currently inconclusive—neither clearly benign nor pathogenic—and does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.637480 | Disordered | 0.558455 | Binding | 0.575 | 0.678 | 0.250 | -4.764 | Likely Benign | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.113 | Likely Benign | -1.57 | Neutral | 0.975 | Probably Damaging | 0.766 | Possibly Damaging | 2.68 | Benign | 0.00 | Affected | 0.3490 | 0.0901 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3538C>A | L1180I 2D 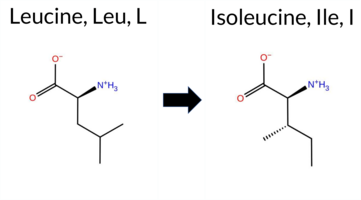 AIThe SynGAP1 missense variant L1180I is not reported in ClinVar and has no gnomAD entry. Prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for the variant, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.559845 | Binding | 0.591 | 0.672 | 0.250 | -4.553 | Likely Benign | 0.697 | Likely Pathogenic | Likely Benign | 0.086 | Likely Benign | -0.67 | Neutral | 0.856 | Possibly Damaging | 0.578 | Possibly Damaging | 2.69 | Benign | 0.00 | Affected | 0.0825 | 0.2708 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||
| c.3538C>G | L1180V 2D  AIThe SynGAP1 missense variant L1180V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls come from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. Grouping by consensus, the majority of tools (five) predict benign, while four predict pathogenic. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. AlphaMissense‑Optimized independently predicts benign. No Foldetta stability assessment is available. Overall, the preponderance of evidence points to a benign effect for the variant, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.559845 | Binding | 0.591 | 0.672 | 0.250 | -4.664 | Likely Benign | 0.741 | Likely Pathogenic | Likely Benign | 0.086 | Likely Benign | -1.00 | Neutral | 0.856 | Possibly Damaging | 0.474 | Possibly Damaging | 2.71 | Benign | 0.00 | Affected | 0.1386 | 0.1991 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3538C>T | L1180F 2D  AIThe SynGAP1 missense variant L1180F is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of high‑accuracy predictions lean toward a benign impact, and there is no conflict with ClinVar status. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.559845 | Binding | 0.591 | 0.672 | 0.250 | -5.370 | Likely Benign | 0.953 | Likely Pathogenic | Ambiguous | 0.077 | Likely Benign | -1.29 | Neutral | 0.749 | Possibly Damaging | 0.444 | Benign | 2.65 | Benign | 0.00 | Affected | 0.0516 | 0.2345 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3539T>A | L1180H 2D  AIThe SynGAP1 missense variant L1180H is not reported in ClinVar and has no gnomAD allele, so its population frequency is unknown. Functional prediction tools show a split: benign calls come from REVEL, PROVEAN, ESM1b, and FATHMM, whereas pathogenic calls come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments highlight AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus leans benign; Foldetta results are not available. Overall, five of nine individual predictors favor pathogenicity, four favor benign, and the consensus tool suggests benign. Thus, the variant is most likely pathogenic based on the preponderance of high‑confidence predictions, and this assessment is not contradicted by ClinVar, which contains no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.559845 | Binding | 0.591 | 0.672 | 0.250 | -5.621 | Likely Benign | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.213 | Likely Benign | -0.22 | Neutral | 0.987 | Probably Damaging | 0.865 | Possibly Damaging | 2.65 | Benign | 0.00 | Affected | 0.0991 | 0.0860 | -2 | -3 | -7.0 | 23.98 | ||||||||||||||||||||||||||||||||||||||
| c.3539T>C | L1180P 2D  AISynGAP1 missense variant L1180P is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, whereas the SGM‑Consensus remains likely benign; Foldetta results are unavailable. Overall, the balance of evidence, including the consensus prediction and the higher number of benign calls, suggests the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.559845 | Binding | 0.591 | 0.672 | 0.250 | -4.564 | Likely Benign | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.213 | Likely Benign | -1.37 | Neutral | 0.992 | Probably Damaging | 0.930 | Probably Damaging | 2.65 | Benign | 0.00 | Affected | 0.3556 | 0.1155 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||
| c.3539T>G | L1180R 2D 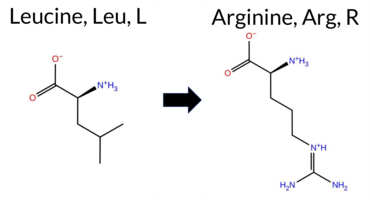 AIThe SynGAP1 missense variant L1180R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is not available. Overall, the majority of evidence—including the SGM Consensus and several benign‑predicting tools—suggests a benign impact. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.626927 | Disordered | 0.559845 | Binding | 0.591 | 0.672 | 0.250 | -4.238 | Likely Benign | 0.920 | Likely Pathogenic | Ambiguous | 0.175 | Likely Benign | -1.58 | Neutral | 0.977 | Probably Damaging | 0.900 | Possibly Damaging | 2.67 | Benign | 0.00 | Affected | 0.1199 | 0.0660 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.353T>G | M118R 2D AIThe SynGAP1 missense variant M118R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign impact for M118R. This conclusion does not contradict any ClinVar annotation, as no ClinVar status is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.676867 | Binding | 0.330 | 0.883 | 0.500 | -3.318 | Likely Benign | 0.698 | Likely Pathogenic | Likely Benign | 0.286 | Likely Benign | -3.17 | Deleterious | 0.697 | Possibly Damaging | 0.202 | Benign | 3.83 | Benign | 0.00 | Affected | 0.2027 | 0.0913 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||||||||||||
| c.3545A>T | E1182V 2D AIThe SynGAP1 missense variant E1182V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas the majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized) predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the preponderance of evidence (seven pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.566480 | Disordered | 0.530232 | Binding | 0.597 | 0.651 | 0.375 | -4.966 | Likely Benign | 0.966 | Likely Pathogenic | Likely Pathogenic | 0.124 | Likely Benign | -3.21 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.59 | Benign | 0.00 | Affected | 0.0447 | 0.6364 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.3562G>A | D1188N 2D 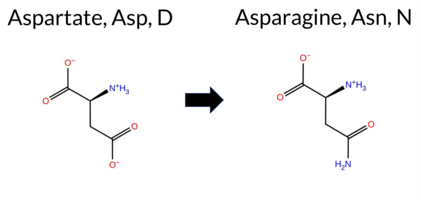 AIThe SynGAP1 D1188N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the balance of evidence favors a pathogenic classification for D1188N, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -5.621 | Likely Benign | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.340 | Likely Benign | -2.50 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 5.47 | Benign | 0.00 | Affected | 0.0767 | 0.4663 | 2 | 1 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3562G>C | D1188H 2D AIThe SynGAP1 D1188H missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, while those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of evaluated tools (seven pathogenic vs. three benign) indicate a pathogenic effect. Thus, the variant is most likely pathogenic, and this prediction does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -6.827 | Likely Benign | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.420 | Likely Benign | -3.27 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 5.41 | Benign | 0.00 | Affected | 0.0892 | 0.5242 | 1 | -1 | 0.3 | 22.05 | |||||||||||||||||||||||||||||||||||||||
| c.3562G>T | D1188Y 2D AIThe SynGAP1 D1188Y missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evaluated tools (seven pathogenic versus three benign) indicate a pathogenic impact. This prediction aligns with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -6.482 | Likely Benign | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.490 | Likely Benign | -4.49 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 5.40 | Benign | 0.00 | Affected | 0.0450 | 0.4714 | -4 | -3 | 2.2 | 48.09 | |||||||||||||||||||||||||||||||||||||||
| c.3563A>C | D1188A 2D 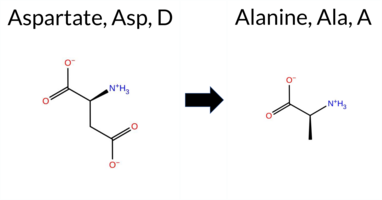 AIThe SynGAP1 D1188A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL, ESM1b, and FATHMM, while those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact. This prediction is consistent with the lack of ClinVar annotation and gnomAD presence, indicating no contradiction with existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -4.369 | Likely Benign | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.439 | Likely Benign | -3.91 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 5.45 | Benign | 0.00 | Affected | 0.2768 | 0.4495 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||||
| c.3563A>G | D1188G 2D 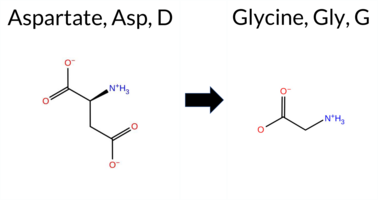 AIThe SynGAP1 D1188G missense change is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus method SGM, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a tie and is therefore inconclusive. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Overall, the majority of evidence points to a pathogenic effect, and this assessment does not conflict with any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -5.286 | Likely Benign | 0.981 | Likely Pathogenic | Likely Pathogenic | 0.440 | Likely Benign | -3.69 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 5.47 | Benign | 0.00 | Affected | 0.2771 | 0.4880 | 1 | -1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.3563A>T | D1188V 2D AIThe SynGAP1 D1188V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the balance of evidence (seven pathogenic versus three benign predictions) indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -4.482 | Likely Benign | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.479 | Likely Benign | -4.13 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 5.49 | Benign | 0.00 | Affected | 0.0592 | 0.4651 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.3564T>A | D1188E 2D 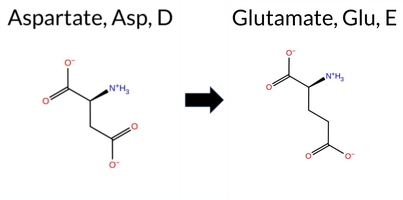 AIThe SynGAP1 missense variant D1188E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized model classifies the variant as pathogenic, whereas the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic interpretation. The variant is most likely pathogenic based on the consensus of computational tools, and this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -2.517 | Likely Benign | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.382 | Likely Benign | -1.93 | Neutral | 0.992 | Probably Damaging | 0.992 | Probably Damaging | 5.50 | Benign | 0.00 | Affected | 0.0954 | 0.4616 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3564T>G | D1188E 2D AIThe SynGAP1 missense variant D1188E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy predictors give a mixed picture: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this residue. Overall, five of the nine evaluated tools predict pathogenicity while four predict benignity, with the high‑accuracy AlphaMissense‑Optimized result supporting the pathogenic prediction. Therefore, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.476583 | Structured | 0.484322 | Uncertain | 0.687 | 0.626 | 0.625 | -2.517 | Likely Benign | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.382 | Likely Benign | -1.93 | Neutral | 0.992 | Probably Damaging | 0.992 | Probably Damaging | 5.50 | Benign | 0.00 | Affected | 0.0954 | 0.4616 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.356A>T | E119V 2D AISynGAP1 missense variant E119V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include REVEL, ESM1b, FATHMM, and polyPhen‑2 HumVar, while pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy methods are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a tie and is therefore unavailable, and Foldetta results are not provided. Consequently, the evidence does not strongly support either outcome. The variant is most likely inconclusive; it does not clearly favor benign or pathogenic status, and this lack of consensus does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.690604 | Disordered | 0.661946 | Binding | 0.346 | 0.881 | 0.750 | -5.696 | Likely Benign | 0.842 | Likely Pathogenic | Ambiguous | 0.151 | Likely Benign | -2.78 | Deleterious | 0.596 | Possibly Damaging | 0.189 | Benign | 3.79 | Benign | 0.00 | Affected | 0.1152 | 0.7753 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3577G>A | D1193N 2D AIThe SynGAP1 missense variant D1193N is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta results are unavailable. Overall, the majority of high‑confidence tools and the consensus prediction favor a benign classification. Thus, the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | -6.472 | Likely Benign | 0.618 | Likely Pathogenic | Likely Benign | 0.334 | Likely Benign | -1.63 | Neutral | 0.856 | Possibly Damaging | 0.652 | Possibly Damaging | 5.44 | Benign | 0.00 | Affected | 0.1051 | 0.4004 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3577G>C | D1193H 2D AIThe SynGAP1 missense variant D1193H is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into three groups: benign predictions from REVEL, PROVEAN, and FATHMM; pathogenic predictions from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default; and uncertain predictions from ESM1b and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized inconclusive, an SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) that is benign, and no Foldetta result available. Overall, the majority of conventional tools predict pathogenicity, while the SGM Consensus suggests benign. Based on the combined evidence, the variant is most likely pathogenic, and this assessment does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | -7.633 | In-Between | 0.900 | Likely Pathogenic | Ambiguous | 0.400 | Likely Benign | -2.31 | Neutral | 0.977 | Probably Damaging | 0.924 | Probably Damaging | 5.41 | Benign | 0.00 | Affected | 0.1239 | 0.4584 | 1 | -1 | 0.3 | 22.05 | |||||||||||||||||||||||||||||||||||||||
| c.3577G>T | D1193Y 2D AIThe SynGAP1 missense variant D1193Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Based on the overall pattern of predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | -8.233 | Likely Pathogenic | 0.883 | Likely Pathogenic | Ambiguous | 0.484 | Likely Benign | -2.94 | Deleterious | 0.992 | Probably Damaging | 0.947 | Probably Damaging | 5.50 | Benign | 0.00 | Affected | 0.0477 | 0.4111 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||
| c.3578A>C | D1193A 2D AIThe SynGAP1 missense variant D1193A is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available result for this variant. Based on the majority of predictions and the consensus from high‑accuracy tools, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | -5.747 | Likely Benign | 0.767 | Likely Pathogenic | Likely Benign | 0.486 | Likely Benign | -2.42 | Neutral | 0.856 | Possibly Damaging | 0.492 | Possibly Damaging | 5.48 | Benign | 0.00 | Affected | 0.2719 | 0.4017 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||||||||||
| c.3578A>G | D1193G 2D AIThe SynGAP1 D1193G missense variant is catalogued in gnomAD (ID 6‑33444613‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority of the four high‑accuracy tools) also favors benign. Foldetta results are unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | 6-33444613-A-G | 1 | 6.21e-7 | -6.506 | Likely Benign | 0.765 | Likely Pathogenic | Likely Benign | 0.480 | Likely Benign | -2.27 | Neutral | 0.924 | Possibly Damaging | 0.652 | Possibly Damaging | 5.42 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2695 | 0.4765 | -1 | 1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||
| c.3578A>T | D1193V 2D AIThe SynGAP1 D1193V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a pathogenic effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default, while only FATHMM predicts a benign outcome. ESM1b and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (two pathogenic vs. one benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | -7.297 | In-Between | 0.855 | Likely Pathogenic | Ambiguous | 0.526 | Likely Pathogenic | -2.92 | Deleterious | 0.977 | Probably Damaging | 0.856 | Possibly Damaging | 5.51 | Benign | 0.00 | Affected | 0.0752 | 0.4174 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.3579T>A | D1193E 2D AIThe SynGAP1 missense variant D1193E is predicted to be benign by the majority of in silico tools. Benign predictions come from REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT classifies the change as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments further support a benign effect: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. ClinVar contains no entry for this variant, and it is absent from gnomAD, so there is no external evidence to contradict the computational predictions. Therefore, the variant is most likely benign, with no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | -3.788 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.307 | Likely Benign | -1.09 | Neutral | 0.008 | Benign | 0.028 | Benign | 5.47 | Benign | 0.00 | Affected | 0.1242 | 0.3957 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3579T>G | D1193E 2D AIThe SynGAP1 missense variant D1193E is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the preponderance of evidence supports a benign classification for D1193E, and this conclusion is consistent with the lack of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | -3.788 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.307 | Likely Benign | -1.09 | Neutral | 0.008 | Benign | 0.028 | Benign | 5.47 | Benign | 0.00 | Affected | 0.1242 | 0.3957 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3580A>T | R1194W 2D AIThe SynGAP1 missense variant R1194W is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: benign predictions are limited to FATHMM, whereas the remaining methods—REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) confirms a likely pathogenic status. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the preponderance of evidence points to the variant being most likely pathogenic, with no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.685117 | Disordered | 0.425297 | Uncertain | 0.796 | 0.602 | 0.375 | -9.583 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.514 | Likely Pathogenic | -2.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 5.41 | Benign | 0.00 | Affected | 0.1252 | 0.3162 | 2 | -3 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.3595G>A | E1199K 2D AIThe SynGAP1 missense variant E1199K (ClinVar ID 1026146.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33446587‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are not available. Overall, the majority of evidence points toward a pathogenic impact, which does not contradict the ClinVar Uncertain classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.538167 | Disordered | 0.444533 | Uncertain | 0.878 | 0.598 | 0.250 | Uncertain | 1 | 6-33446587-G-A | 1 | 6.20e-7 | -10.853 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.171 | Likely Benign | -2.26 | Neutral | 1.000 | Probably Damaging | 0.995 | Probably Damaging | 2.52 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1871 | 0.4072 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||
| c.3595G>C | E1199Q 2D AIThe SynGAP1 missense variant E1199Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default all predict a pathogenic impact. ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, while Foldetta results are unavailable. Overall, the balance of evidence—particularly from the high‑accuracy tools—suggests that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.538167 | Disordered | 0.444533 | Uncertain | 0.878 | 0.598 | 0.250 | -7.428 | In-Between | 0.752 | Likely Pathogenic | Likely Benign | 0.132 | Likely Benign | -1.41 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.70 | Benign | 0.00 | Affected | 0.1000 | 0.3545 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3596A>C | E1199A 2D AIThe SynGAP1 missense variant E1199A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. The high‑accuracy consensus, SGM‑Consensus, also indicates a likely pathogenic outcome, while AlphaMissense‑Optimized is uncertain and Foldetta results are unavailable. Taken together, the majority of evidence points to a pathogenic effect for E1199A, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.538167 | Disordered | 0.444533 | Uncertain | 0.878 | 0.598 | 0.250 | -13.556 | Likely Pathogenic | 0.953 | Likely Pathogenic | Ambiguous | 0.367 | Likely Benign | -4.32 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.48 | Pathogenic | 0.00 | Affected | 0.2858 | 0.3834 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3596A>G | E1199G 2D AIThe SynGAP1 missense change E1199G is not reported in ClinVar (ClinVar ID = None) and has no entries in gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. The high‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for E1199G, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.538167 | Disordered | 0.444533 | Uncertain | 0.878 | 0.598 | 0.250 | -13.414 | Likely Pathogenic | 0.947 | Likely Pathogenic | Ambiguous | 0.360 | Likely Benign | -5.08 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.45 | Pathogenic | 0.00 | Affected | 0.2461 | 0.3960 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||
| c.3596A>T | E1199V 2D AIThe SynGAP1 missense change E1199V is not reported in ClinVar and is absent from gnomAD. Prediction tools that flag the variant as benign include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic effect. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus likewise reports a likely pathogenic outcome. Foldetta results are not available for this variant. Overall, the preponderance of computational evidence points to a pathogenic effect for E1199V, and this conclusion does not conflict with the current ClinVar status, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.538167 | Disordered | 0.444533 | Uncertain | 0.878 | 0.598 | 0.250 | -12.285 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.360 | Likely Benign | -5.14 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.43 | Pathogenic | 0.00 | Affected | 0.0715 | 0.4581 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3597G>C | E1199D 2D AIThe SynGAP1 missense variant E1199D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and FATHMM, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.538167 | Disordered | 0.444533 | Uncertain | 0.878 | 0.598 | 0.250 | -10.917 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.234 | Likely Benign | -2.08 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.52 | Benign | 0.00 | Affected | 0.1728 | 0.2146 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3597G>T | E1199D 2D AIThe SynGAP1 missense variant E1199D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and FATHMM, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.538167 | Disordered | 0.444533 | Uncertain | 0.878 | 0.598 | 0.250 | -10.917 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.236 | Likely Benign | -2.08 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.52 | Benign | 0.00 | Affected | 0.1728 | 0.2146 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3599A>T | E1200V 2D AIThe SynGAP1 E1200V missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2) and Foldetta data are unavailable. Overall, the majority of standard predictors lean toward pathogenicity, but the single high‑accuracy benign prediction and the inconclusive consensus leave the variant’s impact uncertain. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.545602 | Disordered | 0.458056 | Uncertain | 0.889 | 0.596 | 0.250 | -4.987 | Likely Benign | 0.784 | Likely Pathogenic | Likely Benign | 0.274 | Likely Benign | -3.52 | Deleterious | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.63 | Benign | 0.00 | Affected | 0.0498 | 0.4648 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.35G>A | S12N 2D 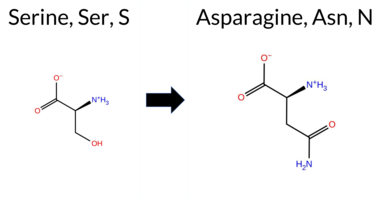 AIThe SynGAP1 missense variant S12N is listed in ClinVar with no submitted interpretation and is present in gnomAD (variant ID 6‑33420299‑G‑A). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized reports benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” No Foldetta stability result is available. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | 6-33420299-G-A | -4.946 | Likely Benign | 0.185 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -0.41 | Neutral | 0.208 | Benign | 0.018 | Benign | 4.10 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1405 | 0.5004 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||||
| c.35G>C | S12T 2D AIThe SynGAP1 missense variant S12T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a benign effect. AlphaMissense‑Optimized independently scores the variant as benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | -4.304 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -0.16 | Neutral | 0.208 | Benign | 0.024 | Benign | 4.14 | Benign | 0.00 | Affected | 0.1487 | 0.6303 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.35G>T | S12I 2D AIThe SynGAP1 missense variant S12I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | -4.838 | Likely Benign | 0.299 | Likely Benign | Likely Benign | 0.121 | Likely Benign | 0.05 | Neutral | 0.659 | Possibly Damaging | 0.072 | Benign | 4.09 | Benign | 0.00 | Affected | 0.0964 | 0.6082 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3602A>T | E1201V 2D AIThe SynGAP1 missense variant E1201V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign change, whereas all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. No Foldetta stability analysis is available for this variant. Overall, the preponderance of evidence from multiple prediction tools and consensus methods indicates that E1201V is most likely pathogenic, and this conclusion is consistent with the absence of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.525368 | Disordered | 0.481868 | Uncertain | 0.870 | 0.596 | 0.250 | -10.865 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.402 | Likely Benign | -5.43 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.59 | Pathogenic | 0.00 | Affected | 0.0455 | 0.6308 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3605T>A | I1202N 2D 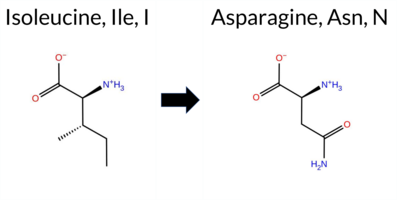 AIThe SynGAP1 missense variant I1202N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Functional prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) is likely pathogenic. Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, with no ClinVar status to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.510422 | Binding | 0.874 | 0.593 | 0.250 | -10.922 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.303 | Likely Benign | -5.65 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.79 | Pathogenic | 0.00 | Affected | 0.1014 | 0.0270 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3605T>G | I1202S 2D 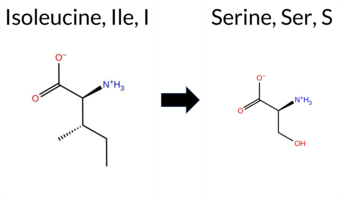 AIThe SynGAP1 missense variant I1202S is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are not available. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.510422 | Binding | 0.874 | 0.593 | 0.250 | -11.877 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.431 | Likely Benign | -4.68 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.3021 | 0.0640 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3613C>A | L1205M 2D  AIThe SynGAP1 missense variant L1205M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the variant as damaging. The high‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple in silico predictors points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.552471 | Binding | 0.880 | 0.576 | 0.375 | -9.793 | Likely Pathogenic | 0.945 | Likely Pathogenic | Ambiguous | 0.231 | Likely Benign | -1.73 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.0627 | 0.2297 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3613C>G | L1205V 2D AIThe SynGAP1 missense variant L1205V is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.552471 | Binding | 0.880 | 0.576 | 0.375 | -12.077 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.194 | Likely Benign | -2.59 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.54 | Pathogenic | 0.00 | Affected | 0.1388 | 0.2289 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3614T>A | L1205Q 2D  AIThe SynGAP1 missense variant L1205Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus agrees. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.552471 | Binding | 0.880 | 0.576 | 0.375 | -14.466 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.453 | Likely Benign | -5.02 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.46 | Pathogenic | 0.00 | Affected | 0.1032 | 0.0558 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3614T>C | L1205P 2D AIThe SynGAP1 missense variant L1205P is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. All available in silico predictors classify the change as pathogenic: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence strongly indicates that the variant is pathogenic, which contradicts the current ClinVar designation of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.552471 | Binding | 0.880 | 0.576 | 0.375 | Uncertain | 1 | -16.878 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.536 | Likely Pathogenic | -5.91 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.45 | Pathogenic | 0.00 | Affected | 0.3559 | 0.1053 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||
| c.3614T>G | L1205R 2D AIThe SynGAP1 missense variant L1205R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) is likely pathogenic. Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.552471 | Binding | 0.880 | 0.576 | 0.375 | -16.706 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.451 | Likely Benign | -5.08 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.46 | Pathogenic | 0.00 | Affected | 0.1081 | 0.0558 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3620A>T | E1207V 2D AIThe SynGAP1 missense change E1207V is not reported in ClinVar (ClinVar ID = None) and has no entries in gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. AlphaMissense‑Optimized is uncertain, providing no definitive direction. High‑accuracy assessments show the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Likely Pathogenic, while AlphaMissense‑Optimized remains uncertain and Foldetta data are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for E1207V, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.604312 | Disordered | 0.562696 | Binding | 0.912 | 0.571 | 0.375 | -9.580 | Likely Pathogenic | 0.821 | Likely Pathogenic | Ambiguous | 0.342 | Likely Benign | -5.00 | Deleterious | 0.999 | Probably Damaging | 0.958 | Probably Damaging | 2.07 | Pathogenic | 0.00 | Affected | 0.0511 | 0.4439 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3622C>T | R1208W 2D AIThe SynGAP1 missense variant R1208W is recorded in gnomAD (variant ID 6‑33446614‑C‑T) but has no ClinVar entry. Prediction tools cluster into two groups: the single benign predictor REVEL, and a consensus of pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labels it “Likely Pathogenic.” No Foldetta stability result is available. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that R1208W is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.604312 | Disordered | 0.566942 | Binding | 0.899 | 0.569 | 0.375 | 6-33446614-C-T | 1 | 6.20e-7 | -12.124 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.192 | Likely Benign | -5.53 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.47 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1247 | 0.2962 | -3 | 2 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||
| c.3625C>A | L1209M 2D AIThe SynGAP1 missense variant L1209M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL and PROVEAN; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this: AlphaMissense‑Optimized remains uncertain, while Foldetta (a combined FoldX‑MD and Rosetta stability analysis) is not available for this residue. Given the predominance of pathogenic calls and the SGM‑Consensus result, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar entry exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.583711 | Binding | 0.899 | 0.574 | 0.375 | -10.605 | Likely Pathogenic | 0.934 | Likely Pathogenic | Ambiguous | 0.171 | Likely Benign | -1.66 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.0676 | 0.2486 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3625C>G | L1209V 2D AIThe SynGAP1 L1209V missense change is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and PROVEAN, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Taken together, the majority of evidence points toward a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.583711 | Binding | 0.899 | 0.574 | 0.375 | -9.962 | Likely Pathogenic | 0.953 | Likely Pathogenic | Ambiguous | 0.152 | Likely Benign | -2.39 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.54 | Pathogenic | 0.00 | Affected | 0.1383 | 0.2289 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3626T>A | L1209Q 2D AIThe SynGAP1 missense variant L1209Q is not listed in ClinVar and has no entry in gnomAD, indicating it is not a common population variant. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign outcome, whereas the remaining predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely pathogenic status. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.583711 | Binding | 0.899 | 0.574 | 0.375 | -12.820 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.379 | Likely Benign | -5.09 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.46 | Pathogenic | 0.00 | Affected | 0.1042 | 0.0558 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3626T>C | L1209P 2D AIThe SynGAP1 missense variant L1209P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is also pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as pathogenic; Foldetta results are unavailable. Based on the overwhelming agreement among pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.583711 | Binding | 0.899 | 0.574 | 0.375 | -17.259 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.448 | Likely Benign | -5.92 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.45 | Pathogenic | 0.00 | Affected | 0.3646 | 0.1053 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||
| c.3626T>G | L1209R 2D AIThe SynGAP1 missense variant L1209R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign outcome, whereas all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely pathogenic status. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its influence is unavailable. Overall, the preponderance of evidence from multiple prediction tools and high‑accuracy methods indicates that the variant is most likely pathogenic, with no conflict from ClinVar status (which is currently unreported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.583711 | Binding | 0.899 | 0.574 | 0.375 | -17.481 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.438 | Likely Benign | -5.12 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.46 | Pathogenic | 0.00 | Affected | 0.1129 | 0.0558 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3634T>A | S1212T 2D AIThe SynGAP1 missense variant S1212T is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; SGM Consensus is unavailable; Foldetta, which combines FoldX‑MD and Rosetta stability calculations, has no reported output for this variant. Overall, the balance of evidence slightly favors a pathogenic interpretation, but the single high‑accuracy benign prediction and the lack of a consensus from SGM and Foldetta leave the assessment uncertain. There is no conflict with ClinVar status, as the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | -4.972 | Likely Benign | 0.759 | Likely Pathogenic | Likely Benign | 0.147 | Likely Benign | -2.19 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.0997 | 0.4618 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3634T>C | S1212P 2D AIThe SynGAP1 missense variant S1212P is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Prediction tools that classify the variant as benign include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic effect. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus likewise reports likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of computational evidence points to the variant being most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | -13.336 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.239 | Likely Benign | -3.79 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.05 | Pathogenic | 0.00 | Affected | 0.1585 | 0.4370 | 1 | -1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||||||
| c.3634T>G | S1212A 2D AIThe SynGAP1 missense variant S1212A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM all predict a pathogenic outcome; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta results are unavailable. Consequently, the collective evidence points to a benign classification for S1212A, and this conclusion does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | -4.705 | Likely Benign | 0.403 | Ambiguous | Likely Benign | 0.109 | Likely Benign | -1.66 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 2.27 | Pathogenic | 0.00 | Affected | 0.3901 | 0.3823 | 1 | 1 | 2.6 | -16.00 | |||||||||||||||||||||||||||||||||||||||
| c.3635C>A | S1212Y 2D AIThe SynGAP1 missense variant S1212Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the convergence of multiple prediction algorithms, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | -12.186 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 0.304 | Likely Benign | -4.55 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.03 | Pathogenic | 0.00 | Affected | 0.0546 | 0.4291 | -3 | -2 | -0.5 | 76.10 | ||||||||||||||||||||||||||||||||||||||
| c.3635C>G | S1212C 2D AIThe SynGAP1 missense variant S1212C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, while the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all indicate pathogenicity. The SGM‑Consensus, a majority‑vote method from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. ESM1b is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available result for this variant. High‑accuracy tools therefore give a benign call from AlphaMissense‑Optimized, a pathogenic call from SGM‑Consensus, and no data from Foldetta. Overall, the preponderance of evidence points to a pathogenic effect, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | -7.938 | In-Between | 0.701 | Likely Pathogenic | Likely Benign | 0.245 | Likely Benign | -3.58 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.04 | Pathogenic | 0.00 | Affected | 0.0693 | 0.4631 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||
| c.3635C>T | S1212F 2D AIThe SynGAP1 missense variant S1212F is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) score—predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence from multiple independent predictors indicates that the variant is most likely pathogenic, which is consistent with its ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.566480 | Disordered | 0.548409 | Binding | 0.852 | 0.565 | 0.500 | Conflicting | 2 | -14.445 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.271 | Likely Benign | -4.52 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.03 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0503 | 0.4579 | -3 | -2 | 3.6 | 60.10 | ||||||||||||||||||||||||||||||||||
| c.3640C>T | R1214W 2D AIThe SynGAP1 missense variant R1214W is listed in ClinVar with an uncertain significance (ClinVar ID 1476244.0) and is present in the gnomAD database (gnomAD ID 6‑33446632‑C‑T). Functional prediction tools cluster into two groups: benign predictions come from REVEL and AlphaMissense‑Optimized, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized indicates a benign effect, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as likely pathogenic. No Foldetta stability analysis is available for this residue. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not conflict with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.506868 | Binding | 0.903 | 0.566 | 0.375 | Uncertain | 1 | 6-33446632-C-T | 2 | 1.24e-6 | -8.799 | Likely Pathogenic | 0.710 | Likely Pathogenic | Likely Benign | 0.143 | Likely Benign | -4.95 | Deleterious | 1.000 | Probably Damaging | 0.983 | Probably Damaging | 2.45 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1186 | 0.2367 | 2 | -3 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||
| c.3644A>T | K1215M 2D AIThe SynGAP1 missense variant K1215M is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign outcome, whereas all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely pathogenic status. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant. Overall, the preponderance of computational evidence indicates that K1215M is most likely pathogenic, and this conclusion is consistent with the absence of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.503613 | Binding | 0.888 | 0.568 | 0.375 | -11.140 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 0.202 | Likely Benign | -4.06 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.35 | Pathogenic | 0.00 | Affected | 0.0922 | 0.3085 | 0 | -1 | 5.8 | 3.02 | ||||||||||||||||||||||||||||||||||||||
| c.3646C>A | L1216M 2D AIThe SynGAP1 missense variant L1216M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs 2 benign), and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward a pathogenic interpretation. This conclusion is not contradicted by ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.580690 | Disordered | 0.504713 | Binding | 0.863 | 0.563 | 0.250 | -6.590 | Likely Benign | 0.806 | Likely Pathogenic | Ambiguous | 0.142 | Likely Benign | -1.40 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.16 | Pathogenic | 0.00 | Affected | 0.0612 | 0.2092 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3646C>G | L1216V 2D AIThe SynGAP1 missense variant L1216V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two agreement groups: benign predictions come from REVEL and PROVEAN, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. Two tools—ESM1b and AlphaMissense‑Optimized—return uncertain results. High‑accuracy assessment further shows that AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.580690 | Disordered | 0.504713 | Binding | 0.863 | 0.563 | 0.250 | -7.842 | In-Between | 0.861 | Likely Pathogenic | Ambiguous | 0.126 | Likely Benign | -2.26 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 2.20 | Pathogenic | 0.00 | Affected | 0.1294 | 0.1883 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3647T>A | L1216Q 2D AIThe SynGAP1 missense variant L1216Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as pathogenic; Foldetta results are not available. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.580690 | Disordered | 0.504713 | Binding | 0.863 | 0.563 | 0.250 | -8.731 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.404 | Likely Benign | -4.12 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.23 | Pathogenic | 0.00 | Affected | 0.0939 | 0.0488 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3647T>C | L1216P 2D AIThe SynGAP1 missense variant L1216P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic. No tool predicts a benign outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as likely pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions strongly suggests that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.580690 | Disordered | 0.504713 | Binding | 0.863 | 0.563 | 0.250 | -16.029 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.524 | Likely Pathogenic | -5.28 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.13 | Pathogenic | 0.00 | Affected | 0.3281 | 0.1048 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||
| c.3647T>G | L1216R 2D AIThe SynGAP1 missense variant L1216R is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus agrees. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.580690 | Disordered | 0.504713 | Binding | 0.863 | 0.563 | 0.250 | -9.700 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.387 | Likely Benign | -4.39 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.16 | Pathogenic | 0.00 | Affected | 0.1071 | 0.0488 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3649G>A | E1217K 2D AIThe SynGAP1 missense variant E1217K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. The high‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is classified as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.599170 | Disordered | 0.493043 | Uncertain | 0.877 | 0.563 | 0.250 | -12.869 | Likely Pathogenic | 0.862 | Likely Pathogenic | Ambiguous | 0.306 | Likely Benign | -3.09 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.1826 | 0.5272 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3649G>C | E1217Q 2D AIThe SynGAP1 missense variant E1217Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of tools (five pathogenic vs. three benign) and the SGM Consensus support a pathogenic classification, which does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.599170 | Disordered | 0.493043 | Uncertain | 0.877 | 0.563 | 0.250 | -8.887 | Likely Pathogenic | 0.398 | Ambiguous | Likely Benign | 0.219 | Likely Benign | -2.16 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.38 | Pathogenic | 0.00 | Affected | 0.0949 | 0.4745 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3650A>C | E1217A 2D AIThe SynGAP1 missense variant E1217A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments show a split: AlphaMissense‑Optimized reports benign, while the SGM‑Consensus (majority vote) reports likely pathogenic; Foldetta results are unavailable. Overall, the preponderance of evidence from multiple independent predictors points to a pathogenic effect for E1217A, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.599170 | Disordered | 0.493043 | Uncertain | 0.877 | 0.563 | 0.250 | -10.446 | Likely Pathogenic | 0.605 | Likely Pathogenic | Likely Benign | 0.261 | Likely Benign | -4.52 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.38 | Pathogenic | 0.00 | Affected | 0.2585 | 0.4401 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3650A>G | E1217G 2D AIThe SynGAP1 missense variant E1217G is reported in gnomAD (ID 6‑33446642‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for E1217G. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.599170 | Disordered | 0.493043 | Uncertain | 0.877 | 0.563 | 0.250 | 6-33446642-A-G | -10.803 | Likely Pathogenic | 0.620 | Likely Pathogenic | Likely Benign | 0.331 | Likely Benign | -5.38 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.35 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2205 | 0.4927 | -2 | 0 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||
| c.3650A>T | E1217V 2D AIThe SynGAP1 missense variant E1217V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: REVEL scores the variant as benign, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all predict pathogenicity. Grouping by consensus, the majority of tools (seven) support a pathogenic effect, while only one tool (REVEL) indicates benign. High‑accuracy assessments further reinforce a deleterious interpretation: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic, and AlphaMissense‑Optimized remains uncertain. No Foldetta stability analysis is available, so it does not influence the conclusion. Overall, the computational evidence overwhelmingly suggests that E1217V is pathogenic, a finding that aligns with its lack of ClinVar annotation and gnomAD presence. Thus, the variant is most likely pathogenic, and this prediction is consistent with its absence from ClinVar and gnomAD. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.599170 | Disordered | 0.493043 | Uncertain | 0.877 | 0.563 | 0.250 | -12.098 | Likely Pathogenic | 0.843 | Likely Pathogenic | Ambiguous | 0.351 | Likely Benign | -5.48 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.33 | Pathogenic | 0.00 | Affected | 0.0579 | 0.5348 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3651A>C | E1217D 2D AIThe SynGAP1 missense variant E1217D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also leans benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Taken together, the preponderance of evidence points to a benign impact for E1217D, and this conclusion is consistent with the absence of any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.493043 | Uncertain | 0.877 | 0.563 | 0.250 | -5.983 | Likely Benign | 0.705 | Likely Pathogenic | Likely Benign | 0.114 | Likely Benign | -2.19 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.51 | Benign | 0.00 | Affected | 0.1745 | 0.3146 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3651A>T | E1217D 2D AIThe SynGAP1 missense variant E1217D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also leans benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Taken together, the preponderance of evidence points to a benign impact for E1217D, and this conclusion is consistent with the absence of any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.599170 | Disordered | 0.493043 | Uncertain | 0.877 | 0.563 | 0.250 | -5.983 | Likely Benign | 0.705 | Likely Pathogenic | Likely Benign | 0.114 | Likely Benign | -2.19 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.51 | Benign | 0.00 | Affected | 0.1745 | 0.3146 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3652G>A | E1218K 2D AIThe SynGAP1 missense variant E1218K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—all predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus indicates it is likely pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from both general and high‑accuracy prediction tools indicates that E1218K is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.483050 | Uncertain | 0.898 | 0.565 | 0.375 | -8.932 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 0.336 | Likely Benign | -3.12 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.28 | Pathogenic | 0.00 | Affected | 0.1672 | 0.4024 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3652G>C | E1218Q 2D AIThe SynGAP1 E1218Q missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and ESM1b, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are less decisive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and thus inconclusive; Foldetta results are not available. Consequently, the majority of conventional predictors lean toward pathogenicity, but the most reliable tools do not provide a clear verdict. The variant is therefore most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.595080 | Disordered | 0.483050 | Uncertain | 0.898 | 0.565 | 0.375 | -6.490 | Likely Benign | 0.857 | Likely Pathogenic | Ambiguous | 0.226 | Likely Benign | -2.09 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.0784 | 0.3666 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3653A>C | E1218A 2D AIThe SynGAP1 missense variant E1218A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus) predict a pathogenic outcome. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic effect. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the overall consensus of the available predictions, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.483050 | Uncertain | 0.898 | 0.565 | 0.375 | -3.698 | Likely Benign | 0.819 | Likely Pathogenic | Ambiguous | 0.370 | Likely Benign | -4.75 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.25 | Pathogenic | 0.00 | Affected | 0.3005 | 0.3969 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3653A>G | E1218G 2D AIThe SynGAP1 missense variant E1218G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all indicate pathogenicity. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely pathogenic. AlphaMissense‑Optimized returns an uncertain result, and no Foldetta (FoldX‑MD/Rosetta) stability assessment is available. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant has not been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.483050 | Uncertain | 0.898 | 0.565 | 0.375 | -5.595 | Likely Benign | 0.864 | Likely Pathogenic | Ambiguous | 0.413 | Likely Benign | -5.61 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.22 | Pathogenic | 0.00 | Affected | 0.2496 | 0.4095 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||
| c.3653A>T | E1218V 2D AISynGAP1 missense variant E1218V is listed in ClinVar with an uncertain significance (ClinVar ID 1015602.0) and is not reported in gnomAD. Functional prediction tools show a split opinion: benign predictions come from REVEL and ESM1b, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. When the high‑accuracy consensus is considered, AlphaMissense‑Optimized is uncertain, but the SGM‑Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as likely pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this change. Overall, the majority of evidence points toward a pathogenic effect, which is consistent with the ClinVar designation of uncertain significance rather than a benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.595080 | Disordered | 0.483050 | Uncertain | 0.898 | 0.565 | 0.375 | Uncertain | 2 | -5.647 | Likely Benign | 0.936 | Likely Pathogenic | Ambiguous | 0.418 | Likely Benign | -5.68 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.21 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0491 | 0.4120 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||
| c.3654G>C | E1218D 2D AIThe SynGAP1 missense variant E1218D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, and no Foldetta stability data are available. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.595080 | Disordered | 0.483050 | Uncertain | 0.898 | 0.565 | 0.375 | -3.574 | Likely Benign | 0.517 | Ambiguous | Likely Benign | 0.138 | Likely Benign | -2.42 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1485 | 0.2475 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3654G>T | E1218D 2D AIThe SynGAP1 missense variant E1218D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, and no Foldetta stability data are available. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.595080 | Disordered | 0.483050 | Uncertain | 0.898 | 0.565 | 0.375 | -3.574 | Likely Benign | 0.517 | Ambiguous | Likely Benign | 0.138 | Likely Benign | -2.42 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1485 | 0.2475 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3655T>A | Y1219N 2D  AIThe SynGAP1 missense variant Y1219N is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess functional impact largely agree on a deleterious effect: SIFT, polyPhen‑2 (HumDiv and HumVar), PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while the only benign prediction comes from REVEL. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as “Likely Pathogenic.” High‑accuracy assessments further support a pathogenic interpretation: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta results are not available, so they do not influence the overall assessment. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.613573 | Disordered | 0.474748 | Uncertain | 0.855 | 0.557 | 0.375 | -11.679 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.366 | Likely Benign | -6.41 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.15 | Pathogenic | 0.00 | Affected | 0.2478 | 0.0573 | -2 | -2 | -2.2 | -49.07 | ||||||||||||||||||||||||||||||||||||||
| c.3655T>C | Y1219H 2D  AIThe SynGAP1 missense variant Y1219H is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic)—all predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence from both general and high‑accuracy tools indicates that the variant is most likely pathogenic, which does not contradict its current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.613573 | Disordered | 0.474748 | Uncertain | 0.855 | 0.557 | 0.375 | Uncertain | 1 | -9.511 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.363 | Likely Benign | -3.62 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.15 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2234 | 0.0573 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||
| c.3655T>G | Y1219D 2D  AIThe SynGAP1 missense variant Y1219D is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta results are not available for this variant. Based on the preponderance of pathogenic predictions, Y1219D is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.613573 | Disordered | 0.474748 | Uncertain | 0.855 | 0.557 | 0.375 | -15.860 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.443 | Likely Benign | -7.11 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.15 | Pathogenic | 0.00 | Affected | 0.4549 | 0.0573 | -4 | -3 | -2.2 | -48.09 | ||||||||||||||||||||||||||||||||||||||
| c.3656A>C | Y1219S 2D  AIThe SynGAP1 missense variant Y1219S is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.613573 | Disordered | 0.474748 | Uncertain | 0.855 | 0.557 | 0.375 | -11.227 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.424 | Likely Benign | -6.05 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.18 | Pathogenic | 0.00 | Affected | 0.5047 | 0.1403 | Weaken | -3 | -2 | 0.5 | -76.10 | |||||||||||||||||||||||||||||||||||||
| c.3656A>G | Y1219C 2D  AIThe SynGAP1 missense variant Y1219C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact. ESM1b is uncertain and does not contribute to a consensus. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic; Foldetta results are unavailable. Overall, the evidence strongly favors a pathogenic classification for Y1219C, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.613573 | Disordered | 0.474748 | Uncertain | 0.855 | 0.557 | 0.375 | -7.776 | In-Between | 0.984 | Likely Pathogenic | Likely Pathogenic | 0.310 | Likely Benign | -5.41 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.23 | Pathogenic | 0.00 | Affected | 0.3446 | 0.1672 | 0 | -2 | 3.8 | -60.04 | ||||||||||||||||||||||||||||||||||||||
| c.3656A>T | Y1219F 2D  AIThe SynGAP1 missense variant Y1219F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments show that the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome, while AlphaMissense‑Optimized remains uncertain. No Foldetta stability data are available, so it does not influence the assessment. Overall, the preponderance of evidence points to a pathogenic effect for Y1219F, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.613573 | Disordered | 0.474748 | Uncertain | 0.855 | 0.557 | 0.375 | -7.202 | In-Between | 0.837 | Likely Pathogenic | Ambiguous | 0.272 | Likely Benign | -2.79 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 2.23 | Pathogenic | 0.00 | Affected | 0.1998 | 0.2583 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||||||||||
| c.3658G>A | E1220K 2D AIThe SynGAP1 missense variant E1220K is listed in gnomAD (6‑33446650‑G‑A) but has no ClinVar entry. Prediction tools that agree on benign impact include REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict pathogenicity. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized scores the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that E1220K is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.444845 | Uncertain | 0.881 | 0.551 | 0.375 | 6-33446650-G-A | 1 | 6.20e-7 | -12.478 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.415 | Likely Benign | -3.46 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.63 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1862 | 0.4046 | 1 | 0 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||
| c.3658G>C | E1220Q 2D AIThe SynGAP1 missense variant E1220Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta data are unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.444845 | Uncertain | 0.881 | 0.551 | 0.375 | -12.066 | Likely Pathogenic | 0.846 | Likely Pathogenic | Ambiguous | 0.276 | Likely Benign | -2.59 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.62 | Pathogenic | 0.00 | Affected | 0.0896 | 0.3997 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3659A>C | E1220A 2D AIThe SynGAP1 missense variant E1220A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus likewise indicates Likely Pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence from multiple in silico predictors indicates that E1220A is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.444845 | Uncertain | 0.881 | 0.551 | 0.375 | -12.798 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.407 | Likely Benign | -5.12 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.62 | Pathogenic | 0.00 | Affected | 0.3345 | 0.4105 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3659A>G | E1220G 2D AIThe SynGAP1 missense variant E1220G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign change, whereas all other evaluated algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence indicates that E1220G is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.444845 | Uncertain | 0.881 | 0.551 | 0.375 | -12.121 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.454 | Likely Benign | -6.05 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 1.61 | Pathogenic | 0.00 | Affected | 0.2791 | 0.4030 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||
| c.3659A>T | E1220V 2D AIThe SynGAP1 missense variant E1220V is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that E1220V is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.444845 | Uncertain | 0.881 | 0.551 | 0.375 | -15.193 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.444 | Likely Benign | -6.05 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.59 | Pathogenic | 0.00 | Affected | 0.0552 | 0.4452 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3660G>C | E1220D 2D AIThe missense change E1220D occurs in the coiled‑coil domain of SynGAP1. ClinVar contains no entry for this variant and it is absent from gnomAD. Functional prediction tools cluster into two groups: a single benign call from REVEL, and a consensus of pathogenic predictions from the remaining methods (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized). The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN, also reports a likely pathogenic outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus likewise indicates pathogenic. Foldetta stability analysis is not available for this variant. Taken together, the preponderance of evidence points to a pathogenic effect, and this assessment does not conflict with the absence of a ClinVar classification. The variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.444845 | Uncertain | 0.881 | 0.551 | 0.375 | -8.820 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.178 | Likely Benign | -2.59 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 1.66 | Pathogenic | 0.00 | Affected | 0.1597 | 0.2810 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3660G>T | E1220D 2D AIThe SynGAP1 missense variant E1220D is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.444845 | Uncertain | 0.881 | 0.551 | 0.375 | -8.820 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.179 | Likely Benign | -2.59 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 1.66 | Pathogenic | 0.00 | Affected | 0.1597 | 0.2810 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3668T>G | L1223R 2D AIThe SynGAP1 missense variant L1223R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as likely pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the preponderance of computational evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.608892 | Disordered | 0.436267 | Uncertain | 0.868 | 0.540 | 0.375 | -15.396 | Likely Pathogenic | 0.966 | Likely Pathogenic | Likely Pathogenic | 0.317 | Likely Benign | -4.14 | Deleterious | 0.999 | Probably Damaging | 0.986 | Probably Damaging | 1.46 | Pathogenic | 0.00 | Affected | 0.1078 | 0.0919 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3676C>A | Q1226K 2D AIThe SynGAP1 missense variant Q1226K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic impact. AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. High‑accuracy evidence therefore points to a likely pathogenic outcome: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus indicates likely pathogenic, and Foldetta data are missing. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -13.233 | Likely Pathogenic | 0.890 | Likely Pathogenic | Ambiguous | 0.212 | Likely Benign | -3.16 | Deleterious | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 1.82 | Pathogenic | 0.00 | Affected | 0.1334 | 0.3199 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||||||||||||
| c.3676C>G | Q1226E 2D AIThe SynGAP1 missense variant Q1226E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, PROVEAN, and AlphaMissense‑Optimized, whereas pathogenic predictions are reported by polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely pathogenic verdict (3/4 pathogenic votes). High‑accuracy assessments further support this: AlphaMissense‑Optimized indicates a benign effect, whereas the SGM‑Consensus remains pathogenic; Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points toward a pathogenic impact, which is consistent with the SGM‑Consensus prediction and does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -11.526 | Likely Pathogenic | 0.625 | Likely Pathogenic | Likely Benign | 0.178 | Likely Benign | -2.13 | Neutral | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.1005 | 0.2297 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3677A>C | Q1226P 2D AIThe SynGAP1 missense variant Q1226P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as likely pathogenic; Foldetta’s protein‑folding stability analysis is unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -13.176 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.418 | Likely Benign | -4.72 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 1.75 | Pathogenic | 0.00 | Affected | 0.1829 | 0.4187 | 0 | -1 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||||||
| c.3677A>G | Q1226R 2D AIThe SynGAP1 missense change Q1226R occurs in a coiled‑coil domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus) all predict a pathogenic impact. The high‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain; the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Pathogenic”; Foldetta data are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -12.260 | Likely Pathogenic | 0.883 | Likely Pathogenic | Ambiguous | 0.301 | Likely Benign | -3.16 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.1166 | 0.1234 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3677A>T | Q1226L 2D AIThe SynGAP1 missense variant Q1226L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious interpretation: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Pathogenic,” AlphaMissense‑Optimized is classified as “Uncertain,” and Foldetta’s protein‑folding stability analysis is unavailable. Taken together, the preponderance of evidence points to a pathogenic effect for Q1226L. This conclusion is not contradicted by ClinVar status, as the variant is currently unreported in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -11.122 | Likely Pathogenic | 0.879 | Likely Pathogenic | Ambiguous | 0.353 | Likely Benign | -5.62 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.77 | Pathogenic | 0.00 | Affected | 0.0493 | 0.4282 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3678G>C | Q1226H 2D AIThe SynGAP1 missense variant Q1226H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic impact. AlphaMissense‑Optimized is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the collective evidence, the variant is most likely pathogenic; this assessment does not contradict any ClinVar status, as no ClinVar entry exists for Q1226H. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -8.363 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.241 | Likely Benign | -3.62 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 1.75 | Pathogenic | 0.00 | Affected | 0.0913 | 0.3230 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3678G>T | Q1226H 2D AIThe SynGAP1 missense variant Q1226H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic impact. AlphaMissense‑Optimized is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the collective evidence, the variant is most likely pathogenic; this assessment does not contradict any ClinVar status, as no ClinVar entry exists for Q1226H. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.432206 | Uncertain | 0.850 | 0.547 | 0.250 | -8.363 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.241 | Likely Benign | -3.62 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 1.75 | Pathogenic | 0.00 | Affected | 0.0913 | 0.3230 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3679G>A | E1227K 2D AIThe SynGAP1 missense variant E1227K is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.513880 | Disordered | 0.433399 | Uncertain | 0.860 | 0.544 | 0.500 | -11.825 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.280 | Likely Benign | -2.94 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1661 | 0.6348 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3679G>C | E1227Q 2D AIThe SynGAP1 missense variant E1227Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, while the SGM‑Consensus remains pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple in silico predictors indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.513880 | Disordered | 0.433399 | Uncertain | 0.860 | 0.544 | 0.500 | -9.212 | Likely Pathogenic | 0.860 | Likely Pathogenic | Ambiguous | 0.277 | Likely Benign | -2.28 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.0761 | 0.6204 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3680A>C | E1227A 2D AIThe SynGAP1 missense variant E1227A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled “Likely Pathogenic.” The Foldetta protein‑folding stability analysis is unavailable for this variant. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.513880 | Disordered | 0.433399 | Uncertain | 0.860 | 0.544 | 0.500 | -9.111 | Likely Pathogenic | 0.961 | Likely Pathogenic | Likely Pathogenic | 0.341 | Likely Benign | -4.63 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.29 | Pathogenic | 0.00 | Affected | 0.3142 | 0.6025 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3680A>G | E1227G 2D AIThe SynGAP1 missense variant E1227G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as Likely Pathogenic. AlphaMissense‑Optimized yields an uncertain result, and Foldetta (FoldX‑MD/Rosetta stability analysis) is not available for this variant. Overall, the preponderance of evidence from high‑accuracy predictors and consensus methods indicates that E1227G is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.513880 | Disordered | 0.433399 | Uncertain | 0.860 | 0.544 | 0.500 | -9.328 | Likely Pathogenic | 0.903 | Likely Pathogenic | Ambiguous | 0.336 | Likely Benign | -5.26 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.28 | Pathogenic | 0.00 | Affected | 0.2725 | 0.5550 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||
| c.3680A>T | E1227V 2D AIThe SynGAP1 missense variant E1227V is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess the variant’s effect fall into two groups: the single benign prediction comes from REVEL, whereas all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—classify it as pathogenic or likely pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus also indicates likely pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from multiple independent prediction tools and high‑accuracy methods indicates that E1227V is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.513880 | Disordered | 0.433399 | Uncertain | 0.860 | 0.544 | 0.500 | -12.852 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.355 | Likely Benign | -5.49 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.25 | Pathogenic | 0.00 | Affected | 0.0405 | 0.6559 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3681A>C | E1227D 2D AIThe SynGAP1 missense variant E1227D is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority of the high‑accuracy tools) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence, including the high‑accuracy tools, points to the variant being most likely benign, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.513880 | Disordered | 0.433399 | Uncertain | 0.860 | 0.544 | 0.500 | -5.675 | Likely Benign | 0.777 | Likely Pathogenic | Likely Benign | 0.172 | Likely Benign | -1.67 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.60 | Benign | 0.00 | Affected | 0.1363 | 0.4161 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3681A>T | E1227D 2D AIThe SynGAP1 missense variant E1227D is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority of the high‑accuracy tools) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence, including the high‑accuracy tools, points to the variant being most likely benign, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.513880 | Disordered | 0.433399 | Uncertain | 0.860 | 0.544 | 0.500 | -5.675 | Likely Benign | 0.777 | Likely Pathogenic | Likely Benign | 0.170 | Likely Benign | -1.67 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.60 | Benign | 0.00 | Affected | 0.1363 | 0.4161 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3683A>G | E1228G 2D AIThe SynGAP1 E1228G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions therefore show one benign call (AlphaMissense‑Optimized) and no decisive pathogenic evidence. Overall, the majority of standard tools (5 pathogenic vs 4 benign) indicate a pathogenic likelihood, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.517562 | Disordered | 0.447051 | Uncertain | 0.892 | 0.546 | 0.500 | -6.273 | Likely Benign | 0.306 | Likely Benign | Likely Benign | 0.249 | Likely Benign | -4.54 | Deleterious | 0.980 | Probably Damaging | 0.721 | Possibly Damaging | 2.44 | Pathogenic | 0.00 | Affected | 0.2598 | 0.4030 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||||||
| c.3683A>T | E1228V 2D AIThe SynGAP1 missense variant E1228V is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL and AlphaMissense‑Optimized, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and the SGM‑Consensus score, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further separate the evidence: AlphaMissense‑Optimized indicates a benign effect, whereas the SGM‑Consensus, derived from a consensus of four high‑confidence predictors, flags the variant as pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this change. Overall, the preponderance of pathogenic predictions, including the SGM‑Consensus, outweighs the benign calls. Therefore, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.517562 | Disordered | 0.447051 | Uncertain | 0.892 | 0.546 | 0.500 | -8.077 | Likely Pathogenic | 0.440 | Ambiguous | Likely Benign | 0.293 | Likely Benign | -4.55 | Deleterious | 0.980 | Probably Damaging | 0.833 | Possibly Damaging | 2.43 | Pathogenic | 0.00 | Affected | 0.0470 | 0.4252 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3694A>C | K1232Q 2D AIThe SynGAP1 missense variant K1232Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of individual predictors (five pathogenic vs four benign) lean toward a pathogenic interpretation. Thus, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict the ClinVar status, which has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -6.905 | Likely Benign | 0.247 | Likely Benign | Likely Benign | 0.102 | Likely Benign | -2.86 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.3457 | 0.1419 | 1 | 1 | 0.4 | -0.04 | |||||||||||||||||||||||||||||||||||||||
| c.3694A>G | K1232E 2D AIThe SynGAP1 missense variant K1232E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as benign, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels it as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -10.690 | Likely Pathogenic | 0.695 | Likely Pathogenic | Likely Benign | 0.165 | Likely Benign | -2.76 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.13 | Pathogenic | 0.00 | Affected | 0.2965 | 0.1039 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3695A>C | K1232T 2D AIThe SynGAP1 missense variant K1232T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains pathogenic; Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -9.276 | Likely Pathogenic | 0.568 | Likely Pathogenic | Likely Benign | 0.189 | Likely Benign | -4.49 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.1846 | 0.3196 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||||||||
| c.3695A>G | K1232R 2D AIThe SynGAP1 missense variant K1232R is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of evidence—including the high‑accuracy tools—points to a benign impact. This conclusion is consistent with the absence of ClinVar annotation and gnomAD data, so there is no contradiction with existing clinical databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -3.146 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.077 | Likely Benign | -1.83 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.3515 | 0.0945 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3695A>T | K1232I 2D  AIThe SynGAP1 K1232I missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. The high‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is classified as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect; this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -12.225 | Likely Pathogenic | 0.896 | Likely Pathogenic | Ambiguous | 0.197 | Likely Benign | -5.98 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.08 | Pathogenic | 0.00 | Affected | 0.0778 | 0.3321 | -2 | -3 | 8.4 | -15.01 | ||||||||||||||||||||||||||||||||||||||
| c.3696A>C | K1232N 2D AIThe SynGAP1 missense variant K1232N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: REVEL scores the variant as benign, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all predict it to be pathogenic. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized returns an uncertain result, and Foldetta data are not available. Based on the overall evidence, the variant is most likely pathogenic, which is consistent with the lack of ClinVar annotation and gnomAD absence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -8.657 | Likely Pathogenic | 0.834 | Likely Pathogenic | Ambiguous | 0.167 | Likely Benign | -3.72 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.2913 | 0.1464 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3696A>T | K1232N 2D AIThe SynGAP1 missense variant K1232N is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus—consistently predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.505461 | Disordered | 0.542907 | Binding | 0.894 | 0.535 | 0.125 | -8.657 | Likely Pathogenic | 0.834 | Likely Pathogenic | Ambiguous | 0.167 | Likely Benign | -3.72 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.2913 | 0.1464 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3698T>A | I1233N 2D AIThe SynGAP1 I1233N missense variant is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and FATHMM. All other evaluated tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic impact. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -9.586 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.193 | Likely Benign | -4.36 | Deleterious | 0.995 | Probably Damaging | 0.913 | Probably Damaging | 2.52 | Benign | 0.00 | Affected | 0.0879 | 0.0340 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3698T>G | I1233S 2D AIThe SynGAP1 missense variant I1233S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and FATHMM, whereas the majority of other predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized—classify the change as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic verdict. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus likewise indicates likely pathogenic. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not conflict with the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -8.066 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.184 | Likely Benign | -3.60 | Deleterious | 0.946 | Possibly Damaging | 0.673 | Possibly Damaging | 2.53 | Benign | 0.00 | Affected | 0.2848 | 0.0910 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.36C>A | S12R 2D AIThe SynGAP1 missense variant S12R is present in gnomAD (ID 6‑33420300‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score all report benign or likely benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors benign. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | 6-33420300-C-A | -4.033 | Likely Benign | 0.500 | Ambiguous | Likely Benign | 0.097 | Likely Benign | -0.30 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.09 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0944 | 0.3678 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.36C>G | S12R 2D AIThe SynGAP1 missense variant S12R is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33420300‑C‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign classification; Foldetta’s protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this does not contradict the ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | Uncertain | 1 | 6-33420300-C-G | 4 | 2.59e-6 | -4.033 | Likely Benign | 0.500 | Ambiguous | Likely Benign | 0.097 | Likely Benign | -0.30 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.09 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0944 | 0.3678 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||
| c.3701T>G | L1234R 2D AIThe SynGAP1 missense change L1234R occurs in a coiled‑coil domain. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that agree on a benign effect are limited to REVEL, which scores the variant as benign. All other evaluated predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—classify the variant as pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.599170 | Disordered | 0.575096 | Binding | 0.844 | 0.527 | 0.125 | -15.015 | Likely Pathogenic | 0.912 | Likely Pathogenic | Ambiguous | 0.211 | Likely Benign | -4.52 | Deleterious | 0.997 | Probably Damaging | 0.939 | Probably Damaging | 1.46 | Pathogenic | 0.00 | Affected | 0.1061 | 0.0849 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3704T>A | M1235K 2D AIThe SynGAP1 missense variant M1235K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of tools, including the high‑accuracy methods, supports a benign classification. This prediction is consistent with the lack of ClinVar evidence and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.690604 | Disordered | 0.577958 | Binding | 0.872 | 0.532 | 0.125 | -6.348 | Likely Benign | 0.269 | Likely Benign | Likely Benign | 0.109 | Likely Benign | -2.37 | Neutral | 0.270 | Benign | 0.089 | Benign | 2.66 | Benign | 0.00 | Affected | 0.1282 | 0.0656 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||||||||||
| c.3704T>G | M1235R 2D AIThe SynGAP1 missense variant M1235R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.690604 | Disordered | 0.577958 | Binding | 0.872 | 0.532 | 0.125 | -5.410 | Likely Benign | 0.246 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -2.37 | Neutral | 0.270 | Benign | 0.089 | Benign | 2.64 | Benign | 0.00 | Affected | 0.1505 | 0.0757 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||||||||||
| c.3707A>T | Q1236L 2D AIThe SynGAP1 missense variant Q1236L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, while Foldetta results are unavailable. Overall, the majority of high‑confidence predictions lean toward a benign impact, and there is no conflict with ClinVar status. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.680603 | Disordered | 0.567914 | Binding | 0.883 | 0.537 | 0.125 | -6.682 | Likely Benign | 0.409 | Ambiguous | Likely Benign | 0.362 | Likely Benign | -4.51 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.66 | Benign | 0.00 | Affected | 0.0507 | 0.3805 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3709T>A | Y1237N 2D AIThe SynGAP1 missense variant Y1237N is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—consistently predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as likely pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.653063 | Disordered | 0.563444 | Binding | 0.842 | 0.535 | 0.125 | -10.505 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.413 | Likely Benign | -7.18 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.44 | Pathogenic | 0.00 | Affected | 0.2428 | 0.0173 | -2 | -2 | -2.2 | -49.07 | ||||||||||||||||||||||||||||||||||||||
| c.3709T>C | Y1237H 2D AIThe SynGAP1 missense variant Y1237H is not reported in ClinVar (ClinVar ID = None) and has no entries in gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic. ESM1b is uncertain, and Foldetta results are unavailable. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. With no conflicting ClinVar evidence, the collective predictions strongly suggest that Y1237H is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.653063 | Disordered | 0.563444 | Binding | 0.842 | 0.535 | 0.125 | -7.985 | In-Between | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.383 | Likely Benign | -4.09 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.45 | Pathogenic | 0.00 | Affected | 0.2221 | 0.0173 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||||||||||
| c.3709T>G | Y1237D 2D AIThe SynGAP1 missense variant Y1237D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Functional prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as likely pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta results are unavailable, so no additional folding‑stability evidence is provided. Overall, the preponderance of predictions indicates that the variant is most likely pathogenic, and this conclusion is consistent with the absence of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.653063 | Disordered | 0.563444 | Binding | 0.842 | 0.535 | 0.125 | -12.849 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.447 | Likely Benign | -8.04 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.44 | Pathogenic | 0.00 | Affected | 0.4353 | 0.0173 | -4 | -3 | -2.2 | -48.09 | ||||||||||||||||||||||||||||||||||||||
| c.3710A>C | Y1237S 2D AIThe SynGAP1 missense variant Y1237S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—classify the variant as pathogenic. The SGM‑Consensus, a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, therefore also predicts pathogenicity. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) is pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of any benign consensus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.653063 | Disordered | 0.563444 | Binding | 0.842 | 0.535 | 0.125 | -10.349 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.463 | Likely Benign | -7.21 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.45 | Pathogenic | 0.00 | Affected | 0.5127 | 0.0891 | Weaken | -3 | -2 | 0.5 | -76.10 | |||||||||||||||||||||||||||||||||||||
| c.3710A>G | Y1237C 2D AIThe SynGAP1 missense variant Y1237C is listed in gnomAD (variant ID 6‑33446702‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: the single benign prediction comes from REVEL, while all other evaluated algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic effect. High‑accuracy assessments reinforce this trend: AlphaMissense‑Optimized reports a pathogenic outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Pathogenic.” No Foldetta stability result is available, so it does not influence the overall assessment. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.653063 | Disordered | 0.563444 | Binding | 0.842 | 0.535 | 0.125 | 6-33446702-A-G | 1 | 6.20e-7 | -8.600 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.429 | Likely Benign | -7.15 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.43 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3354 | 0.1549 | -2 | 0 | 3.8 | -60.04 | |||||||||||||||||||||||||||||||||
| c.3710A>T | Y1237F 2D AIThe SynGAP1 missense variant Y1237F is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Prediction tools that classify the variant as benign include REVEL and ESM1b, whereas the majority of other in silico predictors—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default—label it pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely pathogenic outcome. High‑accuracy assessments are mixed: AlphaMissense‑Optimized returns an uncertain result, SGM‑Consensus confirms a likely pathogenic prediction, and Foldetta data are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar entry exists for Y1237F. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.653063 | Disordered | 0.563444 | Binding | 0.842 | 0.535 | 0.125 | -4.494 | Likely Benign | 0.864 | Likely Pathogenic | Ambiguous | 0.268 | Likely Benign | -3.12 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 1.55 | Pathogenic | 0.00 | Affected | 0.1938 | 0.2424 | 7 | 3 | 4.1 | -16.00 | ||||||||||||||||||||||||||||||||||||||
| c.3715G>A | A1239T 2D AIThe SynGAP1 missense variant A1239T is not reported in ClinVar and has no gnomAD entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the consensus score from SGM‑Consensus is “Likely Benign.” In contrast, two sequence‑based predictors flag it as pathogenic: SIFT and FATHMM. When considering the high‑accuracy subset, AlphaMissense‑Optimized remains benign and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a benign outcome; Foldetta data are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -3.570 | Likely Benign | 0.070 | Likely Benign | Likely Benign | 0.030 | Likely Benign | -1.62 | Neutral | 0.270 | Benign | 0.136 | Benign | 2.37 | Pathogenic | 0.00 | Affected | 0.0957 | 0.5384 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.3715G>C | A1239P 2D AIThe SynGAP1 missense variant A1239P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL and polyPhen‑2 HumVar, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that A1239P is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation (none). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -11.055 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.085 | Likely Benign | -2.62 | Deleterious | 0.784 | Possibly Damaging | 0.390 | Benign | 2.32 | Pathogenic | 0.00 | Affected | 0.1438 | 0.3450 | 1 | -1 | -3.4 | 26.04 | ||||||||||||||||||||||||||||||||||||||
| c.3715G>T | A1239S 2D AIThe SynGAP1 missense variant A1239S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while SIFT and FATHMM predict it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and no Foldetta stability data are available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -2.847 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.019 | Likely Benign | -0.95 | Neutral | 0.003 | Benign | 0.005 | Benign | 2.39 | Pathogenic | 0.00 | Affected | 0.1974 | 0.4095 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||||||||||
| c.3716C>A | A1239D 2D AIThe SynGAP1 missense variant A1239D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes). Foldetta results are unavailable. Overall, the majority of evidence (six benign vs. three pathogenic predictions) supports a benign classification. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -6.177 | Likely Benign | 0.326 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -2.60 | Deleterious | 0.270 | Benign | 0.136 | Benign | 2.36 | Pathogenic | 0.00 | Affected | 0.1521 | 0.2062 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||||
| c.3716C>G | A1239G 2D AIThe SynGAP1 missense variant lies within a coiled‑coil domain. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and the Foldetta stability analysis is unavailable. ClinVar contains no entry for this variant, and it is not present in gnomAD. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -4.188 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.058 | Likely Benign | -2.09 | Neutral | 0.139 | Benign | 0.088 | Benign | 2.35 | Pathogenic | 0.00 | Affected | 0.1707 | 0.3142 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3716C>T | A1239V 2D AIThe A1239V missense change occurs in a coiled‑coil region of SynGAP1. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. ClinVar has no entry for this variant, and it is not present in gnomAD. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.541878 | Disordered | 0.534779 | Binding | 0.887 | 0.542 | 0.250 | -5.914 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.095 | Likely Benign | -2.28 | Neutral | 0.425 | Benign | 0.233 | Benign | 2.35 | Pathogenic | 0.00 | Affected | 0.0771 | 0.4518 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||||
| c.3718C>G | R1240G 2D AIThe SynGAP1 missense variant R1240G is not reported in ClinVar and has no entry in gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus agrees. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the preponderance of evidence indicates that R1240G is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.541878 | Disordered | 0.511333 | Binding | 0.865 | 0.540 | 0.375 | -9.763 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.280 | Likely Benign | -5.48 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.67 | Pathogenic | 0.00 | Affected | 0.3102 | 0.2895 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||
| c.3719G>A | R1240Q 2D AIThe SynGAP1 missense variant R1240Q is reported in gnomAD (variant ID 6-33446711‑G‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect are REVEL and AlphaMissense‑Optimized, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. ESM1b is uncertain. The high‑accuracy consensus (SGM‑Consensus) – a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN – is classified as Likely Pathogenic. AlphaMissense‑Optimized remains benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the preponderance of predictions (six pathogenic vs. two benign) indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.541878 | Disordered | 0.511333 | Binding | 0.865 | 0.540 | 0.375 | 6-33446711-G-A | 2 | 1.24e-6 | -7.110 | In-Between | 0.717 | Likely Pathogenic | Likely Benign | 0.304 | Likely Benign | -3.09 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.69 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2377 | 0.1992 | 1 | 1 | 1.0 | -28.06 | |||||||||||||||||||||||||||||||||
| c.3719G>C | R1240P 2D AIThe SynGAP1 missense variant R1240P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.541878 | Disordered | 0.511333 | Binding | 0.865 | 0.540 | 0.375 | -16.120 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.473 | Likely Benign | -5.45 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.66 | Pathogenic | 0.00 | Affected | 0.2037 | 0.3768 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||||||||
| c.3719G>T | R1240L 2D AIThe SynGAP1 missense variant R1240L is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. No Foldetta stability analysis is available for this variant. Based on the preponderance of pathogenic predictions, R1240L is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.541878 | Disordered | 0.511333 | Binding | 0.865 | 0.540 | 0.375 | -10.181 | Likely Pathogenic | 0.957 | Likely Pathogenic | Likely Pathogenic | 0.372 | Likely Benign | -5.48 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.67 | Pathogenic | 0.00 | Affected | 0.1513 | 0.3394 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3721C>A | L1241M 2D AISynGAP1 missense variant L1241M is listed in ClinVar with an Uncertain significance and is not reported in gnomAD. Functional prediction tools show a split verdict: benign calls come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic calls are made by polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is unresolved (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability predictor that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Consequently, the high‑accuracy tools do not converge on a single interpretation. Overall, the predictions are balanced between benign and pathogenic, leaving the variant’s effect uncertain, which aligns with its ClinVar designation of Uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.545602 | Disordered | 0.488880 | Uncertain | 0.828 | 0.541 | 0.375 | Uncertain | 1 | -5.881 | Likely Benign | 0.782 | Likely Pathogenic | Likely Benign | 0.167 | Likely Benign | -1.43 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.65 | Pathogenic | 0.00 | Affected | 0.0781 | 0.2280 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||
| c.3721C>G | L1241V 2D AIThe SynGAP1 missense variant L1241V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the variant as damaging. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. AlphaMissense‑Optimized yields an uncertain result, and Foldetta (the combined FoldX‑MD and Rosetta stability assessment) is not available for this variant. Overall, the preponderance of evidence from high‑accuracy predictors points to a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.545602 | Disordered | 0.488880 | Uncertain | 0.828 | 0.541 | 0.375 | -8.771 | Likely Pathogenic | 0.866 | Likely Pathogenic | Ambiguous | 0.153 | Likely Benign | -2.33 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.70 | Pathogenic | 0.00 | Affected | 0.1596 | 0.1883 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3722T>A | L1241Q 2D AIThe SynGAP1 missense variant L1241Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: the single benign prediction comes from REVEL, while all other evaluated algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic effect. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Based on the preponderance of pathogenic predictions and the high‑accuracy consensus, the variant is most likely pathogenic, and this conclusion is consistent with the absence of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.545602 | Disordered | 0.488880 | Uncertain | 0.828 | 0.541 | 0.375 | -10.429 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.386 | Likely Benign | -4.65 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.61 | Pathogenic | 0.00 | Affected | 0.1217 | 0.0488 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3722T>C | L1241P 2D AIThe SynGAP1 missense variant L1241P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.545602 | Disordered | 0.488880 | Uncertain | 0.828 | 0.541 | 0.375 | -16.301 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.471 | Likely Benign | -5.42 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.61 | Pathogenic | 0.00 | Affected | 0.3640 | 0.1048 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||
| c.3722T>G | L1241R 2D AIThe SynGAP1 missense variant L1241R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign outcome, whereas the remaining predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely pathogenic status. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools indicates that the variant is most likely pathogenic, and this conclusion is consistent with the absence of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.545602 | Disordered | 0.488880 | Uncertain | 0.828 | 0.541 | 0.375 | -14.178 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.366 | Likely Benign | -4.69 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.62 | Pathogenic | 0.00 | Affected | 0.1204 | 0.0488 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3724G>A | E1242K 2D AIThe SynGAP1 missense variant E1242K is catalogued in gnomAD (6‑33446716‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: the benign‑predicting REVEL score contrasts with a pathogenic consensus from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports “Likely Pathogenic,” and the protein‑folding stability method Foldetta is not available for this variant. Taken together, the majority of evidence points to a pathogenic impact, and this conclusion does not conflict with ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.557691 | Disordered | 0.456349 | Uncertain | 0.870 | 0.549 | 0.500 | 6-33446716-G-A | 1 | 6.20e-7 | -10.075 | Likely Pathogenic | 0.798 | Likely Pathogenic | Ambiguous | 0.179 | Likely Benign | -3.13 | Deleterious | 0.939 | Possibly Damaging | 0.670 | Possibly Damaging | 2.22 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1520 | 0.4024 | 1 | 0 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||
| c.3724G>C | E1242Q 2D AIThe SynGAP1 missense variant E1242Q is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta results are unavailable. Consequently, the aggregate evidence favors a benign effect for E1242Q, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.557691 | Disordered | 0.456349 | Uncertain | 0.870 | 0.549 | 0.500 | -7.036 | In-Between | 0.291 | Likely Benign | Likely Benign | 0.167 | Likely Benign | -2.33 | Neutral | 0.969 | Probably Damaging | 0.843 | Possibly Damaging | 2.21 | Pathogenic | 0.00 | Affected | 0.0763 | 0.3666 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3725A>C | E1242A 2D AIThe SynGAP1 missense variant E1242A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated tools (five pathogenic vs. three benign) predict a pathogenic impact. This prediction does not contradict ClinVar status, as the variant is not yet catalogued there. Thus, based on current computational evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.557691 | Disordered | 0.456349 | Uncertain | 0.870 | 0.549 | 0.500 | -5.654 | Likely Benign | 0.437 | Ambiguous | Likely Benign | 0.187 | Likely Benign | -4.66 | Deleterious | 0.939 | Possibly Damaging | 0.735 | Possibly Damaging | 2.21 | Pathogenic | 0.00 | Affected | 0.2738 | 0.4169 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.3725A>G | E1242G 2D AIThe SynGAP1 missense variant E1242G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and ESM1b, while those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Optimized also predicts a benign outcome, whereas AlphaMissense‑Default is uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple high‑accuracy predictors points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.557691 | Disordered | 0.456349 | Uncertain | 0.870 | 0.549 | 0.500 | -6.674 | Likely Benign | 0.528 | Ambiguous | Likely Benign | 0.214 | Likely Benign | -5.33 | Deleterious | 0.939 | Possibly Damaging | 0.670 | Possibly Damaging | 2.19 | Pathogenic | 0.00 | Affected | 0.2431 | 0.4295 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||||||
| c.3725A>T | E1242V 2D AIThe E1242V missense change occurs in the coiled‑coil domain of SynGAP1. ClinVar has no entry for this variant, and it is not reported in gnomAD. Prediction tools that agree on a benign effect are REVEL and AlphaMissense‑Optimized. Those that predict a pathogenic outcome include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (which is “Likely Pathogenic”). ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic; Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which simply lacks an entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.557691 | Disordered | 0.456349 | Uncertain | 0.870 | 0.549 | 0.500 | -7.456 | In-Between | 0.691 | Likely Pathogenic | Likely Benign | 0.267 | Likely Benign | -5.46 | Deleterious | 0.991 | Probably Damaging | 0.898 | Possibly Damaging | 2.17 | Pathogenic | 0.00 | Affected | 0.0447 | 0.4320 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3726G>C | E1242D 2D AIThe SynGAP1 missense variant E1242D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation—there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.557691 | Disordered | 0.456349 | Uncertain | 0.870 | 0.549 | 0.500 | -2.582 | Likely Benign | 0.158 | Likely Benign | Likely Benign | 0.031 | Likely Benign | -1.96 | Neutral | 0.020 | Benign | 0.018 | Benign | 2.37 | Pathogenic | 0.00 | Affected | 0.1594 | 0.2675 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3726G>T | E1242D 2D AIThe SynGAP1 missense variant E1242D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.557691 | Disordered | 0.456349 | Uncertain | 0.870 | 0.549 | 0.500 | -2.582 | Likely Benign | 0.158 | Likely Benign | Likely Benign | 0.032 | Likely Benign | -1.96 | Neutral | 0.020 | Benign | 0.018 | Benign | 2.37 | Pathogenic | 0.00 | Affected | 0.1594 | 0.2675 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3733G>A | E1245K 2D AIThe SynGAP1 missense variant E1245K is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that E1245K is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.712013 | Disordered | 0.387847 | Uncertain | 0.869 | 0.554 | 0.625 | -11.911 | Likely Pathogenic | 0.968 | Likely Pathogenic | Likely Pathogenic | 0.276 | Likely Benign | -3.22 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.28 | Pathogenic | 0.00 | Affected | 0.1627 | 0.6574 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3733G>C | E1245Q 2D AIThe SynGAP1 missense change E1245Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and PROVEAN, whereas pathogenic predictions are made by polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely pathogenic verdict (3 pathogenic vs. 1 benign). High‑accuracy assessments further support this: AlphaMissense‑Optimized is uncertain, SGM‑Consensus is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) is not available. Consequently, the majority of evidence points to a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.712013 | Disordered | 0.387847 | Uncertain | 0.869 | 0.554 | 0.625 | -11.323 | Likely Pathogenic | 0.804 | Likely Pathogenic | Ambiguous | 0.210 | Likely Benign | -2.29 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.0819 | 0.5952 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3734A>C | E1245A 2D AIThe SynGAP1 missense variant E1245A is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess the variant’s effect largely agree on a deleterious outcome: REVEL is the sole tool that predicts a benign effect, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus likewise indicates a likely pathogenic effect. Foldetta results are not available for this variant. Overall, the consensus of the available predictions indicates that E1245A is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.712013 | Disordered | 0.387847 | Uncertain | 0.869 | 0.554 | 0.625 | -11.433 | Likely Pathogenic | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.311 | Likely Benign | -4.85 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.25 | Pathogenic | 0.00 | Affected | 0.2555 | 0.5721 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3734A>G | E1245G 2D AIThe SynGAP1 missense variant E1245G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. The high‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is classified as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect; this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.712013 | Disordered | 0.387847 | Uncertain | 0.869 | 0.554 | 0.625 | -12.113 | Likely Pathogenic | 0.901 | Likely Pathogenic | Ambiguous | 0.299 | Likely Benign | -5.65 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.22 | Pathogenic | 0.00 | Affected | 0.2145 | 0.5447 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||
| c.3734A>T | E1245V 2D AIThe SynGAP1 missense change E1245V is not reported in ClinVar and is absent from gnomAD. Prediction tools that flag the variant as benign include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic effect. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus likewise reports a likely pathogenic outcome. Foldetta results are not available for this variant. Overall, the preponderance of computational evidence points to a pathogenic effect for E1245V, and this conclusion does not conflict with the current ClinVar status, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.712013 | Disordered | 0.387847 | Uncertain | 0.869 | 0.554 | 0.625 | -12.988 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.319 | Likely Benign | -5.65 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.21 | Pathogenic | 0.00 | Affected | 0.0518 | 0.6856 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3735G>C | E1245D 2D AIThe SynGAP1 missense variant E1245D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and no consensus could be drawn from the SGM Consensus (a tie between pathogenic and benign votes from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the balance of evidence from the available predictors leans toward a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation, as there is no reported ClinVar status to contradict the prediction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.712013 | Disordered | 0.387847 | Uncertain | 0.869 | 0.554 | 0.625 | -6.075 | Likely Benign | 0.928 | Likely Pathogenic | Ambiguous | 0.157 | Likely Benign | -2.46 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1585 | 0.3097 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3735G>T | E1245D 2D AIThe SynGAP1 missense variant E1245D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and no consensus could be drawn from the SGM Consensus (a tie between pathogenic and benign votes from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the balance of evidence from the available predictors leans toward a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.712013 | Disordered | 0.387847 | Uncertain | 0.869 | 0.554 | 0.625 | -6.075 | Likely Benign | 0.928 | Likely Pathogenic | Ambiguous | 0.157 | Likely Benign | -2.46 | Neutral | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1585 | 0.3097 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3739A>G | R1247G 2D AIThe SynGAP1 missense variant R1247G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are REVEL and AlphaMissense‑Optimized, whereas the majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default) predict a pathogenic impact; ESM1b is uncertain and SGM‑Consensus is labeled Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta results are unavailable. Overall, the balance of evidence favors a pathogenic classification, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | -7.920 | In-Between | 0.724 | Likely Pathogenic | Likely Benign | 0.195 | Likely Benign | -5.58 | Deleterious | 0.980 | Probably Damaging | 0.721 | Possibly Damaging | 1.70 | Pathogenic | 0.00 | Affected | 0.3047 | 0.2327 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||
| c.3739A>T | R1247W 2D AIThe SynGAP1 missense variant R1247W is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, whereas the majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely pathogenic outcome. No Foldetta stability result is available. Overall, the balance of evidence points to a pathogenic classification for R1247W, and this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | -9.694 | Likely Pathogenic | 0.676 | Likely Pathogenic | Likely Benign | 0.159 | Likely Benign | -6.38 | Deleterious | 1.000 | Probably Damaging | 0.982 | Probably Damaging | 1.68 | Pathogenic | 0.00 | Affected | 0.0963 | 0.2524 | 2 | -3 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.3740G>A | R1247K 2D  AIThe SynGAP1 missense variant R1247K is reported in gnomAD (6-33446732‑G‑A) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are SIFT and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact. There is no ClinVar classification to contradict this assessment. Thus, based on current predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | 6-33446732-G-A | 4 | 2.48e-6 | -4.713 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.082 | Likely Benign | -2.39 | Neutral | 0.122 | Benign | 0.064 | Benign | 1.80 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3428 | 0.2175 | 2 | 3 | 0.6 | -28.01 | |||||||||||||||||||||||||||||||||
| c.3740G>C | R1247T 2D  AIThe SynGAP1 missense variant R1247T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score, which is labeled Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic. No Foldetta stability result is available. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not conflict with ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | -6.302 | Likely Benign | 0.598 | Likely Pathogenic | Likely Benign | 0.206 | Likely Benign | -4.79 | Deleterious | 0.980 | Probably Damaging | 0.783 | Possibly Damaging | 1.71 | Pathogenic | 0.00 | Affected | 0.1543 | 0.2403 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||
| c.3740G>T | R1247M 2D  AIThe SynGAP1 missense variant R1247M is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts a benign effect, whereas the SGM‑Consensus remains pathogenic; Foldetta data are unavailable. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not conflict with the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | -6.347 | Likely Benign | 0.589 | Likely Pathogenic | Likely Benign | 0.239 | Likely Benign | -4.79 | Deleterious | 1.000 | Probably Damaging | 0.961 | Probably Damaging | 1.68 | Pathogenic | 0.00 | Affected | 0.1039 | 0.2122 | 0 | -1 | 6.4 | -24.99 | ||||||||||||||||||||||||||||||||||||||
| c.3741G>C | R1247S 2D  AIThe SynGAP1 missense variant R1247S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Pathogenic,” and Foldetta data are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for R1247S. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | -6.935 | Likely Benign | 0.851 | Likely Pathogenic | Ambiguous | 0.209 | Likely Benign | -4.79 | Deleterious | 0.961 | Probably Damaging | 0.721 | Possibly Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.2802 | 0.1977 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3741G>T | R1247S 2D AIThe SynGAP1 missense variant R1247S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Pathogenic,” and Foldetta data are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for R1247S. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | -6.935 | Likely Benign | 0.851 | Likely Pathogenic | Ambiguous | 0.209 | Likely Benign | -4.79 | Deleterious | 0.961 | Probably Damaging | 0.721 | Possibly Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.2802 | 0.1977 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3742C>A | L1248I 2D AIThe SynGAP1 missense variant L1248I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, and ESM1b, whereas pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are less decisive: AlphaMissense‑Optimized returns an uncertain result, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable, and Foldetta data are missing. Consequently, the overall evidence leans toward pathogenicity, but the lack of definitive high‑accuracy support and the absence of ClinVar annotation mean the prediction is not contradicted by existing clinical databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.834292 | Disordered | 0.371716 | Uncertain | 0.880 | 0.562 | 0.625 | -6.928 | Likely Benign | 0.872 | Likely Pathogenic | Ambiguous | 0.173 | Likely Benign | -1.56 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.75 | Pathogenic | 0.00 | Affected | 0.0919 | 0.2413 | 2 | 2 | 0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3742C>G | L1248V 2D AIThe SynGAP1 missense variant L1248V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated tools (five pathogenic vs three benign) predict a deleterious impact, and no high‑accuracy consensus or folding‑stability evidence contradicts this. Therefore, the variant is most likely pathogenic, and this assessment does not conflict with the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.834292 | Disordered | 0.371716 | Uncertain | 0.880 | 0.562 | 0.625 | -6.460 | Likely Benign | 0.899 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -2.33 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.73 | Pathogenic | 0.00 | Affected | 0.1510 | 0.1883 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3743T>A | L1248Q 2D AIThe SynGAP1 missense variant L1248Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) confirms this prediction; the Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a pathogenic effect for this variant, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.834292 | Disordered | 0.371716 | Uncertain | 0.880 | 0.562 | 0.625 | -6.471 | Likely Benign | 0.981 | Likely Pathogenic | Likely Pathogenic | 0.364 | Likely Benign | -4.65 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.64 | Pathogenic | 0.00 | Affected | 0.1067 | 0.0688 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3743T>C | L1248P 2D AIThe SynGAP1 missense variant L1248P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus agrees. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.834292 | Disordered | 0.371716 | Uncertain | 0.880 | 0.562 | 0.625 | -14.647 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.410 | Likely Benign | -5.45 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.64 | Pathogenic | 0.00 | Affected | 0.3596 | 0.0848 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||
| c.3743T>G | L1248R 2D AIThe SynGAP1 missense variant L1248R is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. No Foldetta stability analysis is available for this variant. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.834292 | Disordered | 0.371716 | Uncertain | 0.880 | 0.562 | 0.625 | -11.285 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 0.370 | Likely Benign | -4.69 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.64 | Pathogenic | 0.00 | Affected | 0.1222 | 0.0488 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3745A>G | R1249G 2D AIThe SynGAP1 missense variant R1249G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default) predict a pathogenic impact; ESM1b is uncertain and SGM‑Consensus is labeled Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta results are unavailable. Overall, the balance of evidence favors a pathogenic classification, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -7.405 | In-Between | 0.684 | Likely Pathogenic | Likely Benign | 0.206 | Likely Benign | -5.38 | Deleterious | 0.990 | Probably Damaging | 0.828 | Possibly Damaging | 1.70 | Pathogenic | 0.00 | Affected | 0.3087 | 0.2393 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||
| c.3745A>T | R1249W 2D AIThe SynGAP1 missense variant R1249W is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL and AlphaMissense‑Optimized, whereas pathogenic predictions are returned by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments further support a pathogenic interpretation: AlphaMissense‑Optimized predicts benign, but the SGM‑Consensus (majority vote) is pathogenic, and no Foldetta stability data are available. Overall, the majority of evidence points toward a pathogenic effect, which is consistent with the SGM‑Consensus designation and contradicts the benign predictions from a minority of tools. Thus, the variant is most likely pathogenic, and this conclusion aligns with the lack of ClinVar annotation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -9.841 | Likely Pathogenic | 0.477 | Ambiguous | Likely Benign | 0.209 | Likely Benign | -6.18 | Deleterious | 1.000 | Probably Damaging | 0.990 | Probably Damaging | 1.68 | Pathogenic | 0.00 | Affected | 0.1256 | 0.2179 | 2 | -3 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.3746G>A | R1249K 2D AIThe SynGAP1 missense variant R1249K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, SIFT and FATHMM predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; no Foldetta stability data are available, so it does not influence the overall assessment. Overall, the preponderance of evidence points to a benign effect for R1249K, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -5.782 | Likely Benign | 0.205 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -1.99 | Neutral | 0.219 | Benign | 0.191 | Benign | 1.80 | Pathogenic | 0.00 | Affected | 0.3343 | 0.2413 | 3 | 2 | 0.6 | -28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3746G>C | R1249T 2D AIThe SynGAP1 missense variant R1249T is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL and AlphaMissense‑Optimized, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score. The SGM‑Consensus, a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic effect. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM‑Consensus remains pathogenic; Foldetta results are unavailable. Overall, the preponderance of evidence (10 pathogenic vs. 2 benign predictions) points to a pathogenic impact for R1249T. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -8.014 | Likely Pathogenic | 0.712 | Likely Pathogenic | Likely Benign | 0.185 | Likely Benign | -4.59 | Deleterious | 0.990 | Probably Damaging | 0.903 | Possibly Damaging | 1.71 | Pathogenic | 0.00 | Affected | 0.1646 | 0.2458 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||
| c.3746G>T | R1249M 2D AIThe SynGAP1 missense variant R1249M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (which is “Likely Pathogenic” based on a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Benign predictions are limited to REVEL and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains pathogenic. No Foldetta stability analysis is available for this variant. Overall, the preponderance of evidence from multiple in‑silico tools indicates that R1249M is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -8.520 | Likely Pathogenic | 0.643 | Likely Pathogenic | Likely Benign | 0.232 | Likely Benign | -4.59 | Deleterious | 1.000 | Probably Damaging | 0.979 | Probably Damaging | 1.68 | Pathogenic | 0.00 | Affected | 0.1318 | 0.2173 | 0 | -1 | 6.4 | -24.99 | ||||||||||||||||||||||||||||||||||||||
| c.3747G>C | R1249S 2D AIThe SynGAP1 missense variant R1249S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN and labels the variant as Likely Pathogenic. High‑accuracy assessments are limited: AlphaMissense‑Optimized returns an Uncertain result, and the Foldetta stability analysis is not available for this residue. Overall, the majority of evidence points toward a pathogenic effect, and this assessment does not conflict with any ClinVar annotation because no ClinVar entry exists for R1249S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -6.936 | Likely Benign | 0.828 | Likely Pathogenic | Ambiguous | 0.196 | Likely Benign | -4.52 | Deleterious | 0.980 | Probably Damaging | 0.828 | Possibly Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.2777 | 0.1843 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3747G>T | R1249S 2D AIThe SynGAP1 missense variant R1249S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN and labels the variant as Likely Pathogenic. High‑accuracy assessments are limited: AlphaMissense‑Optimized returns an Uncertain result, and the Foldetta stability analysis is not available for this residue. Overall, the majority of evidence points toward a pathogenic effect, and this assessment does not conflict with any ClinVar annotation because no ClinVar entry exists for R1249S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -6.936 | Likely Benign | 0.828 | Likely Pathogenic | Ambiguous | 0.196 | Likely Benign | -4.52 | Deleterious | 0.980 | Probably Damaging | 0.828 | Possibly Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.2777 | 0.1843 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3751C>A | Q1251K 2D AIThe SynGAP1 missense variant Q1251K is catalogued in gnomAD (ID 6‑33446743‑C‑A) but has no ClinVar entry. Functional prediction tools show a split: benign calls from REVEL and FATHMM, whereas the majority—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—label it pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports it as Likely Pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized rates the variant as Uncertain, SGM‑Consensus remains Likely Pathogenic, and Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a pathogenic effect, and this assessment does not conflict with ClinVar, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | 6-33446743-C-A | 1 | 6.20e-7 | -11.113 | Likely Pathogenic | 0.850 | Likely Pathogenic | Ambiguous | 0.208 | Likely Benign | -2.92 | Deleterious | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 2.53 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1418 | 0.2635 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||
| c.3751C>G | Q1251E 2D AIThe SynGAP1 missense variant Q1251E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. Two tools, ESM1b and AlphaMissense‑Default, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it contains two uncertain and two benign calls, and Foldetta data are unavailable. Overall, the balance of evidence leans toward a benign interpretation, and this does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | -7.836 | In-Between | 0.499 | Ambiguous | Likely Benign | 0.183 | Likely Benign | -2.26 | Neutral | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.1281 | 0.1266 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3752A>C | Q1251P 2D AIThe SynGAP1 missense variant Q1251P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | -14.584 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.344 | Likely Benign | -4.45 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 2.43 | Pathogenic | 0.00 | Affected | 0.2418 | 0.4408 | 0 | -1 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||||||
| c.3752A>G | Q1251R 2D AIThe SynGAP1 missense variant Q1251R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of other in silico predictors (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) indicate a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic; this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | -9.456 | Likely Pathogenic | 0.890 | Likely Pathogenic | Ambiguous | 0.236 | Likely Benign | -2.92 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.1257 | 0.1224 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3752A>T | Q1251L 2D AIThe SynGAP1 missense variant Q1251L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b) predict a pathogenic impact; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized remains benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—favors pathogenicity. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently contains no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | -10.298 | Likely Pathogenic | 0.412 | Ambiguous | Likely Benign | 0.279 | Likely Benign | -4.71 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.58 | Benign | 0.00 | Affected | 0.0626 | 0.4605 | -2 | -2 | 7.3 | -14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3753G>C | Q1251H 2D AIThe SynGAP1 missense variant Q1251H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a predominance of pathogenic calls: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all predict deleterious effects, while REVEL predicts a benign outcome. Two tools return uncertain results: ESM1b and AlphaMissense‑Optimized. High‑accuracy assessments further support a harmful interpretation: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic, and AlphaMissense‑Optimized remains inconclusive. No Foldetta stability data are available. Overall, the balance of evidence points to a pathogenic effect for Q1251H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | -7.561 | In-Between | 0.937 | Likely Pathogenic | Ambiguous | 0.176 | Likely Benign | -3.71 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 2.44 | Pathogenic | 0.00 | Affected | 0.1132 | 0.2399 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3753G>T | Q1251H 2D AIThe SynGAP1 missense variant Q1251H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a predominance of pathogenic calls: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all predict deleterious effects, while REVEL predicts a benign outcome. Two tools return uncertain results: ESM1b and AlphaMissense‑Optimized. High‑accuracy assessments further support a harmful interpretation: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic, and AlphaMissense‑Optimized remains inconclusive. No Foldetta stability data are available. Overall, the balance of evidence points to a pathogenic effect for Q1251H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.771762 | Disordered | 0.363872 | Uncertain | 0.869 | 0.551 | 0.875 | -7.561 | In-Between | 0.937 | Likely Pathogenic | Ambiguous | 0.176 | Likely Benign | -3.71 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 2.44 | Pathogenic | 0.00 | Affected | 0.1132 | 0.2399 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3754C>A | Q1252K 2D AIThe SynGAP1 missense variant Q1252K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—predict a pathogenic impact, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the predominance of pathogenic predictions, the variant is most likely pathogenic; this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -13.590 | Likely Pathogenic | 0.878 | Likely Pathogenic | Ambiguous | 0.217 | Likely Benign | -3.22 | Deleterious | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 2.03 | Pathogenic | 0.00 | Affected | 0.1460 | 0.2518 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||||||||||||
| c.3754C>G | Q1252E 2D AIThe SynGAP1 missense variant Q1252E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. three benign) lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -10.181 | Likely Pathogenic | 0.485 | Ambiguous | Likely Benign | 0.152 | Likely Benign | -2.32 | Neutral | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 2.01 | Pathogenic | 0.00 | Affected | 0.1130 | 0.1266 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3755A>C | Q1252P 2D AIThe SynGAP1 missense variant Q1252P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -14.386 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.352 | Likely Benign | -4.75 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 1.95 | Pathogenic | 0.00 | Affected | 0.1998 | 0.3742 | 0 | -1 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||||||
| c.3755A>G | Q1252R 2D AIThe SynGAP1 missense variant Q1252R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: REVEL scores the variant as benign, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all predict pathogenicity. Grouping by consensus, the majority of tools (seven) support a pathogenic effect, while only one tool (REVEL) indicates benign. High‑accuracy assessments further support a deleterious impact: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic; AlphaMissense‑Optimized remains uncertain, and Foldetta data are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus result, the variant is most likely pathogenic. This conclusion aligns with the lack of ClinVar annotation and gnomAD absence, and there is no contradiction with ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -11.890 | Likely Pathogenic | 0.913 | Likely Pathogenic | Ambiguous | 0.249 | Likely Benign | -3.26 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.00 | Pathogenic | 0.00 | Affected | 0.1284 | 0.0854 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||||
| c.3755A>T | Q1252L 2D AIThe SynGAP1 missense variant Q1252L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus—predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -8.110 | Likely Pathogenic | 0.833 | Likely Pathogenic | Ambiguous | 0.295 | Likely Benign | -5.62 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.97 | Pathogenic | 0.00 | Affected | 0.0593 | 0.3689 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3756G>C | Q1252H 2D AIThe SynGAP1 missense variant Q1252H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all classify the variant as deleterious. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -6.891 | Likely Benign | 0.951 | Likely Pathogenic | Ambiguous | 0.175 | Likely Benign | -4.02 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 1.95 | Pathogenic | 0.00 | Affected | 0.1038 | 0.2149 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3756G>T | Q1252H 2D AIThe SynGAP1 missense variant Q1252H is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all classify the variant as damaging. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain, SGM‑Consensus is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is unavailable for this variant. Overall, the preponderance of evidence from multiple in‑silico predictors indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.759478 | Disordered | 0.371411 | Uncertain | 0.850 | 0.544 | 0.875 | -6.891 | Likely Benign | 0.951 | Likely Pathogenic | Ambiguous | 0.175 | Likely Benign | -4.02 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 1.95 | Pathogenic | 0.00 | Affected | 0.1038 | 0.2149 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3761A>T | E1254V 2D AIThe SynGAP1 missense variant E1254V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: REVEL scores the variant as benign, whereas the remaining seven tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) predict it to be pathogenic. Grouping by consensus, the benign prediction is represented only by REVEL, while the pathogenic predictions are supported by the majority of in silico methods. High‑accuracy assessments further reinforce a pathogenic interpretation: AlphaMissense‑Optimized is uncertain, but the SGM‑Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—classifies the variant as likely pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic, which is consistent with the lack of ClinVar annotation and gnomAD absence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.657645 | Disordered | 0.403242 | Uncertain | 0.886 | 0.555 | 0.625 | -9.913 | Likely Pathogenic | 0.814 | Likely Pathogenic | Ambiguous | 0.350 | Likely Benign | -5.15 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.31 | Pathogenic | 0.00 | Affected | 0.0481 | 0.5894 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3763A>C | K1255Q 2D AIThe SynGAP1 missense variant K1255Q is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate pathogenicity. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus also reports a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for K1255Q, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -12.680 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.282 | Likely Benign | -3.19 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.87 | Pathogenic | 0.00 | Affected | 0.3625 | 0.1102 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||||||||||
| c.3763A>G | K1255E 2D AIThe SynGAP1 missense variant K1255E is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are not available. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -15.072 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.308 | Likely Benign | -3.12 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.88 | Pathogenic | 0.00 | Affected | 0.3094 | 0.0877 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3764A>C | K1255T 2D AIThe SynGAP1 missense variant K1255T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as pathogenic; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -9.745 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.360 | Likely Benign | -4.79 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.85 | Pathogenic | 0.00 | Affected | 0.1755 | 0.2628 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||||||||
| c.3764A>G | K1255R 2D AIThe SynGAP1 K1255R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a majority of tools predict a pathogenic impact: polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the change as damaging. The high‑accuracy consensus approach (SGM‑Consensus) – a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN – yields a pathogenic verdict (3 pathogenic vs. 1 benign). AlphaMissense‑Optimized is uncertain, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for K1255R. This prediction is consistent with the lack of ClinVar annotation; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -9.687 | Likely Pathogenic | 0.866 | Likely Pathogenic | Ambiguous | 0.171 | Likely Benign | -2.43 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.93 | Pathogenic | 0.00 | Affected | 0.3727 | 0.0878 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||
| c.3764A>T | K1255M 2D AIThe SynGAP1 missense variant K1255M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign change, whereas all other evaluated algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict pathogenicity. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus confirms a likely pathogenic status. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence from multiple independent predictors points to a pathogenic effect, and this conclusion is consistent with the absence of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -10.554 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.393 | Likely Benign | -4.82 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.82 | Pathogenic | 0.00 | Affected | 0.0776 | 0.3154 | 0 | -1 | 5.8 | 3.02 | ||||||||||||||||||||||||||||||||||||||
| c.3765G>C | K1255N 2D AIThe SynGAP1 missense variant K1255N is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -12.586 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.210 | Likely Benign | -3.99 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.85 | Pathogenic | 0.00 | Affected | 0.2993 | 0.1302 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3765G>T | K1255N 2D AIThe SynGAP1 missense variant K1255N is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the absence of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.637480 | Disordered | 0.417615 | Uncertain | 0.880 | 0.563 | 0.625 | -12.586 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.210 | Likely Benign | -3.99 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.85 | Pathogenic | 0.00 | Affected | 0.2993 | 0.1302 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3766G>A | D1256N 2D AIThe SynGAP1 D1256N missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the variant as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic; this assessment does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -10.375 | Likely Pathogenic | 0.897 | Likely Pathogenic | Ambiguous | 0.290 | Likely Benign | -3.85 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.67 | Pathogenic | 0.00 | Affected | 0.0832 | 0.4219 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3766G>C | D1256H 2D AIThe SynGAP1 missense variant D1256H is predicted to be pathogenic by every available in‑silico tool. Benign predictions are absent; all listed predictors—REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as damaging. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports a pathogenic effect, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. Foldetta results are not available. ClinVar contains no entry for this variant, and it is absent from gnomAD, so there is no existing clinical annotation to contradict the computational predictions. Overall, the evidence strongly suggests the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -14.272 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.508 | Likely Pathogenic | -5.29 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.63 | Pathogenic | 0.00 | Affected | 0.0963 | 0.4798 | 1 | -1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||||||||
| c.3766G>T | D1256Y 2D AIThe SynGAP1 missense variant D1256Y is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools uniformly classify the variant as pathogenic: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a deleterious effect. No tool in the dataset predicts a benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of available predictions indicates that D1256Y is most likely pathogenic, and this conclusion is not contradicted by ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -15.855 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.507 | Likely Pathogenic | -6.92 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.62 | Pathogenic | 0.00 | Affected | 0.0446 | 0.4275 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||
| c.3767A>C | D1256A 2D AIThe SynGAP1 D1256A missense change is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as “Likely Pathogenic.” High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as likely pathogenic; Foldetta results are not available. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -11.665 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.443 | Likely Benign | -6.06 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.66 | Pathogenic | 0.00 | Affected | 0.2830 | 0.4451 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||||||||||
| c.3767A>G | D1256G 2D AIThe SynGAP1 missense variant D1256G is not reported in ClinVar and has no entry in gnomAD. Prediction tools that assess the variant’s effect fall into two groups: the benign‑predicting REVEL score, and a consensus of pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates likely pathogenicity. Foldetta results are unavailable. Overall, the preponderance of evidence from both general and high‑accuracy tools points to a pathogenic effect for D1256G. This conclusion is consistent with the absence of ClinVar annotation, so there is no contradiction with existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -11.341 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.457 | Likely Benign | -5.45 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.65 | Pathogenic | 0.00 | Affected | 0.2886 | 0.5040 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3767A>T | D1256V 2D AIThe SynGAP1 missense variant D1256V is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess functional impact largely agree on a deleterious effect: SIFT, polyPhen‑2 (HumDiv and HumVar), PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while the only benign prediction comes from REVEL. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as “Likely Pathogenic.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status. Foldetta results are not available, so they do not influence the overall assessment. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -14.067 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.446 | Likely Benign | -6.89 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.63 | Pathogenic | 0.00 | Affected | 0.0573 | 0.4407 | -2 | -3 | 7.7 | -15.96 | ||||||||||||||||||||||||||||||||||||||
| c.3768T>A | D1256E 2D AIThe SynGAP1 D1256E missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all indicate pathogenicity. The SGM‑Consensus, which is a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta results are unavailable. Taken together, the consensus of most evidence points to a pathogenic effect, and this conclusion does not contradict any existing ClinVar annotation (none is present). Thus, the variant is most likely pathogenic based on current predictive tools. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -5.806 | Likely Benign | 0.842 | Likely Pathogenic | Ambiguous | 0.149 | Likely Benign | -2.71 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.1022 | 0.4172 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3768T>G | D1256E 2D AIThe SynGAP1 D1256E missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools cluster into two groups: benign predictions come from REVEL and ESM1b, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy consensus, SGM‑Consensus, is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN and therefore reports the variant as likely pathogenic. AlphaMissense‑Optimized is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Taken together, the majority of evidence points to a pathogenic effect, and this assessment does not contradict any ClinVar annotation because none exists. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.549308 | Disordered | 0.445789 | Uncertain | 0.876 | 0.571 | 0.625 | -5.806 | Likely Benign | 0.842 | Likely Pathogenic | Ambiguous | 0.149 | Likely Benign | -2.71 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.72 | Pathogenic | 0.00 | Affected | 0.1022 | 0.4172 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.376T>A | F126I 2D  AIThe SynGAP1 missense variant F126I is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools largely agree that the change is benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify it as benign. Only two tools predict pathogenicity—SIFT and AlphaMissense‑Default. High‑accuracy consensus methods reinforce the benign assessment: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign,” and AlphaMissense‑Optimized itself predicts benign. The protein‑folding stability predictor Foldetta was not available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -2.919 | Likely Benign | 0.735 | Likely Pathogenic | Likely Benign | 0.044 | Likely Benign | -2.20 | Neutral | 0.160 | Benign | 0.045 | Benign | 3.95 | Benign | 0.00 | Affected | 0.2569 | 0.2418 | 1 | 0 | 1.7 | -34.02 | |||||||||||||||||||||||||||||||||||||||
| c.376T>C | F126L 2D  AIThe SynGAP1 missense variant F126L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The high‑accuracy consensus (SGM‑Consensus) classifies the variant as Likely Benign, while the high‑accuracy AlphaMissense‑Optimized result is inconclusive and Foldetta data are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict the ClinVar status, which contains no pathogenic claim. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -0.992 | Likely Benign | 0.948 | Likely Pathogenic | Ambiguous | 0.091 | Likely Benign | -2.27 | Neutral | 0.001 | Benign | 0.001 | Benign | 4.01 | Benign | 0.00 | Affected | 0.2780 | 0.3399 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||||||
| c.376T>G | F126V 2D  AIThe SynGAP1 missense variant F126V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for F126V, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -1.584 | Likely Benign | 0.635 | Likely Pathogenic | Likely Benign | 0.086 | Likely Benign | -2.47 | Neutral | 0.160 | Benign | 0.045 | Benign | 3.98 | Benign | 0.00 | Affected | 0.2496 | 0.2646 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3772C>A | Q1258K 2D AIThe SynGAP1 missense variant Q1258K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: REVEL scores the variant as benign, whereas the majority of other in silico predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—classify it as pathogenic. Grouping by consensus, the benign prediction is represented only by REVEL, while the pathogenic predictions are supported by seven distinct algorithms. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized returns an uncertain result, SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labels the variant as likely pathogenic, and Foldetta data are not available. Taken together, the preponderance of evidence from multiple pathogenic predictors and the SGM‑Consensus suggests that the variant is most likely pathogenic, which is consistent with the absence of a ClinVar entry and gnomAD observation. Thus, the variant is most likely pathogenic, and this prediction does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | -10.927 | Likely Pathogenic | 0.912 | Likely Pathogenic | Ambiguous | 0.227 | Likely Benign | -3.19 | Deleterious | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 2.03 | Pathogenic | 0.00 | Affected | 0.1151 | 0.2761 | 1 | 1 | -0.4 | 0.04 | ||||||||||||||||||||||||||||||||||||||
| c.3772C>G | Q1258E 2D AIThe SynGAP1 missense variant Q1258E is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, PROVEAN, and AlphaMissense‑Optimized, whereas pathogenic predictions are reported by polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely pathogenic verdict (3/4 pathogenic votes). High‑accuracy assessments further support this: AlphaMissense‑Optimized indicates a benign effect, whereas the SGM‑Consensus remains pathogenic; Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points toward a pathogenic impact, which is consistent with the lack of ClinVar annotation and gnomAD absence. Thus, the variant is most likely pathogenic, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | -9.894 | Likely Pathogenic | 0.666 | Likely Pathogenic | Likely Benign | 0.206 | Likely Benign | -2.39 | Neutral | 0.985 | Probably Damaging | 0.981 | Probably Damaging | 2.01 | Pathogenic | 0.00 | Affected | 0.0958 | 0.1897 | 2 | 2 | 0.0 | 0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3773A>C | Q1258P 2D AIThe SynGAP1 missense variant Q1258P is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | -16.904 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.440 | Likely Benign | -4.79 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 1.95 | Pathogenic | 0.00 | Affected | 0.1727 | 0.4187 | 0 | -1 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||||||
| c.3773A>G | Q1258R 2D AIThe SynGAP1 missense variant Q1258R is listed in ClinVar with an uncertain significance (ClinVar ID 3359527.0) and is not observed in gnomAD. Functional prediction tools largely agree on a deleterious effect: pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default, while only REVEL predicts a benign outcome. The high‑accuracy predictors give the following results: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic classification; Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, has no available output for this variant. Based on the preponderance of pathogenic predictions and the SGM Consensus, the variant is most likely pathogenic, which is consistent with its ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | Uncertain | 1 | -10.971 | Likely Pathogenic | 0.931 | Likely Pathogenic | Ambiguous | 0.316 | Likely Benign | -3.19 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 2.00 | Pathogenic | 0.00 | Affected | 0.1027 | 0.0991 | 1 | 1 | -1.0 | 28.06 | ||||||||||||||||||||||||||||||||||||
| c.3773A>T | Q1258L 2D AIThe SynGAP1 missense variant Q1258L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: REVEL scores the variant as benign, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all predict pathogenicity. Grouping by consensus, the majority of tools (seven) predict pathogenic, while only one tool (REVEL) predicts benign. High‑accuracy assessments further support a deleterious effect: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic; AlphaMissense‑Optimized remains uncertain, and Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic. This conclusion aligns with the lack of ClinVar annotation and gnomAD absence, indicating no conflicting evidence from population databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | -10.302 | Likely Pathogenic | 0.895 | Likely Pathogenic | Ambiguous | 0.341 | Likely Benign | -5.55 | Deleterious | 0.994 | Probably Damaging | 0.988 | Probably Damaging | 1.97 | Pathogenic | 0.00 | Affected | 0.0448 | 0.4137 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3774G>C | Q1258H 2D AIThe SynGAP1 missense variant Q1258H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of other in silico predictors (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also leans pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic; this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | -5.465 | Likely Benign | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.172 | Likely Benign | -3.99 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 1.95 | Pathogenic | 0.00 | Affected | 0.0735 | 0.3066 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3774G>T | Q1258H 2D AIThe SynGAP1 missense variant Q1258H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) is likely pathogenic; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.486429 | Structured | 0.525814 | Binding | 0.859 | 0.577 | 0.250 | -5.465 | Likely Benign | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.172 | Likely Benign | -3.99 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 1.95 | Pathogenic | 0.00 | Affected | 0.0735 | 0.3066 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3776T>A | I1259N 2D AIThe SynGAP1 missense variant I1259N is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) all indicate likely pathogenicity. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus also reports a likely pathogenic classification. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from multiple in silico predictors points to a pathogenic effect for I1259N. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.494003 | Structured | 0.576405 | Binding | 0.885 | 0.574 | 0.250 | -10.979 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.459 | Likely Benign | -3.55 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.51 | Benign | 0.00 | Affected | 0.0799 | 0.0340 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3778A>C | K1260Q 2D AIThe SynGAP1 missense variant K1260Q is listed in ClinVar with no submitted interpretation and is present in gnomAD (ID 6‑33446770‑A‑C). Functional prediction tools cluster into two groups: benign predictions come from REVEL, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. ESM1b is uncertain. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts benign, but the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic verdict, and Foldetta results are unavailable. Overall, the majority of evidence points to pathogenicity, and this conclusion does not contradict the ClinVar status, which remains unclassified. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | 6-33446770-A-C | -7.830 | In-Between | 0.325 | Likely Benign | Likely Benign | 0.367 | Likely Benign | -3.06 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.36 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3597 | 0.1102 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||||||||
| c.3778A>G | K1260E 2D AIThe SynGAP1 missense variant K1260E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—predict a pathogenic impact, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus remains pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions point to a pathogenic effect, and there is no ClinVar annotation to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | -10.913 | Likely Pathogenic | 0.846 | Likely Pathogenic | Ambiguous | 0.462 | Likely Benign | -3.06 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.38 | Pathogenic | 0.00 | Affected | 0.3129 | 0.0877 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3779A>C | K1260T 2D AIThe SynGAP1 missense variant K1260T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (which is “Likely Pathogenic” based on a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Benign predictions are limited to REVEL and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains pathogenic. No Foldetta stability analysis is available for this variant. Overall, the preponderance of evidence from multiple in‑silico tools indicates that K1260T is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | -10.099 | Likely Pathogenic | 0.772 | Likely Pathogenic | Likely Benign | 0.387 | Likely Benign | -4.59 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.33 | Pathogenic | 0.00 | Affected | 0.1859 | 0.3028 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||||||||
| c.3779A>G | K1260R 2D AIThe SynGAP1 missense variant K1260R is catalogued in gnomAD (ID 6‑33446771‑A‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; those that predict pathogenicity are polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the collective predictions point to a benign impact, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | 6-33446771-A-G | 1 | 6.20e-7 | -6.033 | Likely Benign | 0.135 | Likely Benign | Likely Benign | 0.254 | Likely Benign | -2.29 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.40 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3655 | 0.0878 | 2 | 3 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||
| c.3779A>T | K1260M 2D AIThe SynGAP1 missense variant K1260M is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the majority of algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—classify the substitution as pathogenic. High‑accuracy assessments further support a deleterious impact: the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports “Likely Pathogenic”; AlphaMissense‑Optimized yields an uncertain result, and Foldetta’s stability prediction is unavailable. Taken together, the evidence overwhelmingly points to a pathogenic effect for K1260M. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | -10.938 | Likely Pathogenic | 0.887 | Likely Pathogenic | Ambiguous | 0.400 | Likely Benign | -4.72 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.29 | Pathogenic | 0.00 | Affected | 0.0746 | 0.3366 | 0 | -1 | 5.8 | 3.02 | ||||||||||||||||||||||||||||||||||||||
| c.377T>A | F126Y 2D  AIThe SynGAP1 missense variant F126Y has no ClinVar entry and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized independently predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -3.519 | Likely Benign | 0.509 | Ambiguous | Likely Benign | 0.044 | Likely Benign | -1.29 | Neutral | 0.851 | Possibly Damaging | 0.221 | Benign | 3.90 | Benign | 0.00 | Affected | 0.1631 | 0.1907 | 7 | 3 | -4.1 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.377T>C | F126S 2D  AIThe SynGAP1 missense variant F126S is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools largely agree that the change is benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM all classify it as benign. In contrast, SIFT and AlphaMissense‑Default predict a pathogenic effect. AlphaMissense‑Optimized returns an uncertain result, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available prediction for this residue. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” Overall, the majority of high‑accuracy tools (REVEL, PROVEAN, polyPhen‑2, ESM1b, FATHMM, and SGM‑Consensus) support a benign interpretation, while only two tools (SIFT, AlphaMissense‑Default) suggest pathogenicity. Given the preponderance of benign predictions and the absence of ClinVar evidence, the variant is most likely benign and does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -0.234 | Likely Benign | 0.844 | Likely Pathogenic | Ambiguous | 0.086 | Likely Benign | -2.46 | Neutral | 0.160 | Benign | 0.045 | Benign | 3.96 | Benign | 0.00 | Affected | 0.4833 | 0.0000 | -3 | -2 | -3.6 | -60.10 | |||||||||||||||||||||||||||||||||||||||
| c.377T>G | F126C 2D  AIThe SynGAP1 missense variant F126C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, more tools predict pathogenicity (five) than benignity (three), and no ClinVar evidence contradicts this assessment. Thus, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -2.553 | Likely Benign | 0.824 | Likely Pathogenic | Ambiguous | 0.109 | Likely Benign | -3.19 | Deleterious | 0.952 | Possibly Damaging | 0.570 | Possibly Damaging | 3.88 | Benign | 0.00 | Affected | 0.2682 | 0.1270 | -4 | -2 | -0.3 | -44.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3780G>C | K1260N 2D AIThe SynGAP1 missense variant K1260N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) all classify the variant as pathogenic or likely pathogenic. AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. Based on the consensus of high‑accuracy predictors, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | -9.053 | Likely Pathogenic | 0.915 | Likely Pathogenic | Ambiguous | 0.157 | Likely Benign | -3.39 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.42 | Pathogenic | 0.00 | Affected | 0.3064 | 0.1302 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3780G>T | K1260N 2D AIThe SynGAP1 missense variant K1260N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) all classify the variant as pathogenic or likely pathogenic. AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. Based on the consensus of high‑accuracy predictors, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.509769 | Disordered | 0.625808 | Binding | 0.890 | 0.575 | 0.250 | -9.053 | Likely Pathogenic | 0.915 | Likely Pathogenic | Ambiguous | 0.157 | Likely Benign | -3.39 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.42 | Pathogenic | 0.00 | Affected | 0.3064 | 0.1302 | 1 | 0 | 0.4 | -14.07 | ||||||||||||||||||||||||||||||||||||||
| c.3784A>C | I1262L 2D  AIThe SynGAP1 missense variant I1262L is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and PROVEAN, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b remains uncertain. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (pathogenic), and PROVEAN (benign)—also indicates pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic impact for I1262L, and this conclusion does not contradict the current ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -7.334 | In-Between | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.269 | Likely Benign | -1.66 | Neutral | 0.981 | Probably Damaging | 0.970 | Probably Damaging | 1.94 | Pathogenic | 0.00 | Affected | 0.0735 | 0.3246 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3784A>G | I1262V 2D 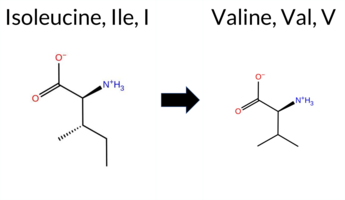 AIThe SynGAP1 I1262V missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that agree on a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments are less decisive: AlphaMissense‑Optimized rates the variant as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta folding‑stability data are unavailable. Overall, the majority of standard predictors lean toward pathogenicity, but the most reliable tools provide no clear verdict. Thus, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -6.773 | Likely Benign | 0.814 | Likely Pathogenic | Ambiguous | 0.237 | Likely Benign | -0.82 | Neutral | 0.958 | Probably Damaging | 0.970 | Probably Damaging | 1.99 | Pathogenic | 0.00 | Affected | 0.1049 | 0.3270 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3784A>T | I1262F 2D 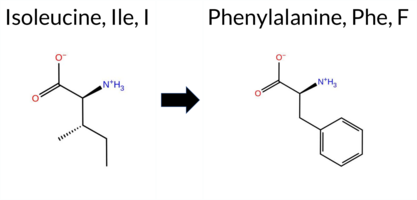 AIThe SynGAP1 missense variant I1262F is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess functional impact uniformly indicate a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic. No tool in the dataset predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the evidence strongly supports that the variant is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -11.343 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.515 | Likely Pathogenic | -3.32 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.81 | Pathogenic | 0.00 | Affected | 0.0532 | 0.2731 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3785T>A | I1262N 2D AIThe SynGAP1 missense variant I1262N is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign outcome, whereas all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely pathogenic status. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant. Overall, the preponderance of computational evidence indicates that I1262N is most likely pathogenic, and this conclusion is consistent with the absence of any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -14.512 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.463 | Likely Benign | -5.81 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.79 | Pathogenic | 0.00 | Affected | 0.0940 | 0.0340 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3785T>C | I1262T 2D 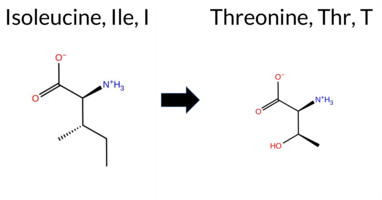 AIThe SynGAP1 missense variant I1262T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic. No tool in the dataset predicts a benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely pathogenic status. The Foldetta stability analysis is not available for this variant. Overall, the consensus of all available predictions points to a pathogenic effect, and this conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -11.985 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.514 | Likely Pathogenic | -4.14 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.81 | Pathogenic | 0.00 | Affected | 0.1109 | 0.1419 | 0 | -1 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||||||
| c.3785T>G | I1262S 2D AIThe SynGAP1 missense variant I1262S is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion is not contradicted by ClinVar status, which currently has no entry for I1262S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -15.167 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.483 | Likely Benign | -4.98 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.2943 | 0.1110 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3786C>G | I1262M 2D 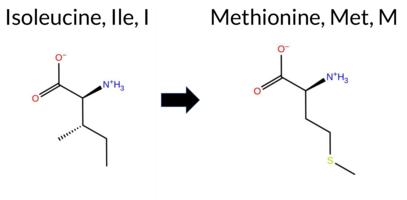 AIThe SynGAP1 missense variant I1262M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and PROVEAN, while pathogenic predictions are made by polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic effect. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus also leans pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -9.081 | Likely Pathogenic | 0.965 | Likely Pathogenic | Likely Pathogenic | 0.248 | Likely Benign | -2.49 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.81 | Pathogenic | 0.00 | Affected | 0.0671 | 0.2726 | 2 | 1 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3787A>C | I1263L 2D AIThe SynGAP1 missense variant I1263L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign, two pathogenic). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -1.210 | Likely Benign | 0.699 | Likely Pathogenic | Likely Benign | 0.194 | Likely Benign | -1.66 | Neutral | 0.011 | Benign | 0.022 | Benign | 1.94 | Pathogenic | 0.00 | Affected | 0.0717 | 0.2890 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3787A>G | I1263V 2D AIThe SynGAP1 missense variant I1263V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two benign vs. two pathogenic votes), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -4.230 | Likely Benign | 0.729 | Likely Pathogenic | Likely Benign | 0.221 | Likely Benign | -0.83 | Neutral | 0.437 | Benign | 0.170 | Benign | 1.99 | Pathogenic | 0.00 | Affected | 0.1136 | 0.3102 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3787A>T | I1263F 2D AIThe SynGAP1 missense variant I1263F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta results are unavailable. Because the majority of evidence points to a deleterious effect and there is no ClinVar annotation to contradict this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -5.887 | Likely Benign | 0.952 | Likely Pathogenic | Ambiguous | 0.335 | Likely Benign | -3.32 | Deleterious | 0.968 | Probably Damaging | 0.637 | Possibly Damaging | 1.81 | Pathogenic | 0.00 | Affected | 0.0494 | 0.2375 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3788T>A | I1263N 2D AIThe SynGAP1 missense variant I1263N is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -9.158 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.379 | Likely Benign | -5.80 | Deleterious | 0.995 | Probably Damaging | 0.913 | Probably Damaging | 1.79 | Pathogenic | 0.00 | Affected | 0.1041 | 0.0340 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3788T>C | I1263T 2D AIThe SynGAP1 missense variant I1263T is listed in ClinVar with an “Uncertain” status and is present in the gnomAD database (ID 6‑33446780‑T‑C). Functional prediction tools largely agree on a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report pathogenicity, while only ESM1b predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a pathogenic effect, which aligns with the ClinVar designation of uncertainty but does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | Uncertain | 1 | 6-33446780-T-C | 2 | 1.24e-6 | -6.564 | Likely Benign | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.529 | Likely Pathogenic | -4.15 | Deleterious | 0.946 | Possibly Damaging | 0.673 | Possibly Damaging | 1.81 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1221 | 0.1051 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||||
| c.3788T>G | I1263S 2D AIThe SynGAP1 missense variant I1263S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -8.074 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.493 | Likely Benign | -4.97 | Deleterious | 0.984 | Probably Damaging | 0.825 | Possibly Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.3242 | 0.0910 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3789T>G | I1263M 2D AIThe SynGAP1 missense variant I1263M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs 2 benign). High‑accuracy methods are not available: AlphaMissense‑Optimized is benign, but AlphaMissense‑Default is pathogenic; Foldetta results are missing. Overall, the majority of predictions (five pathogenic vs four benign) lean toward a pathogenic impact. This conclusion does not contradict ClinVar status, as the variant has no existing ClinVar classification. Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -2.839 | Likely Benign | 0.701 | Likely Pathogenic | Likely Benign | 0.291 | Likely Benign | -2.49 | Neutral | 0.968 | Probably Damaging | 0.789 | Possibly Damaging | 1.81 | Pathogenic | 0.00 | Affected | 0.0654 | 0.2170 | 2 | 1 | -2.6 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.378C>A | F126L 2D AIThe SynGAP1 missense variant F126L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The high‑accuracy consensus (SGM‑Consensus) classifies the variant as Likely Benign, while the high‑accuracy AlphaMissense‑Optimized result is inconclusive and Foldetta data are unavailable. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict the ClinVar status, which contains no pathogenic claim. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -0.992 | Likely Benign | 0.948 | Likely Pathogenic | Ambiguous | 0.114 | Likely Benign | -2.27 | Neutral | 0.001 | Benign | 0.001 | Benign | 4.01 | Benign | 0.00 | Affected | 0.2780 | 0.3399 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||||||
| c.378C>G | F126L 2D AIThe SynGAP1 missense variant F126L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The high‑accuracy consensus (SGM‑Consensus) classifies the variant as Likely Benign, while the high‑accuracy AlphaMissense‑Optimized result is inconclusive and Foldetta data are unavailable. Overall, the majority of evidence points to a benign impact; the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -0.992 | Likely Benign | 0.948 | Likely Pathogenic | Ambiguous | 0.114 | Likely Benign | -2.27 | Neutral | 0.001 | Benign | 0.001 | Benign | 4.01 | Benign | 0.00 | Affected | 0.2780 | 0.3399 | 2 | 0 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3793A>G | R1265G 2D AIThe SynGAP1 missense variant R1265G is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity all converge on a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all label the change as pathogenic. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic, while a Foldetta stability analysis is unavailable. Based on the unanimous pathogenic predictions and the lack of any benign calls, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status (which has no entry). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | -12.732 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.523 | Likely Pathogenic | -5.80 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 2.27 | Pathogenic | 0.00 | Affected | 0.3520 | 0.3759 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||
| c.3793A>T | R1265W 2D AIThe SynGAP1 missense variant R1265W is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity uniformly classify the variant as deleterious: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a pathogenic effect. No tool in the dataset predicts a benign outcome, so the benign group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized indicates a pathogenic change, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. Foldetta results are not available, so they do not influence the conclusion. Overall, the variant is most likely pathogenic based on the consensus of predictive tools, and this assessment is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | -14.584 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.505 | Likely Pathogenic | -6.54 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.23 | Pathogenic | 0.00 | Affected | 0.1166 | 0.3868 | 2 | -3 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.3794G>A | R1265K 2D AIThe SynGAP1 missense variant R1265K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the available predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | -5.223 | Likely Benign | 0.951 | Likely Pathogenic | Ambiguous | 0.344 | Likely Benign | -2.49 | Neutral | 0.992 | Probably Damaging | 0.987 | Probably Damaging | 2.38 | Pathogenic | 0.00 | Affected | 0.4829 | 0.3632 | 3 | 2 | 0.6 | -28.01 | |||||||||||||||||||||||||||||||||||||||
| c.3794G>C | R1265T 2D AIThe SynGAP1 missense variant R1265T is reported in ClinVar as Pathogenic (ClinVar ID 522047.0) and is not found in gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic. No tool in the dataset predicts a benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a Likely Pathogenic verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of all available predictions supports a pathogenic classification, which is consistent with the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | Likely Pathogenic | 1 | -10.129 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.529 | Likely Pathogenic | -4.97 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 2.29 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1947 | 0.4618 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||
| c.3794G>T | R1265M 2D AIThe SynGAP1 missense variant R1265M is not reported in ClinVar and has no entries in gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus confirms a likely pathogenic status; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that R1265M is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | -13.657 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.495 | Likely Benign | -4.97 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.25 | Pathogenic | 0.00 | Affected | 0.1442 | 0.4082 | 0 | -1 | 6.4 | -24.99 | ||||||||||||||||||||||||||||||||||||||
| c.3795G>C | R1265S 2D AIThe SynGAP1 missense variant R1265S is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | -9.874 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.289 | Likely Benign | -4.96 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.3525 | 0.3993 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3795G>T | R1265S 2D AIThe SynGAP1 missense variant R1265S is not reported in ClinVar and has no entry in gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | -9.874 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.289 | Likely Benign | -4.96 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.3525 | 0.3993 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||
| c.3796C>A | L1266M 2D AIThe SynGAP1 missense variant L1266M is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and PROVEAN, whereas a larger set—polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict pathogenicity. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments are mixed: AlphaMissense‑Optimized yields an uncertain result, SGM‑Consensus remains likely pathogenic, and Foldetta data are unavailable. Overall, the majority of evidence points toward a pathogenic effect for L1266M. This conclusion does not conflict with ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.802655 | Binding | 0.868 | 0.602 | 0.000 | -8.257 | Likely Pathogenic | 0.938 | Likely Pathogenic | Ambiguous | 0.126 | Likely Benign | -1.67 | Neutral | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 2.12 | Pathogenic | 0.00 | Affected | 0.0627 | 0.3113 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3796C>G | L1266V 2D AIThe SynGAP1 missense variant L1266V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign outcome, whereas all other evaluated algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.802655 | Binding | 0.868 | 0.602 | 0.000 | -10.024 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.125 | Likely Benign | -2.53 | Deleterious | 0.995 | Probably Damaging | 0.890 | Possibly Damaging | 2.16 | Pathogenic | 0.00 | Affected | 0.1301 | 0.3118 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3797T>A | L1266Q 2D AIThe SynGAP1 missense variant L1266Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, which scores the variant as benign. All other evaluated predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—classify the variant as pathogenic or likely pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates likely pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence points to a pathogenic effect for L1266Q, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.802655 | Binding | 0.868 | 0.602 | 0.000 | -16.101 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.374 | Likely Benign | -5.02 | Deleterious | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.12 | Pathogenic | 0.00 | Affected | 0.0906 | 0.1249 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||||||
| c.3797T>C | L1266P 2D AIThe SynGAP1 missense variant L1266P is reported in gnomAD (6‑33447845‑T‑C) but has no ClinVar entry. Prediction tools cluster into two groups: benign predictions are made only by REVEL, whereas all other evaluated algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic effect. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized returns a pathogenic score, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Pathogenic.” The Foldetta stability analysis is unavailable for this variant. Overall, the consensus of both general and high‑accuracy predictors points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.802655 | Binding | 0.868 | 0.602 | 0.000 | 6-33447845-T-C | -16.566 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.424 | Likely Benign | -5.83 | Deleterious | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3150 | 0.2283 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||
| c.3797T>G | L1266R 2D AISynGAP1 missense variant L1266R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized scores the variant as pathogenic, and the SGM‑Consensus confirms a likely pathogenic classification. Foldetta results are unavailable for this variant. Overall, the preponderance of evidence from multiple in silico predictors and high‑accuracy tools points to a pathogenic effect, with no conflict from ClinVar status (which has no entry). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.802655 | Binding | 0.868 | 0.602 | 0.000 | -16.676 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.391 | Likely Benign | -5.04 | Deleterious | 0.996 | Probably Damaging | 0.951 | Probably Damaging | 2.13 | Pathogenic | 0.00 | Affected | 0.1063 | 0.0891 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||
| c.3799A>C | M1267L 2D AIThe SynGAP1 missense variant M1267L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, the majority of evidence (five pathogenic‑predicting tools versus three benign) indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -1.766 | Likely Benign | 0.399 | Ambiguous | Likely Benign | 0.293 | Likely Benign | -2.52 | Deleterious | 0.813 | Possibly Damaging | 0.456 | Possibly Damaging | 2.36 | Pathogenic | 0.00 | Affected | 0.1392 | 0.3626 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3799A>G | M1267V 2D AIThe SynGAP1 missense variant M1267V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated tools (five pathogenic vs. three benign) indicate a pathogenic effect. This prediction is not contradicted by ClinVar status, as the variant is not yet catalogued there. Thus, based on current computational evidence, the M1267V variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -2.249 | Likely Benign | 0.537 | Ambiguous | Likely Benign | 0.254 | Likely Benign | -3.34 | Deleterious | 0.959 | Probably Damaging | 0.672 | Possibly Damaging | 2.35 | Pathogenic | 0.00 | Affected | 0.3057 | 0.3241 | 2 | 1 | 2.3 | -32.06 | |||||||||||||||||||||||||||||||||||||||
| c.3799A>T | M1267L 2D AIThe SynGAP1 missense variant M1267L is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence (five pathogenic‑predicting tools versus three benign‑predicting tools) indicates that M1267L is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -1.766 | Likely Benign | 0.399 | Ambiguous | Likely Benign | 0.293 | Likely Benign | -2.52 | Deleterious | 0.813 | Possibly Damaging | 0.456 | Possibly Damaging | 2.36 | Pathogenic | 0.00 | Affected | 0.1392 | 0.3626 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.379C>T | R127W 2D AISynGAP1 missense variant R127W is listed in ClinVar with an Uncertain significance status and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 HumVar, ESM1b, and FATHMM, while pathogenic calls come from PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is Uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta stability analysis is unavailable. Consequently, the evidence does not favor a clear benign or pathogenic outcome; the predictions are balanced and align with the ClinVar designation of Uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.497853 | Structured | 0.711716 | Binding | 0.333 | 0.870 | 0.625 | Uncertain | 1 | -4.776 | Likely Benign | 0.806 | Likely Pathogenic | Ambiguous | 0.118 | Likely Benign | -2.98 | Deleterious | 0.989 | Probably Damaging | 0.420 | Benign | 3.88 | Benign | 0.00 | Affected | 0.1583 | 0.3005 | 2 | -3 | 3.6 | 30.03 | ||||||||||||||||||||||||||||||||||||||
| c.37A>C | I13L 2D AIThe SynGAP1 missense variant I13L is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign outcome. Foldetta results are not available, so they do not influence the assessment. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | -2.144 | Likely Benign | 0.100 | Likely Benign | Likely Benign | 0.095 | Likely Benign | 0.07 | Neutral | 0.002 | Benign | 0.001 | Benign | 4.19 | Benign | 0.00 | Affected | 0.1055 | 0.4923 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.37A>G | I13V 2D AIThe SynGAP1 missense variant I13V is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the ClinVar designation of uncertain significance rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | Uncertain | 2 | -2.497 | Likely Benign | 0.105 | Likely Benign | Likely Benign | 0.110 | Likely Benign | 0.01 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.25 | Benign | 0.00 | Affected | 0.1548 | 0.4315 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||
| c.37A>T | I13F 2D AIThe SynGAP1 missense variant I13F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | -3.359 | Likely Benign | 0.126 | Likely Benign | Likely Benign | 0.134 | Likely Benign | -0.09 | Neutral | 0.107 | Benign | 0.010 | Benign | 4.04 | Benign | 0.00 | Affected | 0.0654 | 0.3972 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3800T>A | M1267K 2D AIThe SynGAP1 missense variant M1267K is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, favors a pathogenic outcome (3/4 pathogenic). AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. Overall, the majority of evidence points to a deleterious effect, classifying the variant as most likely pathogenic. This assessment does not conflict with ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -6.415 | Likely Benign | 0.917 | Likely Pathogenic | Ambiguous | 0.366 | Likely Benign | -5.01 | Deleterious | 0.884 | Possibly Damaging | 0.581 | Possibly Damaging | 2.31 | Pathogenic | 0.00 | Affected | 0.1373 | 0.0488 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||||||||||
| c.3800T>C | M1267T 2D AIThe SynGAP1 missense variant M1267T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) likewise indicates likely pathogenicity. No Foldetta stability prediction is available for this variant. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -5.315 | Likely Benign | 0.958 | Likely Pathogenic | Likely Pathogenic | 0.270 | Likely Benign | -5.00 | Deleterious | 0.982 | Probably Damaging | 0.672 | Possibly Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.2036 | 0.1934 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||||||
| c.3800T>G | M1267R 2D AIThe SynGAP1 missense variant M1267R is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. Grouping by consensus, two tools predict benign and six predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM‑Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—classifies the variant as likely pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic impact, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -4.990 | Likely Benign | 0.899 | Likely Pathogenic | Ambiguous | 0.366 | Likely Benign | -5.03 | Deleterious | 0.982 | Probably Damaging | 0.757 | Possibly Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1555 | 0.0837 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||||||||||
| c.3801G>A | M1267I 2D AIThe SynGAP1 missense variant M1267I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote). High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for M1267I. This conclusion is not contradicted by ClinVar status, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -1.432 | Likely Benign | 0.936 | Likely Pathogenic | Ambiguous | 0.205 | Likely Benign | -3.33 | Deleterious | 0.959 | Probably Damaging | 0.672 | Possibly Damaging | 2.33 | Pathogenic | 0.00 | Affected | 0.1254 | 0.2741 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3801G>C | M1267I 2D AIThe SynGAP1 missense variant M1267I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) remains Likely Pathogenic; Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for M1267I. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -1.432 | Likely Benign | 0.936 | Likely Pathogenic | Ambiguous | 0.205 | Likely Benign | -3.33 | Deleterious | 0.959 | Probably Damaging | 0.672 | Possibly Damaging | 2.33 | Pathogenic | 0.00 | Affected | 0.1254 | 0.2741 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3801G>T | M1267I 2D AIThe SynGAP1 missense variant M1267I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) remains Likely Pathogenic; Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for M1267I. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -1.432 | Likely Benign | 0.936 | Likely Pathogenic | Ambiguous | 0.205 | Likely Benign | -3.33 | Deleterious | 0.959 | Probably Damaging | 0.672 | Possibly Damaging | 2.33 | Pathogenic | 0.00 | Affected | 0.1254 | 0.2741 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||
| c.3805G>A | V1269M 2D  AIThe SynGAP1 missense variant V1269M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (majority vote) is pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for V1269M, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.787464 | Binding | 0.843 | 0.647 | 0.125 | -3.743 | Likely Benign | 0.977 | Likely Pathogenic | Likely Pathogenic | 0.300 | Likely Benign | -2.53 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.08 | Pathogenic | 0.00 | Affected | 0.0582 | 0.3676 | 2 | 1 | -2.3 | 32.06 | ||||||||||||||||||||||||||||||||||||||
| c.3805G>C | V1269L 2D  AIThe SynGAP1 missense variant V1269L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of other in silico predictors (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy tools points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.787464 | Binding | 0.843 | 0.647 | 0.125 | -3.572 | Likely Benign | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.299 | Likely Benign | -2.53 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.0683 | 0.4082 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3805G>T | V1269L 2D AIThe SynGAP1 missense variant V1269L is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of other in silico predictors (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of predictions points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.787464 | Binding | 0.843 | 0.647 | 0.125 | -3.572 | Likely Benign | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.299 | Likely Benign | -2.53 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.0683 | 0.4082 | 2 | 1 | -0.4 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3806T>A | V1269E 2D  AIThe SynGAP1 missense change V1269E is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that flag the variant as benign include only REVEL, whereas the remaining predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently classify it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a “Likely Pathogenic” outcome. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as likely pathogenic; the Foldetta stability analysis is unavailable. Overall, the preponderance of evidence points to a pathogenic effect, which does not contradict the ClinVar designation of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.787464 | Binding | 0.843 | 0.647 | 0.125 | Uncertain | 1 | -11.418 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.403 | Likely Benign | -5.05 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.09 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0899 | 0.1557 | -2 | -2 | -7.7 | 29.98 | ||||||||||||||||||||||||||||||||||
| c.3806T>C | V1269A 2D  AIThe SynGAP1 missense variant V1269A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas a larger group predicts pathogenicity: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). AlphaMissense‑Optimized is uncertain, and Foldetta results are unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and no Foldetta data. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.433034 | Structured | 0.787464 | Binding | 0.843 | 0.647 | 0.125 | -6.115 | Likely Benign | 0.954 | Likely Pathogenic | Ambiguous | 0.291 | Likely Benign | -3.38 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.15 | Pathogenic | 0.00 | Affected | 0.2699 | 0.2312 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||||||||||||||||
| c.3806T>G | V1269G 2D AIThe SynGAP1 missense variant V1269G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, whereas all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic outcome. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus is pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.433034 | Structured | 0.787464 | Binding | 0.843 | 0.647 | 0.125 | -9.927 | Likely Pathogenic | 0.969 | Likely Pathogenic | Likely Pathogenic | 0.420 | Likely Benign | -5.91 | Deleterious | 0.995 | Probably Damaging | 0.999 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.2180 | 0.2557 | -1 | -3 | -4.6 | -42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3808G>A | E1270K 2D AIThe SynGAP1 missense variant E1270K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: REVEL predicts a benign outcome, whereas all other evaluated algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the preponderance of evidence indicates that E1270K is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.771865 | Binding | 0.805 | 0.659 | 0.250 | -12.549 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.413 | Likely Benign | -3.37 | Deleterious | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 2.07 | Pathogenic | 0.00 | Affected | 0.1780 | 0.6276 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||||||||
| c.3808G>C | E1270Q 2D AIThe SynGAP1 missense variant E1270Q is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Prediction tools that agree on a benign effect include only REVEL, which scores the substitution as benign. In contrast, the majority of in silico predictors—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—classify the change as pathogenic. The high‑accuracy AlphaMissense‑Optimized assessment is uncertain, while the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic outcome. Foldetta predictions are unavailable for this variant. Overall, the preponderance of pathogenic predictions, together with the SGM Consensus result, indicates that E1270Q is most likely pathogenic; this conclusion does not contradict ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.771865 | Binding | 0.805 | 0.659 | 0.250 | -8.645 | Likely Pathogenic | 0.935 | Likely Pathogenic | Ambiguous | 0.330 | Likely Benign | -2.53 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.06 | Pathogenic | 0.00 | Affected | 0.0919 | 0.5858 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||
| c.3809A>C | E1270A 2D AIThe SynGAP1 missense change E1270A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: REVEL is the only score that flags the variant as benign, whereas PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) all predict pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports a pathogenic effect, and the SGM‑Consensus also indicates a likely pathogenic outcome. No Foldetta (FoldX‑MD/Rosetta stability) result is available, so it does not influence the assessment. Overall, the preponderance of evidence points to a pathogenic impact for E1270A, and this conclusion is consistent with the absence of a ClinVar entry (i.e., no contradictory clinical classification). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.771865 | Binding | 0.805 | 0.659 | 0.250 | -12.081 | Likely Pathogenic | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.388 | Likely Benign | -5.04 | Deleterious | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 2.06 | Pathogenic | 0.00 | Affected | 0.3024 | 0.5126 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||
| c.3809A>G | E1270G 2D AIThe SynGAP1 missense variant E1270G is listed in gnomAD (ID 6‑33447857‑A‑G) and has no ClinVar entry. Prediction tools cluster into two groups: the single benign predictor REVEL, and a consensus of pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as likely pathogenic, while Foldetta’s protein‑folding stability analysis is unavailable. Based on the preponderance of pathogenic predictions and the absence of a benign consensus, the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.771865 | Binding | 0.805 | 0.659 | 0.250 | 6-33447857-A-G | -12.022 | Likely Pathogenic | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.385 | Likely Benign | -5.90 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 2.05 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2415 | 0.5452 | -2 | 0 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||
| c.3809A>T | E1270V 2D AIThe SynGAP1 missense variant E1270V is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus agrees. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that E1270V is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.771865 | Binding | 0.805 | 0.659 | 0.250 | -13.293 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.408 | Likely Benign | -5.90 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 0.0570 | 0.6461 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||
| c.3810G>C | E1270D 2D AIThe SynGAP1 missense variant E1270D is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.771865 | Binding | 0.805 | 0.659 | 0.250 | -9.379 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.220 | Likely Benign | -2.53 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.26 | Pathogenic | 0.00 | Affected | 0.1655 | 0.3271 | 3 | 2 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3810G>T | E1270D 2D AIThe SynGAP1 missense variant E1270D is catalogued in gnomAD (6-33447858‑G‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: the single benign prediction comes from REVEL, while all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Pathogenic.” The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence indicates that E1270D is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.771865 | Binding | 0.805 | 0.659 | 0.250 | 6-33447858-G-T | -9.379 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.219 | Likely Benign | -2.53 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.26 | Pathogenic | 0.00 | Affected | 0.1655 | 0.3271 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||
| c.3811G>A | E1271K 2D AIThe SynGAP1 missense variant E1271K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized, while pathogenic calls come from PROVEAN, polyPhen‑2 HumDiv, SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further reveal AlphaMissense‑Optimized as benign, whereas Foldetta (combining FoldX‑MD and Rosetta) has no available result. Overall, the preponderance of evidence—both from general predictors and the SGM Consensus—leans toward pathogenicity. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.767529 | Binding | 0.832 | 0.666 | 0.375 | -2.295 | Likely Benign | 0.689 | Likely Pathogenic | Likely Benign | 0.192 | Likely Benign | -3.24 | Deleterious | 0.905 | Possibly Damaging | 0.433 | Benign | 2.07 | Pathogenic | 0.00 | Affected | 0.1780 | 0.5888 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3811G>C | E1271Q 2D AIThe SynGAP1 missense variant E1271Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are not available. Overall, the majority of predictions (six benign vs. four pathogenic) support a benign classification. This conclusion is not contradicted by ClinVar, which has no entry for this variant. Thus, based on the available computational evidence, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.767529 | Binding | 0.832 | 0.666 | 0.375 | -3.085 | Likely Benign | 0.282 | Likely Benign | Likely Benign | 0.175 | Likely Benign | -2.40 | Neutral | 0.951 | Possibly Damaging | 0.617 | Possibly Damaging | 2.06 | Pathogenic | 0.00 | Affected | 0.0955 | 0.5470 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3812A>C | E1271A 2D AIThe SynGAP1 missense variant E1271A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessment shows AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves as pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of conventional tools and the high‑accuracy consensus favor a pathogenic interpretation. This prediction does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.483068 | Structured | 0.767529 | Binding | 0.832 | 0.666 | 0.375 | -3.817 | Likely Benign | 0.375 | Ambiguous | Likely Benign | 0.298 | Likely Benign | -4.79 | Deleterious | 0.951 | Possibly Damaging | 0.433 | Benign | 2.06 | Pathogenic | 0.00 | Affected | 0.2985 | 0.4959 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3812A>G | E1271G 2D AIThe SynGAP1 missense variant E1271G is catalogued in gnomAD (ID 6‑33447860‑A‑G) but has no entry in ClinVar. Functional prediction tools show a split consensus: benign calls come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts a benign effect, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (two pathogenic versus one benign vote). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence, including the SGM Consensus, indicates a pathogenic impact. This prediction is not contradicted by ClinVar, as no ClinVar classification exists for E1271G. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.483068 | Structured | 0.767529 | Binding | 0.832 | 0.666 | 0.375 | 6-33447860-A-G | -4.857 | Likely Benign | 0.393 | Ambiguous | Likely Benign | 0.288 | Likely Benign | -5.37 | Deleterious | 0.905 | Possibly Damaging | 0.538 | Possibly Damaging | 2.05 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2513 | 0.5485 | -2 | 0 | 3.1 | -72.06 | 10.1016/j.ajhg.2020.11.011 | ||||||||||||||||||||||||||||||||||||
| c.3812A>T | E1271V 2D AISynGAP1 missense variant E1271V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and ESM1b, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. AlphaMissense‑Optimized yields an uncertain result, and no Foldetta stability assessment is available. Considering the majority of high‑confidence tools and the consensus score, the variant is most likely pathogenic. This assessment does not contradict any ClinVar annotation, as no ClinVar entry exists for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.767529 | Binding | 0.832 | 0.666 | 0.375 | -6.961 | Likely Benign | 0.848 | Likely Pathogenic | Ambiguous | 0.303 | Likely Benign | -5.64 | Deleterious | 0.995 | Probably Damaging | 0.846 | Possibly Damaging | 2.02 | Pathogenic | 0.00 | Affected | 0.0620 | 0.6106 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.3813G>C | E1271D 2D AIThe SynGAP1 missense variant E1271D is not reported in ClinVar and has no entry in gnomAD, indicating it is not catalogued in these databases. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Benign; the Foldetta stability prediction is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.767529 | Binding | 0.832 | 0.666 | 0.375 | -4.816 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.122 | Likely Benign | -1.59 | Neutral | 0.004 | Benign | 0.008 | Benign | 2.26 | Pathogenic | 0.00 | Affected | 0.1729 | 0.3471 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3813G>T | E1271D 2D AIThe SynGAP1 missense variant E1271D is not reported in ClinVar and has no entry in gnomAD, indicating it is not catalogued in these databases. Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Benign; the Foldetta stability prediction is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.767529 | Binding | 0.832 | 0.666 | 0.375 | -4.816 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.121 | Likely Benign | -1.59 | Neutral | 0.004 | Benign | 0.008 | Benign | 2.26 | Pathogenic | 0.00 | Affected | 0.1729 | 0.3471 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3814G>A | E1272K 2D AIThe SynGAP1 E1272K missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools cluster into two groups: benign predictions are made by REVEL and ESM1b, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; the SGM‑Consensus score is “Likely Pathogenic.” High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Pathogenic, and Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact. Thus, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.766082 | Binding | 0.799 | 0.677 | 0.500 | -4.227 | Likely Benign | 0.956 | Likely Pathogenic | Likely Pathogenic | 0.312 | Likely Benign | -3.37 | Deleterious | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 2.28 | Pathogenic | 0.00 | Affected | 0.1544 | 0.5488 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3814G>C | E1272Q 2D AIThe SynGAP1 E1272Q missense variant is catalogued in gnomAD (variant ID 6‑33447862‑G‑C) but has no ClinVar entry. Prediction tools cluster into two groups: benign calls come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, a majority‑vote aggregator of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further show AlphaMissense‑Optimized as benign, SGM‑Consensus as Likely Pathogenic, and Foldetta (a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the preponderance of evidence (seven pathogenic versus three benign predictions) indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.766082 | Binding | 0.799 | 0.677 | 0.500 | 6-33447862-G-C | 1 | 6.45e-7 | -3.000 | Likely Benign | 0.651 | Likely Pathogenic | Likely Benign | 0.207 | Likely Benign | -2.53 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.25 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0746 | 0.5258 | 2 | 2 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||
| c.3815A>C | E1272A 2D AIThe SynGAP1 missense variant E1272A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus indicates a likely pathogenic outcome; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic impact for E1272A. This prediction is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.766082 | Binding | 0.799 | 0.677 | 0.500 | -0.783 | Likely Benign | 0.779 | Likely Pathogenic | Likely Benign | 0.261 | Likely Benign | -5.05 | Deleterious | 0.997 | Probably Damaging | 0.989 | Probably Damaging | 2.26 | Pathogenic | 0.00 | Affected | 0.3243 | 0.5547 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.3815A>G | E1272G 2D AIThe SynGAP1 missense variant E1272G is catalogued in gnomAD (ID 6‑33447863‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL and ESM1b, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score, which is labeled Likely Pathogenic. The high‑accuracy AlphaMissense‑Optimized assessment is uncertain, and the Foldetta protein‑folding stability analysis is not available for this residue. Overall, the majority of evidence points toward a deleterious effect, so the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for E1272G. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.766082 | Binding | 0.799 | 0.677 | 0.500 | 6-33447863-A-G | -1.919 | Likely Benign | 0.863 | Likely Pathogenic | Ambiguous | 0.287 | Likely Benign | -5.89 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 2.22 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2804 | 0.5072 | -2 | 0 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||
| c.3815A>T | E1272V 2D AIThe SynGAP1 missense variant E1272V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all indicate pathogenicity. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. AlphaMissense‑Optimized returns an uncertain result, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available prediction for this variant. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic; this assessment does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.766082 | Binding | 0.799 | 0.677 | 0.500 | -3.628 | Likely Benign | 0.934 | Likely Pathogenic | Ambiguous | 0.278 | Likely Benign | -5.90 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.22 | Pathogenic | 0.00 | Affected | 0.0424 | 0.5894 | -2 | -2 | 7.7 | -29.98 | |||||||||||||||||||||||||||||||||||||||
| c.3816G>C | E1272D 2D AIThe SynGAP1 missense variant E1272D has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus) all predict a pathogenic impact. High‑accuracy assessments further show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely pathogenic; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a pathogenic effect. There is no ClinVar classification to contradict this conclusion, so the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.766082 | Binding | 0.799 | 0.677 | 0.500 | -4.781 | Likely Benign | 0.751 | Likely Pathogenic | Likely Benign | 0.189 | Likely Benign | -2.53 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.27 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1450 | 0.3471 | 2 | 3 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||
| c.3816G>T | E1272D 2D AIThe SynGAP1 missense variant E1272D is listed in ClinVar with no submitted interpretation and is present in gnomAD (variant ID 6‑33447864‑G‑T). Functional prediction tools show mixed results: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score. The high‑accuracy AlphaMissense‑Optimized tool classifies the change as benign, while the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic effect. No Foldetta stability assessment is available for this variant. Overall, the majority of conventional predictors and the consensus score lean toward pathogenicity, which is consistent with the SGM‑Consensus designation but contradicts the benign calls from AlphaMissense‑Optimized and a few other tools. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion aligns with the SGM‑Consensus prediction rather than the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.766082 | Binding | 0.799 | 0.677 | 0.500 | 6-33447864-G-T | -4.781 | Likely Benign | 0.751 | Likely Pathogenic | Likely Benign | 0.188 | Likely Benign | -2.53 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.27 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1450 | 0.3471 | 2 | 3 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||
| c.3817C>A | L1273M 2D AIThe SynGAP1 missense variant L1273M is catalogued in gnomAD (ID 6‑33447865‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect for the variant. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.773625 | Binding | 0.747 | 0.677 | 0.500 | 6-33447865-C-A | -5.375 | Likely Benign | 0.503 | Ambiguous | Likely Benign | 0.161 | Likely Benign | -1.68 | Neutral | 0.983 | Probably Damaging | 0.874 | Possibly Damaging | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0824 | 0.3932 | 2 | 4 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||
| c.3817C>G | L1273V 2D AIThe SynGAP1 missense variant L1273V is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (two pathogenic versus one benign vote). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence, particularly the SGM Consensus, indicates that the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.773625 | Binding | 0.747 | 0.677 | 0.500 | -6.014 | Likely Benign | 0.539 | Ambiguous | Likely Benign | 0.111 | Likely Benign | -2.50 | Deleterious | 0.773 | Possibly Damaging | 0.287 | Benign | 2.20 | Pathogenic | 0.00 | Affected | 0.1556 | 0.3178 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3818T>A | L1273Q 2D AIThe SynGAP1 missense variant L1273Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic and the SGM‑Consensus is labeled Likely Pathogenic; Foldetta results are unavailable. Based on the preponderance of evidence, the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.599170 | Disordered | 0.773625 | Binding | 0.747 | 0.677 | 0.500 | -6.813 | Likely Benign | 0.957 | Likely Pathogenic | Likely Pathogenic | 0.423 | Likely Benign | -5.05 | Deleterious | 0.997 | Probably Damaging | 0.950 | Probably Damaging | 2.13 | Pathogenic | 0.00 | Affected | 0.1139 | 0.1119 | -2 | -2 | -7.3 | 14.97 | |||||||||||||||||||||||||||||||||||||||
| c.3818T>C | L1273P 2D AIThe SynGAP1 missense variant L1273P is not reported in ClinVar (ClinVar status: not listed) but is present in the gnomAD database (gnomAD ID: 6‑33447866‑T‑C). Functional prediction tools uniformly indicate a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic. No tool in the dataset predicts a benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the consensus of predictive algorithms strongly supports a pathogenic classification, and this assessment does not contradict the ClinVar status, which simply lacks an entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.599170 | Disordered | 0.773625 | Binding | 0.747 | 0.677 | 0.500 | 6-33447866-T-C | -9.443 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.523 | Likely Pathogenic | -5.87 | Deleterious | 0.997 | Probably Damaging | 0.950 | Probably Damaging | 2.12 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3836 | 0.1343 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||
| c.3818T>G | L1273R 2D AIThe SynGAP1 missense variant L1273R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default all classify the variant as damaging. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Based on the preponderance of pathogenic predictions and the SGM‑Consensus outcome, the variant is most likely pathogenic; this assessment is not contradicted by any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.599170 | Disordered | 0.773625 | Binding | 0.747 | 0.677 | 0.500 | -6.252 | Likely Benign | 0.946 | Likely Pathogenic | Ambiguous | 0.431 | Likely Benign | -5.05 | Deleterious | 0.997 | Probably Damaging | 0.934 | Probably Damaging | 2.13 | Pathogenic | 0.00 | Affected | 0.1369 | 0.0761 | -3 | -2 | -8.3 | 43.03 | |||||||||||||||||||||||||||||||||||||||
| c.3820C>T | R1274C 2D 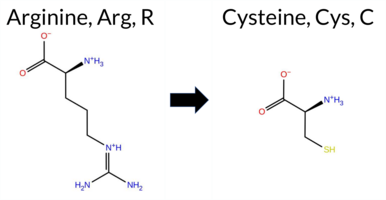 AIThe SynGAP1 missense variant R1274C is listed in ClinVar with an “Uncertain” significance and is present in gnomAD (ID 6‑33447868‑C‑T). Prediction tools that agree on benign impact include REVEL, ESM1b, and AlphaMissense‑Optimized, while those that predict pathogenicity are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). AlphaMissense‑Default remains uncertain, and Foldetta results are unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus as pathogenic, and no Foldetta data to weigh in. Overall, the majority of evaluated tools (seven pathogenic vs. three benign) suggest a pathogenic effect. This consensus does not contradict the ClinVar “Uncertain” status, which remains inconclusive. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.613573 | Disordered | 0.779985 | Binding | 0.746 | 0.688 | 0.625 | Uncertain | 1 | 6-33447868-C-T | -6.467 | Likely Benign | 0.439 | Ambiguous | Likely Benign | 0.170 | Likely Benign | -5.22 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.46 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3232 | 0.1517 | -4 | -3 | 7.0 | -53.05 | |||||||||||||||||||||||||||||||||||
| c.3823C>T | R1275W 2D AIThe SynGAP1 missense variant R1275W is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447871‑C‑T). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas a majority (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b) predict a pathogenic impact; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a split vote (2 pathogenic, 1 benign, 1 uncertain). Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the balance of evidence leans toward a pathogenic classification, and this assessment does not contradict ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.648219 | Disordered | 0.790317 | Binding | 0.723 | 0.697 | 0.500 | 6-33447871-C-T | 3 | 1.93e-6 | -9.895 | Likely Pathogenic | 0.513 | Ambiguous | Likely Benign | 0.161 | Likely Benign | -4.76 | Deleterious | 0.999 | Probably Damaging | 0.875 | Possibly Damaging | 2.51 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1269 | 0.2372 | -3 | 2 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||||
| c.3826G>A | D1276N 2D AIThe SynGAP1 missense variant D1276N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, more tools (five) predict pathogenicity than benign (three), and the high‑accuracy consensus also leans pathogenic. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | 3.371 | Likely Benign | 0.473 | Ambiguous | Likely Benign | 0.242 | Likely Benign | -3.68 | Deleterious | 0.899 | Possibly Damaging | 0.581 | Possibly Damaging | 1.22 | Pathogenic | 0.00 | Affected | 0.0883 | 0.5213 | 2 | 1 | 0.0 | -0.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3826G>C | D1276H 2D AIThe SynGAP1 missense variant D1276H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates majority votes from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic. High‑accuracy assessments further highlight discordance: AlphaMissense‑Optimized predicts a benign effect, whereas the SGM‑Consensus (a high‑accuracy consensus) indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions support a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | 0.715 | Likely Benign | 0.778 | Likely Pathogenic | Likely Benign | 0.321 | Likely Benign | -5.08 | Deleterious | 0.996 | Probably Damaging | 0.898 | Possibly Damaging | 1.19 | Pathogenic | 0.00 | Affected | 0.1077 | 0.5697 | 1 | -1 | 0.3 | 22.05 | |||||||||||||||||||||||||||||||||||||||
| c.3826G>T | D1276Y 2D AIThe SynGAP1 missense variant D1276Y is catalogued in gnomAD (ID 6‑33447874‑G‑T) but has no ClinVar entry. Functional prediction tools fall into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus indicates a likely pathogenic effect; Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not contradict any ClinVar status, as none is currently reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | 6-33447874-G-T | -1.558 | Likely Benign | 0.768 | Likely Pathogenic | Likely Benign | 0.325 | Likely Benign | -6.66 | Deleterious | 0.999 | Probably Damaging | 0.928 | Probably Damaging | 1.18 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.0385 | 0.5082 | -3 | -4 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||
| c.3827A>C | D1276A 2D AIThe SynGAP1 missense variant D1276A has no ClinVar entry and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further support this split: AlphaMissense‑Optimized reports a benign effect, whereas the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. No Foldetta stability analysis is available for this residue. Overall, the majority of computational evidence points to a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently contains no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | -0.008 | Likely Benign | 0.710 | Likely Pathogenic | Likely Benign | 0.319 | Likely Benign | -5.87 | Deleterious | 0.816 | Possibly Damaging | 0.495 | Possibly Damaging | 1.21 | Pathogenic | 0.00 | Affected | 0.3458 | 0.5050 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||||
| c.3827A>G | D1276G 2D AIThe SynGAP1 missense variant D1276G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains Likely Pathogenic; Foldetta results are unavailable. Overall, the majority of predictions and the SGM‑Consensus support a pathogenic interpretation, and there is no ClinVar record to contradict this assessment. Thus, the variant is most likely pathogenic based on the available computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | 0.509 | Likely Benign | 0.601 | Likely Pathogenic | Likely Benign | 0.293 | Likely Benign | -4.93 | Deleterious | 0.899 | Possibly Damaging | 0.655 | Possibly Damaging | 1.21 | Pathogenic | 0.00 | Affected | 0.3429 | 0.5247 | 1 | -1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.3827A>T | D1276V 2D AIThe SynGAP1 D1276V missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default) predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is unavailable for this variant. Based on the preponderance of pathogenic predictions and the SGM‑Consensus result, the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | -0.725 | Likely Benign | 0.851 | Likely Pathogenic | Ambiguous | 0.331 | Likely Benign | -6.66 | Deleterious | 0.984 | Probably Damaging | 0.825 | Possibly Damaging | 1.19 | Pathogenic | 0.00 | Affected | 0.0614 | 0.5207 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.3828C>A | D1276E 2D AIThe SynGAP1 missense variant D1276E is catalogued in gnomAD (ID 6‑33447876‑C‑A) but has no ClinVar submission. Functional prediction tools show a split assessment: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Optimized, whereas pathogenic calls arise from PROVEAN, SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. A high‑accuracy consensus (SGM) that aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a pathogenic verdict. AlphaMissense‑Optimized alone predicts benign, and no Foldetta stability data are available. Overall, the majority of individual predictors favor a benign effect, but the SGM consensus contradicts this by labeling the variant pathogenic. Because ClinVar contains no classification, there is no external evidence to resolve the discrepancy. Thus, based on the current computational evidence, the variant is most likely benign, though the SGM consensus suggests a possible pathogenic interpretation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | 6-33447876-C-A | -0.388 | Likely Benign | 0.416 | Ambiguous | Likely Benign | 0.091 | Likely Benign | -2.64 | Deleterious | 0.027 | Benign | 0.020 | Benign | 1.26 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1027 | 0.5365 | 2 | 3 | 0.0 | 14.03 | |||||||||||||||||||||||||||||||||||||
| c.3828C>G | D1276E 2D AIThe SynGAP1 missense variant D1276E is not reported in ClinVar and is absent from gnomAD. Consensus from routine in silico predictors shows a split: benign calls from REVEL, polyPhen‑2 (HumDiv and HumVar), and ESM1b, versus pathogenic calls from PROVEAN, SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments give a benign result from AlphaMissense‑Optimized, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic. Foldetta, a protein‑folding stability method, has no available output for this residue. Overall, the balance of evidence tilts toward a benign interpretation, and this conclusion is not in conflict with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.802156 | Binding | 0.636 | 0.705 | 0.625 | -0.388 | Likely Benign | 0.416 | Ambiguous | Likely Benign | 0.092 | Likely Benign | -2.64 | Deleterious | 0.027 | Benign | 0.020 | Benign | 1.26 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1027 | 0.5365 | 2 | 3 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||
| c.3829C>A | H1277N 2D AIThe SynGAP1 missense variant H1277N is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447877‑C‑A). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | 6-33447877-C-A | -3.347 | Likely Benign | 0.193 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -4.96 | Deleterious | 0.224 | Benign | 0.120 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1562 | 0.1250 | 1 | 2 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||
| c.3829C>G | H1277D 2D AIThe SynGAP1 missense variant H1277D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447877‑C‑G). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), and ESM1b, while those that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. AlphaMissense‑Default is uncertain, whereas AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a pathogenic outcome (two pathogenic, one benign, one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy and consensus predictions indicate a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | 6-33447877-C-G | -4.632 | Likely Benign | 0.537 | Ambiguous | Likely Benign | 0.172 | Likely Benign | -6.38 | Deleterious | 0.411 | Benign | 0.091 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2389 | 0.1266 | -1 | 1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||
| c.3829C>T | H1277Y 2D AIThe SynGAP1 missense variant H1277Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the balance of evidence (5 benign vs 4 pathogenic predictions) leans toward a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | -4.288 | Likely Benign | 0.232 | Likely Benign | Likely Benign | 0.136 | Likely Benign | -4.32 | Deleterious | 0.812 | Possibly Damaging | 0.298 | Benign | 2.36 | Pathogenic | 0.00 | Affected | 0.0742 | 0.3034 | 0 | 2 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3830A>C | H1277P 2D AIThe SynGAP1 missense variant H1277P is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447878‑A‑C). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, and this is consistent with the lack of a ClinVar pathogenic annotation. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | 6-33447878-A-C | -3.829 | Likely Benign | 0.324 | Likely Benign | Likely Benign | 0.237 | Likely Benign | -7.14 | Deleterious | 0.586 | Possibly Damaging | 0.287 | Benign | 2.12 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2266 | 0.3393 | -2 | 0 | 1.6 | -40.02 | |||||||||||||||||||||||||||||||||||||
| c.3830A>G | H1277R 2D AIThe SynGAP1 missense variant H1277R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | -2.940 | Likely Benign | 0.306 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -5.60 | Deleterious | 0.259 | Benign | 0.066 | Benign | 2.15 | Pathogenic | 0.00 | Affected | 0.1710 | 0.1562 | 2 | 0 | -1.3 | 19.05 | ||||||||||||||||||||||||||||||||||||||||
| c.3830A>T | H1277L 2D AIThe SynGAP1 missense variant H1277L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | -2.949 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.215 | Likely Benign | -7.93 | Deleterious | 0.224 | Benign | 0.091 | Benign | 2.17 | Pathogenic | 0.00 | Affected | 0.0822 | 0.4158 | -2 | -3 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3831C>A | H1277Q 2D AIThe SynGAP1 missense variant H1277Q is reported in gnomAD (ID 6‑33447879‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b; pathogenic predictions come from PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta data are unavailable. Overall, the majority of evidence points toward a benign effect, and this conclusion does not conflict with the absence of a ClinVar classification. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | 6-33447879-C-A | -3.323 | Likely Benign | 0.325 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -5.34 | Deleterious | 0.004 | Benign | 0.010 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1311 | 0.2351 | 0 | 3 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||
| c.3831C>G | H1277Q 2D AIThe SynGAP1 missense variant H1277Q is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b; pathogenic predictions come from PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs. 2 pathogenic). Foldetta results are not available. Overall, the majority of evidence points toward a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | -3.323 | Likely Benign | 0.325 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -5.34 | Deleterious | 0.004 | Benign | 0.010 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1311 | 0.2351 | 0 | 3 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3838A>C | M1280L 2D AIThe SynGAP1 missense variant M1280L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for M1280L, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -1.391 | Likely Benign | 0.092 | Likely Benign | Likely Benign | 0.204 | Likely Benign | -2.23 | Neutral | 0.052 | Benign | 0.017 | Benign | 2.40 | Pathogenic | 0.00 | Affected | 0.1113 | 0.3186 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3838A>G | M1280V 2D AIThe SynGAP1 missense variant M1280V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for M1280V, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -2.414 | Likely Benign | 0.063 | Likely Benign | Likely Benign | 0.240 | Likely Benign | -2.24 | Neutral | 0.004 | Benign | 0.008 | Benign | 2.36 | Pathogenic | 0.00 | Affected | 0.2568 | 0.3137 | 2 | 1 | 2.3 | -32.06 | |||||||||||||||||||||||||||||||||||||||
| c.3838A>T | M1280L 2D AIThe SynGAP1 missense variant M1280L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a benign impact. The predictions do not contradict any ClinVar status, as no ClinVar claim exists for this variant. Thus, based on current computational predictions, the M1280L variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -1.391 | Likely Benign | 0.092 | Likely Benign | Likely Benign | 0.204 | Likely Benign | -2.23 | Neutral | 0.052 | Benign | 0.017 | Benign | 2.40 | Pathogenic | 0.00 | Affected | 0.1113 | 0.3186 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3839T>A | M1280K 2D AIThe SynGAP1 missense variant M1280K is not reported in ClinVar and has no entry in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and consensus methods points to a benign impact for M1280K, and this conclusion does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -2.963 | Likely Benign | 0.223 | Likely Benign | Likely Benign | 0.213 | Likely Benign | -1.42 | Neutral | 0.126 | Benign | 0.041 | Benign | 2.37 | Pathogenic | 0.00 | Affected | 0.1096 | 0.0488 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||||||||||||
| c.3839T>C | M1280T 2D AIThe SynGAP1 missense variant M1280T is catalogued in gnomAD (6‑33447887‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign or likely benign outcome. In contrast, SIFT and FATHMM predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic designation. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | 6-33447887-T-C | 3 | 1.93e-6 | -2.305 | Likely Benign | 0.112 | Likely Benign | Likely Benign | 0.158 | Likely Benign | -2.26 | Neutral | 0.369 | Benign | 0.120 | Benign | 2.34 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1814 | 0.2031 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||
| c.3839T>G | M1280R 2D AIThe SynGAP1 missense variant M1280R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for M1280R, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -2.347 | Likely Benign | 0.209 | Likely Benign | Likely Benign | 0.272 | Likely Benign | -1.97 | Neutral | 0.001 | Benign | 0.001 | Benign | 2.44 | Pathogenic | 0.00 | Affected | 0.1339 | 0.0837 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.3840G>A | M1280I 2D AIThe SynGAP1 missense variant M1280I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -3.723 | Likely Benign | 0.251 | Likely Benign | Likely Benign | 0.166 | Likely Benign | -2.59 | Deleterious | 0.059 | Benign | 0.041 | Benign | 2.35 | Pathogenic | 0.00 | Affected | 0.0995 | 0.2468 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3840G>C | M1280I 2D AIThe SynGAP1 missense variant M1280I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -3.723 | Likely Benign | 0.251 | Likely Benign | Likely Benign | 0.166 | Likely Benign | -2.59 | Deleterious | 0.059 | Benign | 0.041 | Benign | 2.35 | Pathogenic | 0.00 | Affected | 0.0995 | 0.2468 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3840G>T | M1280I 2D AIThe SynGAP1 missense variant M1280I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | -3.723 | Likely Benign | 0.251 | Likely Benign | Likely Benign | 0.166 | Likely Benign | -2.59 | Deleterious | 0.059 | Benign | 0.041 | Benign | 2.35 | Pathogenic | 0.00 | Affected | 0.0995 | 0.2468 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3845A>G | E1282G 2D AIThe SynGAP1 missense variant E1282G is reported in gnomAD (ID 6‑33447893‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Taken together, the overwhelming majority of predictions support a benign impact, and there is no ClinVar classification to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.817364 | Binding | 0.465 | 0.725 | 0.875 | 6-33447893-A-G | -2.649 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.219 | Likely Benign | -1.25 | Neutral | 0.000 | Benign | 0.001 | Benign | 2.68 | Benign | 0.00 | Affected | 0.2729 | 0.5284 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||
| c.3851T>G | L1284R 2D AIThe SynGAP1 missense variant L1284R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic impact. This assessment does not contradict ClinVar status, as the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.812494 | Disordered | 0.824557 | Binding | 0.441 | 0.748 | 0.875 | -4.203 | Likely Benign | 0.156 | Likely Benign | Likely Benign | 0.163 | Likely Benign | -3.11 | Deleterious | 0.990 | Probably Damaging | 0.722 | Possibly Damaging | 2.48 | Pathogenic | 0.00 | Affected | 0.1216 | 0.0488 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3857A>T | E1286V 2D AIThe SynGAP1 missense variant E1286V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑benign/2‑pathogenic split. High‑accuracy methods show AlphaMissense‑Optimized as benign; Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. four benign) indicate that the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.817022 | Binding | 0.544 | 0.765 | 0.750 | -4.195 | Likely Benign | 0.275 | Likely Benign | Likely Benign | 0.259 | Likely Benign | -3.90 | Deleterious | 0.960 | Probably Damaging | 0.679 | Possibly Damaging | 2.42 | Pathogenic | 0.00 | Affected | 0.0828 | 0.5614 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3860C>A | P1287H 2D AIThe SynGAP1 missense variant P1287H is recorded in gnomAD (6‑33447908‑C‑A) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, SGM‑Consensus is likely benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) is not available for this variant. Based on the aggregate predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.813701 | Binding | 0.538 | 0.777 | 0.750 | 6-33447908-C-A | -3.606 | Likely Benign | 0.135 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -1.36 | Neutral | 0.938 | Possibly Damaging | 0.596 | Possibly Damaging | 2.80 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1306 | 0.3448 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.3862A>C | K1288Q 2D AIThe SynGAP1 missense variant K1288Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a 2‑to‑2 split and is therefore inconclusive. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence is divided, with an equal number of benign and pathogenic calls, leaving the variant’s clinical significance uncertain. This uncertainty does not contradict ClinVar, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | -2.369 | Likely Benign | 0.172 | Likely Benign | Likely Benign | 0.173 | Likely Benign | -2.51 | Deleterious | 0.991 | Probably Damaging | 0.987 | Probably Damaging | 2.11 | Pathogenic | 0.00 | Affected | 0.4149 | 0.0657 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3862A>G | K1288E 2D AISynGAP1 missense variant K1288E is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33447910‑A‑G). Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and AlphaMissense‑Optimized, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this change. Overall, the preponderance of evidence points to a pathogenic effect, which is consistent with the ClinVar designation of uncertain significance rather than a clear benign classification. Thus, the variant is most likely pathogenic, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | Uncertain | 1 | 6-33447910-A-G | 5 | 3.22e-6 | -2.751 | Likely Benign | 0.407 | Ambiguous | Likely Benign | 0.185 | Likely Benign | -3.27 | Deleterious | 0.979 | Probably Damaging | 0.973 | Probably Damaging | 2.13 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3627 | 0.0676 | 1 | 0 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||
| c.3863A>C | K1288T 2D AIThe SynGAP1 missense variant K1288T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, more tools (five) predict pathogenicity than benign (three), and the high‑accuracy consensus also leans pathogenic. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | -2.616 | Likely Benign | 0.477 | Ambiguous | Likely Benign | 0.227 | Likely Benign | -4.84 | Deleterious | 0.991 | Probably Damaging | 0.982 | Probably Damaging | 2.09 | Pathogenic | 0.00 | Affected | 0.2066 | 0.2614 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||||||||||
| c.3863A>G | K1288R 2D AIThe SynGAP1 missense variant K1288R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | -2.367 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 0.141 | Likely Benign | -2.22 | Neutral | 0.979 | Probably Damaging | 0.973 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 0.4189 | 0.0782 | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.3863A>T | K1288M 2D AIThe SynGAP1 missense variant K1288M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus indicates a likely pathogenic outcome; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of computational predictions (seven pathogenic vs. three benign) point to a pathogenic impact for K1288M. This conclusion is not contradicted by ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | -3.355 | Likely Benign | 0.660 | Likely Pathogenic | Likely Benign | 0.246 | Likely Benign | -4.89 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.06 | Pathogenic | 0.00 | Affected | 0.0985 | 0.3140 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||||||||||||
| c.3864G>C | K1288N 2D AIThe SynGAP1 missense variant K1288N has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and ESM1b, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no result for this variant. High‑accuracy assessments therefore show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta as unavailable. Based on the overall distribution of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | -3.670 | Likely Benign | 0.801 | Likely Pathogenic | Ambiguous | 0.128 | Likely Benign | -4.05 | Deleterious | 0.991 | Probably Damaging | 0.987 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.3513 | 0.1101 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3864G>T | K1288N 2D AIThe SynGAP1 missense variant K1288N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and ESM1b, while the majority of tools predict a pathogenic effect: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports the variant as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy assessments therefore show an uncertain AlphaMissense‑Optimized prediction, a Likely Pathogenic SGM‑Consensus, and no Foldetta data. Overall, the preponderance of evidence points to a pathogenic impact, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.827927 | Disordered | 0.814714 | Binding | 0.538 | 0.784 | 0.625 | -3.670 | Likely Benign | 0.801 | Likely Pathogenic | Ambiguous | 0.128 | Likely Benign | -4.05 | Deleterious | 0.991 | Probably Damaging | 0.987 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.3513 | 0.1101 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.3868A>T | R1290W 2D AIThe SynGAP1 missense variant R1290W is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote) as benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the preponderance of evidence from multiple independent predictors points to a benign impact for R1290W, and this conclusion is consistent with the lack of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.844138 | Binding | 0.567 | 0.795 | 0.625 | -5.672 | Likely Benign | 0.294 | Likely Benign | Likely Benign | 0.163 | Likely Benign | -5.15 | Deleterious | 0.994 | Probably Damaging | 0.920 | Probably Damaging | 2.58 | Benign | 0.00 | Affected | 0.0992 | 0.2724 | 2 | -3 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3869G>T | R1290M 2D AIThe SynGAP1 missense variant R1290M is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of reliable predictions indicate a benign impact, and this conclusion does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.844138 | Binding | 0.567 | 0.795 | 0.625 | -4.661 | Likely Benign | 0.390 | Ambiguous | Likely Benign | 0.143 | Likely Benign | -3.47 | Deleterious | 0.977 | Probably Damaging | 0.796 | Possibly Damaging | 2.60 | Benign | 0.00 | Affected | 0.1059 | 0.2899 | 0 | -1 | 6.4 | -24.99 | ||||||||||||||||||||||||||||||||||||||||
| c.3872T>G | L1291R 2D AIThe SynGAP1 missense variant L1291R is not reported in ClinVar and is present in gnomAD (ID 6‑33447920‑T‑G). Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessment shows AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive and Foldetta results are unavailable. Overall, the majority of conventional predictors indicate a pathogenic effect, whereas the single high‑accuracy tool suggests benign. No ClinVar annotation contradicts these findings. Thus, based on the collective evidence, the variant is most likely pathogenic, though the high‑accuracy prediction introduces uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.827927 | Disordered | 0.863458 | Binding | 0.579 | 0.797 | 0.625 | 6-33447920-T-G | 1 | 6.44e-7 | -3.977 | Likely Benign | 0.302 | Likely Benign | Likely Benign | 0.287 | Likely Benign | -4.29 | Deleterious | 0.990 | Probably Damaging | 0.856 | Possibly Damaging | 2.49 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1160 | 0.1049 | -2 | -3 | -8.3 | 43.03 | |||||||||||||||||||||||||||||||||||
| c.3875T>A | L1292H 2D AIThe SynGAP1 missense variant L1292H is not reported in ClinVar (ClinVar status: None) but is present in gnomAD (ID 6‑33447923‑T‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the predictions are split, with a slight tilt toward pathogenicity (five pathogenic vs. four benign). Thus, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.882643 | Binding | 0.586 | 0.800 | 0.625 | 6-33447923-T-A | -5.692 | Likely Benign | 0.335 | Likely Benign | Likely Benign | 0.202 | Likely Benign | -4.42 | Deleterious | 0.992 | Probably Damaging | 0.820 | Possibly Damaging | 2.46 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1271 | 0.0649 | -3 | -2 | -7.0 | 23.98 | |||||||||||||||||||||||||||||||||||||
| c.3875T>G | L1292R 2D AIThe SynGAP1 missense variant L1292R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, five tools predict pathogenicity while four predict benignity, giving a slight tilt toward a pathogenic interpretation. This prediction does not contradict ClinVar status, as the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.882643 | Binding | 0.586 | 0.800 | 0.625 | -4.249 | Likely Benign | 0.238 | Likely Benign | Likely Benign | 0.223 | Likely Benign | -4.34 | Deleterious | 0.971 | Probably Damaging | 0.773 | Possibly Damaging | 2.46 | Pathogenic | 0.00 | Affected | 0.1286 | 0.0849 | -3 | -2 | -8.3 | 43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3877G>A | D1293N 2D AIThe SynGAP1 missense variant D1293N is reported in gnomAD (6‑33447925‑G‑A) but has no ClinVar entry. Functional prediction tools split in a 5‑to‑4 ratio: benign calls come from REVEL, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic calls come from PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | 6-33447925-G-A | 5 | 3.22e-6 | -3.983 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.175 | Likely Benign | -3.30 | Deleterious | 0.950 | Possibly Damaging | 0.414 | Benign | 2.33 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1264 | 0.3421 | 1 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||
| c.3877G>C | D1293H 2D AIThe SynGAP1 missense variant D1293H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. three benign) and the SGM Consensus support a pathogenic classification, whereas AlphaMissense‑Optimized suggests benign. The variant is most likely pathogenic based on the collective evidence, and this conclusion does not contradict any ClinVar status because the variant is not yet reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | -4.422 | Likely Benign | 0.391 | Ambiguous | Likely Benign | 0.278 | Likely Benign | -4.59 | Deleterious | 0.984 | Probably Damaging | 0.888 | Possibly Damaging | 2.17 | Pathogenic | 0.00 | Affected | 0.1609 | 0.3731 | 1 | -1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||||||||||
| c.3877G>T | D1293Y 2D AIThe SynGAP1 missense variant D1293Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. three benign) and the SGM Consensus support a pathogenic classification, with no ClinVar entry to contradict this assessment. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | -5.452 | Likely Benign | 0.392 | Ambiguous | Likely Benign | 0.310 | Likely Benign | -6.15 | Deleterious | 0.995 | Probably Damaging | 0.897 | Possibly Damaging | 2.15 | Pathogenic | 0.00 | Affected | 0.0627 | 0.3577 | -4 | -3 | 2.2 | 48.09 | ||||||||||||||||||||||||||||||||||||||||
| c.3878A>C | D1293A 2D AIThe SynGAP1 missense variant D1293A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the balance of evidence (5 benign vs 4 pathogenic predictions) leans toward a benign classification. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | -2.251 | Likely Benign | 0.197 | Likely Benign | Likely Benign | 0.302 | Likely Benign | -5.19 | Deleterious | 0.770 | Possibly Damaging | 0.255 | Benign | 2.20 | Pathogenic | 0.00 | Affected | 0.3267 | 0.3447 | 0 | -2 | 5.3 | -44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.3878A>G | D1293G 2D AIThe SynGAP1 missense variant D1293G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the balance of evidence (5 benign vs 4 pathogenic predictions, with the most accurate tool indicating benign) suggests the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | -3.060 | Likely Benign | 0.261 | Likely Benign | Likely Benign | 0.332 | Likely Benign | -4.91 | Deleterious | 0.872 | Possibly Damaging | 0.399 | Benign | 2.19 | Pathogenic | 0.00 | Affected | 0.3022 | 0.3944 | 1 | -1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3878A>T | D1293V 2D AIThe SynGAP1 missense variant D1293V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of available predictions (five pathogenic vs. four benign) lean toward a pathogenic impact. This assessment does not contradict ClinVar status, as the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | -3.817 | Likely Benign | 0.328 | Likely Benign | Likely Benign | 0.343 | Likely Benign | -6.03 | Deleterious | 0.960 | Probably Damaging | 0.679 | Possibly Damaging | 2.16 | Pathogenic | 0.00 | Affected | 0.0896 | 0.3603 | -2 | -3 | 7.7 | -15.96 | ||||||||||||||||||||||||||||||||||||||||
| c.3879C>A | D1293E 2D AIThe SynGAP1 missense variant D1293E is reported in gnomAD (variant ID 6‑33447927‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, SIFT and FATHMM predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments further support benignity: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | 6-33447927-C-A | -2.627 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.042 | Likely Benign | -1.86 | Neutral | 0.011 | Benign | 0.008 | Benign | 2.39 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1512 | 0.3425 | 2 | 3 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||
| c.3879C>G | D1293E 2D AIThe SynGAP1 missense variant D1293E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for D1293E, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.779859 | Disordered | 0.892346 | Binding | 0.569 | 0.801 | 0.625 | -2.627 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.86 | Neutral | 0.011 | Benign | 0.008 | Benign | 2.39 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1512 | 0.3425 | 2 | 3 | 0.0 | 14.03 | |||||||||||||||||||||||||||||||||||||
| c.3880G>A | A1294T 2D AIThe SynGAP1 missense variant A1294T is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447928‑G‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy predictions show AlphaMissense‑Optimized as benign, while SGM Consensus and Foldetta remain unavailable. Overall, five tools predict pathogenicity versus four predicting benign, and the lack of ClinVar evidence does not contradict these findings. Thus, the variant is most likely pathogenic based on the current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.895011 | Binding | 0.565 | 0.806 | 0.625 | 6-33447928-G-A | -3.966 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.290 | Likely Benign | -3.04 | Deleterious | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 2.18 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1244 | 0.5479 | 0 | 1 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||
| c.3880G>C | A1294P 2D AIThe SynGAP1 missense variant A1294P is catalogued in gnomAD (6‑33447928‑G‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—yields a 2‑to‑2 split and is therefore inconclusive; Foldetta results are not available. Overall, the balance of evidence leans toward a pathogenic effect, though the single high‑accuracy benign call and the inconclusive consensus temper certainty. This assessment does not conflict with ClinVar, which currently contains no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.895011 | Binding | 0.565 | 0.806 | 0.625 | 6-33447928-G-C | 1 | 6.45e-7 | -2.688 | Likely Benign | 0.220 | Likely Benign | Likely Benign | 0.367 | Likely Benign | -3.79 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1682 | 0.3207 | -1 | 1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||
| c.3880G>T | A1294S 2D AIThe SynGAP1 missense variant A1294S is reported in ClinVar as not yet classified (no ClinVar ID) and is present in gnomAD (variant ID 6-33447928‑G‑T). Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates benign. No Foldetta stability analysis is available. Overall, the majority of evidence points to a benign effect for A1294S, and this conclusion does not contradict the current ClinVar status, which remains unclassified. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.895011 | Binding | 0.565 | 0.806 | 0.625 | 6-33447928-G-T | -3.229 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.276 | Likely Benign | -2.21 | Neutral | 0.997 | Probably Damaging | 0.991 | Probably Damaging | 2.19 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2302 | 0.3990 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||||||||
| c.3881C>A | A1294D 2D AIThe SynGAP1 missense variant A1294D is listed in gnomAD (ID 6‑33447929‑C‑A) but has no ClinVar entry. Functional prediction tools show mixed results: benign calls come from REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessment further clarifies the picture: AlphaMissense‑Optimized predicts a benign effect, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans toward pathogenicity. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a pathogenic effect, which is consistent with the lack of a benign ClinVar classification and the presence of the variant in a general population database. Thus, the variant is most likely pathogenic, and this prediction does not contradict any ClinVar status because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.895011 | Binding | 0.565 | 0.806 | 0.625 | 6-33447929-C-A | -4.179 | Likely Benign | 0.369 | Ambiguous | Likely Benign | 0.224 | Likely Benign | -4.17 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1893 | 0.2262 | -2 | 0 | -5.3 | 44.01 | |||||||||||||||||||||||||||||||||||||
| c.3881C>G | A1294G 2D AIThe SynGAP1 missense variant A1294G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy predictors (AlphaMissense‑Optimized) indicate a benign outcome, while the consensus of other tools is split. Thus, the variant is most likely benign based on current predictions, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.895011 | Binding | 0.565 | 0.806 | 0.625 | -3.242 | Likely Benign | 0.132 | Likely Benign | Likely Benign | 0.201 | Likely Benign | -3.06 | Deleterious | 0.992 | Probably Damaging | 0.983 | Probably Damaging | 2.15 | Pathogenic | 0.00 | Affected | 0.1980 | 0.2695 | 1 | 0 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3881C>T | A1294V 2D AIThe SynGAP1 missense variant A1294V is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447929‑C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑benign/2‑pathogenic split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools specifically show AlphaMissense‑Optimized as benign, SGM Consensus as inconclusive, and Foldetta as unavailable. Overall, the predictions are mixed, with one more pathogenic than benign calls. Thus, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.895011 | Binding | 0.565 | 0.806 | 0.625 | 6-33447929-C-T | 1 | 6.45e-7 | -3.842 | Likely Benign | 0.105 | Likely Benign | Likely Benign | 0.164 | Likely Benign | -3.16 | Deleterious | 0.997 | Probably Damaging | 0.983 | Probably Damaging | 2.22 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1033 | 0.4413 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||||||||
| c.3883C>A | Q1295K 2D AIThe SynGAP1 missense variant Q1295K is not reported in ClinVar (ClinVar status: not listed) but is present in gnomAD (gnomAD ID: 6‑33447931‑C‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. AlphaMissense‑Default is uncertain. The high‑accuracy consensus (SGM Consensus) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a pathogenic prediction (2 pathogenic vs. 1 benign vs. 1 uncertain). AlphaMissense‑Optimized remains benign, while Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. three benign) point to a pathogenic effect. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | 6-33447931-C-A | -3.419 | Likely Benign | 0.343 | Ambiguous | Likely Benign | 0.313 | Likely Benign | -3.30 | Deleterious | 0.843 | Possibly Damaging | 0.893 | Possibly Damaging | 2.38 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1782 | 0.3517 | 1 | 1 | -0.4 | 0.04 | |||||||||||||||||||||||||||||||||||||
| c.3883C>G | Q1295E 2D AIThe SynGAP1 missense variant Q1295E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | -3.192 | Likely Benign | 0.177 | Likely Benign | Likely Benign | 0.264 | Likely Benign | -2.46 | Neutral | 0.843 | Possibly Damaging | 0.848 | Possibly Damaging | 2.29 | Pathogenic | 0.00 | Affected | 0.1428 | 0.1492 | 2 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3884A>C | Q1295P 2D AIThe SynGAP1 missense variant Q1295P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of predictions (5/9) indicate pathogenicity, while 4/9 suggest benignity. Thus, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | -2.511 | Likely Benign | 0.168 | Likely Benign | Likely Benign | 0.372 | Likely Benign | -4.95 | Deleterious | 0.968 | Probably Damaging | 0.954 | Probably Damaging | 2.23 | Pathogenic | 0.00 | Affected | 0.2299 | 0.4738 | 0 | -1 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||||||||
| c.3884A>G | Q1295R 2D AIThe SynGAP1 missense variant Q1295R is catalogued in gnomAD (ID 6‑33447932‑A‑G) but has no ClinVar entry. Functional prediction tools show discordant results: benign calls come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments are mixed: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta data are unavailable. Overall, the balance of evidence slightly favors a pathogenic interpretation, though the presence of several benign predictions and the lack of a ClinVar classification mean the variant’s clinical significance remains uncertain. No contradiction exists with ClinVar status, as no ClinVar claim is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | 6-33447932-A-G | -3.342 | Likely Benign | 0.300 | Likely Benign | Likely Benign | 0.351 | Likely Benign | -3.30 | Deleterious | 0.925 | Possibly Damaging | 0.932 | Probably Damaging | 2.44 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1432 | 0.1348 | 1 | 1 | -1.0 | 28.06 | |||||||||||||||||||||||||||||||||||||
| c.3884A>T | Q1295L 2D AIThe SynGAP1 missense variant Q1295L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact for Q1295L, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | -2.862 | Likely Benign | 0.348 | Ambiguous | Likely Benign | 0.379 | Likely Benign | -5.63 | Deleterious | 0.925 | Possibly Damaging | 0.932 | Probably Damaging | 2.25 | Pathogenic | 0.00 | Affected | 0.0810 | 0.5672 | -2 | -2 | 7.3 | -14.97 | ||||||||||||||||||||||||||||||||||||||||
| c.3885G>C | Q1295H 2D AIThe SynGAP1 missense variant Q1295H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show discordant results: benign calls come from REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further highlight this split: AlphaMissense‑Optimized predicts a benign effect, whereas the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome. No Foldetta stability analysis is available for this residue. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict any existing ClinVar annotation, as none is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | -4.851 | Likely Benign | 0.588 | Likely Pathogenic | Likely Benign | 0.349 | Likely Benign | -4.08 | Deleterious | 0.991 | Probably Damaging | 0.986 | Probably Damaging | 2.24 | Pathogenic | 0.00 | Affected | 0.1345 | 0.3203 | 3 | 0 | 0.3 | 9.01 | |||||||||||||||||||||||||||||||||||||||
| c.3885G>T | Q1295H 2D AIThe SynGAP1 missense variant Q1295H is catalogued in gnomAD (ID 6‑33447933‑G‑T) but has no ClinVar submission. Functional prediction tools show a split: benign calls come from REVEL, ESM1b, and AlphaMissense‑Optimized, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. When predictions are grouped, 3 tools predict benign and 6 predict pathogenic. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized remains benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this substitution. Overall, the preponderance of evidence points to a pathogenic effect, and this assessment does not conflict with ClinVar, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.852992 | Disordered | 0.892719 | Binding | 0.499 | 0.801 | 0.625 | 6-33447933-G-T | -4.851 | Likely Benign | 0.588 | Likely Pathogenic | Likely Benign | 0.349 | Likely Benign | -4.08 | Deleterious | 0.991 | Probably Damaging | 0.986 | Probably Damaging | 2.24 | Pathogenic | 0.00 | Affected | 0.1345 | 0.3203 | 3 | 0 | 0.3 | 9.01 | ||||||||||||||||||||||||||||||||||||||
| c.3896T>A | L1299H 2D AIThe SynGAP1 missense variant L1299H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.771762 | Disordered | 0.896323 | Binding | 0.398 | 0.832 | 0.750 | -4.586 | Likely Benign | 0.300 | Likely Benign | Likely Benign | 0.317 | Likely Benign | -3.83 | Deleterious | 0.992 | Probably Damaging | 0.750 | Possibly Damaging | 2.67 | Benign | 0.00 | Affected | 0.1293 | 0.1273 | -2 | -3 | -7.0 | 23.98 | |||||||||||||||||||||||||||||||||||||||
| c.38T>A | I13N 2D AIThe SynGAP1 missense variant I13N is reported in gnomAD (ID 6‑33420302‑T‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labels it “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign; Foldetta results are not available. Overall, the consensus of the majority of tools, including the high‑accuracy methods, indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | 6-33420302-T-A | -3.725 | Likely Benign | 0.237 | Likely Benign | Likely Benign | 0.096 | Likely Benign | -0.16 | Neutral | 0.056 | Benign | 0.005 | Benign | 4.02 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1220 | 0.1100 | -3 | -2 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||
| c.38T>C | I13T 2D AIThe SynGAP1 missense variant I13T is listed in gnomAD (ID 6‑33420302‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign and the SGM‑Consensus is “Likely Benign.” No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | 6-33420302-T-C | -2.856 | Likely Benign | 0.385 | Ambiguous | Likely Benign | 0.132 | Likely Benign | -0.02 | Neutral | 0.024 | Benign | 0.003 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1401 | 0.1778 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||||
| c.38T>G | I13S 2D AIThe SynGAP1 missense variant I13S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus methods also support a benign classification: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for I13S, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | -1.756 | Likely Benign | 0.187 | Likely Benign | Likely Benign | 0.264 | Likely Benign | 0.20 | Neutral | 0.024 | Benign | 0.002 | Benign | 4.06 | Benign | 0.00 | Affected | 0.3292 | 0.1682 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3904T>A | L1302M 2D AIThe SynGAP1 missense variant L1302M is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; Foldetta results are unavailable. Overall, the majority of predictions (six benign vs. four pathogenic) support a benign classification. There is no ClinVar entry to contradict this assessment, so the variant is most likely benign based on the available computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | -5.246 | Likely Benign | 0.131 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -1.34 | Neutral | 0.960 | Probably Damaging | 0.799 | Possibly Damaging | 1.54 | Pathogenic | 0.00 | Affected | 0.0929 | 0.3439 | 4 | 2 | -1.9 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3904T>G | L1302V 2D AIThe SynGAP1 missense variant L1302V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, AlphaMissense‑Optimized, and ESM1b. Tools that predict a pathogenic effect are SIFT and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | -4.140 | Likely Benign | 0.082 | Likely Benign | Likely Benign | 0.214 | Likely Benign | -1.71 | Neutral | 0.004 | Benign | 0.022 | Benign | 1.58 | Pathogenic | 0.00 | Affected | 0.1603 | 0.2686 | 2 | 1 | 0.4 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3905T>C | L1302S 2D 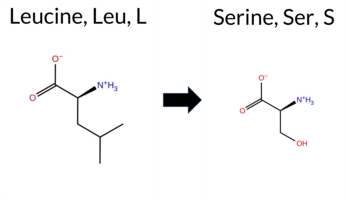 AIThe SynGAP1 missense variant L1302S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools specifically show AlphaMissense‑Optimized as benign, while SGM Consensus and Foldetta remain unavailable. Overall, the majority of predictions (five pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | -3.411 | Likely Benign | 0.196 | Likely Benign | Likely Benign | 0.307 | Likely Benign | -4.17 | Deleterious | 0.960 | Probably Damaging | 0.726 | Possibly Damaging | 1.50 | Pathogenic | 0.00 | Affected | 0.3237 | 0.0982 | -3 | -2 | -4.6 | -26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3905T>G | L1302W 2D 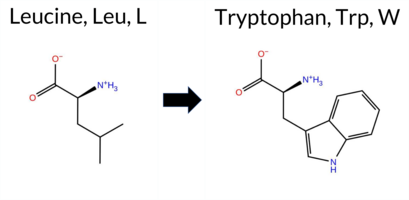 AIThe SynGAP1 missense variant L1302W is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM; ESM1b is uncertain and not counted. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. three benign) lean toward pathogenicity, and this is consistent with the SGM Consensus result. Therefore, the variant is most likely pathogenic based on current computational evidence, and this does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | -7.133 | In-Between | 0.297 | Likely Benign | Likely Benign | 0.290 | Likely Benign | -4.20 | Deleterious | 0.996 | Probably Damaging | 0.946 | Probably Damaging | 1.49 | Pathogenic | 0.00 | Affected | 0.0671 | 0.2530 | -2 | -2 | -4.7 | 73.05 | ||||||||||||||||||||||||||||||||||||||||
| c.3906G>C | L1302F 2D AISynGAP1 missense variant L1302F is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments indicate AlphaMissense‑Optimized predicts benign, whereas the SGM consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta stability analysis is unavailable. Overall, the majority of predictions (five pathogenic versus four benign) lean toward a pathogenic effect. Thus, the variant is most likely pathogenic, a conclusion that contrasts with its current ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | Uncertain | 1 | -5.674 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.211 | Likely Benign | -2.70 | Deleterious | 0.960 | Probably Damaging | 0.657 | Possibly Damaging | 1.53 | Pathogenic | 0.00 | Affected | 0.0721 | 0.2648 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3906G>T | L1302F 2D AIThe SynGAP1 missense variant L1302F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools that are available indicate a benign outcome (AlphaMissense‑Optimized) while the consensus and stability analyses are missing. Consequently, the overall prediction is ambiguous, but the most reliable evidence points toward a benign effect, and this does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.889642 | Binding | 0.429 | 0.842 | 0.875 | -5.674 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.215 | Likely Benign | -2.70 | Deleterious | 0.960 | Probably Damaging | 0.657 | Possibly Damaging | 1.53 | Pathogenic | 0.00 | Affected | 0.0721 | 0.2648 | 2 | 0 | -1.0 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.3916A>C | N1306H 2D 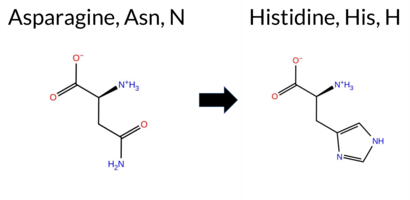 AIThe SynGAP1 missense variant N1306H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is labeled “Likely Benign”). In contrast, tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for this variant, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -2.949 | Likely Benign | 0.159 | Likely Benign | Likely Benign | 0.240 | Likely Benign | -3.42 | Deleterious | 0.961 | Probably Damaging | 0.825 | Possibly Damaging | 2.54 | Benign | 0.00 | Affected | 0.1705 | 0.7846 | 2 | 1 | 0.3 | 23.04 | |||||||||||||||||||||||||||||||||||||||
| c.3916A>G | N1306D 2D  AIThe SynGAP1 missense variant N1306D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -2.089 | Likely Benign | 0.207 | Likely Benign | Likely Benign | 0.143 | Likely Benign | -3.43 | Deleterious | 0.001 | Benign | 0.010 | Benign | 2.58 | Benign | 0.00 | Affected | 0.2018 | 0.4992 | 2 | 1 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3916A>T | N1306Y 2D  AIThe SynGAP1 missense variant N1306Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote) as benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the preponderance of evidence points to a benign impact for the variant, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -4.193 | Likely Benign | 0.298 | Likely Benign | Likely Benign | 0.273 | Likely Benign | -5.59 | Deleterious | 0.961 | Probably Damaging | 0.825 | Possibly Damaging | 2.54 | Benign | 0.00 | Affected | 0.0732 | 0.6658 | -2 | -2 | 2.2 | 49.07 | |||||||||||||||||||||||||||||||||||||||
| c.3917A>C | N1306T 2D  AIThe SynGAP1 missense variant N1306T is reported in gnomAD (ID 6‑33451791‑A‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, while pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | 6-33451791-A-C | -1.863 | Likely Benign | 0.156 | Likely Benign | Likely Benign | 0.240 | Likely Benign | -4.17 | Deleterious | 0.697 | Possibly Damaging | 0.399 | Benign | 2.59 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1588 | 0.8327 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||||||||
| c.3917A>G | N1306S 2D  AIThe SynGAP1 missense variant N1306S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, while Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -1.344 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.233 | Likely Benign | -3.49 | Deleterious | 0.319 | Benign | 0.242 | Benign | 2.69 | Benign | 0.00 | Affected | 0.4120 | 0.7826 | 1 | 1 | 2.7 | -27.03 | |||||||||||||||||||||||||||||||||||||||
| c.3917A>T | N1306I 2D AIThe SynGAP1 missense variant N1306I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic predictions come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -3.234 | Likely Benign | 0.296 | Likely Benign | Likely Benign | 0.188 | Likely Benign | -6.15 | Deleterious | 0.890 | Possibly Damaging | 0.761 | Possibly Damaging | 2.56 | Benign | 0.00 | Affected | 0.0837 | 0.6672 | -2 | -3 | 8.0 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3918C>A | N1306K 2D  AIThe SynGAP1 missense variant N1306K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -3.021 | Likely Benign | 0.585 | Likely Pathogenic | Likely Benign | 0.140 | Likely Benign | -4.00 | Deleterious | 0.532 | Possibly Damaging | 0.327 | Benign | 2.61 | Benign | 0.00 | Affected | 0.2375 | 0.6234 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.3918C>G | N1306K 2D AIThe SynGAP1 missense variant N1306K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.795062 | Disordered | 0.902190 | Binding | 0.367 | 0.888 | 0.875 | -3.021 | Likely Benign | 0.585 | Likely Pathogenic | Likely Benign | 0.177 | Likely Benign | -4.00 | Deleterious | 0.532 | Possibly Damaging | 0.327 | Benign | 2.61 | Benign | 0.00 | Affected | 0.2375 | 0.6234 | 1 | 0 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||||
| c.391G>C | G131R 2D  AIThe SynGAP1 missense variant G131R is listed in ClinVar with an “Uncertain” significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive due to a 2‑to‑2 split and therefore does not contribute evidence. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence tools predict a pathogenic effect, and the variant is most likely pathogenic based on current predictions, which does not contradict the ClinVar “Uncertain” status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.724779 | Binding | 0.302 | 0.891 | 0.250 | Uncertain | 1 | -6.564 | Likely Benign | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.099 | Likely Benign | -3.82 | Deleterious | 0.983 | Probably Damaging | 0.656 | Possibly Damaging | 3.92 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1070 | 0.4667 | -2 | -3 | -4.1 | 99.14 | ||||||||||||||||||||||||||||||||||||
| c.391G>T | G131C 2D  AIThe SynGAP1 missense variant G131C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM Consensus again indicates Likely Pathogenic; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of tools predict pathogenicity, and this aligns with the SGM Consensus. Therefore, the variant is most likely pathogenic based on the available predictions, and this conclusion is consistent with the lack of ClinVar reporting rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.724779 | Binding | 0.302 | 0.891 | 0.250 | -8.288 | Likely Pathogenic | 0.727 | Likely Pathogenic | Likely Benign | 0.155 | Likely Benign | -4.42 | Deleterious | 0.998 | Probably Damaging | 0.840 | Possibly Damaging | 3.88 | Benign | 0.00 | Affected | 0.1339 | 0.4356 | -3 | -3 | 2.9 | 46.09 | |||||||||||||||||||||||||||||||||||||||
| c.3922C>A | R1308S 2D AIThe SynGAP1 missense variant R1308S is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The high‑accuracy AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evaluated tools (five pathogenic vs. three benign) indicate a pathogenic effect. This prediction is not contradicted by ClinVar status, which has no entry for the variant. Thus, based on current computational evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.741537 | Disordered | 0.930652 | Binding | 0.378 | 0.904 | 0.750 | -2.180 | Likely Benign | 0.419 | Ambiguous | Likely Benign | 0.307 | Likely Benign | -3.76 | Deleterious | 0.982 | Probably Damaging | 0.982 | Probably Damaging | 2.36 | Pathogenic | 0.00 | Affected | 0.3070 | 0.4744 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.3922C>G | R1308G 2D AIThe SynGAP1 missense variant R1308G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign vs. two pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic interpretation. This assessment does not contradict any ClinVar status, as the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.741537 | Disordered | 0.930652 | Binding | 0.378 | 0.904 | 0.750 | -2.503 | Likely Benign | 0.180 | Likely Benign | Likely Benign | 0.300 | Likely Benign | -4.34 | Deleterious | 0.982 | Probably Damaging | 0.982 | Probably Damaging | 2.33 | Pathogenic | 0.00 | Affected | 0.3528 | 0.3830 | -3 | -2 | 4.1 | -99.14 | ||||||||||||||||||||||||||||||||||||||||
| c.3922C>T | R1308C 2D AIThe SynGAP1 missense variant R1308C is listed in ClinVar with an “Uncertain” significance and is present in the gnomAD database (ID 6‑33451796‑C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. The high‑accuracy consensus (SGM Consensus) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a pathogenic verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact, and this assessment does not contradict the current ClinVar “Uncertain” status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.741537 | Disordered | 0.930652 | Binding | 0.378 | 0.904 | 0.750 | Conflicting | 2 | 6-33451796-C-T | 4 | 2.48e-6 | -4.994 | Likely Benign | 0.421 | Ambiguous | Likely Benign | 0.352 | Likely Benign | -4.89 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 2.31 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.3139 | 0.4274 | -4 | -3 | 7.0 | -53.05 | |||||||||||||||||||||||||||||||||
| c.3923G>A | R1308H 2D  AISynGAP1 missense variant R1308H is listed in ClinVar as benign and is present in gnomAD (ID 6‑33451797‑G‑A). Functional prediction tools show mixed results: benign calls come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments give a benign result from AlphaMissense‑Optimized; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic), and Foldetta stability analysis is unavailable. Overall, the majority of standard predictors favor a pathogenic effect, whereas the most accurate single‑tool prediction is benign. Thus, the variant is most likely pathogenic according to the aggregate predictions, which contradicts the benign classification in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.741537 | Disordered | 0.930652 | Binding | 0.378 | 0.904 | 0.750 | Likely Benign | 1 | 6-33451797-G-A | 3 | 1.86e-6 | -3.586 | Likely Benign | 0.201 | Likely Benign | Likely Benign | 0.319 | Likely Benign | -3.12 | Deleterious | 0.998 | Probably Damaging | 0.991 | Probably Damaging | 2.33 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2955 | 0.2515 | 2 | 0 | 1.3 | -19.05 | |||||||||||||||||||||||||||||||||
| c.3923G>C | R1308P 2D AIThe SynGAP1 missense variant R1308P is not reported in ClinVar but is present in gnomAD (ID 6‑33451797‑G‑C). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑vs‑2 split and therefore inconclusive. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Because the available predictions are evenly divided between benign and pathogenic, the overall computational assessment is inconclusive; the variant is neither clearly benign nor clearly pathogenic. This lack of consensus does not contradict ClinVar, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.741537 | Disordered | 0.930652 | Binding | 0.378 | 0.904 | 0.750 | 6-33451797-G-C | -1.320 | Likely Benign | 0.216 | Likely Benign | Likely Benign | 0.158 | Likely Benign | -4.62 | Deleterious | 0.995 | Probably Damaging | 0.992 | Probably Damaging | 2.32 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2070 | 0.5020 | -2 | 0 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||
| c.3923G>T | R1308L 2D AIThe SynGAP1 missense variant R1308L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs. 2 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic impact. This assessment does not contradict ClinVar status, as the variant has not yet been classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.741537 | Disordered | 0.930652 | Binding | 0.378 | 0.904 | 0.750 | -1.838 | Likely Benign | 0.257 | Likely Benign | Likely Benign | 0.360 | Likely Benign | -4.04 | Deleterious | 0.982 | Probably Damaging | 0.982 | Probably Damaging | 2.39 | Pathogenic | 0.00 | Affected | 0.1751 | 0.4930 | -3 | -2 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3926T>A | V1309E 2D AIThe SynGAP1 missense variant V1309E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.712013 | Disordered | 0.948596 | Binding | 0.402 | 0.907 | 0.750 | -3.393 | Likely Benign | 0.333 | Likely Benign | Likely Benign | 0.294 | Likely Benign | -2.40 | Neutral | 0.767 | Possibly Damaging | 0.473 | Possibly Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.1098 | 0.1727 | -2 | -2 | -7.7 | 29.98 | |||||||||||||||||||||||||||||||||||||||
| c.3926T>G | V1309G 2D AIThe SynGAP1 missense variant V1309G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact, and this conclusion does not contradict any ClinVar annotation because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.712013 | Disordered | 0.948596 | Binding | 0.402 | 0.907 | 0.750 | -2.618 | Likely Benign | 0.160 | Likely Benign | Likely Benign | 0.326 | Likely Benign | -3.40 | Deleterious | 0.391 | Benign | 0.767 | Possibly Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.1901 | 0.2144 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.392G>T | G131V 2D  AIThe SynGAP1 missense variant G131V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a tie and is therefore unavailable, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no result for this variant. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any existing ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.724779 | Binding | 0.302 | 0.891 | 0.250 | -6.248 | Likely Benign | 0.795 | Likely Pathogenic | Ambiguous | 0.199 | Likely Benign | -4.25 | Deleterious | 0.983 | Probably Damaging | 0.559 | Possibly Damaging | 3.92 | Benign | 0.00 | Affected | 0.1168 | 0.4175 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3934G>A | A1312T 2D AISynGAP1 missense variant A1312T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.767246 | Disordered | 0.971112 | Binding | 0.392 | 0.902 | 0.750 | -4.137 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.155 | Likely Benign | -0.65 | Neutral | 0.991 | Probably Damaging | 0.989 | Probably Damaging | 3.17 | Benign | 0.00 | Affected | 0.1865 | 0.7600 | 1 | 0 | -2.5 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.3934G>C | A1312P 2D AIThe SynGAP1 missense variant A1312P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.767246 | Disordered | 0.971112 | Binding | 0.392 | 0.902 | 0.750 | -2.488 | Likely Benign | 0.076 | Likely Benign | Likely Benign | 0.222 | Likely Benign | -1.64 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 3.12 | Benign | 0.00 | Affected | 0.2208 | 0.5820 | 1 | -1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3934G>T | A1312S 2D AIThe SynGAP1 missense variant A1312S is reported in gnomAD (variant ID 6‑33451808‑G‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: six methods (REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) and the SGM‑Consensus score all predict a benign effect, while three tools (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT) predict pathogenicity. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns “Benign” and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” No Foldetta stability data are available, so it does not influence the conclusion. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar status (none is listed). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.767246 | Disordered | 0.971112 | Binding | 0.392 | 0.902 | 0.750 | 6-33451808-G-T | 1 | 6.21e-7 | -3.906 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.123 | Likely Benign | -1.03 | Neutral | 0.978 | Probably Damaging | 0.983 | Probably Damaging | 3.21 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2850 | 0.6211 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||||||
| c.3935C>A | A1312D 2D AIThe SynGAP1 missense variant A1312D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors a benign outcome (2 benign vs. 1 pathogenic, with one uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence, particularly from the high‑accuracy tools, indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.767246 | Disordered | 0.971112 | Binding | 0.392 | 0.902 | 0.750 | -4.419 | Likely Benign | 0.398 | Ambiguous | Likely Benign | 0.272 | Likely Benign | -2.74 | Deleterious | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 3.11 | Benign | 0.00 | Affected | 0.2289 | 0.2993 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||||||||||||
| c.3935C>G | A1312G 2D AIThe SynGAP1 missense variant A1312G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to the variant being most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.767246 | Disordered | 0.971112 | Binding | 0.392 | 0.902 | 0.750 | -3.735 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.157 | Likely Benign | -2.25 | Neutral | 0.978 | Probably Damaging | 0.983 | Probably Damaging | 3.15 | Benign | 0.00 | Affected | 0.2241 | 0.5058 | 1 | 0 | -2.2 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3935C>T | A1312V 2D AIThe SynGAP1 missense variant A1312V is reported in gnomAD (ID 6‑33451809‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: six methods (REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) and the SGM‑Consensus score all predict a benign effect, while three tools (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT) predict pathogenicity. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns “Benign” and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” No Foldetta stability data are available, so it does not influence the conclusion. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar status (none is listed). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.767246 | Disordered | 0.971112 | Binding | 0.392 | 0.902 | 0.750 | 6-33451809-C-T | 4 | 2.48e-6 | -4.585 | Likely Benign | 0.109 | Likely Benign | Likely Benign | 0.231 | Likely Benign | -1.74 | Neutral | 0.991 | Probably Damaging | 0.983 | Probably Damaging | 3.15 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1495 | 0.6343 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||
| c.3946A>C | N1316H 2D AIThe SynGAP1 missense variant N1316H is reported in gnomAD (ID 6‑33451820‑A‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status, as none is currently assigned to the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451820-A-C | -4.151 | Likely Benign | 0.247 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -1.87 | Neutral | 0.939 | Possibly Damaging | 0.496 | Possibly Damaging | 3.95 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1535 | 0.6636 | 1 | 2 | 0.3 | 23.04 | ||||||||||||||||||||||||||||||||||||
| c.3946A>G | N1316D 2D AIThe SynGAP1 missense variant N1316D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451820‑A‑G). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also leans toward benign, with two benign and one pathogenic prediction among the four considered. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence indicates a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451820-A-G | -2.717 | Likely Benign | 0.364 | Ambiguous | Likely Benign | 0.057 | Likely Benign | -2.80 | Deleterious | 0.225 | Benign | 0.084 | Benign | 3.96 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2095 | 0.3745 | 1 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||
| c.3946A>T | N1316Y 2D AIThe SynGAP1 missense variant N1316Y is reported in gnomAD (variant ID 6‑33451820‑A‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, polyPhen2_HumVar, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Pathogenic predictions come from polyPhen2_HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is available). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451820-A-T | -5.579 | Likely Benign | 0.392 | Ambiguous | Likely Benign | 0.101 | Likely Benign | -2.48 | Neutral | 0.939 | Possibly Damaging | 0.396 | Benign | 3.90 | Benign | 0.00 | Affected | 3.77 | 5 | 0.0671 | 0.5365 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||||||||
| c.3947A>C | N1316T 2D AIThe SynGAP1 missense variant N1316T is reported in gnomAD (ID 6‑33451821‑A‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. When predictions are grouped by consensus, the benign‑predicating tools outnumber the pathogenic ones. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451821-A-C | -3.080 | Likely Benign | 0.315 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -3.18 | Deleterious | 0.127 | Benign | 0.045 | Benign | 3.96 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1450 | 0.6621 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||||||||
| c.3947A>G | N1316S 2D AIThe SynGAP1 missense variant N1316S is reported in gnomAD (ID 6‑33451821‑A‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. No Foldetta stability data are available, so it does not influence the assessment. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451821-A-G | 1 | 6.25e-7 | -2.906 | Likely Benign | 0.169 | Likely Benign | Likely Benign | 0.084 | Likely Benign | -2.69 | Deleterious | 0.004 | Benign | 0.003 | Benign | 4.03 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3937 | 0.6307 | 1 | 1 | 2.7 | -27.03 | ||||||||||||||||||||||||||||||||||
| c.3947A>T | N1316I 2D AIThe SynGAP1 missense variant N1316I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | -4.829 | Likely Benign | 0.635 | Likely Pathogenic | Likely Benign | 0.147 | Likely Benign | -2.86 | Deleterious | 0.009 | Benign | 0.004 | Benign | 3.91 | Benign | 0.00 | Affected | 0.0719 | 0.5354 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.3948T>A | N1316K 2D AIThe SynGAP1 missense variant N1316K is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451822‑T‑A). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none). Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451822-T-A | -3.815 | Likely Benign | 0.699 | Likely Pathogenic | Likely Benign | 0.044 | Likely Benign | -3.27 | Deleterious | 0.414 | Benign | 0.063 | Benign | 4.00 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2065 | 0.5500 | 0 | 1 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||
| c.3948T>G | N1316K 2D AIThe SynGAP1 missense variant N1316K is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451822‑T‑G). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign + 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.899122 | Disordered | 0.971970 | Binding | 0.380 | 0.885 | 0.750 | 6-33451822-T-G | -3.815 | Likely Benign | 0.699 | Likely Pathogenic | Likely Benign | 0.044 | Likely Benign | -3.27 | Deleterious | 0.414 | Benign | 0.063 | Benign | 4.00 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2065 | 0.5500 | 0 | 1 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||
| c.3949G>A | G1317S 2D  AIThe SynGAP1 missense variant G1317S is listed in ClinVar with an uncertain significance and is present in the gnomAD database. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized returns a benign prediction, and the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the computational evidence overwhelmingly points to a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | Conflicting | 3 | 6-33451823-G-A | 1 | 6.26e-7 | -3.522 | Likely Benign | 0.145 | Likely Benign | Likely Benign | 0.092 | Likely Benign | -2.45 | Neutral | 0.127 | Benign | 0.045 | Benign | 4.08 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2546 | 0.4935 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||
| c.3949G>C | G1317R 2D AIThe SynGAP1 missense variant G1317R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. No Foldetta stability prediction is available for this variant. Overall, the computational evidence overwhelmingly suggests that G1317R is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | -3.646 | Likely Benign | 0.619 | Likely Pathogenic | Likely Benign | 0.176 | Likely Benign | -2.17 | Neutral | 0.834 | Possibly Damaging | 0.307 | Benign | 4.04 | Benign | 0.00 | Affected | 0.1018 | 0.3791 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||||||
| c.3949G>T | G1317C 2D AIThe SynGAP1 missense variant G1317C is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451823‑G‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts benign, while Foldetta results are unavailable. Overall, the majority of reliable predictions favor a benign impact, and this is consistent with the lack of ClinVar evidence. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | 6-33451823-G-T | -5.850 | Likely Benign | 0.342 | Ambiguous | Likely Benign | 0.162 | Likely Benign | -3.83 | Deleterious | 0.939 | Possibly Damaging | 0.570 | Possibly Damaging | 3.99 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1345 | 0.3753 | -3 | -3 | 2.9 | 46.09 | |||||||||||||||||||||||||||||||||||||
| c.394T>A | F132I 2D AISynGAP1 missense variant F132I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic, two benign) and Foldetta results are unavailable. Because the majority of consensus tools predict benign and no ClinVar evidence contradicts this, the variant is most likely benign, although the pathogenic signal from AlphaMissense‑Optimized and the inconclusive SGM Consensus leave some uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -6.245 | Likely Benign | 0.974 | Likely Pathogenic | Likely Pathogenic | 0.211 | Likely Benign | -3.23 | Deleterious | 0.084 | Benign | 0.020 | Benign | 3.36 | Benign | 0.00 | Affected | 0.2306 | 0.1698 | 1 | 0 | 1.7 | -34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.394T>C | F132L 2D AIThe SynGAP1 missense variant F132L is not reported in ClinVar (status: none) but is present in gnomAD (ID 6‑33432691‑T‑C). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs 2 benign), and Foldetta (combining FoldX‑MD and Rosetta) has no available output. Overall, the majority of consensus tools lean toward a benign interpretation, and the single high‑accuracy pathogenic prediction is counterbalanced by inconclusive consensus and missing folding‑stability data. Thus, the variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | 6-33432691-T-C | -5.401 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.165 | Likely Benign | -2.99 | Deleterious | 0.000 | Benign | 0.002 | Benign | 3.46 | Benign | 0.00 | Affected | 3.61 | 5 | 0.2564 | 0.2669 | 0 | 2 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||||
| c.394T>G | F132V 2D AIThe SynGAP1 missense variant F132V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictions indicate a pathogenic effect, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -7.637 | In-Between | 0.979 | Likely Pathogenic | Likely Pathogenic | 0.328 | Likely Benign | -3.86 | Deleterious | 0.084 | Benign | 0.034 | Benign | 3.38 | Benign | 0.00 | Affected | 0.2349 | 0.1725 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3950G>A | G1317D 2D  AIThe SynGAP1 missense variant G1317D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451824‑G‑A). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and SIFT. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign prediction (2 benign vs. 1 pathogenic, with one uncertain). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | 6-33451824-G-A | -4.860 | Likely Benign | 0.520 | Ambiguous | Likely Benign | 0.081 | Likely Benign | -3.54 | Deleterious | 0.588 | Possibly Damaging | 0.212 | Benign | 4.03 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2115 | 0.2859 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||||
| c.3950G>C | G1317A 2D  AIThe SynGAP1 missense variant G1317A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion is consistent with the lack of ClinVar annotation. Thus, the variant is most likely benign and does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | -3.761 | Likely Benign | 0.190 | Likely Benign | Likely Benign | 0.049 | Likely Benign | -2.62 | Deleterious | 0.001 | Benign | 0.002 | Benign | 4.08 | Benign | 0.00 | Affected | 0.3582 | 0.4284 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3950G>T | G1317V 2D AIThe SynGAP1 missense variant G1317V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | -4.604 | Likely Benign | 0.286 | Likely Benign | Likely Benign | 0.050 | Likely Benign | -2.99 | Deleterious | 0.004 | Benign | 0.004 | Benign | 4.05 | Benign | 0.00 | Affected | 0.1132 | 0.3635 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3953T>A | L1318Q 2D AIThe SynGAP1 missense variant L1318Q is reported in gnomAD (variant ID 6‑33451827‑T‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 HumDiv and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates benign. No Foldetta stability analysis is available, so it does not influence the conclusion. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.968271 | Binding | 0.399 | 0.865 | 0.750 | 6-33451827-T-A | 1 | 6.32e-7 | -3.445 | Likely Benign | 0.120 | Likely Benign | Likely Benign | 0.077 | Likely Benign | -2.06 | Neutral | 0.834 | Possibly Damaging | 0.307 | Benign | 4.04 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1410 | 0.0903 | -2 | -2 | -7.3 | 14.97 | ||||||||||||||||||||||||||||||||||
| c.3953T>C | L1318P 2D AIThe SynGAP1 missense variant L1318P is listed in gnomAD (ID 6‑33451827‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, while pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support benignity: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for this variant, and this conclusion does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.887230 | Disordered | 0.968271 | Binding | 0.399 | 0.865 | 0.750 | 6-33451827-T-C | -2.307 | Likely Benign | 0.116 | Likely Benign | Likely Benign | 0.126 | Likely Benign | -2.90 | Deleterious | 0.813 | Possibly Damaging | 0.212 | Benign | 3.98 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3424 | 0.1411 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||
| c.3958C>A | P1320T 2D AIThe SynGAP1 missense variant P1320T is reported in gnomAD (variant ID 6‑33451832‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: the majority (REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a benign effect, while a minority (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT) predict pathogenicity. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | 6-33451832-C-A | 1 | 6.42e-7 | -5.355 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -0.86 | Neutral | 0.994 | Probably Damaging | 0.981 | Probably Damaging | 4.21 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1787 | 0.6392 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||
| c.3958C>G | P1320A 2D AIThe SynGAP1 missense variant P1320A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P1320A, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | -4.099 | Likely Benign | 0.055 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -0.92 | Neutral | 0.980 | Probably Damaging | 0.956 | Probably Damaging | 4.25 | Benign | 0.00 | Affected | 0.3517 | 0.5583 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3958C>T | P1320S 2D AIThe SynGAP1 missense variant P1320S is listed in ClinVar (ID 469160.0) with an “Uncertain” clinical significance and is present in gnomAD (variant ID 6‑33451832‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this is not in conflict with the ClinVar “Uncertain” status. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | Uncertain | 1 | 6-33451832-C-T | 2 | 1.28e-6 | -4.928 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -0.69 | Neutral | 0.980 | Probably Damaging | 0.968 | Probably Damaging | 4.25 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3390 | 0.5814 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||
| c.3959C>A | P1320H 2D AIThe SynGAP1 missense variant P1320H is listed in gnomAD (ID 6‑33451833‑C‑A) and has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | 6-33451833-C-A | 2 | 1.29e-6 | -6.296 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -1.05 | Neutral | 0.998 | Probably Damaging | 0.990 | Probably Damaging | 4.17 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1955 | 0.5461 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||
| c.3959C>G | P1320R 2D AIThe SynGAP1 missense variant P1320R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | -5.204 | Likely Benign | 0.160 | Likely Benign | Likely Benign | 0.142 | Likely Benign | -1.39 | Neutral | 0.994 | Probably Damaging | 0.987 | Probably Damaging | 4.25 | Benign | 0.00 | Affected | 0.1558 | 0.4052 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3959C>T | P1320L 2D AIThe SynGAP1 missense variant P1320L is reported in gnomAD (variant ID 6‑33451833‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also reports it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta’s protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect for P1320L, and this conclusion is not contradicted by any ClinVar classification (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | 6-33451833-C-T | -5.187 | Likely Benign | 0.094 | Likely Benign | Likely Benign | 0.115 | Likely Benign | -1.22 | Neutral | 0.994 | Probably Damaging | 0.981 | Probably Damaging | 4.18 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2622 | 0.6744 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||
| c.395T>A | F132Y 2D AIThe SynGAP1 missense variant F132Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of available predictions (five benign vs three pathogenic) suggest a benign impact. This consensus does not contradict any ClinVar annotation, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -8.220 | Likely Pathogenic | 0.839 | Likely Pathogenic | Ambiguous | 0.165 | Likely Benign | -1.98 | Neutral | 0.272 | Benign | 0.126 | Benign | 3.39 | Benign | 0.00 | Affected | 0.1635 | 0.1526 | 7 | 3 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.395T>C | F132S 2D AIThe SynGAP1 missense variant F132S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the balance of evidence (five pathogenic vs four benign predictions) indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -4.177 | Likely Benign | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.329 | Likely Benign | -4.54 | Deleterious | 0.567 | Possibly Damaging | 0.249 | Benign | 3.31 | Benign | 0.00 | Affected | 0.4869 | 0.0000 | -3 | -2 | -3.6 | -60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.395T>G | F132C 2D AIThe SynGAP1 missense variant F132C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Pathogenic” based on a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (derived from the four high‑accuracy predictors) also indicates likely pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -10.013 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.331 | Likely Benign | -4.55 | Deleterious | 0.938 | Possibly Damaging | 0.498 | Possibly Damaging | 3.29 | Benign | 0.00 | Affected | 0.2677 | 0.0925 | -4 | -2 | -0.3 | -44.04 | |||||||||||||||||||||||||||||||||||||||
| c.3967C>A | P1323T 2D AIThe SynGAP1 missense variant P1323T is reported in gnomAD (ID 6‑33451841‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as tolerated or benign. Only two algorithms—polyPhen‑2 HumDiv and SIFT—suggest a damaging impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates benign. Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with any ClinVar classification, as none is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451841-C-A | -6.296 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -0.37 | Neutral | 0.588 | Possibly Damaging | 0.227 | Benign | 3.95 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1988 | 0.5161 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3967C>G | P1323A 2D AIThe SynGAP1 missense variant P1323A is catalogued in gnomAD (ID 6‑33451841‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes, while the single pathogenic signal comes from SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign; Foldetta results are not available. Overall, the consensus of available predictions points to a benign impact for P1323A, and this conclusion is not contradicted by ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451841-C-G | -4.802 | Likely Benign | 0.053 | Likely Benign | Likely Benign | 0.040 | Likely Benign | -0.73 | Neutral | 0.011 | Benign | 0.017 | Benign | 3.92 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3743 | 0.3976 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||||
| c.3967C>T | P1323S 2D AIThe SynGAP1 missense variant P1323S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | -6.007 | Likely Benign | 0.072 | Likely Benign | Likely Benign | 0.077 | Likely Benign | -0.52 | Neutral | 0.588 | Possibly Damaging | 0.122 | Benign | 3.88 | Benign | 0.00 | Affected | 0.3758 | 0.4371 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.3968C>A | P1323Q 2D AIThe SynGAP1 missense variant P1323Q is reported in gnomAD (ID 6‑33451842‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that P1323Q is most likely benign, and this conclusion is not contradicted by any ClinVar classification (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451842-C-A | -6.134 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.033 | Likely Benign | -0.85 | Neutral | 0.712 | Possibly Damaging | 0.328 | Benign | 3.82 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1791 | 0.4657 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3968C>G | P1323R 2D AIThe SynGAP1 missense variant P1323R is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that P1323R is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | -5.785 | Likely Benign | 0.159 | Likely Benign | Likely Benign | 0.074 | Likely Benign | -0.67 | Neutral | 0.002 | Benign | 0.003 | Benign | 3.82 | Benign | 0.00 | Affected | 0.1725 | 0.3015 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3968C>T | P1323L 2D AIThe SynGAP1 missense variant P1323L is reported in gnomAD (variant ID 6‑33451842‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority‑vote) is benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451842-C-T | 3 | 1.95e-6 | -6.005 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.045 | Likely Benign | -1.04 | Neutral | 0.414 | Benign | 0.175 | Benign | 3.82 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2650 | 0.6052 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||
| c.396C>A | F132L 2D AIThe SynGAP1 missense variant F132L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic). Foldetta, a protein‑folding‑stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools (AlphaMissense‑Optimized and AlphaMissense‑Default) both predict pathogenicity, while the consensus and stability analyses are inconclusive or missing. Overall, the majority of individual predictors and the high‑accuracy tools lean toward a pathogenic effect, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -5.401 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.137 | Likely Benign | -2.99 | Deleterious | 0.000 | Benign | 0.002 | Benign | 3.46 | Benign | 0.00 | Affected | 3.61 | 5 | 0.2564 | 0.2669 | 0 | 2 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||||
| c.396C>G | F132L 2D AIThe SynGAP1 missense variant F132L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools (AlphaMissense‑Optimized and AlphaMissense‑Default) both predict pathogenicity, whereas the consensus and stability analyses are inconclusive or missing. Overall, the majority of individual predictors and the two high‑accuracy tools suggest a pathogenic effect, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -5.401 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.137 | Likely Benign | -2.99 | Deleterious | 0.000 | Benign | 0.002 | Benign | 3.46 | Benign | 0.00 | Affected | 3.61 | 5 | 0.2564 | 0.2669 | 0 | 2 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||||
| c.3970C>A | P1324T 2D AIThe SynGAP1 missense variant P1324T is listed in gnomAD (ID 6‑33451844‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the available predictions points to a benign effect, and this conclusion does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | 6-33451844-C-A | -5.289 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.072 | Likely Benign | 0.95 | Neutral | 0.588 | Possibly Damaging | 0.175 | Benign | 4.43 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1863 | 0.4761 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3970C>G | P1324A 2D AIThe SynGAP1 missense variant P1324A is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the consensus of the majority of tools and the high‑accuracy predictions point to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | -4.416 | Likely Benign | 0.056 | Likely Benign | Likely Benign | 0.030 | Likely Benign | -0.57 | Neutral | 0.011 | Benign | 0.017 | Benign | 4.30 | Benign | 0.00 | Affected | 0.3489 | 0.3776 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3970C>T | P1324S 2D AIThe SynGAP1 missense variant P1324S is listed in ClinVar as Benign (ClinVar ID 1137951.0) and is present in gnomAD (ID 6‑33451844‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote) also benign; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely benign, which is consistent with its ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | Likely Benign | 1 | 6-33451844-C-T | 5 | 3.26e-6 | -5.451 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.049 | Likely Benign | 0.35 | Neutral | 0.225 | Benign | 0.092 | Benign | 4.33 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3505 | 0.4450 | 1 | -1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||
| c.3971C>A | P1324Q 2D AIThe SynGAP1 missense variant P1324Q is reported in gnomAD (ID 6‑33451845‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools, including the high‑accuracy predictors, points to a benign effect. This prediction is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | 6-33451845-C-A | -5.577 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.028 | Likely Benign | -0.69 | Neutral | 0.027 | Benign | 0.010 | Benign | 4.27 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1744 | 0.4736 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3971C>G | P1324R 2D AIThe SynGAP1 missense variant P1324R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | -5.718 | Likely Benign | 0.154 | Likely Benign | Likely Benign | 0.053 | Likely Benign | -0.80 | Neutral | 0.414 | Benign | 0.133 | Benign | 4.28 | Benign | 0.00 | Affected | 0.1723 | 0.3613 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3971C>T | P1324L 2D AIThe SynGAP1 missense variant P1324L is reported in gnomAD (ID 6‑33451845‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | 6-33451845-C-T | -5.549 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.072 | Likely Benign | -1.17 | Neutral | 0.414 | Benign | 0.175 | Benign | 4.26 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2507 | 0.5864 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||
| c.3973C>A | P1325T 2D AIThe SynGAP1 missense variant P1325T is reported in gnomAD (ID 6‑33451847‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign effect for P1325T, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451847-C-A | -5.880 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.135 | Likely Benign | -0.14 | Neutral | 0.000 | Benign | 0.001 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1814 | 0.5150 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3973C>G | P1325A 2D AIThe SynGAP1 missense variant P1325A is reported in gnomAD (ID 6‑33451847‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only SIFT predicts a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy methods confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign impact for P1325A, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451847-C-G | -3.786 | Likely Benign | 0.058 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -0.95 | Neutral | 0.019 | Benign | 0.004 | Benign | 4.10 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3650 | 0.4016 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||||
| c.3973C>T | P1325S 2D AIThe SynGAP1 missense variant P1325S is reported in gnomAD (ID 6‑33451847‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451847-C-T | 1 | 6.43e-7 | -5.273 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.053 | Likely Benign | 0.29 | Neutral | 0.010 | Benign | 0.007 | Benign | 4.10 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3657 | 0.4360 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3974C>A | P1325H 2D AIThe SynGAP1 missense variant P1325H is reported in gnomAD (variant ID 6‑33451848‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv and SIFT. The high‑accuracy consensus methods reinforce the benign assessment: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” No result is available from the Foldetta stability analysis. Overall, the majority of evidence points to a benign effect. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451848-C-A | -6.970 | Likely Benign | 0.136 | Likely Benign | Likely Benign | 0.056 | Likely Benign | 0.10 | Neutral | 0.704 | Possibly Damaging | 0.187 | Benign | 4.04 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2103 | 0.4364 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.3974C>G | P1325R 2D AIThe SynGAP1 missense variant P1325R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and there is no conflict with ClinVar status, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | -5.427 | Likely Benign | 0.173 | Likely Benign | Likely Benign | 0.051 | Likely Benign | -0.33 | Neutral | 0.182 | Benign | 0.029 | Benign | 4.06 | Benign | 0.00 | Affected | 0.1668 | 0.3081 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.3974C>T | P1325L 2D AIThe SynGAP1 missense variant P1325L is listed in ClinVar (ID 1720534.0) with an uncertain significance designation and is present in gnomAD (variant ID 6‑33451848‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect for P1325L, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | Uncertain | 1 | 6-33451848-C-T | -5.256 | Likely Benign | 0.085 | Likely Benign | Likely Benign | 0.146 | Likely Benign | -1.05 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.05 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2616 | 0.6073 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||
| c.3976C>A | P1326T 2D AIThe SynGAP1 missense variant P1326T is reported in gnomAD (ID 6‑33451850‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority of the same four high‑accuracy tools) is benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that P1326T is most likely benign, and this assessment does not contradict any ClinVar classification because no ClinVar variant is listed. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | 6-33451850-C-A | -5.755 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.135 | Likely Benign | -0.26 | Neutral | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 3.71 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1955 | 0.5517 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3976C>G | P1326A 2D AIThe SynGAP1 missense variant P1326A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | -4.450 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.097 | Likely Benign | -0.76 | Neutral | 0.996 | Probably Damaging | 0.986 | Probably Damaging | 3.73 | Benign | 0.00 | Affected | 0.3728 | 0.4333 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.3976C>T | P1326S 2D AIThe SynGAP1 missense variant P1326S is listed in ClinVar with no submitted interpretation and is present in gnomAD (ID 6‑33451850‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence—including the high‑accuracy tools—supports a benign classification. This conclusion does not contradict the ClinVar status, which currently has no pathogenic assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | 6-33451850-C-T | -5.221 | Likely Benign | 0.121 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -0.35 | Neutral | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 3.74 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3722 | 0.4563 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||
| c.3977C>A | P1326Q 2D AIThe SynGAP1 missense variant P1326Q is listed in ClinVar (ID 2806103.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33451851‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar “Uncertain” classification, which remains inconclusive. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | Uncertain | 1 | 6-33451851-C-A | 1 | 6.40e-7 | -5.422 | Likely Benign | 0.128 | Likely Benign | Likely Benign | 0.138 | Likely Benign | -0.86 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.62 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1769 | 0.4457 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||
| c.3977C>G | P1326R 2D AIThe SynGAP1 missense variant P1326R is listed in ClinVar (ID 2429486.0) with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote) also benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | Uncertain | 1 | -5.097 | Likely Benign | 0.240 | Likely Benign | Likely Benign | 0.133 | Likely Benign | -0.82 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.62 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1710 | 0.2808 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||
| c.3977C>T | P1326L 2D AIThe SynGAP1 missense variant P1326L is listed in ClinVar (ID 1004879.0) with an “Uncertain” clinical significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as “Likely Benign,” and AlphaMissense‑Optimized also predicts benign. No Foldetta (FoldX‑MD/ Rosetta) stability result is available for this variant. Overall, the majority of evidence—including the high‑confidence SGM consensus and AlphaMissense‑Optimized prediction—supports a benign classification, which does not contradict the current ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | Uncertain | 1 | -5.541 | Likely Benign | 0.115 | Likely Benign | Likely Benign | 0.117 | Likely Benign | -1.06 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.62 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2723 | 0.6113 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||
| c.3982C>T | R1328W 2D AISynGAP1 missense variant R1328W is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33451856‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default all predict a pathogenic outcome; ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta stability analysis is unavailable. Taken together, the majority of evidence points to a benign effect, which does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.887230 | Disordered | 0.911775 | Binding | 0.360 | 0.762 | 0.875 | Uncertain | 1 | 6-33451856-C-T | 4 | 2.56e-6 | -7.022 | In-Between | 0.779 | Likely Pathogenic | Likely Benign | 0.125 | Likely Benign | -2.40 | Neutral | 0.997 | Probably Damaging | 0.756 | Possibly Damaging | 4.04 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1349 | 0.2906 | -3 | 2 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||
| c.3985C>A | L1329M 2D AIThe SynGAP1 missense change L1329M is recorded in gnomAD (ID 6‑33451859‑C‑A) but has no ClinVar entry. Functional prediction tools split into two consensus groups: benign‑predicted by REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic‑predicted by polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized scores benign, and the SGM‑Consensus likewise indicates benign. Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.924905 | Binding | 0.336 | 0.748 | 0.875 | 6-33451859-C-A | -5.493 | Likely Benign | 0.780 | Likely Pathogenic | Likely Benign | 0.087 | Likely Benign | -1.16 | Neutral | 0.994 | Probably Damaging | 0.990 | Probably Damaging | 3.10 | Benign | 0.00 | Affected | 3.77 | 5 | 0.0914 | 0.4273 | 2 | 4 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||
| c.3985C>G | L1329V 2D AIThe SynGAP1 missense variant L1329V is catalogued in gnomAD (6‑33451859‑C‑G) but has no ClinVar entry. Functional prediction tools show mixed results: benign calls come from REVEL, PROVEAN, ESM1b, and FATHMM, whereas pathogenic calls are reported by polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. When predictions are grouped by consensus, the majority of standard algorithms (REVEL, PROVEAN, ESM1b, FATHMM) support a benign effect, while a minority (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default) suggest pathogenicity. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts a benign outcome, and the SGM‑Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign effect, and this conclusion is consistent with the lack of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.924905 | Binding | 0.336 | 0.748 | 0.875 | 6-33451859-C-G | 1 | 6.40e-7 | -4.209 | Likely Benign | 0.770 | Likely Pathogenic | Likely Benign | 0.077 | Likely Benign | -1.55 | Neutral | 0.980 | Probably Damaging | 0.952 | Probably Damaging | 3.18 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1571 | 0.3579 | 1 | 2 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||
| c.3986T>A | L1329Q 2D AIThe SynGAP1 missense variant L1329Q is reported in gnomAD (ID 6‑33451860‑T‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a deleterious effect, indicating that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.924905 | Binding | 0.336 | 0.748 | 0.875 | 6-33451860-T-A | -4.106 | Likely Benign | 0.956 | Likely Pathogenic | Likely Pathogenic | 0.157 | Likely Benign | -3.31 | Deleterious | 0.994 | Probably Damaging | 0.993 | Probably Damaging | 3.05 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1382 | 0.1505 | -2 | -2 | -7.3 | 14.97 | |||||||||||||||||||||||||||||||||||||
| c.3986T>C | L1329P 2D AIThe SynGAP1 missense variant L1329P is listed in gnomAD (ID 6‑33451860‑T‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, while pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a tie and is therefore inconclusive. No Foldetta stability assessment is available. Overall, the majority of high‑confidence predictors (five pathogenic vs three benign) lean toward a pathogenic effect. Because ClinVar contains no classification, there is no conflict with existing clinical annotation. Thus, based on current in silico evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.924905 | Binding | 0.336 | 0.748 | 0.875 | 6-33451860-T-C | -2.903 | Likely Benign | 0.951 | Likely Pathogenic | Ambiguous | 0.165 | Likely Benign | -3.74 | Deleterious | 0.994 | Probably Damaging | 0.993 | Probably Damaging | 3.04 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3396 | 0.1794 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||||
| c.3986T>G | L1329R 2D AIThe SynGAP1 missense variant L1329R is catalogued in gnomAD (ID 6‑33451860‑T‑G) but has no ClinVar submission. Functional prediction tools cluster into two groups: benign predictions from REVEL, ESM1b, and FATHMM; pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized returns an uncertain result. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive due to a 2‑to‑2 split. No Foldetta stability assessment is available. Overall, the majority of high‑confidence predictors (five pathogenic vs three benign) lean toward a pathogenic effect. Because ClinVar contains no classification, there is no conflict with existing clinical annotations. Thus, based on current in silico evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.924905 | Binding | 0.336 | 0.748 | 0.875 | 6-33451860-T-G | -3.636 | Likely Benign | 0.923 | Likely Pathogenic | Ambiguous | 0.141 | Likely Benign | -3.16 | Deleterious | 0.994 | Probably Damaging | 0.990 | Probably Damaging | 3.05 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1594 | 0.1547 | -2 | -3 | -8.3 | 43.03 | |||||||||||||||||||||||||||||||||||||
| c.398T>C | L133P 2D AIThe SynGAP1 missense variant L133P is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. ESM1b remains uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (benign), and PROVEAN (pathogenic), yields a pathogenic consensus. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.422041 | Structured | 0.718429 | Binding | 0.320 | 0.896 | 0.250 | -7.744 | In-Between | 0.982 | Likely Pathogenic | Likely Pathogenic | 0.342 | Likely Benign | -3.16 | Deleterious | 0.700 | Possibly Damaging | 0.483 | Possibly Damaging | 3.52 | Benign | 0.00 | Affected | 0.3270 | 0.1048 | -3 | -3 | -5.4 | -16.04 | ||||||||||||||||||||||||||||||||||||||||
| c.398T>G | L133R 2D AIThe SynGAP1 missense variant L133R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome (3 pathogenic vs. 1 benign votes). AlphaMissense‑Optimized independently predicts pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence supports a pathogenic classification for L133R, and this assessment does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.422041 | Structured | 0.718429 | Binding | 0.320 | 0.896 | 0.250 | -8.857 | Likely Pathogenic | 0.965 | Likely Pathogenic | Likely Pathogenic | 0.389 | Likely Benign | -2.57 | Deleterious | 0.002 | Benign | 0.005 | Benign | 3.55 | Benign | 0.00 | Affected | 0.1522 | 0.0479 | -3 | -2 | -8.3 | 43.03 | |||||||||||||||||||||||||||||||||||||||
| c.3991A>C | I1331L 2D AIThe SynGAP1 missense variant I1331L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), which collectively classify the variant as likely benign. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized result is uncertain, and Foldetta stability analysis is unavailable. Overall, the balance of evidence leans toward a benign interpretation, with no conflict with ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -2.450 | Likely Benign | 0.892 | Likely Pathogenic | Ambiguous | 0.180 | Likely Benign | -1.04 | Neutral | 0.762 | Possibly Damaging | 0.785 | Possibly Damaging | 3.45 | Benign | 0.00 | Affected | 0.0752 | 0.3616 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.3991A>G | I1331V 2D AIThe SynGAP1 missense variant I1331V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain,” SGM‑Consensus as “Likely Benign,” and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -2.799 | Likely Benign | 0.899 | Likely Pathogenic | Ambiguous | 0.138 | Likely Benign | -0.51 | Neutral | 0.581 | Possibly Damaging | 0.785 | Possibly Damaging | 3.59 | Benign | 0.00 | Affected | 0.1091 | 0.3074 | 4 | 3 | -0.3 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3991A>T | I1331F 2D AIThe SynGAP1 missense variant I1331F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain” and the SGM‑Consensus as “Likely Benign”; Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign interpretation, with no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -4.220 | Likely Benign | 0.946 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -2.46 | Neutral | 0.984 | Probably Damaging | 0.969 | Probably Damaging | 3.31 | Benign | 0.00 | Affected | 0.0522 | 0.3225 | 1 | 0 | -1.7 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.3992T>A | I1331N 2D AIThe SynGAP1 missense variant I1331N is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451866‑T‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of evaluated tools predict a pathogenic impact, and there is no conflict with ClinVar status. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | 6-33451866-T-A | -3.788 | Likely Benign | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.237 | Likely Benign | -4.09 | Deleterious | 0.984 | Probably Damaging | 0.979 | Probably Damaging | 3.29 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1012 | 0.0666 | -3 | -2 | -8.0 | 0.94 | |||||||||||||||||||||||||||||||||||||
| c.3992T>C | I1331T 2D AIThe SynGAP1 missense variant I1331T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence (seven pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -2.953 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.170 | Likely Benign | -2.69 | Deleterious | 0.947 | Possibly Damaging | 0.950 | Probably Damaging | 3.32 | Benign | 0.00 | Affected | 0.1115 | 0.1550 | 0 | -1 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||||||||
| c.3992T>G | I1331S 2D AIThe SynGAP1 I1331S variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence (six pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -3.014 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.230 | Likely Benign | -3.40 | Deleterious | 0.984 | Probably Damaging | 0.969 | Probably Damaging | 3.30 | Benign | 0.00 | Affected | 0.2929 | 0.0836 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3993T>G | I1331M 2D AIThe SynGAP1 I1331M missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on benign impact include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and Foldetta results are unavailable. Considering the majority of consensus‑based and individual predictors, the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -4.202 | Likely Benign | 0.883 | Likely Pathogenic | Ambiguous | 0.143 | Likely Benign | -1.69 | Neutral | 0.984 | Probably Damaging | 0.979 | Probably Damaging | 3.32 | Benign | 0.00 | Affected | 0.0698 | 0.3220 | 2 | 1 | -2.6 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.3994A>C | T1332P 2D AIThe SynGAP1 missense variant T1332P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is tied 2‑2 and thus unavailable, and Foldetta results are not provided. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any existing ClinVar status because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | -3.404 | Likely Benign | 0.918 | Likely Pathogenic | Ambiguous | 0.238 | Likely Benign | -3.12 | Deleterious | 0.994 | Probably Damaging | 0.981 | Probably Damaging | 2.95 | Benign | 0.00 | Affected | 0.2185 | 0.5308 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||||||
| c.3994A>G | T1332A 2D AIThe SynGAP1 missense variant T1332A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) indicate a pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | -3.521 | Likely Benign | 0.887 | Likely Pathogenic | Ambiguous | 0.150 | Likely Benign | -2.85 | Deleterious | 0.953 | Possibly Damaging | 0.935 | Probably Damaging | 3.02 | Benign | 0.00 | Affected | 0.4042 | 0.4766 | 1 | 0 | 2.5 | -30.03 | ||||||||||||||||||||||||||||||||||||||||
| c.3994A>T | T1332S 2D AIThe SynGAP1 missense variant T1332S is reported in gnomAD (ID 6‑33451868‑A‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as “Likely Benign” (three benign votes versus one pathogenic). High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect; this conclusion is not contradicted by ClinVar, which contains no classification for T1332S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | 6-33451868-A-T | -3.085 | Likely Benign | 0.674 | Likely Pathogenic | Likely Benign | 0.163 | Likely Benign | -2.29 | Neutral | 0.980 | Probably Damaging | 0.935 | Probably Damaging | 3.00 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3451 | 0.5007 | 1 | 1 | -0.1 | -14.03 | ||||||||||||||||||||||||||||||||||||
| c.3995C>A | T1332K 2D AIThe SynGAP1 missense variant T1332K is catalogued in gnomAD (ID 6‑33451869‑C‑A) but has no ClinVar submission. Functional prediction tools cluster into two groups: benign predictions from REVEL, ESM1b, and FATHMM; pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. A high‑accuracy consensus (SGM) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a 2‑to‑2 split, leaving the consensus inconclusive. No Foldetta stability assessment is available. Overall, the majority of evidence (five pathogenic versus three benign) points to a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | 6-33451869-C-A | -3.264 | Likely Benign | 0.935 | Likely Pathogenic | Ambiguous | 0.247 | Likely Benign | -3.48 | Deleterious | 0.998 | Probably Damaging | 0.989 | Probably Damaging | 2.96 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1424 | 0.4376 | -1 | 0 | -3.2 | 27.07 | |||||||||||||||||||||||||||||||||||||
| c.3995C>G | T1332R 2D AIThe SynGAP1 missense variant T1332R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie and therefore unavailable; Foldetta results are not provided. Overall, the balance of evidence (five pathogenic versus three benign predictions) indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | -3.354 | Likely Benign | 0.878 | Likely Pathogenic | Ambiguous | 0.271 | Likely Benign | -3.59 | Deleterious | 0.998 | Probably Damaging | 0.993 | Probably Damaging | 2.96 | Benign | 0.00 | Affected | 0.1173 | 0.3867 | -1 | -1 | -3.8 | 55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3995C>T | T1332M 2D  AISynGAP1 missense variant T1332M is listed as Benign in ClinVar (ID 794425) and is present in gnomAD (6‑33451869‑C‑T). Functional prediction tools show mixed results: benign calls come from REVEL, ESM1b, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized returned an uncertain result, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie, and no Foldetta stability data are available. Overall, the majority of evidence points toward a pathogenic effect, which contradicts the ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | Likely Benign | 1 | 6-33451869-C-T | 20 | 1.86e-5 | -4.107 | Likely Benign | 0.948 | Likely Pathogenic | Ambiguous | 0.252 | Likely Benign | -3.63 | Deleterious | 1.000 | Probably Damaging | 0.991 | Probably Damaging | 2.95 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1551 | 0.6513 | -1 | -1 | 2.6 | 30.09 | |||||||||||||||||||||||||||||||||
| c.3997G>A | E1333K 2D AISynGAP1 missense variant E1333K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, and Foldetta results are unavailable. Overall, the consensus of multiple benign‑predicting tools and the SGM‑Consensus suggests a benign outcome, whereas a subset of tools indicates pathogenicity. Thus, the variant is most likely benign based on the current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.953319 | Binding | 0.347 | 0.746 | 0.750 | -5.038 | Likely Benign | 0.942 | Likely Pathogenic | Ambiguous | 0.244 | Likely Benign | -2.49 | Neutral | 0.980 | Probably Damaging | 0.956 | Probably Damaging | 2.84 | Benign | 0.00 | Affected | 0.2728 | 0.7491 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.3997G>C | E1333Q 2D AIThe SynGAP1 missense variant E1333Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (3 benign vs. 1 pathogenic votes) also indicates benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the preponderance of evidence points to a benign effect for E1333Q, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.953319 | Binding | 0.347 | 0.746 | 0.750 | -4.034 | Likely Benign | 0.746 | Likely Pathogenic | Likely Benign | 0.217 | Likely Benign | -1.84 | Neutral | 0.980 | Probably Damaging | 0.968 | Probably Damaging | 2.83 | Benign | 0.00 | Affected | 0.1548 | 0.7617 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.3998A>C | E1333A 2D AIThe SynGAP1 missense variant E1333A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returned an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2). Foldetta, which would assess protein‑folding stability, has no available output for this variant. Overall, more tools predict pathogenicity than benignity, and no ClinVar entry contradicts this assessment. Thus, the variant is most likely pathogenic based on the current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.953319 | Binding | 0.347 | 0.746 | 0.750 | -3.636 | Likely Benign | 0.834 | Likely Pathogenic | Ambiguous | 0.254 | Likely Benign | -3.76 | Deleterious | 0.980 | Probably Damaging | 0.956 | Probably Damaging | 2.83 | Benign | 0.00 | Affected | 0.4177 | 0.7382 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3998A>G | E1333G 2D AIThe SynGAP1 missense variant E1333G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie and therefore unavailable; Foldetta, which would evaluate protein‑folding stability, has no result for this variant. Overall, more tools predict pathogenicity than benignity, and no ClinVar entry contradicts this assessment. Thus, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.953319 | Binding | 0.347 | 0.746 | 0.750 | -4.504 | Likely Benign | 0.801 | Likely Pathogenic | Ambiguous | 0.260 | Likely Benign | -4.31 | Deleterious | 0.994 | Probably Damaging | 0.968 | Probably Damaging | 2.83 | Benign | 0.00 | Affected | 0.3101 | 0.6217 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.3998A>T | E1333V 2D AIThe SynGAP1 E1333V missense change is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs two benign votes), and Foldetta results are unavailable. Overall, the majority of evidence (six pathogenic vs three benign) points to a pathogenic impact. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar assertion exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.930790 | Disordered | 0.953319 | Binding | 0.347 | 0.746 | 0.750 | -4.322 | Likely Benign | 0.974 | Likely Pathogenic | Likely Pathogenic | 0.289 | Likely Benign | -4.30 | Deleterious | 0.994 | Probably Damaging | 0.981 | Probably Damaging | 2.81 | Benign | 0.00 | Affected | 0.0994 | 0.7615 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||||
| c.3999G>C | E1333D 2D AIThe SynGAP1 missense variant E1333D is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for E1333D, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.953319 | Binding | 0.347 | 0.746 | 0.750 | -4.239 | Likely Benign | 0.651 | Likely Pathogenic | Likely Benign | 0.117 | Likely Benign | -1.78 | Neutral | 0.953 | Possibly Damaging | 0.935 | Probably Damaging | 2.88 | Benign | 0.00 | Affected | 0.1989 | 0.5029 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.3999G>T | E1333D 2D AIThe SynGAP1 missense variant E1333D is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors indicates a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all predict a likely benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT classify the change as pathogenic, and AlphaMissense‑Default also predicts pathogenicity. High‑accuracy tools specifically give a benign prediction for AlphaMissense‑Optimized and a likely benign result for the SGM‑Consensus; Foldetta data are unavailable. Overall, the majority of evidence supports a benign interpretation, and this is consistent with the absence of a ClinVar pathogenic claim. Thus, the variant is most likely benign, and this is consistent with the lack of a ClinVar pathogenic claim. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.953319 | Binding | 0.347 | 0.746 | 0.750 | -4.239 | Likely Benign | 0.651 | Likely Pathogenic | Likely Benign | 0.117 | Likely Benign | -1.78 | Neutral | 0.953 | Possibly Damaging | 0.935 | Probably Damaging | 2.88 | Benign | 0.00 | Affected | 0.1989 | 0.5029 | 3 | 2 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||||||
| c.39C>G | I13M 2D AIThe SynGAP1 missense variant I13M is reported in gnomAD (ID 6‑33420303‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus itself is likely benign. Foldetta results are not available, so they do not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | 6-33420303-C-G | 1 | 6.49e-7 | -4.097 | Likely Benign | 0.170 | Likely Benign | Likely Benign | 0.093 | Likely Benign | 0.16 | Neutral | 0.296 | Benign | 0.022 | Benign | 4.04 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0883 | 0.3806 | 1 | 2 | -2.6 | 18.03 | ||||||||||||||||||||||||||||||||||
| c.3G>A | M1I 2D AIThe SynGAP1 missense variant M1I is listed in ClinVar (ID 833646.0) with an uncertain significance annotation and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM all classify the substitution as benign, while SIFT uniquely predicts it to be pathogenic. The consensus score from the SGM framework, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy predictors are incomplete: AlphaMissense‑Optimized and the Foldetta stability assessment are unavailable for this variant. Taking the collective evidence into account, the variant is most likely benign, and this assessment does not conflict with the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.540766 | Binding | 0.354 | 0.924 | 0.875 | Conflicting | 3 | -5.397 | Likely Benign | 0.227 | Likely Benign | -0.17 | Neutral | 0.001 | Benign | 0.000 | Benign | 4.25 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1655 | 0.4008 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||||
| c.4000A>C | N1334H 2D AIThe SynGAP1 missense variant N1334H is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of reliable tools and the high‑accuracy consensus predict a benign impact. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | -4.954 | Likely Benign | 0.496 | Ambiguous | Likely Benign | 0.164 | Likely Benign | -2.99 | Deleterious | 0.985 | Probably Damaging | 0.927 | Probably Damaging | 3.50 | Benign | 0.00 | Affected | 0.1963 | 0.6246 | 2 | 1 | 0.3 | 23.04 | ||||||||||||||||||||||||||||||||||||||||
| c.4000A>G | N1334D 2D AIThe SynGAP1 missense variant N1334D (ClinVar ID 3653769.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33451874‑A‑G). Functional prediction tools show a split: benign calls come from REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic calls come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments give a benign result from AlphaMissense‑Optimized, an inconclusive SGM Consensus (a 2‑vs‑2 majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and no available Foldetta stability data. Overall, the majority of predictions (5/10) indicate pathogenicity, and the high‑accuracy tools do not overturn this trend. Therefore, the variant is most likely pathogenic, which does not contradict its ClinVar status of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | Uncertain | 1 | 6-33451874-A-G | -4.584 | Likely Benign | 0.674 | Likely Pathogenic | Likely Benign | 0.126 | Likely Benign | -3.06 | Deleterious | 0.886 | Possibly Damaging | 0.522 | Possibly Damaging | 3.55 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2444 | 0.3307 | 1 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||
| c.4000A>T | N1334Y 2D AIThe SynGAP1 missense variant N1334Y is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—yields a tie and is therefore inconclusive; Foldetta results are unavailable. Overall, the majority of conventional tools predict a pathogenic impact, but the single high‑accuracy benign prediction and the inconclusive consensus suggest uncertainty. Consequently, the variant is most likely pathogenic based on the prevailing evidence, and this assessment does not contradict any ClinVar annotation because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | -5.916 | Likely Benign | 0.770 | Likely Pathogenic | Likely Benign | 0.249 | Likely Benign | -4.62 | Deleterious | 0.985 | Probably Damaging | 0.852 | Possibly Damaging | 3.49 | Benign | 0.00 | Affected | 0.0716 | 0.5447 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||||||||||||
| c.4001A>C | N1334T 2D AIThe SynGAP1 missense variant N1334T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs. two benign votes). Foldetta, which evaluates protein‑folding stability, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and there is no ClinVar entry to contradict this assessment. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | -4.249 | Likely Benign | 0.576 | Likely Pathogenic | Likely Benign | 0.103 | Likely Benign | -3.45 | Deleterious | 0.047 | Benign | 0.063 | Benign | 3.59 | Benign | 0.00 | Affected | 0.1600 | 0.5519 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||||||||||||
| c.4001A>G | N1334S 2D AIThe SynGAP1 missense variant N1334S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign consensus (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | -3.989 | Likely Benign | 0.409 | Ambiguous | Likely Benign | 0.107 | Likely Benign | -2.94 | Deleterious | 0.557 | Possibly Damaging | 0.348 | Benign | 3.56 | Benign | 0.00 | Affected | 0.3542 | 0.4708 | 1 | 1 | 2.7 | -27.03 | ||||||||||||||||||||||||||||||||||||||||
| c.4001A>T | N1334I 2D AIThe SynGAP1 missense variant N1334I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the balance of evidence (seven pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | -5.880 | Likely Benign | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.193 | Likely Benign | -5.06 | Deleterious | 0.985 | Probably Damaging | 0.721 | Possibly Damaging | 3.50 | Benign | 0.00 | Affected | 0.0861 | 0.5491 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.4002C>A | N1334K 2D AIThe SynGAP1 missense variant N1334K is listed in gnomAD (ID 6‑33451876‑C‑A) but has no ClinVar record. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The high‑accuracy consensus (SGM Consensus) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN is inconclusive (2 pathogenic vs 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, more tools (five) predict pathogenicity than benign (three), and the high‑accuracy methods do not overturn this trend. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | 6-33451876-C-A | -4.875 | Likely Benign | 0.934 | Likely Pathogenic | Ambiguous | 0.109 | Likely Benign | -3.49 | Deleterious | 0.979 | Probably Damaging | 0.756 | Possibly Damaging | 3.54 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2355 | 0.5163 | 0 | 1 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||
| c.4002C>G | N1334K 2D AIThe SynGAP1 missense variant N1334K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, whereas pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are limited: AlphaMissense‑Optimized remains inconclusive; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑to‑2 tie and therefore unavailable; Foldetta predictions are not provided. Overall, the balance of evidence favors a pathogenic effect for N1334K. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.960403 | Binding | 0.406 | 0.734 | 0.875 | -4.875 | Likely Benign | 0.934 | Likely Pathogenic | Ambiguous | 0.120 | Likely Benign | -3.49 | Deleterious | 0.979 | Probably Damaging | 0.756 | Possibly Damaging | 3.54 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2355 | 0.5163 | 0 | 1 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||
| c.4003G>A | G1335S 2D AIThe SynGAP1 missense variant G1335S is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33451877‑G‑A). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that G1335S is most likely pathogenic, which is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | Conflicting | 2 | 6-33451877-G-A | 3 | 2.37e-6 | -4.495 | Likely Benign | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.362 | Likely Benign | -3.79 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 2.04 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2452 | 0.5895 | 1 | 0 | -0.4 | 30.03 | ||||||||||||||||||||||||||||||||
| c.4003G>C | G1335R 2D AIThe SynGAP1 missense variant G1335R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely converge on a deleterious effect: benign predictions come from REVEL and ESM1b, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Pathogenic. Foldetta results are not available. Overall, the preponderance of evidence supports a pathogenic classification for G1335R, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | -4.921 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.394 | Likely Benign | -5.06 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 0.1071 | 0.5024 | -3 | -2 | -4.1 | 99.14 | |||||||||||||||||||||||||||||||||||||||
| c.4003G>T | G1335C 2D AIThe SynGAP1 missense variant G1335C is listed in gnomAD (variant ID 6-33451877‑G‑T) but has no ClinVar entry. Functional prediction tools split into two groups: benign predictions come from REVEL and ESM1b, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta results are not available for this variant. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | 6-33451877-G-T | 1 | 7.91e-7 | -6.878 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.426 | Likely Benign | -5.51 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.01 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1376 | 0.4513 | -3 | -3 | 2.9 | 46.09 | ||||||||||||||||||||||||||||||||||
| c.4004G>A | G1335D 2D AIThe SynGAP1 missense variant G1335D is reported in ClinVar as “not listed” and is present in gnomAD (variant ID 6‑33451878‑G‑A). Functional prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta results are not available for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and the SGM‑Consensus suggests that G1335D is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | 6-33451878-G-A | -5.687 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.404 | Likely Benign | -4.42 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2120 | 0.3702 | -1 | 1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||
| c.4004G>C | G1335A 2D AIThe SynGAP1 missense variant G1335A is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also likely pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for G1335A, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | -4.942 | Likely Benign | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.357 | Likely Benign | -3.69 | Deleterious | 0.996 | Probably Damaging | 0.992 | Probably Damaging | 2.05 | Pathogenic | 0.00 | Affected | 0.3383 | 0.4953 | 1 | 0 | 2.2 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.4004G>T | G1335V 2D AIThe SynGAP1 missense variant G1335V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact, and this assessment does not conflict with the ClinVar status, which currently has no entry for G1335V. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | -4.982 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.394 | Likely Benign | -5.65 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 0.1124 | 0.4195 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.4006G>A | E1336K 2D AIThe SynGAP1 missense variant E1336K is listed in ClinVar (ID 984837) with an “Uncertain” status and is present in gnomAD (6‑33451880‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic impact are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains likely benign; Foldetta results are unavailable. Overall, the balance of evidence leans toward a benign effect, and this conclusion does not contradict the ClinVar designation, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | Conflicting | 3 | 6-33451880-G-A | 6 | 4.20e-6 | -4.697 | Likely Benign | 0.977 | Likely Pathogenic | Likely Pathogenic | 0.272 | Likely Benign | -2.44 | Neutral | 0.748 | Possibly Damaging | 0.079 | Benign | 3.23 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2630 | 0.7501 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||||
| c.4006G>C | E1336Q 2D AIThe SynGAP1 missense change E1336Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | -4.113 | Likely Benign | 0.769 | Likely Pathogenic | Likely Benign | 0.159 | Likely Benign | -1.88 | Neutral | 0.731 | Possibly Damaging | 0.301 | Benign | 3.21 | Benign | 0.00 | Affected | 0.1432 | 0.7427 | 2 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||||||
| c.4007A>C | E1336A 2D AIThe SynGAP1 missense variant E1336A is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Overall, the majority of evidence (five benign vs. three pathogenic) supports a benign classification. This prediction does not contradict any ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | -3.545 | Likely Benign | 0.879 | Likely Pathogenic | Ambiguous | 0.191 | Likely Benign | -3.78 | Deleterious | 0.345 | Benign | 0.099 | Benign | 3.22 | Benign | 0.00 | Affected | 0.4000 | 0.7385 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||||||||||||
| c.4007A>G | E1336G 2D AIThe SynGAP1 missense variant E1336G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized result is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 vs 2). Foldetta, which would assess protein‑folding stability, has no available result for this variant. Overall, the balance of evidence (five benign vs three pathogenic predictions) suggests the variant is most likely benign. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | -3.574 | Likely Benign | 0.932 | Likely Pathogenic | Ambiguous | 0.211 | Likely Benign | -4.36 | Deleterious | 0.345 | Benign | 0.109 | Benign | 3.20 | Benign | 0.00 | Affected | 0.3092 | 0.6170 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||||||||||||
| c.4007A>T | E1336V 2D AIThe SynGAP1 missense variant E1336V has no ClinVar record (ClinVar status: None) and is not present in gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is reported as uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a tie (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Consequently, the variant’s predicted impact is ambiguous, with an equal split between benign and pathogenic calls and no evidence from ClinVar to contradict this uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | -3.367 | Likely Benign | 0.932 | Likely Pathogenic | Ambiguous | 0.221 | Likely Benign | -4.46 | Deleterious | 0.789 | Possibly Damaging | 0.348 | Benign | 3.18 | Benign | 0.00 | Affected | 0.0991 | 0.7425 | -2 | -2 | 7.7 | -29.98 | ||||||||||||||||||||||||||||||||||||||||
| c.4008G>C | E1336D 2D AIThe SynGAP1 missense variant E1336D is listed in ClinVar (ID 3323942.0) as benign and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus result is a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yielding a benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, consistent with the ClinVar benign designation. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | Likely Benign | 1 | -3.344 | Likely Benign | 0.596 | Likely Pathogenic | Likely Benign | 0.062 | Likely Benign | -1.92 | Neutral | 0.001 | Benign | 0.003 | Benign | 3.30 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1995 | 0.5101 | 2 | 3 | 0.0 | -14.03 | |||||||||||||||||||||||||||||||||||
| c.4008G>T | E1336D 2D AIThe SynGAP1 missense variant E1336D is catalogued in gnomAD (ID 6‑33451882‑G‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign or likely benign outcome. Only two tools—SIFT and AlphaMissense‑Default—predict pathogenicity. High‑accuracy assessments further support benignity: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus also reports it as likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign classification, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.865454 | Disordered | 0.973342 | Binding | 0.336 | 0.717 | 0.750 | 6-33451882-G-T | -3.344 | Likely Benign | 0.596 | Likely Pathogenic | Likely Benign | 0.065 | Likely Benign | -1.92 | Neutral | 0.001 | Benign | 0.003 | Benign | 3.30 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1995 | 0.5101 | 2 | 3 | 0.0 | -14.03 | ||||||||||||||||||||||||||||||||||||
| c.4009T>A | F1337I 2D AIThe SynGAP1 missense variant F1337I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the majority of evaluated tools (seven pathogenic vs. three benign) indicate a pathogenic effect. This prediction is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -2.761 | Likely Benign | 0.982 | Likely Pathogenic | Likely Pathogenic | 0.252 | Likely Benign | -3.32 | Deleterious | 0.947 | Possibly Damaging | 0.950 | Probably Damaging | 2.77 | Benign | 0.00 | Affected | 0.2574 | 0.2875 | 1 | 0 | 1.7 | -34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.4009T>C | F1337L 2D AIThe SynGAP1 missense variant F1337L is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the majority of evaluated predictors (six pathogenic vs. three benign) indicate a likely pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -2.396 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.186 | Likely Benign | -3.17 | Deleterious | 0.880 | Possibly Damaging | 0.899 | Possibly Damaging | 2.81 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2765 | 0.3543 | 0 | 2 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||||
| c.4009T>G | F1337V 2D AIThe SynGAP1 missense variant F1337V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs two benign), and Foldetta results are unavailable. Overall, the majority of evidence (six pathogenic vs three benign) points to a pathogenic impact. The variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -2.425 | Likely Benign | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.335 | Likely Benign | -3.94 | Deleterious | 0.947 | Possibly Damaging | 0.932 | Probably Damaging | 2.76 | Benign | 0.00 | Affected | 0.2470 | 0.2902 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.400A>C | S134R 2D AIThe SynGAP1 missense variant S134R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -9.053 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.292 | Likely Benign | -2.54 | Deleterious | 0.380 | Benign | 0.147 | Benign | 3.84 | Benign | 0.00 | Affected | 0.0719 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.400A>G | S134G 2D AIThe SynGAP1 missense variant S134G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus (SGM‑Consensus) is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yielding a “Likely Benign” classification. AlphaMissense‑Optimized also reports a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple independent predictors and the high‑accuracy consensus indicates that the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -6.722 | Likely Benign | 0.457 | Ambiguous | Likely Benign | 0.120 | Likely Benign | -2.19 | Neutral | 0.034 | Benign | 0.035 | Benign | 3.84 | Benign | 0.00 | Affected | 0.2268 | 0.4008 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.400A>T | S134C 2D AIThe SynGAP1 missense variant S134C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen2_HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen2_HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains pathogenic; Foldetta results are unavailable. Overall, the majority of tools and the consensus prediction lean toward pathogenicity, which is not contradicted by ClinVar (no entry) but is opposed by the AlphaMissense‑Optimized benign call. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -9.184 | Likely Pathogenic | 0.774 | Likely Pathogenic | Likely Benign | 0.247 | Likely Benign | -3.12 | Deleterious | 0.876 | Possibly Damaging | 0.417 | Benign | 3.79 | Benign | 0.00 | Affected | 0.0767 | 0.5151 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.4010T>A | F1337Y 2D AIThe SynGAP1 missense variant F1337Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the balance of evidence, including the SGM‑Consensus and the majority of individual predictors, leans toward a benign interpretation. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -3.481 | Likely Benign | 0.855 | Likely Pathogenic | Ambiguous | 0.202 | Likely Benign | -1.80 | Neutral | 0.947 | Possibly Damaging | 0.899 | Possibly Damaging | 2.82 | Benign | 0.00 | Affected | 0.1722 | 0.2754 | 7 | 3 | -4.1 | 16.00 | |||||||||||||||||||||||||||||||||||||||
| c.4010T>C | F1337S 2D AIThe SynGAP1 missense variant F1337S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence (six pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -2.641 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.327 | Likely Benign | -4.65 | Deleterious | 0.984 | Probably Damaging | 0.969 | Probably Damaging | 2.74 | Benign | 0.00 | Affected | 0.4478 | 0.0743 | -3 | -2 | -3.6 | -60.10 | ||||||||||||||||||||||||||||||||||||||||
| c.4010T>G | F1337C 2D AIThe SynGAP1 missense variant F1337C is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence (six pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -4.628 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.357 | Likely Benign | -4.57 | Deleterious | 0.996 | Probably Damaging | 0.984 | Probably Damaging | 2.73 | Benign | 0.00 | Affected | 0.2619 | 0.1905 | -4 | -2 | -0.3 | -44.04 | ||||||||||||||||||||||||||||||||||||||||
| c.4011C>A | F1337L 2D AIThe SynGAP1 missense variant F1337L is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451885‑C‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of evaluated tools predict a pathogenic impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Thus, the variant is most likely pathogenic based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | 6-33451885-C-A | -2.396 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.111 | Likely Benign | -3.17 | Deleterious | 0.880 | Possibly Damaging | 0.899 | Possibly Damaging | 2.81 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2765 | 0.3543 | 0 | 2 | 1.0 | -34.02 | |||||||||||||||||||||||||||||||||||||
| c.4011C>G | F1337L 2D AIThe SynGAP1 missense variant F1337L is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, whereas pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic versus two benign votes), and Foldetta data are unavailable. With a predominance of pathogenic calls and a single high‑confidence pathogenic prediction, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar entry exists for F1337L. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -2.396 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.122 | Likely Benign | -3.17 | Deleterious | 0.880 | Possibly Damaging | 0.899 | Possibly Damaging | 2.81 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2765 | 0.3543 | 0 | 2 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||||
| c.4015A>C | N1339H 2D AIThe SynGAP1 missense variant N1339H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, SGM‑Consensus likewise indicates likely benign, while Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the preponderance of evidence from both general and high‑accuracy predictors, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is assigned). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | -3.487 | Likely Benign | 0.338 | Likely Benign | Likely Benign | 0.234 | Likely Benign | -2.98 | Deleterious | 0.994 | Probably Damaging | 0.987 | Probably Damaging | 2.87 | Benign | 0.00 | Affected | 0.1585 | 0.7644 | 2 | 1 | 0.3 | 23.04 | |||||||||||||||||||||||||||||||||||||||
| c.4015A>G | N1339D 2D AIThe SynGAP1 missense variant N1339D is catalogued in gnomAD (ID 6‑33451889‑A‑G) but has no ClinVar submission. Functional prediction tools show mixed results: benign calls come from REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. When aggregated into a consensus, the four contributing scores (AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) split evenly, leaving the SGM Consensus inconclusive. High‑accuracy assessments further indicate a benign outcome from AlphaMissense‑Optimized; the SGM Consensus and Foldetta predictions are unavailable. Overall, the majority of individual tools predict pathogenicity, and the high‑accuracy benign prediction does not overturn this trend. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | 6-33451889-A-G | -2.533 | Likely Benign | 0.634 | Likely Pathogenic | Likely Benign | 0.186 | Likely Benign | -3.04 | Deleterious | 0.980 | Probably Damaging | 0.956 | Probably Damaging | 2.92 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2087 | 0.4721 | 1 | 2 | 0.0 | 0.98 | |||||||||||||||||||||||||||||||||||||
| c.4015A>T | N1339Y 2D AIThe SynGAP1 missense variant N1339Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic) and Foldetta data are unavailable. Overall, the majority of conventional predictors lean toward pathogenicity, but the single high‑accuracy benign prediction and the inconclusive consensus leave the variant’s impact uncertain. Thus, the variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | -3.808 | Likely Benign | 0.578 | Likely Pathogenic | Likely Benign | 0.319 | Likely Benign | -4.83 | Deleterious | 0.994 | Probably Damaging | 0.990 | Probably Damaging | 2.87 | Benign | 0.00 | Affected | 0.0697 | 0.6573 | -2 | -2 | 2.2 | 49.07 | ||||||||||||||||||||||||||||||||||||||||
| c.4016A>C | N1339T 2D AIThe SynGAP1 missense variant N1339T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, and Foldetta results are unavailable. Based on the most reliable predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | -2.682 | Likely Benign | 0.448 | Ambiguous | Likely Benign | 0.251 | Likely Benign | -3.31 | Deleterious | 0.980 | Probably Damaging | 0.956 | Probably Damaging | 2.89 | Benign | 0.00 | Affected | 0.1443 | 0.7598 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||||||||||||
| c.4016A>G | N1339S 2D AIThe SynGAP1 missense variant N1339S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | -2.331 | Likely Benign | 0.197 | Likely Benign | Likely Benign | 0.178 | Likely Benign | -2.66 | Deleterious | 0.980 | Probably Damaging | 0.935 | Probably Damaging | 2.91 | Benign | 0.00 | Affected | 0.3735 | 0.7283 | 1 | 1 | 2.7 | -27.03 | |||||||||||||||||||||||||||||||||||||||
| c.4016A>T | N1339I 2D AIThe SynGAP1 missense variant N1339I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs four benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic based on current computational evidence, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | -3.104 | Likely Benign | 0.740 | Likely Pathogenic | Likely Benign | 0.306 | Likely Benign | -5.25 | Deleterious | 0.994 | Probably Damaging | 0.987 | Probably Damaging | 2.87 | Benign | 0.00 | Affected | 0.0747 | 0.6271 | -2 | -3 | 8.0 | -0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.4017C>A | N1339K 2D AIThe SynGAP1 missense variant N1339K is listed in gnomAD (ID 6‑33451891‑C‑A) but has no ClinVar record. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized returns an uncertain result. The high‑accuracy consensus (SGM Consensus) – a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN – is inconclusive (two pathogenic, two benign). Foldetta, a protein‑folding stability method, has no reported output for this variant. Overall, the majority of available tools (five pathogenic vs. three benign) predict a deleterious effect. The variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | 6-33451891-C-A | -3.009 | Likely Benign | 0.872 | Likely Pathogenic | Ambiguous | 0.169 | Likely Benign | -3.56 | Deleterious | 0.980 | Probably Damaging | 0.968 | Probably Damaging | 2.90 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2201 | 0.6507 | 0 | 1 | -0.4 | 14.07 | |||||||||||||||||||||||||||||||||||||
| c.4017C>G | N1339K 2D AISynGAP1 missense variant N1339K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain, the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑to‑2 tie, and Foldetta results are unavailable. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.771762 | Disordered | 0.977585 | Binding | 0.396 | 0.687 | 1.000 | -3.009 | Likely Benign | 0.872 | Likely Pathogenic | Ambiguous | 0.181 | Likely Benign | -3.56 | Deleterious | 0.980 | Probably Damaging | 0.968 | Probably Damaging | 2.90 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2201 | 0.6507 | 0 | 1 | -0.4 | 14.07 | ||||||||||||||||||||||||||||||||||||||
| c.401G>A | S134N 2D AIThe SynGAP1 missense variant S134N is listed in ClinVar with an “Uncertain” status (ClinVar ID 2819575.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Those that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The remaining tools—AlphaMissense‑Optimized and the SGM‑Consensus—are inconclusive; the consensus score is “Likely Benign” based on a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely benign, and Foldetta (combining FoldX‑MD and Rosetta) has no available result. Overall, the balance of evidence points to a benign impact, which does not contradict the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | Uncertain | 1 | -5.534 | Likely Benign | 0.813 | Likely Pathogenic | Ambiguous | 0.075 | Likely Benign | -1.62 | Neutral | 0.001 | Benign | 0.002 | Benign | 3.90 | Benign | 0.00 | Affected | 3.61 | 5 | 0.0964 | 0.4381 | 1 | 1 | -2.7 | 27.03 | |||||||||||||||||||||||||||||||||||
| c.401G>C | S134T 2D AIThe SynGAP1 missense variant S134T is listed in gnomAD (ID 6‑33432698‑G‑C) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact; this conclusion is not contradicted by ClinVar, which contains no classification for S134T. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | 6-33432698-G-C | 1 | 6.32e-7 | -5.187 | Likely Benign | 0.563 | Ambiguous | Likely Benign | 0.069 | Likely Benign | -1.76 | Neutral | 0.034 | Benign | 0.047 | Benign | 3.85 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1083 | 0.5326 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.401G>T | S134I 2D AIThe SynGAP1 missense variant S134I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -8.860 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.230 | Likely Benign | -3.27 | Deleterious | 0.041 | Benign | 0.031 | Benign | 3.81 | Benign | 0.00 | Affected | 0.0683 | 0.5191 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.4025A>T | D1342V 2D AIThe SynGAP1 missense variant D1342V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign and the SGM‑Consensus also indicating a likely benign outcome; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.981682 | Binding | 0.316 | 0.678 | 0.875 | -2.890 | Likely Benign | 0.317 | Likely Benign | Likely Benign | 0.089 | Likely Benign | -1.27 | Neutral | 0.588 | Possibly Damaging | 0.212 | Benign | 4.01 | Benign | 0.00 | Affected | 0.1214 | 0.5617 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.4027C>A | H1343N 2D AIThe SynGAP1 missense variant H1343N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -3.077 | Likely Benign | 0.081 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -1.09 | Neutral | 0.444 | Benign | 0.071 | Benign | 4.06 | Benign | 0.00 | Affected | 0.2398 | 0.3299 | 2 | 1 | -0.3 | -23.04 | |||||||||||||||||||||||||||||||||||||||
| c.4027C>G | H1343D 2D AIThe SynGAP1 missense variant H1343D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only SIFT predicts a pathogenic impact. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. AlphaMissense‑Optimized also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -3.136 | Likely Benign | 0.179 | Likely Benign | Likely Benign | 0.051 | Likely Benign | -1.29 | Neutral | 0.444 | Benign | 0.071 | Benign | 4.07 | Benign | 0.00 | Affected | 0.3011 | 0.2403 | 1 | -1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||||
| c.4027C>T | H1343Y 2D AIThe SynGAP1 missense variant H1343Y is reported in gnomAD (ID 6‑33451901‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact for H1343Y, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | 6-33451901-C-T | -3.479 | Likely Benign | 0.139 | Likely Benign | Likely Benign | 0.040 | Likely Benign | -0.85 | Neutral | 0.444 | Benign | 0.071 | Benign | 4.06 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1030 | 0.4792 | 2 | 0 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||||||||
| c.4028A>C | H1343P 2D AIThe SynGAP1 missense variant H1343P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -2.696 | Likely Benign | 0.094 | Likely Benign | Likely Benign | 0.126 | Likely Benign | -1.23 | Neutral | 0.659 | Possibly Damaging | 0.104 | Benign | 4.02 | Benign | 0.00 | Affected | 0.2333 | 0.4220 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||||||||||||
| c.4028A>G | H1343R 2D AIThe SynGAP1 missense variant H1343R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -2.179 | Likely Benign | 0.105 | Likely Benign | Likely Benign | 0.069 | Likely Benign | -1.16 | Neutral | 0.659 | Possibly Damaging | 0.071 | Benign | 4.08 | Benign | 0.00 | Affected | 0.2301 | 0.2602 | 2 | 0 | -1.3 | 19.05 | |||||||||||||||||||||||||||||||||||||||
| c.4028A>T | H1343L 2D AIThe SynGAP1 missense variant H1343L is reported in gnomAD (ID 6‑33451902‑A‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the available predictions points to a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | 6-33451902-A-T | -1.552 | Likely Benign | 0.132 | Likely Benign | Likely Benign | 0.058 | Likely Benign | -1.28 | Neutral | 0.053 | Benign | 0.012 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1222 | 0.5869 | -3 | -2 | 7.0 | -23.98 | ||||||||||||||||||||||||||||||||||||
| c.4029C>A | H1343Q 2D AIThe SynGAP1 missense variant H1343Q is reported in gnomAD (ID 6‑33451903‑C‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. No Foldetta stability result is available. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | 6-33451903-C-A | -2.900 | Likely Benign | 0.111 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -1.04 | Neutral | 0.659 | Possibly Damaging | 0.104 | Benign | 4.06 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2126 | 0.3716 | 0 | 3 | -0.3 | -9.01 | ||||||||||||||||||||||||||||||||||||
| c.4029C>G | H1343Q 2D AIThe SynGAP1 missense variant H1343Q is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that predict a pathogenic effect are polyPhen2_HumDiv and SIFT. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of predictions, including the high‑accuracy tools, suggest that H1343Q is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.983646 | Binding | 0.350 | 0.677 | 0.875 | -2.900 | Likely Benign | 0.111 | Likely Benign | Likely Benign | 0.033 | Likely Benign | -1.04 | Neutral | 0.659 | Possibly Damaging | 0.104 | Benign | 4.06 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2126 | 0.3716 | 0 | 3 | -0.3 | -9.01 | |||||||||||||||||||||||||||||||||||||
| c.402C>A | S134R 2D AIThe SynGAP1 missense variant S134R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict pathogenicity are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized also predicts Pathogenic, while the SGM‑Consensus remains Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -9.053 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.146 | Likely Benign | -2.54 | Deleterious | 0.380 | Benign | 0.147 | Benign | 3.84 | Benign | 0.00 | Affected | 0.0719 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.402C>G | S134R 2D AIThe SynGAP1 missense variant S134R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict pathogenicity are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized also predicts Pathogenic, while the SGM‑Consensus remains Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -9.053 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.146 | Likely Benign | -2.54 | Deleterious | 0.380 | Benign | 0.147 | Benign | 3.84 | Benign | 0.00 | Affected | 0.0719 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.406C>G | R136G 2D AIThe SynGAP1 missense variant R136G is not reported in ClinVar and is absent from gnomAD. Consensus from standard in‑silico predictors shows a split: benign calls come from REVEL, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic calls arise from PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.433034 | Structured | 0.657394 | Binding | 0.351 | 0.894 | 0.250 | -10.641 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.246 | Likely Benign | -3.46 | Deleterious | 0.487 | Possibly Damaging | 0.211 | Benign | 3.47 | Benign | 0.00 | Affected | 0.3581 | 0.3585 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.406C>T | R136W 2D AIThe SynGAP1 missense variant R136W is listed in ClinVar (ID 1479441.0) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts a deleterious effect, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a pathogenic impact, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.433034 | Structured | 0.657394 | Binding | 0.351 | 0.894 | 0.250 | Uncertain | 2 | -10.453 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.237 | Likely Benign | -4.71 | Deleterious | 0.965 | Probably Damaging | 0.416 | Benign | 3.45 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1189 | 0.3827 | 2 | -3 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||||
| c.407G>C | R136P 2D AIThe SynGAP1 missense variant R136P is listed in ClinVar (ID 579340.0) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.433034 | Structured | 0.657394 | Binding | 0.351 | 0.894 | 0.250 | Uncertain | 1 | -11.952 | Likely Pathogenic | 0.981 | Likely Pathogenic | Likely Pathogenic | 0.277 | Likely Benign | -3.72 | Deleterious | 0.910 | Possibly Damaging | 0.578 | Possibly Damaging | 3.47 | Benign | 0.00 | Affected | 3.61 | 5 | 0.2063 | 0.4426 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||
| c.409C>A | L137I 2D AIThe SynGAP1 missense variant L137I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the majority of predictions (5 pathogenic vs. 3 benign) indicate a likely pathogenic impact. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.553315 | Disordered | 0.639549 | Binding | 0.377 | 0.897 | 0.375 | -8.870 | Likely Pathogenic | 0.914 | Likely Pathogenic | Ambiguous | 0.113 | Likely Benign | -1.22 | Neutral | 0.993 | Probably Damaging | 0.967 | Probably Damaging | 3.73 | Benign | 0.00 | Affected | 0.0932 | 0.3102 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||||
| c.409C>G | L137V 2D AIThe SynGAP1 missense variant L137V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictors (six out of nine) indicate a pathogenic impact, whereas three predict benign. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.553315 | Disordered | 0.639549 | Binding | 0.377 | 0.897 | 0.375 | -8.621 | Likely Pathogenic | 0.969 | Likely Pathogenic | Likely Pathogenic | 0.118 | Likely Benign | -1.75 | Neutral | 0.993 | Probably Damaging | 0.967 | Probably Damaging | 3.73 | Benign | 0.00 | Affected | 0.1533 | 0.2612 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.40C>A | P14T 2D AIThe SynGAP1 missense variant P14T is reported in gnomAD (ID 6‑33420304‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also reports likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for P14T, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | 6-33420304-C-A | -3.261 | Likely Benign | 0.150 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -0.27 | Neutral | 0.212 | Benign | 0.014 | Benign | 4.20 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1912 | 0.6658 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.40C>G | P14A 2D AIThe SynGAP1 missense variant P14A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign status: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. Foldetta results are unavailable. Overall, the majority of computational evidence indicates that P14A is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | -2.919 | Likely Benign | 0.096 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -0.01 | Neutral | 0.100 | Benign | 0.007 | Benign | 4.24 | Benign | 0.00 | Affected | 0.3928 | 0.5543 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.40C>T | P14S 2D AIThe SynGAP1 missense variant P14S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | -2.352 | Likely Benign | 0.123 | Likely Benign | Likely Benign | 0.068 | Likely Benign | -0.02 | Neutral | 0.212 | Benign | 0.014 | Benign | 4.23 | Benign | 0.00 | Affected | 0.3935 | 0.5938 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.410T>A | L137Q 2D AIThe SynGAP1 missense variant L137Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM. All other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled “Likely Pathogenic.” The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.553315 | Disordered | 0.639549 | Binding | 0.377 | 0.897 | 0.375 | -12.246 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.356 | Likely Benign | -3.43 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 3.60 | Benign | 0.00 | Affected | 0.1137 | 0.1049 | -2 | -2 | -7.3 | 14.97 | |||||||||||||||||||||||||||||||||||||||
| c.410T>C | L137P 2D AIThe SynGAP1 missense variant L137P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus indicates it is likely pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.553315 | Disordered | 0.639549 | Binding | 0.377 | 0.897 | 0.375 | -11.067 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.377 | Likely Benign | -3.77 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 3.59 | Benign | 0.00 | Affected | 0.3620 | 0.1177 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||||||
| c.410T>G | L137R 2D AIThe SynGAP1 missense variant L137R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM. All other evaluated tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.553315 | Disordered | 0.639549 | Binding | 0.377 | 0.897 | 0.375 | -11.595 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.334 | Likely Benign | -3.50 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | 3.60 | Benign | 0.00 | Affected | 0.1300 | 0.0691 | -3 | -2 | -8.3 | 43.03 | |||||||||||||||||||||||||||||||||||||||
| c.413A>T | K138I 2D AIThe SynGAP1 missense variant K138I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.590140 | Disordered | 0.619482 | Binding | 0.349 | 0.901 | 0.375 | -9.366 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.295 | Likely Benign | -4.74 | Deleterious | 0.535 | Possibly Damaging | 0.259 | Benign | 3.53 | Benign | 0.00 | Affected | 0.1003 | 0.3054 | -2 | -3 | 8.4 | -15.01 | |||||||||||||||||||||||||||||||||||||||
| c.419C>A | S140Y 2D AIThe SynGAP1 missense variant S140Y is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default—predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the S140Y variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.587898 | Binding | 0.321 | 0.896 | 0.625 | -13.071 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.252 | Likely Benign | -3.88 | Deleterious | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 3.48 | Benign | 0.00 | Affected | 0.0597 | 0.5849 | -3 | -2 | -0.5 | 76.10 | |||||||||||||||||||||||||||||||||||||||
| c.419C>T | S140F 2D AIThe SynGAP1 missense variant S140F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic or likely pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports “Likely Pathogenic.” High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote) also indicates likely pathogenic. Foldetta results are unavailable. Overall, the consensus of the available predictions points to a pathogenic effect for S140F, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.587898 | Binding | 0.321 | 0.896 | 0.625 | -10.824 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.246 | Likely Benign | -3.83 | Deleterious | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 3.48 | Benign | 0.00 | Affected | 0.0565 | 0.5988 | -3 | -2 | 3.6 | 60.10 | |||||||||||||||||||||||||||||||||||||||
| c.41C>A | P14H 2D AIThe SynGAP1 missense variant P14H is listed in gnomAD (ID 6‑33420305‑C‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Overall, the majority of predictions—including the high‑accuracy tools—indicate that P14H is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | 6-33420305-C-A | -3.747 | Likely Benign | 0.231 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -0.27 | Neutral | 0.742 | Possibly Damaging | 0.091 | Benign | 4.15 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1999 | 0.5176 | -2 | 0 | -1.6 | 40.02 | ||||||||||||||||||||||||||||||||||||
| c.41C>G | P14R 2D AIThe SynGAP1 missense variant P14R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The high‑accuracy consensus methods give a benign verdict: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” No result is available from Foldetta, so its folding‑stability assessment is not considered. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of any ClinVar classification. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | -2.794 | Likely Benign | 0.253 | Likely Benign | Likely Benign | 0.183 | Likely Benign | 0.13 | Neutral | 0.486 | Possibly Damaging | 0.032 | Benign | 4.20 | Benign | 0.00 | Affected | 0.1597 | 0.3536 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.41C>T | P14L 2D AIThe SynGAP1 missense variant P14L is catalogued in gnomAD (ID 6‑33420305‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes, while the single pathogenic call comes from SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise favors benign; Foldetta results are unavailable. Overall, the consensus of available predictions points to a benign impact for P14L, and this conclusion is not contradicted by ClinVar status, which currently lacks an entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | 6-33420305-C-T | -3.277 | Likely Benign | 0.332 | Likely Benign | Likely Benign | 0.153 | Likely Benign | -0.53 | Neutral | 0.062 | Benign | 0.004 | Benign | 4.19 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2618 | 0.7197 | -3 | -3 | 5.4 | 16.04 | ||||||||||||||||||||||||||||||||||||
| c.422T>A | I141N 2D AIThe SynGAP1 missense variant I141N is not reported in ClinVar and is absent from gnomAD. Consensus from standard prediction algorithms shows a split: benign predictions come from REVEL, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic. Foldetta stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic impact for I141N. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.465241 | Structured | 0.577021 | Binding | 0.367 | 0.877 | 0.500 | -12.417 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.258 | Likely Benign | -3.90 | Deleterious | 0.799 | Possibly Damaging | 0.424 | Benign | 3.54 | Benign | 0.00 | Affected | 0.1047 | 0.0270 | -2 | -3 | -8.0 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.422T>C | I141T 2D AIThe SynGAP1 missense variant I141T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its contribution is unavailable. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.465241 | Structured | 0.577021 | Binding | 0.367 | 0.877 | 0.500 | -10.580 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.251 | Likely Benign | -2.81 | Deleterious | 0.272 | Benign | 0.167 | Benign | 3.57 | Benign | 0.00 | Affected | 0.1165 | 0.1214 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||||||||||||
| c.422T>G | I141S 2D AIThe SynGAP1 missense variant I141S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus also as Likely Pathogenic; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic impact for I141S, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.465241 | Structured | 0.577021 | Binding | 0.367 | 0.877 | 0.500 | -11.874 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.267 | Likely Benign | -3.28 | Deleterious | 0.567 | Possibly Damaging | 0.249 | Benign | 3.56 | Benign | 0.00 | Affected | 0.2885 | 0.0840 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.424A>C | K142Q 2D AIThe SynGAP1 missense variant K142Q is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. No Foldetta stability analysis is available for this variant. Overall, the preponderance of computational evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -11.295 | Likely Pathogenic | 0.979 | Likely Pathogenic | Likely Pathogenic | 0.191 | Likely Benign | -2.50 | Deleterious | 0.700 | Possibly Damaging | 0.383 | Benign | 3.50 | Benign | 0.00 | Affected | 0.4232 | 0.1132 | 1 | 1 | 0.4 | -0.04 | |||||||||||||||||||||||||||||||||||||||
| c.424A>G | K142E 2D AIThe SynGAP1 missense variant K142E is not reported in ClinVar (no ClinVar entry) and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions point to a pathogenic effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -14.450 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.249 | Likely Benign | -2.59 | Deleterious | 0.247 | Benign | 0.125 | Benign | 3.49 | Benign | 0.00 | Affected | 0.3644 | 0.0952 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.425A>C | K142T 2D AIThe SynGAP1 missense variant K142T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -12.013 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.251 | Likely Benign | -3.77 | Deleterious | 0.247 | Benign | 0.166 | Benign | 3.47 | Benign | 0.00 | Affected | 0.2034 | 0.2708 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.425A>G | K142R 2D AIThe SynGAP1 missense variant K142R is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also favors benign (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -9.327 | Likely Pathogenic | 0.361 | Ambiguous | Likely Benign | 0.157 | Likely Benign | -1.87 | Neutral | 0.399 | Benign | 0.212 | Benign | 3.65 | Benign | 0.00 | Affected | 0.4459 | 0.0858 | 3 | 2 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||||||||
| c.425A>T | K142I 2D AIThe SynGAP1 missense variant K142I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus also indicates Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -14.597 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.296 | Likely Benign | -4.81 | Deleterious | 0.700 | Possibly Damaging | 0.403 | Benign | 3.44 | Benign | 0.00 | Affected | 0.1005 | 0.3173 | -2 | -3 | 8.4 | -15.01 | |||||||||||||||||||||||||||||||||||||||
| c.426A>C | K142N 2D AIThe SynGAP1 missense variant K142N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -12.169 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.097 | Likely Benign | -3.13 | Deleterious | 0.700 | Possibly Damaging | 0.383 | Benign | 3.47 | Benign | 0.00 | Affected | 0.3586 | 0.1437 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.426A>T | K142N 2D AIThe SynGAP1 missense variant K142N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.461924 | Structured | 0.558796 | Binding | 0.374 | 0.859 | 0.500 | -12.169 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 0.096 | Likely Benign | -3.13 | Deleterious | 0.700 | Possibly Damaging | 0.383 | Benign | 3.47 | Benign | 0.00 | Affected | 0.3586 | 0.1437 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.427C>G | R143G 2D AIThe SynGAP1 missense variant R143G is not reported in ClinVar and is absent from gnomAD. In silico predictors that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Predictors that agree on a pathogenic effect include PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the preponderance of evidence from multiple independent tools points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, as no ClinVar annotation exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.575842 | Disordered | 0.538584 | Binding | 0.338 | 0.838 | 0.625 | -12.686 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.151 | Likely Benign | -3.86 | Deleterious | 0.319 | Benign | 0.124 | Benign | 3.51 | Benign | 0.00 | Affected | 0.3502 | 0.3547 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.428G>A | R143Q 2D AIThe SynGAP1 missense variant R143Q is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33432725‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs 2 benign), and Foldetta stability analysis is unavailable. Overall, the majority of predictions (5 pathogenic vs 4 benign) and the pathogenic call from AlphaMissense‑Optimized suggest that the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.575842 | Disordered | 0.538584 | Binding | 0.338 | 0.838 | 0.625 | 6-33432725-G-A | 2 | 1.35e-6 | -12.110 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.181 | Likely Benign | -2.48 | Neutral | 0.678 | Possibly Damaging | 0.176 | Benign | 3.53 | Benign | 0.00 | Affected | 3.61 | 5 | 0.3255 | 0.2678 | 1 | 1 | 1.0 | -28.06 | |||||||||||||||||||||||||||||||||||
| c.428G>C | R143P 2D AIThe SynGAP1 missense variant R143P is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33432725‑G‑C). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as Likely Pathogenic; the Foldetta stability analysis is unavailable. Overall, the majority of computational evidence points to a pathogenic impact, and this is consistent with the ClinVar designation of “Uncertain” rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.575842 | Disordered | 0.538584 | Binding | 0.338 | 0.838 | 0.625 | Uncertain | 2 | 6-33432725-G-C | 2 | 1.35e-6 | -14.564 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.292 | Likely Benign | -3.74 | Deleterious | 0.001 | Benign | 0.000 | Benign | 3.49 | Benign | 0.00 | Affected | 3.61 | 5 | 0.2063 | 0.4457 | -2 | 0 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||
| c.428G>T | R143L 2D AIThe SynGAP1 missense variant R143L is listed in ClinVar with no submitted interpretation and is present in gnomAD (ID 6‑33432725‑G‑T). Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic; the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.575842 | Disordered | 0.538584 | Binding | 0.338 | 0.838 | 0.625 | 6-33432725-G-T | 1 | 6.77e-7 | -14.250 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.316 | Likely Benign | -4.37 | Deleterious | 0.319 | Benign | 0.124 | Benign | 3.52 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1758 | 0.4886 | -2 | -3 | 8.3 | -43.03 | ||||||||||||||||||||||||||||||||||
| c.430A>C | T144P 2D AIThe SynGAP1 missense variant T144P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict pathogenicity are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely pathogenic, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of predictions and the consensus call indicate a pathogenic effect. This conclusion is not contradicted by ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | -11.920 | Likely Pathogenic | 0.775 | Likely Pathogenic | Likely Benign | 0.159 | Likely Benign | -2.66 | Deleterious | 0.000 | Benign | 0.000 | Benign | 3.76 | Benign | 0.00 | Affected | 0.2508 | 0.5271 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||||||||||||
| c.430A>G | T144A 2D AIThe SynGAP1 missense variant T144A is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also reports a benign prediction. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | -4.007 | Likely Benign | 0.440 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -2.24 | Neutral | 0.000 | Benign | 0.001 | Benign | 3.95 | Benign | 0.00 | Affected | 0.4777 | 0.4131 | 1 | 0 | 2.5 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.430A>T | T144S 2D AIThe SynGAP1 missense variant T144S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default, while ESM1b remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for T144S. This conclusion does not contradict ClinVar, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | -7.730 | In-Between | 0.672 | Likely Pathogenic | Likely Benign | 0.081 | Likely Benign | -1.90 | Neutral | 0.018 | Benign | 0.016 | Benign | 3.84 | Benign | 0.00 | Affected | 0.4039 | 0.4183 | 1 | 1 | -0.1 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.431C>A | T144K 2D AIThe SynGAP1 T144K variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a pathogenic impact, and this assessment does not contradict any ClinVar status because none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | -15.436 | Likely Pathogenic | 0.956 | Likely Pathogenic | Likely Pathogenic | 0.164 | Likely Benign | -2.83 | Deleterious | 0.180 | Benign | 0.063 | Benign | 3.76 | Benign | 0.00 | Affected | 0.1462 | 0.3179 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||||||||||||
| c.431C>G | T144R 2D AIThe SynGAP1 missense variant T144R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign calls come from REVEL, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic effect. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the aggregate predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | -13.331 | Likely Pathogenic | 0.922 | Likely Pathogenic | Ambiguous | 0.177 | Likely Benign | -2.83 | Deleterious | 0.609 | Possibly Damaging | 0.150 | Benign | 3.75 | Benign | 0.00 | Affected | 0.1188 | 0.2823 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.431C>T | T144M 2D AIThe SynGAP1 missense variant T144M is listed in ClinVar with an “Uncertain” status (ClinVar ID 2231966.0) and is present in the gnomAD database (gnomAD ID 6‑33432728‑C‑T). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Pathogenic.” High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain,” SGM‑Consensus as “Likely Pathogenic,” and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of computational predictions lean toward a pathogenic impact, and this assessment does not contradict the ClinVar designation of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | Uncertain | 2 | 6-33432728-C-T | 2 | 1.30e-6 | -11.228 | Likely Pathogenic | 0.922 | Likely Pathogenic | Ambiguous | 0.118 | Likely Benign | -3.16 | Deleterious | 0.913 | Possibly Damaging | 0.333 | Benign | 3.73 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1504 | 0.6510 | -1 | -1 | 2.6 | 30.09 | ||||||||||||||||||||||||||||||||
| c.433A>C | K145Q 2D AIThe SynGAP1 missense variant K145Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Computational predictions are split: benign calls come from REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls come from polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy tools give no definitive verdict: the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie, and Foldetta results are unavailable. Consequently, the variant is neither clearly benign nor pathogenic according to current predictions, and there is no ClinVar annotation to contradict this ambiguous computational assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -9.676 | Likely Pathogenic | 0.955 | Likely Pathogenic | Ambiguous | 0.163 | Likely Benign | -2.34 | Neutral | 0.700 | Possibly Damaging | 0.383 | Benign | 3.65 | Benign | 0.00 | Affected | 0.4233 | 0.1478 | 1 | 1 | 0.4 | -0.04 | ||||||||||||||||||||||||||||||||||||||||
| c.433A>G | K145E 2D AIThe SynGAP1 missense variant K145E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions arise from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of high‑confidence tools predict a pathogenic impact, and this conclusion does not contradict any ClinVar annotation (none is present). Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -12.571 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.245 | Likely Benign | -2.51 | Deleterious | 0.247 | Benign | 0.125 | Benign | 3.68 | Benign | 0.00 | Affected | 0.3638 | 0.1179 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||||||||||||
| c.434A>C | K145T 2D AIThe SynGAP1 missense variant K145T is not reported in ClinVar and is absent from gnomAD. Computational assessment shows a split: benign predictions from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, whereas pathogenic predictions come from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN, classifies the change as Likely Pathogenic. High‑accuracy tools further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus itself is a high‑confidence prediction; Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic impact for K145T. Thus, the variant is most likely pathogenic, and there is no ClinVar annotation to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -8.519 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 0.224 | Likely Benign | -3.54 | Deleterious | 0.247 | Benign | 0.166 | Benign | 3.63 | Benign | 0.00 | Affected | 0.1965 | 0.3611 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.434A>G | K145R 2D AIThe SynGAP1 missense variant K145R is listed in gnomAD (ID 6‑33432731‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify the change as benign or likely benign. Only SIFT predicts a pathogenic outcome, while ESM1b remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status, as none is currently assigned to the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | 6-33432731-A-G | 1 | 6.20e-7 | -7.685 | In-Between | 0.214 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -1.58 | Neutral | 0.399 | Benign | 0.212 | Benign | 3.71 | Benign | 0.00 | Affected | 3.61 | 5 | 0.4661 | 0.1242 | 2 | 3 | -0.6 | 28.01 | ||||||||||||||||||||||||||||||||||
| c.434A>T | K145M 2D AIThe SynGAP1 missense variant K145M is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence indicates that K145M is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -9.884 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.245 | Likely Benign | -3.47 | Deleterious | 0.964 | Probably Damaging | 0.650 | Possibly Damaging | 3.59 | Benign | 0.00 | Affected | 0.1041 | 0.4427 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||||||||||||
| c.435G>C | K145N 2D AIThe SynGAP1 missense variant K145N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as Likely Pathogenic; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic impact for K145N, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -8.718 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.109 | Likely Benign | -2.76 | Deleterious | 0.700 | Possibly Damaging | 0.383 | Benign | 3.65 | Benign | 0.00 | Affected | 0.3430 | 0.1832 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.435G>T | K145N 2D AIThe SynGAP1 missense variant K145N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL (score 0.45), polyPhen‑2 HumVar (benign), and FATHMM (benign). Pathogenic predictions arise from PROVEAN (deleterious), polyPhen‑2 HumDiv (probably damaging), SIFT (deleterious), ESM1b (damaging), AlphaMissense‑Default (pathogenic), and AlphaMissense‑Optimized (pathogenic). The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic effect, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.671169 | Disordered | 0.516174 | Binding | 0.321 | 0.835 | 0.625 | -8.718 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.109 | Likely Benign | -2.76 | Deleterious | 0.700 | Possibly Damaging | 0.383 | Benign | 3.65 | Benign | 0.00 | Affected | 0.3430 | 0.1832 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.436T>A | S146T 2D AIThe SynGAP1 missense variant S146T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a tie (two pathogenic, two benign) and thus unavailable; Foldetta results are not provided. Overall, the majority of evidence (five benign vs. three pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.541878 | Disordered | 0.508612 | Binding | 0.349 | 0.837 | 0.625 | -9.795 | Likely Pathogenic | 0.786 | Likely Pathogenic | Ambiguous | 0.113 | Likely Benign | -2.09 | Neutral | 0.084 | Benign | 0.042 | Benign | 3.65 | Benign | 0.00 | Affected | 0.1267 | 0.5075 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.436T>C | S146P 2D AIThe SynGAP1 missense variant S146P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions arise from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates likely pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for S146P, and this conclusion is not contradicted by ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.541878 | Disordered | 0.508612 | Binding | 0.349 | 0.837 | 0.625 | -13.292 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.215 | Likely Benign | -3.11 | Deleterious | 0.392 | Benign | 0.230 | Benign | 3.60 | Benign | 0.00 | Affected | 0.1934 | 0.4627 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.436T>G | S146A 2D AIThe SynGAP1 missense variant S146A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.541878 | Disordered | 0.508612 | Binding | 0.349 | 0.837 | 0.625 | -14.042 | Likely Pathogenic | 0.721 | Likely Pathogenic | Likely Benign | 0.097 | Likely Benign | -1.81 | Neutral | 0.000 | Benign | 0.002 | Benign | 3.71 | Benign | 0.00 | Affected | 0.4435 | 0.3708 | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.437C>T | S146L 2D  AIThe SynGAP1 missense variant S146L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) which classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are not available. Overall, the majority of predictions indicate a pathogenic impact, and this is consistent with the lack of ClinVar annotation—there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.541878 | Disordered | 0.508612 | Binding | 0.349 | 0.837 | 0.625 | -15.179 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.130 | Likely Benign | -3.77 | Deleterious | 0.084 | Benign | 0.063 | Benign | 3.63 | Benign | 0.00 | Affected | 0.0856 | 0.4975 | -3 | -2 | 4.6 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.43G>A | A15T 2D AIThe SynGAP1 missense variant A15T is listed in ClinVar (ID 1925632.0) with an “Uncertain” clinical significance and is present in gnomAD (6‑33420307‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as “Likely Benign”; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | Uncertain | 1 | 6-33420307-G-A | 4 | 2.60e-6 | -3.720 | Likely Benign | 0.125 | Likely Benign | Likely Benign | 0.086 | Likely Benign | -0.08 | Neutral | 0.602 | Possibly Damaging | 0.017 | Benign | 4.16 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2053 | 0.7423 | 1 | 0 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||||
| c.43G>C | A15P 2D AIThe SynGAP1 missense variant A15P is listed in ClinVar (ID 3688743.0) with an *Uncertain* clinical significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as *Likely Benign*. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | Uncertain | 1 | -3.436 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.146 | Likely Benign | -0.23 | Neutral | 0.880 | Possibly Damaging | 0.123 | Benign | 4.09 | Benign | 0.00 | Affected | 0.2573 | 0.6988 | 1 | -1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||||
| c.43G>T | A15S 2D AIThe SynGAP1 missense variant A15S is reported in ClinVar as “Not submitted” and is present in gnomAD (variant ID 6-33420307‑G‑T). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign effect for A15S, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | 6-33420307-G-T | -2.925 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.074 | Likely Benign | 0.11 | Neutral | 0.122 | Benign | 0.010 | Benign | 4.17 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3115 | 0.5830 | 1 | 1 | -2.6 | 16.00 | ||||||||||||||||||||||||||||||||||||
| c.443C>A | P148H 2D AIThe SynGAP1 missense variant P148H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple in silico predictors points to a pathogenic effect for P148H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.653063 | Disordered | 0.500109 | Binding | 0.372 | 0.837 | 0.625 | -11.034 | Likely Pathogenic | 0.961 | Likely Pathogenic | Likely Pathogenic | 0.169 | Likely Benign | -3.21 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.90 | Benign | 0.00 | Affected | 0.1881 | 0.3847 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.445A>C | K149Q 2D AIThe SynGAP1 missense variant K149Q is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2). AlphaMissense‑Optimized predicts pathogenic, while Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the balance of evidence (five pathogenic vs. four benign predictions) indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -11.430 | Likely Pathogenic | 0.980 | Likely Pathogenic | Likely Pathogenic | 0.190 | Likely Benign | -1.99 | Neutral | 0.535 | Possibly Damaging | 0.310 | Benign | 3.61 | Benign | 0.00 | Affected | 0.5243 | 0.1454 | Weaken | 1 | 1 | 0.4 | -0.04 | |||||||||||||||||||||||||||||||||||||||
| c.445A>G | K149E 2D AIThe SynGAP1 missense variant K149E is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields an equal split (2 vs 2) and is therefore considered unavailable. AlphaMissense‑Optimized, a high‑accuracy predictor, classifies the variant as pathogenic, while Foldetta (combining FoldX‑MD and Rosetta) has no available result for this variant. Overall, the majority of standard tools (five benign vs. four pathogenic) lean toward a benign interpretation, and this is consistent with the lack of ClinVar evidence. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -12.148 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.231 | Likely Benign | -2.17 | Neutral | 0.141 | Benign | 0.062 | Benign | 3.59 | Benign | 0.00 | Affected | 0.4636 | 0.1022 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.446A>C | K149T 2D AIThe SynGAP1 missense variant K149T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar), and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta results are unavailable, so no stability evidence is provided. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -11.948 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.260 | Likely Benign | -3.53 | Deleterious | 0.141 | Benign | 0.091 | Benign | 3.56 | Benign | 0.00 | Affected | 0.2398 | 0.3473 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||||||||||||
| c.446A>G | K149R 2D AIThe SynGAP1 missense variant K149R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that K149R is most likely benign, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -8.237 | Likely Pathogenic | 0.406 | Ambiguous | Likely Benign | 0.143 | Likely Benign | -1.70 | Neutral | 0.247 | Benign | 0.125 | Benign | 3.70 | Benign | 0.00 | Affected | 0.5639 | 0.1169 | Weaken | 3 | 2 | -0.6 | 28.01 | |||||||||||||||||||||||||||||||||||||||
| c.446A>T | K149I 2D AIThe SynGAP1 missense variant K149I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; the Foldetta protein‑folding stability analysis is unavailable. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -14.426 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.305 | Likely Benign | -4.72 | Deleterious | 0.535 | Possibly Damaging | 0.403 | Benign | 3.53 | Benign | 0.00 | Affected | 0.1477 | 0.3965 | -2 | -3 | 8.4 | -15.01 | |||||||||||||||||||||||||||||||||||||||
| c.447A>C | K149N 2D AIThe SynGAP1 missense variant K149N is not reported in ClinVar and has no entry in gnomAD, indicating it is not catalogued in these databases. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a majority pathogenic vote (3 pathogenic vs. 1 benign) and is labeled “Likely Pathogenic.” AlphaMissense‑Optimized independently predicts pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, is not available for this variant. Overall, the majority of computational evidence points to a pathogenic impact, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -9.275 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.121 | Likely Benign | -2.85 | Deleterious | 0.141 | Benign | 0.123 | Benign | 3.56 | Benign | 0.00 | Affected | 0.4427 | 0.1559 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.447A>T | K149N 2D AIThe SynGAP1 missense variant K149N is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions arise from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also indicates likely pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect for K149N, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.562014 | Disordered | 0.501681 | Binding | 0.302 | 0.839 | 0.625 | -9.275 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.121 | Likely Benign | -2.85 | Deleterious | 0.141 | Benign | 0.123 | Benign | 3.56 | Benign | 0.00 | Affected | 0.4427 | 0.1559 | 1 | 0 | 0.4 | -14.07 | |||||||||||||||||||||||||||||||||||||||
| c.448C>A | L150I 2D AIThe SynGAP1 missense variant L150I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments are less definitive: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign), and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) indicate a likely pathogenic impact for the variant. This conclusion is not contradicted by ClinVar status, as the variant has no existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.494003 | Structured | 0.505752 | Binding | 0.299 | 0.839 | 0.625 | -10.118 | Likely Pathogenic | 0.886 | Likely Pathogenic | Ambiguous | 0.080 | Likely Benign | -1.25 | Neutral | 0.993 | Probably Damaging | 0.967 | Probably Damaging | 3.74 | Benign | 0.00 | Affected | 0.0894 | 0.3468 | 2 | 2 | 0.7 | 0.00 | ||||||||||||||||||||||||||||||||||||||||
| c.448C>G | L150V 2D AIThe SynGAP1 missense variant L150V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool returns an uncertain result, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (5 pathogenic vs. 3 benign) indicate a pathogenic effect. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.494003 | Structured | 0.505752 | Binding | 0.299 | 0.839 | 0.625 | -10.375 | Likely Pathogenic | 0.927 | Likely Pathogenic | Ambiguous | 0.113 | Likely Benign | -1.84 | Neutral | 0.993 | Probably Damaging | 0.967 | Probably Damaging | 3.74 | Benign | 0.00 | Affected | 0.1473 | 0.3178 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.448C>T | L150F 2D AIThe SynGAP1 missense variant L150F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts Pathogenic, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta results are not available for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and the SGM‑Consensus suggests that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.494003 | Structured | 0.505752 | Binding | 0.299 | 0.839 | 0.625 | -12.459 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.105 | Likely Benign | -2.56 | Deleterious | 0.998 | Probably Damaging | 0.991 | Probably Damaging | 3.69 | Benign | 0.00 | Affected | 0.0580 | 0.3112 | 2 | 0 | -1.0 | 34.02 | |||||||||||||||||||||||||||||||||||||||
| c.449T>A | L150H 2D AIThe SynGAP1 missense variant L150H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic (3 pathogenic vs. 1 benign). AlphaMissense‑Optimized alone also predicts Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple independent predictors indicates that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.494003 | Structured | 0.505752 | Binding | 0.299 | 0.839 | 0.625 | -14.892 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.303 | Likely Benign | -4.09 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 3.64 | Benign | 0.00 | Affected | 0.1011 | 0.0519 | -2 | -3 | -7.0 | 23.98 | |||||||||||||||||||||||||||||||||||||||
| c.449T>C | L150P 2D AIThe SynGAP1 missense variant L150P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus indicates it is likely pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.494003 | Structured | 0.505752 | Binding | 0.299 | 0.839 | 0.625 | -12.881 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.320 | Likely Benign | -3.90 | Deleterious | 0.998 | Probably Damaging | 0.995 | Probably Damaging | 3.64 | Benign | 0.00 | Affected | 0.3557 | 0.1543 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||||||
| c.449T>G | L150R 2D AIThe SynGAP1 missense variant L150R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) all indicate pathogenicity. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from multiple in silico predictors and high‑accuracy tools points to a pathogenic effect for the SynGAP1 L150R variant, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.494003 | Structured | 0.505752 | Binding | 0.299 | 0.839 | 0.625 | -14.879 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.290 | Likely Benign | -3.16 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | 3.67 | Benign | 0.00 | Affected | 0.1275 | 0.0719 | -3 | -2 | -8.3 | 43.03 | |||||||||||||||||||||||||||||||||||||||
| c.44C>A | A15E 2D  AIThe SynGAP1 missense variant A15E is reported in gnomAD (ID 6‑33420308‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | 6-33420308-C-A | -3.423 | Likely Benign | 0.277 | Likely Benign | Likely Benign | 0.169 | Likely Benign | 0.46 | Neutral | 0.406 | Benign | 0.040 | Benign | 4.13 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1563 | 0.1908 | -1 | 0 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||
| c.44C>G | A15G 2D AIThe SynGAP1 missense variant A15G is reported in gnomAD (ID 6‑33420308‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also reports likely benign; Foldetta results are not available. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | 6-33420308-C-G | 3 | 1.95e-6 | -3.261 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.084 | Likely Benign | -0.04 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.12 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2753 | 0.5196 | 0 | 1 | -2.2 | -14.03 | ||||||||||||||||||||||||||||||||||
| c.44C>T | A15V 2D AIThe SynGAP1 missense variant A15V is listed in ClinVar (ID 1801174.0) with an “Uncertain” status and is present in gnomAD (6‑33420308‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while polyPhen‑2 HumDiv and SIFT predict a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this does not contradict the ClinVar designation, which remains uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | Uncertain | 1 | 6-33420308-C-T | 1 | 6.49e-7 | -3.560 | Likely Benign | 0.161 | Likely Benign | Likely Benign | 0.105 | Likely Benign | 0.20 | Neutral | 0.602 | Possibly Damaging | 0.015 | Benign | 4.19 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1809 | 0.7669 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||
| c.451G>C | D151H 2D AIThe SynGAP1 missense variant D151H is listed in ClinVar with an “Uncertain” status and is present in gnomAD (variant ID 6‑33432748‑G‑C). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as “Likely Pathogenic.” No Foldetta stability result is available for this variant. Overall, the majority of computational evidence indicates that D151H is most likely pathogenic, which does not contradict the current ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.529623 | Disordered | 0.503277 | Binding | 0.342 | 0.841 | 0.625 | Uncertain | 1 | 6-33432748-G-C | 2 | 1.26e-6 | -11.747 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.335 | Likely Benign | -3.90 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 3.86 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1543 | 0.8419 | -1 | 1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||
| c.451G>T | D151Y 2D AIThe SynGAP1 D151Y variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labels it as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the D151Y variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.529623 | Disordered | 0.503277 | Binding | 0.342 | 0.841 | 0.625 | -13.174 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.340 | Likely Benign | -5.16 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 3.85 | Benign | 0.00 | Affected | 0.0567 | 0.7560 | -4 | -3 | 2.2 | 48.09 | |||||||||||||||||||||||||||||||||||||||
| c.452A>T | D151V 2D AIThe SynGAP1 D151V variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Pathogenic” based on a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is likely pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from multiple in silico predictors indicates that D151V is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.529623 | Disordered | 0.503277 | Binding | 0.342 | 0.841 | 0.625 | -11.927 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.380 | Likely Benign | -5.19 | Deleterious | 0.998 | Probably Damaging | 0.994 | Probably Damaging | 3.88 | Benign | 0.00 | Affected | 0.0864 | 0.7781 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.454C>G | R152G 2D AIThe SynGAP1 missense variant R152G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that R152G is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.618285 | Disordered | 0.500158 | Binding | 0.319 | 0.842 | 0.625 | -11.043 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.169 | Likely Benign | -4.13 | Deleterious | 0.993 | Probably Damaging | 0.982 | Probably Damaging | 3.83 | Benign | 0.00 | Affected | 0.3633 | 0.3956 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.454C>T | R152W 2D AIThe SynGAP1 missense variant R152W is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.618285 | Disordered | 0.500158 | Binding | 0.319 | 0.842 | 0.625 | -11.783 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.163 | Likely Benign | -4.66 | Deleterious | 1.000 | Probably Damaging | 0.990 | Probably Damaging | 3.79 | Benign | 0.00 | Affected | 0.1550 | 0.4564 | 2 | -3 | 3.6 | 30.03 | |||||||||||||||||||||||||||||||||||||||
| c.455G>A | R152Q 2D AIThe SynGAP1 missense variant R152Q is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33432752‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic, two benign). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus remains unavailable, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no reported result for this variant. Overall, the preponderance of evidence (seven pathogenic versus three benign predictions) indicates that the variant is most likely pathogenic, which does not contradict the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.618285 | Disordered | 0.500158 | Binding | 0.319 | 0.842 | 0.625 | Uncertain | 1 | 6-33432752-G-A | 5 | 3.14e-6 | -10.336 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.181 | Likely Benign | -2.34 | Neutral | 0.997 | Probably Damaging | 0.968 | Probably Damaging | 3.89 | Benign | 0.00 | Affected | 3.61 | 5 | 0.3618 | 0.2996 | 1 | 1 | 1.0 | -28.06 | |||||||||||||||||||||||||||||||||
| c.455G>C | R152P 2D AIThe SynGAP1 missense variant R152P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic outcome. The SGM‑Consensus, a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is also pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as pathogenic; Foldetta results are unavailable. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.618285 | Disordered | 0.500158 | Binding | 0.319 | 0.842 | 0.625 | -11.499 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.332 | Likely Benign | -4.00 | Deleterious | 0.998 | Probably Damaging | 0.992 | Probably Damaging | 3.82 | Benign | 0.00 | Affected | 0.2302 | 0.4838 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.455G>T | R152L 2D AIThe SynGAP1 missense variant R152L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is also pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence indicates that R152L is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.618285 | Disordered | 0.500158 | Binding | 0.319 | 0.842 | 0.625 | -11.501 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.331 | Likely Benign | -3.85 | Deleterious | 0.993 | Probably Damaging | 0.982 | Probably Damaging | 3.83 | Benign | 0.00 | Affected | 0.2032 | 0.5249 | -3 | -2 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||||
| c.463A>C | S155R 2D AIThe SynGAP1 missense variant S155R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default—predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the S155R variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -11.939 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.250 | Likely Benign | -2.74 | Deleterious | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.88 | Benign | 0.00 | Affected | 0.0712 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.463A>G | S155G 2D AIThe SynGAP1 missense variant S155G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic, two benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools specifically indicate a benign outcome from AlphaMissense‑Optimized, while SGM Consensus and Foldetta are unavailable. Overall, the balance of evidence leans toward a pathogenic interpretation, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -9.243 | Likely Pathogenic | 0.628 | Likely Pathogenic | Likely Benign | 0.152 | Likely Benign | -1.84 | Neutral | 0.956 | Probably Damaging | 0.931 | Probably Damaging | 3.81 | Benign | 0.00 | Affected | 0.2416 | 0.3976 | 1 | 0 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||||||||||||
| c.463A>T | S155C 2D AIThe SynGAP1 missense variant S155C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, while the SGM‑Consensus remains Likely Pathogenic; Foldetta results are unavailable. Overall, the majority of predictions lean toward pathogenicity, but this conclusion conflicts with the benign prediction from AlphaMissense‑Optimized and the lack of ClinVar evidence. Thus, the variant is most likely pathogenic based on the prevailing predictions, though the evidence is not unequivocal. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -11.903 | Likely Pathogenic | 0.756 | Likely Pathogenic | Likely Benign | 0.238 | Likely Benign | -2.53 | Deleterious | 0.999 | Probably Damaging | 0.990 | Probably Damaging | 3.78 | Benign | 0.00 | Affected | 0.0826 | 0.5681 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.464G>A | S155N 2D AIThe SynGAP1 missense variant S155N is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, while those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give mixed or unavailable results: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no reported result. Overall, the majority of available predictions (five pathogenic vs. three benign) indicate a pathogenic impact. There is no ClinVar entry to contradict this assessment, so the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -9.428 | Likely Pathogenic | 0.907 | Likely Pathogenic | Ambiguous | 0.166 | Likely Benign | -1.69 | Neutral | 0.981 | Probably Damaging | 0.954 | Probably Damaging | 3.84 | Benign | 0.00 | Affected | 0.1033 | 0.4549 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||||||||
| c.464G>C | S155T 2D AIThe SynGAP1 missense variant S155T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors a benign outcome. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of reliable predictors and the consensus analysis indicate that S155T is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -8.635 | Likely Pathogenic | 0.523 | Ambiguous | Likely Benign | 0.153 | Likely Benign | -1.60 | Neutral | 0.956 | Probably Damaging | 0.931 | Probably Damaging | 3.84 | Benign | 0.00 | Affected | 0.1151 | 0.5694 | 1 | 1 | 0.1 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.464G>T | S155I 2D AIThe SynGAP1 missense variant S155I is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict it to be pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized scores the variant as pathogenic, the SGM‑Consensus indicates a likely pathogenic status, and the Foldetta stability analysis is unavailable. Taken together, the majority of evidence points to a pathogenic impact for S155I. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -11.298 | Likely Pathogenic | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.264 | Likely Benign | -2.70 | Deleterious | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 3.81 | Benign | 0.00 | Affected | 0.0764 | 0.5711 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.465C>A | S155R 2D AIThe SynGAP1 missense variant S155R is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict it to be pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized scores the variant as pathogenic, the SGM‑Consensus indicates a likely pathogenic status, and a Foldetta stability analysis is not available. Taken together, the majority of evidence points to a pathogenic impact for S155R. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -11.939 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.160 | Likely Benign | -2.74 | Deleterious | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.88 | Benign | 0.00 | Affected | 0.0712 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.465C>G | S155R 2D AIThe SynGAP1 missense variant S155R is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict it to be pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized scores the variant as pathogenic, the SGM‑Consensus indicates a likely pathogenic status, and a Foldetta stability analysis is not available. Taken together, the majority of evidence points to a pathogenic impact for S155R. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -11.939 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.160 | Likely Benign | -2.74 | Deleterious | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.88 | Benign | 0.00 | Affected | 0.0712 | 0.3528 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||||
| c.466T>A | F156I 2D AIThe SynGAP1 missense variant F156I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 pathogenic vs. 2 benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictors (six out of nine) indicate a pathogenic impact, whereas three predict benign. Consequently, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -10.505 | Likely Pathogenic | 0.959 | Likely Pathogenic | Likely Pathogenic | 0.194 | Likely Benign | -2.38 | Neutral | 0.981 | Probably Damaging | 0.966 | Probably Damaging | 3.99 | Benign | 0.00 | Affected | 0.1960 | 0.1938 | 1 | 0 | 1.7 | -34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.466T>C | F156L 2D AIThe SynGAP1 missense variant F156L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas tools that agree on a pathogenic effect include polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) leans toward benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of conventional tools predict pathogenicity, but the high‑accuracy consensus is ambiguous. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -7.437 | In-Between | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.180 | Likely Benign | -2.31 | Neutral | 0.956 | Probably Damaging | 0.931 | Probably Damaging | 4.04 | Benign | 0.00 | Affected | 0.2067 | 0.3109 | 2 | 0 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.466T>G | F156V 2D AIThe SynGAP1 missense variant F156V is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—classify the change as pathogenic. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is uncertain, but the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic status. Foldetta, a protein‑folding stability predictor that integrates FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the preponderance of evidence points to a pathogenic effect for F156V, and this conclusion does not conflict with the current ClinVar annotation, which is absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -11.945 | Likely Pathogenic | 0.950 | Likely Pathogenic | Ambiguous | 0.246 | Likely Benign | -2.91 | Deleterious | 0.981 | Probably Damaging | 0.954 | Probably Damaging | 4.03 | Benign | 0.00 | Affected | 0.2177 | 0.2166 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.467T>A | F156Y 2D AIThe SynGAP1 missense variant F156Y is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas a majority of tools (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a tie (2 pathogenic vs. 2 benign) and thus unavailable; Foldetta predictions are not provided. Overall, the balance of evidence (5 pathogenic vs. 3 benign predictions) indicates that the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -11.811 | Likely Pathogenic | 0.869 | Likely Pathogenic | Ambiguous | 0.136 | Likely Benign | -1.51 | Neutral | 0.981 | Probably Damaging | 0.931 | Probably Damaging | 3.96 | Benign | 0.00 | Affected | 0.1363 | 0.1419 | 7 | 3 | -4.1 | 16.00 | ||||||||||||||||||||||||||||||||||||||||
| c.467T>C | F156S 2D AIThe SynGAP1 missense variant F156S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is “Likely Pathogenic” based on a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus (derived from the same set of high‑accuracy predictors) also indicates a likely pathogenic outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -14.082 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.282 | Likely Benign | -3.62 | Deleterious | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.97 | Benign | 0.00 | Affected | 0.5183 | 0.0200 | Weaken | -3 | -2 | -3.6 | -60.10 | ||||||||||||||||||||||||||||||||||||||
| c.467T>G | F156C 2D AIThe SynGAP1 missense variant F156C (ClinVar ID 3651386) is annotated as benign in ClinVar and is not reported in gnomAD. Functional prediction tools show a split opinion: benign calls come from REVEL and FATHMM, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus is labeled likely pathogenic. No Foldetta stability analysis is available for this variant. Overall, the majority of computational evidence points toward a pathogenic effect, which contradicts the benign classification in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | Likely Benign | 1 | -13.658 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.297 | Likely Benign | -3.54 | Deleterious | 0.999 | Probably Damaging | 0.990 | Probably Damaging | 3.92 | Benign | 0.00 | Affected | 0.3030 | 0.1291 | -4 | -2 | -0.3 | -44.04 | |||||||||||||||||||||||||||||||||||||
| c.468T>A | F156L 2D AIThe SynGAP1 missense variant F156L is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas tools that agree on a pathogenic effect include polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) leans toward benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of conventional tools predict pathogenicity, but the high‑accuracy consensus is ambiguous. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -7.437 | In-Between | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.164 | Likely Benign | -2.31 | Neutral | 0.956 | Probably Damaging | 0.931 | Probably Damaging | 4.04 | Benign | 0.00 | Affected | 0.2067 | 0.3109 | 2 | 0 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.468T>G | F156L 2D AISynGAP1 missense variant F156L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (uncertain), FATHMM (benign), and PROVEAN (benign)—leans toward benign. Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the preponderance of conventional tools indicates a pathogenic effect, and the high‑accuracy predictions are mixed; no ClinVar entry exists to contradict the assessment. Therefore, the variant is most likely pathogenic based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -7.437 | In-Between | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.163 | Likely Benign | -2.31 | Neutral | 0.956 | Probably Damaging | 0.931 | Probably Damaging | 4.04 | Benign | 0.00 | Affected | 0.2067 | 0.3109 | 2 | 0 | 1.0 | -34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.469C>A | R157S 2D AIThe SynGAP1 R157S missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, while the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. ESM1b remains uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic majority vote (2 pathogenic, 1 benign, 1 uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that R157S is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.483068 | Structured | 0.523978 | Binding | 0.306 | 0.777 | 0.375 | -7.573 | In-Between | 0.977 | Likely Pathogenic | Likely Pathogenic | 0.211 | Likely Benign | -2.82 | Deleterious | 0.993 | Probably Damaging | 0.982 | Probably Damaging | 3.85 | Benign | 0.00 | Affected | 0.3484 | 0.2563 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.469C>G | R157G 2D AIThe SynGAP1 R157G missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized yields an Uncertain result, SGM‑Consensus indicates Likely Pathogenic, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) is unavailable for this variant. Overall, the majority of evidence points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.523978 | Binding | 0.306 | 0.777 | 0.375 | -9.125 | Likely Pathogenic | 0.913 | Likely Pathogenic | Ambiguous | 0.252 | Likely Benign | -3.52 | Deleterious | 0.993 | Probably Damaging | 0.982 | Probably Damaging | 3.80 | Benign | 0.00 | Affected | 0.3892 | 0.2567 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.469C>T | R157C 2D AIThe SynGAP1 missense variant R157C is listed in gnomAD (ID 6‑33432766‑C‑T) but has no ClinVar entry. Functional prediction tools split in two groups: benign predictions come from REVEL and FATHMM, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score (which is “Likely Pathogenic”). AlphaMissense‑Optimized returns an uncertain result, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available prediction for this variant. High‑accuracy assessments therefore indicate a likely pathogenic consensus from SGM‑Consensus, an uncertain AlphaMissense‑Optimized score, and no Foldetta data. Overall, the majority of evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.523978 | Binding | 0.306 | 0.777 | 0.375 | 6-33432766-C-T | 7 | 4.34e-6 | -11.524 | Likely Pathogenic | 0.880 | Likely Pathogenic | Ambiguous | 0.237 | Likely Benign | -4.02 | Deleterious | 1.000 | Probably Damaging | 0.990 | Probably Damaging | 3.77 | Benign | 0.00 | Affected | 3.74 | 4 | 0.3696 | 0.2092 | -3 | -4 | 7.0 | -53.05 | ||||||||||||||||||||||||||||||||||
| c.46A>C | M16L 2D AIThe SynGAP1 missense variant M16L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | -1.277 | Likely Benign | 0.249 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -0.33 | Neutral | 0.001 | Benign | 0.000 | Benign | 4.37 | Benign | 0.00 | Affected | 0.1805 | 0.4629 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.46A>G | M16V 2D AIThe SynGAP1 missense variant M16V is reported in gnomAD (ID 6‑33420310‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact for M16V, and this conclusion does not contradict any ClinVar classification, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | 6-33420310-A-G | 16 | 1.05e-5 | -1.595 | Likely Benign | 0.128 | Likely Benign | Likely Benign | 0.073 | Likely Benign | -0.07 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.30 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3561 | 0.3937 | 1 | 2 | 2.3 | -32.06 | ||||||||||||||||||||||||||||||||||
| c.46A>T | M16L 2D AIThe SynGAP1 missense variant M16L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | -1.277 | Likely Benign | 0.249 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -0.33 | Neutral | 0.001 | Benign | 0.000 | Benign | 4.37 | Benign | 0.00 | Affected | 0.1805 | 0.4629 | 4 | 2 | 1.9 | -18.03 | |||||||||||||||||||||||||||||||||||||||
| c.470G>A | R157H 2D AIThe SynGAP1 missense variant R157H (ClinVar ID 2065231.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33432767‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—yields a tie and is therefore inconclusive. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant. Overall, the balance of predictions leans toward pathogenic, but the high‑accuracy tools do not provide a definitive verdict. This assessment does not contradict the ClinVar status, which remains Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.483068 | Structured | 0.523978 | Binding | 0.306 | 0.777 | 0.375 | Uncertain | 1 | 6-33432767-G-A | 1 | 6.20e-7 | -10.235 | Likely Pathogenic | 0.604 | Likely Pathogenic | Likely Benign | 0.254 | Likely Benign | -2.23 | Neutral | 0.999 | Probably Damaging | 0.987 | Probably Damaging | 3.80 | Benign | 0.00 | Affected | 3.74 | 4 | 0.2981 | 0.1449 | 2 | 0 | 1.3 | -19.05 | |||||||||||||||||||||||||||||||||
| c.470G>C | R157P 2D AIThe SynGAP1 missense variant R157P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM. All other evaluated tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM‑Consensus indicates it is likely pathogenic, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic; this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.523978 | Binding | 0.306 | 0.777 | 0.375 | -11.463 | Likely Pathogenic | 0.968 | Likely Pathogenic | Likely Pathogenic | 0.450 | Likely Benign | -3.38 | Deleterious | 0.998 | Probably Damaging | 0.992 | Probably Damaging | 3.79 | Benign | 0.00 | Affected | 0.2438 | 0.3438 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.470G>T | R157L 2D AIThe SynGAP1 missense variant R157L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default all classify the variant as damaging. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.483068 | Structured | 0.523978 | Binding | 0.306 | 0.777 | 0.375 | -8.978 | Likely Pathogenic | 0.950 | Likely Pathogenic | Ambiguous | 0.330 | Likely Benign | -3.13 | Deleterious | 0.993 | Probably Damaging | 0.982 | Probably Damaging | 3.81 | Benign | 0.00 | Affected | 0.2015 | 0.3709 | -3 | -2 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||||
| c.475A>C | I159L 2D AIThe SynGAP1 missense variant I159L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. ESM1b is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for I159L, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -7.936 | In-Between | 0.195 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -0.39 | Neutral | 0.904 | Possibly Damaging | 0.847 | Possibly Damaging | 4.04 | Benign | 0.00 | Affected | 0.0776 | 0.2652 | 2 | 2 | -0.7 | 0.00 | |||||||||||||||||||||||||||||||||||||||
| c.475A>G | I159V 2D AIThe SynGAP1 I159V missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b; AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign consensus (2 benign vs. 1 pathogenic, with one uncertain). Foldetta results are unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -9.714 | Likely Pathogenic | 0.384 | Ambiguous | Likely Benign | 0.113 | Likely Benign | -0.25 | Neutral | 0.803 | Possibly Damaging | 0.847 | Possibly Damaging | 3.98 | Benign | 0.00 | Affected | 0.1111 | 0.2736 | 4 | 3 | -0.3 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.475A>T | I159F 2D AIThe SynGAP1 missense variant I159F is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign (2 benign vs. 1 pathogenic, 1 uncertain). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the balance of evidence, particularly from the high‑accuracy tools, points to a benign impact. This conclusion is not contradicted by any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -9.530 | Likely Pathogenic | 0.447 | Ambiguous | Likely Benign | 0.200 | Likely Benign | -1.41 | Neutral | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.87 | Benign | 0.00 | Affected | 0.0490 | 0.2446 | 1 | 0 | -1.7 | 34.02 | ||||||||||||||||||||||||||||||||||||||||
| c.476T>A | I159N 2D AIThe SynGAP1 missense variant I159N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default all predict a pathogenic impact. The high‑accuracy AlphaMissense‑Optimized result is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑vs‑2 split. Foldetta, which would assess protein‑folding stability, has no available output for this variant. Overall, the majority of tools (five out of eight) predict pathogenicity, while three predict benign and one is uncertain. Thus, the variant is most likely pathogenic based on current computational predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -14.684 | Likely Pathogenic | 0.891 | Likely Pathogenic | Ambiguous | 0.218 | Likely Benign | -1.93 | Neutral | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 3.82 | Benign | 0.00 | Affected | 0.0919 | 0.0342 | -2 | -3 | -8.0 | 0.94 | ||||||||||||||||||||||||||||||||||||||||
| c.476T>C | I159T 2D AIThe SynGAP1 missense variant I159T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that predict a pathogenic effect comprise polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy AlphaMissense‑Optimized predicts pathogenic, while AlphaMissense‑Default also predicts pathogenic. Given that six of the seven individual tools predict pathogenicity versus three predicting benign, the variant is most likely pathogenic. This prediction does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -12.422 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | 0.218 | Likely Benign | -1.69 | Neutral | 0.981 | Probably Damaging | 0.966 | Probably Damaging | 3.86 | Benign | 0.00 | Affected | 0.1069 | 0.0685 | 0 | -1 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||||||||
| c.476T>G | I159S 2D AIThe SynGAP1 missense variant I159S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool rates the variant as uncertain, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -13.684 | Likely Pathogenic | 0.905 | Likely Pathogenic | Ambiguous | 0.241 | Likely Benign | -1.70 | Neutral | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.85 | Benign | 0.00 | Affected | 0.2854 | 0.0712 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.477C>G | I159M 2D AIThe SynGAP1 missense variant I159M is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical databases. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -9.838 | Likely Pathogenic | 0.142 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -0.48 | Neutral | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 3.89 | Benign | 0.00 | Affected | 0.0678 | 0.2500 | 2 | 1 | -2.6 | 18.03 | |||||||||||||||||||||||||||||||||||||||
| c.478C>A | L160M 2D AIThe SynGAP1 missense variant L160M is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive because it yields a 2‑to‑2 split. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; the SGM Consensus is unavailable; and Foldetta, which combines FoldX‑MD and Rosetta stability calculations, has no reported output for this variant. Overall, the balance of evidence leans toward a pathogenic interpretation, but the single high‑accuracy benign prediction and the lack of a ClinVar assertion mean the variant’s clinical significance remains uncertain. This assessment does not contradict any existing ClinVar status, as none is available. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.526760 | Binding | 0.275 | 0.728 | 0.125 | -11.120 | Likely Pathogenic | 0.723 | Likely Pathogenic | Likely Benign | 0.097 | Likely Benign | -0.94 | Neutral | 0.877 | Possibly Damaging | 0.580 | Possibly Damaging | 3.85 | Benign | 0.00 | Affected | 0.0837 | 0.3613 | 4 | 2 | -1.9 | 18.03 | ||||||||||||||||||||||||||||||||||||||||
| c.478C>G | L160V 2D AIThe SynGAP1 missense variant L160V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of predictions (six benign vs. three pathogenic) indicate that the variant is most likely benign. This conclusion is consistent with the lack of ClinVar evidence and gnomAD presence, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.526760 | Binding | 0.275 | 0.728 | 0.125 | -12.506 | Likely Pathogenic | 0.729 | Likely Pathogenic | Likely Benign | 0.055 | Likely Benign | -1.44 | Neutral | 0.247 | Benign | 0.113 | Benign | 3.90 | Benign | 0.00 | Affected | 0.1553 | 0.3319 | 2 | 1 | 0.4 | -14.03 | ||||||||||||||||||||||||||||||||||||||||
| c.479T>A | L160Q 2D AIThe SynGAP1 missense variant L160Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.454136 | Structured | 0.526760 | Binding | 0.275 | 0.728 | 0.125 | -16.626 | Likely Pathogenic | 0.973 | Likely Pathogenic | Likely Pathogenic | 0.227 | Likely Benign | -2.83 | Deleterious | 0.700 | Possibly Damaging | 0.483 | Possibly Damaging | 3.87 | Benign | 0.00 | Affected | 0.1248 | 0.1060 | -2 | -2 | -7.3 | 14.97 | |||||||||||||||||||||||||||||||||||||||
| c.479T>C | L160P 2D AIThe SynGAP1 missense variant L160P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a majority pathogenic vote (3 pathogenic vs. 1 benign) and is labeled “Likely Pathogenic.” AlphaMissense‑Optimized independently predicts pathogenicity. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.454136 | Structured | 0.526760 | Binding | 0.275 | 0.728 | 0.125 | -15.939 | Likely Pathogenic | 0.969 | Likely Pathogenic | Likely Pathogenic | 0.283 | Likely Benign | -3.06 | Deleterious | 0.001 | Benign | 0.000 | Benign | 3.86 | Benign | 0.00 | Affected | 0.3589 | 0.1484 | -3 | -3 | -5.4 | -16.04 | |||||||||||||||||||||||||||||||||||||||
| c.479T>G | L160R 2D AIThe SynGAP1 missense variant L160R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. The protein‑folding stability method Foldetta did not provide a result, so its status is unavailable. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.454136 | Structured | 0.526760 | Binding | 0.275 | 0.728 | 0.125 | -14.539 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | 0.190 | Likely Benign | -2.83 | Deleterious | 0.700 | Possibly Damaging | 0.483 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.1482 | 0.0702 | -3 | -2 | -8.3 | 43.03 | |||||||||||||||||||||||||||||||||||||||
| c.47T>A | M16K 2D AIThe SynGAP1 missense variant M16K is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational predictions and the high‑accuracy consensus suggest that M16K is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | -2.567 | Likely Benign | 0.635 | Likely Pathogenic | Likely Benign | 0.185 | Likely Benign | -0.37 | Neutral | 0.003 | Benign | 0.001 | Benign | 4.23 | Benign | 0.00 | Affected | 0.1984 | 0.1112 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||||||||||||
| c.47T>C | M16T 2D AIThe SynGAP1 missense variant M16T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign and the SGM‑Consensus classifies it as likely benign. No Foldetta stability analysis is available for this residue, so folding‑stability evidence is lacking. Overall, the majority of computational evidence indicates that M16T is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | -2.060 | Likely Benign | 0.515 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -0.46 | Neutral | 0.006 | Benign | 0.001 | Benign | 4.23 | Benign | 0.00 | Affected | 0.2385 | 0.2545 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.47T>G | M16R 2D AIThe SynGAP1 missense variant M16R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign, while the only pathogenic call comes from SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as likely benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus also indicates likely benign, and no Foldetta stability data are available. Taken together, the preponderance of evidence supports a benign classification for M16R, and this conclusion does not conflict with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | -1.475 | Likely Benign | 0.485 | Ambiguous | Likely Benign | 0.191 | Likely Benign | -0.25 | Neutral | 0.014 | Benign | 0.003 | Benign | 4.21 | Benign | 0.00 | Affected | 0.1992 | 0.1093 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.481C>A | P161T 2D AIThe SynGAP1 missense variant P161T has no ClinVar entry and is not reported in gnomAD. Computational predictors fall into two groups: benign predictions come from REVEL, polyPhen‑2 HumVar, and FATHMM, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy tools further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect, and this assessment is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -8.759 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 0.153 | Likely Benign | -3.77 | Deleterious | 0.535 | Possibly Damaging | 0.310 | Benign | 3.92 | Benign | 0.00 | Affected | 0.1891 | 0.5038 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.481C>G | P161A 2D AIThe SynGAP1 missense variant P161A is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. When predictions are grouped by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Overall, the balance of evidence, especially the SGM Consensus and the majority of individual predictors, indicates that P161A is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -9.012 | Likely Pathogenic | 0.926 | Likely Pathogenic | Ambiguous | 0.079 | Likely Benign | -3.52 | Deleterious | 0.247 | Benign | 0.091 | Benign | 3.95 | Benign | 0.00 | Affected | 0.3600 | 0.4123 | 1 | -1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||||||
| c.481C>T | P161S 2D AIThe SynGAP1 missense variant P161S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta results are unavailable. Based on the overall distribution of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -8.550 | Likely Pathogenic | 0.945 | Likely Pathogenic | Ambiguous | 0.085 | Likely Benign | -3.63 | Deleterious | 0.700 | Possibly Damaging | 0.383 | Benign | 3.94 | Benign | 0.00 | Affected | 0.3628 | 0.4597 | 1 | -1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||||
| c.482C>A | P161H 2D AIThe SynGAP1 missense variant P161H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN)—all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus reports it as Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple in silico predictors indicates that P161H is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -12.103 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 0.291 | Likely Benign | -4.22 | Deleterious | 0.964 | Probably Damaging | 0.650 | Possibly Damaging | 3.89 | Benign | 0.00 | Affected | 0.2060 | 0.4039 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.482C>G | P161R 2D AIThe SynGAP1 missense variant P161R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -13.014 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.302 | Likely Benign | -4.16 | Deleterious | 0.700 | Possibly Damaging | 0.483 | Possibly Damaging | 3.93 | Benign | 0.00 | Affected | 0.1716 | 0.2900 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.482C>T | P161L 2D AIThe SynGAP1 missense variant P161L is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus (SGM‑Consensus) derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN classifies the variant as Likely Pathogenic. AlphaMissense‑Optimized also predicts Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the majority of predictions (seven pathogenic vs. four benign) indicate that P161L is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.509769 | Disordered | 0.520000 | Binding | 0.256 | 0.713 | 0.375 | -12.159 | Likely Pathogenic | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.236 | Likely Benign | -4.48 | Deleterious | 0.001 | Benign | 0.003 | Benign | 3.92 | Benign | 0.00 | Affected | 0.2424 | 0.5902 | -3 | -3 | 5.4 | 16.04 | |||||||||||||||||||||||||||||||||||||||
| c.488T>C | F163S 2D AIThe SynGAP1 missense variant F163S has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus reports it as Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the preponderance of evidence indicates that F163S is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.433034 | Structured | 0.513928 | Binding | 0.325 | 0.686 | 0.375 | -13.338 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.273 | Likely Benign | -2.64 | Deleterious | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 4.05 | Benign | 0.00 | Affected | 0.5240 | 0.0358 | Weaken | -3 | -2 | -3.6 | -60.10 | ||||||||||||||||||||||||||||||||||||||
| c.488T>G | F163C 2D AIThe SynGAP1 missense variant F163C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM. All other evaluated tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus indicates a likely pathogenic status. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of predictions, including the high‑accuracy tools, point to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.433034 | Structured | 0.513928 | Binding | 0.325 | 0.686 | 0.375 | -12.221 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | 0.282 | Likely Benign | -2.99 | Deleterious | 0.999 | Probably Damaging | 0.990 | Probably Damaging | 4.01 | Benign | 0.00 | Affected | 0.2860 | 0.1232 | -4 | -2 | -0.3 | -44.04 | |||||||||||||||||||||||||||||||||||||||
| c.48G>A | M16I 2D AIThe SynGAP1 missense variant M16I is listed in ClinVar with an “Uncertain” status (ClinVar ID 1424213.0) and is present in gnomAD (6‑33420312‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | Uncertain | 1 | 6-33420312-G-A | 1 | 6.49e-7 | -2.198 | Likely Benign | 0.722 | Likely Pathogenic | Likely Benign | 0.057 | Likely Benign | -0.15 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1606 | 0.3877 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||
| c.48G>C | M16I 2D AIThe SynGAP1 missense variant M16I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational predictions and the high‑accuracy consensus suggest that M16I is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | -2.198 | Likely Benign | 0.722 | Likely Pathogenic | Likely Benign | 0.057 | Likely Benign | -0.15 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1606 | 0.3877 | 2 | 1 | 2.6 | -18.03 | |||||||||||||||||||||||||||||||||||||
| c.48G>T | M16I 2D AIThe SynGAP1 missense variant M16I is not reported in ClinVar (ClinVar ID = None) but is present in gnomAD (ID = 6‑33420312‑G‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | 6-33420312-G-T | -2.198 | Likely Benign | 0.722 | Likely Pathogenic | Likely Benign | 0.057 | Likely Benign | -0.15 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1606 | 0.3877 | 2 | 1 | 2.6 | -18.03 | ||||||||||||||||||||||||||||||||||||
| c.490C>G | R164G 2D AIThe SynGAP1 missense variant R164G has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a split opinion: benign calls come from REVEL, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: the SGM‑Consensus remains Likely Pathogenic, AlphaMissense‑Optimized is inconclusive, and the Foldetta stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic impact. Because there is no ClinVar classification to oppose this, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.414856 | Structured | 0.512396 | Binding | 0.317 | 0.666 | 0.250 | -12.416 | Likely Pathogenic | 0.879 | Likely Pathogenic | Ambiguous | 0.190 | Likely Benign | -3.01 | Deleterious | 0.487 | Possibly Damaging | 0.272 | Benign | 3.77 | Benign | 0.00 | Affected | 0.3657 | 0.3631 | -3 | -2 | 4.1 | -99.14 | |||||||||||||||||||||||||||||||||||||||
| c.491G>A | R164Q 2D AISynGAP1 missense variant R164Q is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33432788‑G‑A). Functional prediction tools show mixed results: benign predictions come from REVEL, PROVEAN, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic predictions come from polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments indicate that AlphaMissense‑Optimized predicts a benign effect, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split; Foldetta results are not available. Overall, the balance of evidence slightly favors a benign interpretation, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.414856 | Structured | 0.512396 | Binding | 0.317 | 0.666 | 0.250 | Uncertain | 1 | 6-33432788-G-A | 2 | 1.24e-6 | -11.208 | Likely Pathogenic | 0.600 | Likely Pathogenic | Likely Benign | 0.184 | Likely Benign | -1.86 | Neutral | 0.957 | Probably Damaging | 0.342 | Benign | 3.82 | Benign | 0.00 | Affected | 3.74 | 4 | 0.3607 | 0.2711 | 1 | 1 | 1.0 | -28.06 | |||||||||||||||||||||||||||||||||
| c.491G>C | R164P 2D AIThe SynGAP1 missense variant R164P is reported in gnomAD (ID 6‑33432788‑G‑C) but has no ClinVar entry. Functional prediction tools split in two groups: benign predictions come from REVEL and FATHMM, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. AlphaMissense‑Optimized returns an Uncertain result, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available output for this variant. Overall, the majority of high‑confidence tools predict pathogenicity, and this assessment does not contradict any ClinVar status (none is available). Therefore, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.414856 | Structured | 0.512396 | Binding | 0.317 | 0.666 | 0.250 | 6-33432788-G-C | 1 | 6.20e-7 | -12.792 | Likely Pathogenic | 0.898 | Likely Pathogenic | Ambiguous | 0.339 | Likely Benign | -3.42 | Deleterious | 0.910 | Possibly Damaging | 0.578 | Possibly Damaging | 3.77 | Benign | 0.00 | Affected | 3.74 | 4 | 0.2408 | 0.4730 | -2 | 0 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||||
| c.491G>T | R164L 2D AIThe SynGAP1 missense variant R164L is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls arise from PROVEAN, SIFT, ESM1b and AlphaMissense‑Default. Grouping by consensus, four tools predict benign and four predict pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Taken together, the majority of evidence, including the SGM Consensus, points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for R164L. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.414856 | Structured | 0.512396 | Binding | 0.317 | 0.666 | 0.250 | -10.794 | Likely Pathogenic | 0.910 | Likely Pathogenic | Ambiguous | 0.274 | Likely Benign | -3.37 | Deleterious | 0.001 | Benign | 0.003 | Benign | 3.80 | Benign | 0.00 | Affected | 0.2137 | 0.5154 | -3 | -2 | 8.3 | -43.03 | |||||||||||||||||||||||||||||||||||||||
| c.493A>C | S165R 2D AIThe SynGAP1 missense variant S165R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs four benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -9.527 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.293 | Likely Benign | -1.89 | Neutral | 0.567 | Possibly Damaging | 0.249 | Benign | 4.01 | Benign | 0.00 | Affected | 0.1003 | 0.3969 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.493A>G | S165G 2D AIThe SynGAP1 missense variant S165G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b is uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable. Overall, the preponderance of evidence points to the variant being most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -7.691 | In-Between | 0.223 | Likely Benign | Likely Benign | 0.182 | Likely Benign | -1.17 | Neutral | 0.272 | Benign | 0.086 | Benign | 3.99 | Benign | 0.00 | Affected | 0.2622 | 0.4441 | 1 | 0 | 0.4 | -30.03 | |||||||||||||||||||||||||||||||||||||||
| c.493A>T | S165C 2D AIThe SynGAP1 missense variant S165C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact for S165C, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -8.425 | Likely Pathogenic | 0.367 | Ambiguous | Likely Benign | 0.304 | Likely Benign | -1.92 | Neutral | 0.938 | Possibly Damaging | 0.498 | Possibly Damaging | 3.94 | Benign | 0.00 | Affected | 0.1306 | 0.5534 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.494G>A | S165N 2D AIThe SynGAP1 missense variant S165N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and ESM1b. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for S165N. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -9.041 | Likely Pathogenic | 0.509 | Ambiguous | Likely Benign | 0.074 | Likely Benign | -0.49 | Neutral | 0.532 | Possibly Damaging | 0.229 | Benign | 4.03 | Benign | 0.00 | Affected | 0.1589 | 0.4862 | 1 | 1 | -2.7 | 27.03 | ||||||||||||||||||||||||||||||||||||||||
| c.494G>C | S165T 2D AIThe SynGAP1 missense variant S165T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect. The variant is most likely benign, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -6.952 | Likely Benign | 0.205 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -0.94 | Neutral | 0.155 | Benign | 0.064 | Benign | 4.02 | Benign | 0.00 | Affected | 0.1754 | 0.5779 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.494G>T | S165I 2D AISynGAP1 missense variant S165I has no ClinVar record and is absent from gnomAD. Individual prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains Likely Pathogenic; Foldetta stability analysis is unavailable. Overall, the majority of single‑tool predictions favor a benign effect, whereas the consensus score suggests pathogenicity. Consequently, the variant is most likely benign according to the bulk of evidence, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -11.304 | Likely Pathogenic | 0.774 | Likely Pathogenic | Likely Benign | 0.259 | Likely Benign | -2.53 | Deleterious | 0.084 | Benign | 0.031 | Benign | 3.96 | Benign | 0.00 | Affected | 0.0943 | 0.5027 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.495T>A | S165R 2D AIThe SynGAP1 missense variant S165R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 vs 2). High‑accuracy AlphaMissense‑Optimized predicts pathogenic, while Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. four benign) indicate a pathogenic effect. There is no ClinVar annotation to contradict this assessment, so the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -9.527 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.177 | Likely Benign | -1.89 | Neutral | 0.567 | Possibly Damaging | 0.249 | Benign | 4.01 | Benign | 0.00 | Affected | 0.1003 | 0.3969 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.495T>G | S165R 2D AIThe SynGAP1 missense variant S165R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect include polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. High‑accuracy tools specifically show AlphaMissense‑Optimized as pathogenic, while SGM Consensus and Foldetta are unavailable. Based on the available predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -9.527 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.177 | Likely Benign | -1.89 | Neutral | 0.567 | Possibly Damaging | 0.249 | Benign | 4.01 | Benign | 0.00 | Affected | 0.1003 | 0.3969 | 0 | -1 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||||||
| c.499G>A | D167N 2D AIThe SynGAP1 missense variant D167N is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33432796‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Tools that predict a pathogenic effect are SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM Consensus, which takes a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic, two benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the available predictions, the variant is most likely benign; this assessment does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.502306 | Binding | 0.377 | 0.667 | 0.375 | 6-33432796-G-A | 3 | 1.86e-6 | -11.939 | Likely Pathogenic | 0.843 | Likely Pathogenic | Ambiguous | 0.097 | Likely Benign | -2.32 | Neutral | 0.141 | Benign | 0.123 | Benign | 3.96 | Benign | 0.00 | Affected | 3.74 | 4 | 0.1222 | 0.7330 | 1 | 2 | 0.0 | -0.98 | |||||||||||||||||||||||||||||||||||
| c.499G>C | D167H 2D AIThe SynGAP1 missense variant D167H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. Foldetta results are unavailable, so they do not influence the overall assessment. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.502306 | Binding | 0.377 | 0.667 | 0.375 | -12.606 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | 0.338 | Likely Benign | -3.29 | Deleterious | 0.898 | Possibly Damaging | 0.557 | Possibly Damaging | 3.93 | Benign | 0.00 | Affected | 0.1478 | 0.7563 | 1 | -1 | 0.3 | 22.05 | |||||||||||||||||||||||||||||||||||||||
| c.499G>T | D167Y 2D AIThe SynGAP1 missense variant D167Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and FATHMM, whereas the remaining eight predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized—and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate pathogenicity. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus is labeled Likely Pathogenic. No Foldetta stability analysis is available for this residue. Overall, the majority of evidence points to a pathogenic impact for D167Y, and this conclusion is not contradicted by any ClinVar annotation, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.502306 | Binding | 0.377 | 0.667 | 0.375 | -14.228 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.414 | Likely Benign | -4.23 | Deleterious | 0.898 | Possibly Damaging | 0.557 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.0493 | 0.6358 | -4 | -3 | 2.2 | 48.09 | |||||||||||||||||||||||||||||||||||||||
| c.49T>A | S17T 2D AIThe SynGAP1 missense variant S17T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect. The variant is most likely benign, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | -3.782 | Likely Benign | 0.197 | Likely Benign | Likely Benign | 0.061 | Likely Benign | -0.58 | Neutral | 0.052 | Benign | 0.004 | Benign | 4.10 | Benign | 0.00 | Affected | 0.1667 | 0.6933 | 1 | 1 | 0.1 | 14.03 | |||||||||||||||||||||||||||||||||||||||
| c.49T>C | S17P 2D AIThe SynGAP1 missense variant S17P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus, SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a benign effect. AlphaMissense‑Optimized independently scores the variant as benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | -3.251 | Likely Benign | 0.256 | Likely Benign | Likely Benign | 0.210 | Likely Benign | -0.65 | Neutral | 0.212 | Benign | 0.010 | Benign | 4.01 | Benign | 0.00 | Affected | 0.2406 | 0.6473 | 1 | -1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||||||
| c.49T>G | S17A 2D AIThe SynGAP1 missense variant S17A is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta results are unavailable. Overall, the majority of computational evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.483068 | Structured | 0.452228 | Uncertain | 0.341 | 0.910 | 0.375 | -3.140 | Likely Benign | 0.166 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -0.34 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.15 | Benign | 0.00 | Affected | 0.5276 | 0.5824 | Strenghten | 1 | 1 | 2.6 | -16.00 | ||||||||||||||||||||||||||||||||||||||
| c.4A>C | S2R 2D AIThe SynGAP1 missense variant S2R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.543646 | Binding | 0.382 | 0.922 | 0.750 | -3.684 | Likely Benign | 0.426 | Ambiguous | Likely Benign | 0.062 | Likely Benign | -0.44 | Neutral | 0.117 | Benign | 0.008 | Benign | 4.05 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0996 | 0.4503 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.4A>G | S2G 2D AIThe SynGAP1 missense variant S2G is reported in gnomAD (ID 6‑33420268‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign effect for this variant, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.543646 | Binding | 0.382 | 0.922 | 0.750 | 6-33420268-A-G | 1 | 6.58e-7 | -4.273 | Likely Benign | 0.124 | Likely Benign | Likely Benign | 0.079 | Likely Benign | -0.48 | Neutral | 0.012 | Benign | 0.002 | Benign | 4.10 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2872 | 0.5495 | 0 | 1 | 0.4 | -30.03 | ||||||||||||||||||||||||||||||||||
| c.4A>T | S2C 2D AIThe SynGAP1 missense variant S2C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.543646 | Binding | 0.382 | 0.922 | 0.750 | -5.328 | Likely Benign | 0.187 | Likely Benign | Likely Benign | 0.067 | Likely Benign | 0.16 | Neutral | 0.916 | Possibly Damaging | 0.091 | Benign | 4.01 | Benign | 0.00 | Affected | 0.1145 | 0.6462 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.500A>C | D167A 2D AIThe SynGAP1 D167A missense variant has no ClinVar entry and is not reported in gnomAD. Functional prediction tools split evenly: benign calls come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls come from PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessment shows AlphaMissense‑Optimized as uncertain, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability predictor, has no available result for this residue. Taken together, the preponderance of evidence, especially the SGM Consensus, points to a pathogenic effect. This conclusion is consistent with the absence of a ClinVar classification, so there is no contradiction with existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.502306 | Binding | 0.377 | 0.667 | 0.375 | -13.473 | Likely Pathogenic | 0.921 | Likely Pathogenic | Ambiguous | 0.291 | Likely Benign | -3.61 | Deleterious | 0.141 | Benign | 0.056 | Benign | 3.97 | Benign | 0.00 | Affected | 0.3429 | 0.6549 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||||
| c.500A>G | D167G 2D AIThe SynGAP1 missense variant D167G is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, while Foldetta (combining FoldX‑MD and Rosetta outputs) is not available for this variant. Overall, the majority of evidence points to a pathogenic impact. The prediction is consistent with the absence of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.502306 | Binding | 0.377 | 0.667 | 0.375 | -11.806 | Likely Pathogenic | 0.898 | Likely Pathogenic | Ambiguous | 0.338 | Likely Benign | -3.20 | Deleterious | 0.141 | Benign | 0.091 | Benign | 3.93 | Benign | 0.00 | Affected | 0.3580 | 0.6750 | 1 | -1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||||||||
| c.500A>T | D167V 2D AIThe SynGAP1 D167V variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic impact are PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. The Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.502306 | Binding | 0.377 | 0.667 | 0.375 | -14.388 | Likely Pathogenic | 0.967 | Likely Pathogenic | Likely Pathogenic | 0.396 | Likely Benign | -4.03 | Deleterious | 0.535 | Possibly Damaging | 0.247 | Benign | 3.92 | Benign | 0.00 | Affected | 0.0709 | 0.6905 | -2 | -3 | 7.7 | -15.96 | |||||||||||||||||||||||||||||||||||||||
| c.501C>A | D167E 2D AIThe SynGAP1 D167E missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive because it receives two benign votes and two uncertain votes, yielding no clear majority. Foldetta, which would provide a protein‑folding stability estimate, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.502306 | Binding | 0.377 | 0.667 | 0.375 | -7.636 | In-Between | 0.516 | Ambiguous | Likely Benign | 0.067 | Likely Benign | -1.52 | Neutral | 0.063 | Benign | 0.062 | Benign | 4.07 | Benign | 0.00 | Affected | 0.1392 | 0.7144 | 3 | 2 | 0.0 | 14.03 | ||||||||||||||||||||||||||||||||||||||||
Found 8751 rows. Show 800 rows per page. Page 3/11 « Previous | Next »