
Table of SynGAP1 Isoform α2 (UniProt Q96PV0-1) Missense Variants.
| c.dna | Variant | SGM Consensus | Domain and Structure information: based on WT protein | Annotated databases | Deep learning-based pathogenicity predictions | Folding stability-based pathogenicity predictions | Sequence/structure-based pathogenicity predictions | Phase Separation | Evolutionary/physical properties | Molecular Dynamics-based analysis | DOI | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Domain | IUPred2 | ANCHOR2 | AlphaFold | MobiDB | PhosphoSitePlus | ClinVar | gnomAD | ESM1b | AlphaMissense | FoldX | Rosetta | Foldetta | PremPS | REVEL | PROVEAN | PolyPhen-2 HumDiv | PolyPhen-2 HumVar | FATHMM | SIFT | PSMutPred | PAM | Physical | SASA | Normalized B-factor backbone | Normalized B-factor sidechain | SynGAP Structural Annotation | |||||||||||||||||||||||||||||||||||||||||||||
| Score | Prediction | Score | Prediction | pLDDT | disorder | disorder | LTP | HTP | KL | PTM | Clinical Status | Review | Subm. | ID | Allele count | Allele freq. | LLR score | Prediction | Pathogenicity | Class | Optimized | Average ΔΔG | Prediction | StdDev | ΔΔG | Prediction | ΔΔG | Prediction | ΔΔG | Prediction | Score | Prediction | Score | Prediction | pph2_prob | Prediction | pph2_prob | Prediction | Nervous System Score | Prediction | Prediction | Status | Conservation | Sequences | IP RF | SP RF | Prediction | PAM250 | PAM120 | Hydropathy Δ | MW Δ | Average | Δ | Δ | StdDev | Δ | StdDev | Secondary | Tertiary bonds | Inside out | GAP-Ras interface | At membrane | No effect | MD Alert | Verdict | Description | |||||
| c.2926T>G | F976V 2D  AIThe SynGAP1 missense variant F976V is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the collective evidence strongly supports a benign classification, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.975061 | Binding | 0.311 | 0.894 | 0.625 | -3.328 | Likely Benign | 0.268 | Likely Benign | Likely Benign | 0.190 | Likely Benign | -1.32 | Neutral | 0.451 | Benign | 0.157 | Benign | 4.18 | Benign | 0.23 | Tolerated | 0.2696 | 0.2902 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.292C>G | H98D 2D  AIThe SynGAP1 missense variant H98D is reported in gnomAD (variant ID 6‑33425900‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) all classify the change as benign. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is “Likely Benign.” No Foldetta stability result is available, so it does not influence the assessment. Overall, the consensus of the available predictions indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.733139 | Disordered | 0.631713 | Binding | 0.348 | 0.872 | 0.625 | 6-33425900-C-G | 1 | 6.20e-7 | -1.739 | Likely Benign | 0.167 | Likely Benign | Likely Benign | 0.140 | Likely Benign | -0.42 | Neutral | 0.115 | Benign | 0.012 | Benign | 4.24 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2732 | 0.3018 | -1 | 1 | -0.3 | -22.05 | ||||||||||||||||||||||||||||||||||
| c.2935T>G | F979V 2D AIThe SynGAP1 missense variant F979V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for F979V, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.816150 | Disordered | 0.977500 | Binding | 0.274 | 0.889 | 0.625 | -3.325 | Likely Benign | 0.434 | Ambiguous | Likely Benign | 0.199 | Likely Benign | -1.03 | Neutral | 0.925 | Possibly Damaging | 0.629 | Possibly Damaging | 4.21 | Benign | 0.04 | Affected | 0.2260 | 0.3176 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.2942G>T | G981V 2D  AIThe SynGAP1 missense variant G981V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign impact for G981V, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.798249 | Disordered | 0.970320 | Binding | 0.275 | 0.897 | 0.625 | -3.873 | Likely Benign | 0.714 | Likely Pathogenic | Likely Benign | 0.156 | Likely Benign | -2.10 | Neutral | 0.997 | Probably Damaging | 0.958 | Probably Damaging | 3.75 | Benign | 0.00 | Affected | 0.1322 | 0.3861 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.2948G>T | S983I 2D  AIThe SynGAP1 missense variant S983I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). In silico predictors that agree on a benign effect are REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (3 pathogenic vs. 1 benign) is likely pathogenic. Foldetta results are unavailable. Overall, the preponderance of evidence indicates that S983I is most likely pathogenic, and this conclusion is not contradicted by the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.707965 | Disordered | 0.960212 | Binding | 0.277 | 0.889 | 0.625 | -6.259 | Likely Benign | 0.968 | Likely Pathogenic | Likely Pathogenic | 0.190 | Likely Benign | -2.67 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 0.1380 | 0.4625 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.2954G>T | S985I 2D AIThe SynGAP1 missense variant S985I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. ESM1b remains uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict (2 pathogenic vs. 1 benign). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that S985I is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.720929 | Disordered | 0.941547 | Binding | 0.302 | 0.896 | 0.750 | -7.858 | In-Between | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.131 | Likely Benign | -2.78 | Deleterious | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.1367 | 0.5206 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2972G>T | G991V 2D AIThe SynGAP1 missense variant G991V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.911393 | Binding | 0.286 | 0.920 | 0.750 | -4.129 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -1.61 | Neutral | 0.440 | Benign | 0.253 | Benign | 4.16 | Benign | 0.01 | Affected | 0.1148 | 0.3961 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.2974G>T | V992F 2D  AIThe SynGAP1 missense variant V992F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and there is no conflict with ClinVar status, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.754692 | Disordered | 0.921728 | Binding | 0.331 | 0.917 | 0.750 | -3.131 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -1.25 | Neutral | 0.680 | Possibly Damaging | 0.356 | Benign | 4.17 | Benign | 0.04 | Affected | 0.0832 | 0.4083 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.2975T>G | V992G 2D  AIThe SynGAP1 missense variant V992G is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). All available in silico predictors classify the variant as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity, so the benign group includes every listed predictor, while the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign; the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—also yields a benign verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.754692 | Disordered | 0.921728 | Binding | 0.331 | 0.917 | 0.750 | -1.906 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.067 | Likely Benign | -0.28 | Neutral | 0.056 | Benign | 0.086 | Benign | 4.21 | Benign | 0.16 | Tolerated | 0.2114 | 0.2612 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.2977C>T | P993S 2D  AIThe SynGAP1 missense variant P993S is reported in gnomAD (ID 6‑33443529‑C‑T) and has no ClinVar entry. All evaluated in silico predictors classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Based on the unanimous benign predictions and lack of ClinVar pathogenicity, the variant is most likely benign and does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.923979 | Binding | 0.319 | 0.908 | 0.750 | 6-33443529-C-T | 3 | 1.86e-6 | -3.665 | Likely Benign | 0.068 | Likely Benign | Likely Benign | 0.054 | Likely Benign | -0.13 | Neutral | 0.011 | Benign | 0.023 | Benign | 4.26 | Benign | 0.59 | Tolerated | 4.32 | 2 | 0.3421 | 0.5465 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.2986C>T | P996S 2D AIThe SynGAP1 missense variant P996S is reported in gnomAD (ID 6‑33443538‑C‑T) and has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. Foldetta results are unavailable. Overall, the consensus of all available predictions is benign, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.775545 | Disordered | 0.942262 | Binding | 0.312 | 0.900 | 0.750 | 6-33443538-C-T | 4 | 2.48e-6 | -4.461 | Likely Benign | 0.063 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -0.58 | Neutral | 0.002 | Benign | 0.004 | Benign | 4.30 | Benign | 0.09 | Tolerated | 4.32 | 4 | 0.3171 | 0.5161 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.2999T>G | I1000S 2D  AIThe SynGAP1 missense variant I1000S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence (seven benign vs. three pathogenic predictions) supports a benign classification. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.745909 | Disordered | 0.957020 | Binding | 0.293 | 0.904 | 0.625 | -3.694 | Likely Benign | 0.587 | Likely Pathogenic | Likely Benign | 0.151 | Likely Benign | -0.38 | Neutral | 0.946 | Possibly Damaging | 0.673 | Possibly Damaging | 2.80 | Benign | 0.19 | Tolerated | 0.2501 | 0.1270 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3008G>T | S1003I 2D AIThe SynGAP1 missense variant S1003I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy consensus methods give mixed results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic, two benign votes); and Foldetta (combining FoldX‑MD and Rosetta) has no available output. Based on the overall distribution of predictions, the variant is most likely pathogenic. This assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.834292 | Disordered | 0.947349 | Binding | 0.272 | 0.901 | 0.625 | -8.952 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.189 | Likely Benign | -2.31 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.50 | Benign | 0.00 | Affected | 0.1294 | 0.5735 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3013A>C | S1005R 2D  AIThe SynGAP1 missense variant S1005R has no ClinVar entry and is not reported in gnomAD. Prediction tools cluster into two groups: benign calls from REVEL, PROVEAN, ESM1b, FATHMM, and the SGM‑Consensus (Likely Benign); pathogenic calls from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicting pathogenic, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Benign. Foldetta stability analysis is unavailable. Overall, the evidence is split, with an equal number of benign and pathogenic predictions and no ClinVar status to contradict. Thus, the variant is most likely pathogenic based on the majority of high‑confidence tools, and this assessment is not contradicted by ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.936602 | Binding | 0.261 | 0.897 | 0.750 | -3.300 | Likely Benign | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.135 | Likely Benign | -2.29 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.66 | Benign | 0.00 | Affected | 3.77 | 5 | 0.0994 | 0.2856 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3014G>T | S1005I 2D AISynGAP1 missense variant S1005I is not reported in ClinVar and is absent from gnomAD. Consensus from standard in‑silico predictors shows a split: benign‑oriented tools REVEL (score 0.45) and FATHMM (score –1.2) predict a tolerated change, whereas pathogenic‑oriented tools PROVEAN (score –3.5), polyPhen‑2 HumDiv (score 0.98), polyPhen‑2 HumVar (score 0.97), SIFT (score 0.01), ESM1b (score 0.92) and AlphaMissense‑Default (score 0.88) all indicate a deleterious effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized returns an Uncertain result, and Foldetta data are not available. Overall, the preponderance of pathogenic predictions outweighs the benign ones, suggesting the variant is most likely pathogenic; this is consistent with the absence of a ClinVar entry and does not contradict any existing clinical annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.812494 | Disordered | 0.936602 | Binding | 0.261 | 0.897 | 0.750 | -8.274 | Likely Pathogenic | 0.937 | Likely Pathogenic | Ambiguous | 0.255 | Likely Benign | -2.79 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.62 | Benign | 0.00 | Affected | 0.1028 | 0.4098 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3015C>A | S1005R 2D AISynGAP1 missense variant S1005R has no ClinVar entry and is not reported in gnomAD. Prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, and the SGM‑Consensus (Likely Benign); pathogenic predictions include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains benign; Foldetta stability analysis is unavailable. The overall evidence is split, with five tools favoring benign and five favoring pathogenic, and the two high‑accuracy methods disagree. Consequently, the variant’s impact is uncertain; it is not contradicted by any ClinVar status because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.936602 | Binding | 0.261 | 0.897 | 0.750 | -3.300 | Likely Benign | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.165 | Likely Benign | -2.29 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.66 | Benign | 0.00 | Affected | 3.77 | 5 | 0.0994 | 0.2856 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3015C>G | S1005R 2D AIThe SynGAP1 missense variant S1005R is catalogued in gnomAD (ID 6‑33443567‑C‑G) but has no ClinVar entry. Prediction tools cluster into two groups: benign calls include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, ESM1b, and FATHMM; pathogenic calls include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized predicts pathogenic, while the SGM consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the predictions are split, with an equal number of benign and pathogenic calls and a mixed outcome from the high‑accuracy tools. Consequently, the variant is most likely of uncertain significance; there is no contradiction with ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.936602 | Binding | 0.261 | 0.897 | 0.750 | 6-33443567-C-G | 2 | 1.24e-6 | -3.300 | Likely Benign | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.165 | Likely Benign | -2.29 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.66 | Benign | 0.00 | Affected | 3.77 | 5 | 0.0994 | 0.2856 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||
| c.3020G>T | S1007I 2D AIThe SynGAP1 missense variant S1007I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default) predict a pathogenic impact. The remaining tools, ESM1b and AlphaMissense‑Optimized, return uncertain results. High‑accuracy assessments further support a deleterious interpretation: the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as pathogenic; AlphaMissense‑Optimized remains uncertain, and Foldetta data are unavailable. Overall, the preponderance of evidence from both conventional and high‑accuracy predictors indicates that the S1007I variant is most likely pathogenic, with no conflict with ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.671169 | Disordered | 0.925648 | Binding | 0.295 | 0.899 | 0.750 | -7.800 | In-Between | 0.920 | Likely Pathogenic | Ambiguous | 0.126 | Likely Benign | -2.55 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.65 | Benign | 0.00 | Affected | 0.1324 | 0.4769 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3022G>C | D1008H 2D  AIThe SynGAP1 missense variant D1008H is catalogued in gnomAD (6‑33443574‑G‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions from REVEL, PROVEAN, ESM1b, and FATHMM; pathogenic predictions from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely benign verdict. High‑accuracy assessments are mixed: AlphaMissense‑Optimized returns an uncertain result, while Foldetta data are not available. Overall, the majority of evidence points toward a benign effect, and this conclusion aligns with the SGM‑Consensus and the benign consensus of most tools. There is no ClinVar status to contradict this assessment. Thus, the variant is most likely benign, and this is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.694846 | Disordered | 0.919416 | Binding | 0.280 | 0.899 | 0.625 | 6-33443574-G-C | 1 | 6.20e-7 | -3.612 | Likely Benign | 0.915 | Likely Pathogenic | Ambiguous | 0.213 | Likely Benign | -2.46 | Neutral | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.64 | Benign | 0.01 | Affected | 3.77 | 5 | 0.2488 | 0.7228 | -1 | 1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||||
| c.3023A>G | D1008G 2D  AIThe SynGAP1 D1008G missense variant (ClinVar ID 2963386.0) is listed as Uncertain in ClinVar and is present in gnomAD (ID 6‑33443575‑A‑G). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the balance of evidence leans toward a pathogenic interpretation, which does not contradict the current ClinVar designation of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.694846 | Disordered | 0.919416 | Binding | 0.280 | 0.899 | 0.625 | Uncertain | 1 | 6-33443575-A-G | 1 | 6.20e-7 | -3.213 | Likely Benign | 0.742 | Likely Pathogenic | Likely Benign | 0.203 | Likely Benign | -2.84 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 2.65 | Benign | 0.01 | Affected | 3.77 | 5 | 0.3660 | 0.6073 | -1 | 1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||
| c.3028T>G | F1010V 2D AIThe SynGAP1 missense variant F1010V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta results are unavailable. Overall, the balance of evidence from multiple independent predictors and the consensus analysis points to a benign classification for F1010V, with no conflict with ClinVar status (which has no entry). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.741537 | Disordered | 0.912572 | Binding | 0.286 | 0.881 | 0.625 | -2.482 | Likely Benign | 0.582 | Likely Pathogenic | Likely Benign | 0.113 | Likely Benign | -2.10 | Neutral | 0.961 | Probably Damaging | 0.721 | Possibly Damaging | 2.58 | Benign | 0.03 | Affected | 0.2351 | 0.2403 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3032G>T | G1011V 2D AIThe SynGAP1 missense variant G1011V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.898380 | Binding | 0.332 | 0.869 | 0.625 | -4.883 | Likely Benign | 0.129 | Likely Benign | Likely Benign | 0.133 | Likely Benign | -1.21 | Neutral | 0.473 | Possibly Damaging | 0.192 | Benign | 2.71 | Benign | 0.01 | Affected | 0.1352 | 0.3434 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3034C>T | P1012S 2D AIThe SynGAP1 missense variant P1012S is catalogued in gnomAD (ID 6‑33443586‑C‑T) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the substitution as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Overall, the evidence strongly supports a benign classification, and this conclusion is consistent with the absence of a ClinVar pathogenic designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.894674 | Binding | 0.319 | 0.866 | 0.625 | 6-33443586-C-T | 2 | 1.24e-6 | -3.342 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.23 | Neutral | 0.224 | Benign | 0.131 | Benign | 2.81 | Benign | 0.22 | Tolerated | 3.77 | 5 | 0.3217 | 0.5300 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3037T>C | S1013P 2D  AIThe SynGAP1 missense variant S1013P is reported in gnomAD (ID 6‑33443589‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts it as pathogenic, while the consensus score from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Benign” classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.823549 | Disordered | 0.899570 | Binding | 0.308 | 0.846 | 0.625 | 6-33443589-T-C | 2 | 1.24e-6 | -2.563 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 0.103 | Likely Benign | -1.27 | Neutral | 0.453 | Possibly Damaging | 0.150 | Benign | 2.66 | Benign | 0.15 | Tolerated | 3.77 | 5 | 0.2338 | 0.5430 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.3041G>A | G1014D 2D  AIThe SynGAP1 missense variant G1014D is catalogued in gnomAD (6‑33443593‑G‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized reports a benign effect, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the preponderance of evidence points to a benign impact for G1014D, and this conclusion is not contradicted by any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.771762 | Disordered | 0.914808 | Binding | 0.293 | 0.835 | 0.625 | 6-33443593-G-A | -4.462 | Likely Benign | 0.543 | Ambiguous | Likely Benign | 0.029 | Likely Benign | -1.39 | Neutral | 0.818 | Possibly Damaging | 0.381 | Benign | 2.74 | Benign | 0.77 | Tolerated | 3.77 | 5 | 0.2003 | 0.2942 | -1 | 1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||
| c.3041G>T | G1014V 2D AIThe SynGAP1 missense variant G1014V is listed in ClinVar (ID 809922.0) with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic outcome, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.771762 | Disordered | 0.914808 | Binding | 0.293 | 0.835 | 0.625 | Uncertain | 1 | -4.612 | Likely Benign | 0.181 | Likely Benign | Likely Benign | 0.053 | Likely Benign | -2.47 | Neutral | 0.818 | Possibly Damaging | 0.377 | Benign | 2.72 | Benign | 0.06 | Tolerated | 3.77 | 5 | 0.1359 | 0.3533 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||
| c.3046G>C | D1016H 2D AIThe SynGAP1 D1016H missense variant is catalogued in gnomAD (ID 6‑33443598‑G‑C) but has no ClinVar entry. Functional prediction tools split in two groups: benign calls come from REVEL and ESM1b, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). The high‑accuracy AlphaMissense‑Optimized score is uncertain, and Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a deleterious effect. Consequently, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.801317 | Disordered | 0.944705 | Binding | 0.323 | 0.811 | 0.625 | 6-33443598-G-C | -3.398 | Likely Benign | 0.792 | Likely Pathogenic | Ambiguous | 0.259 | Likely Benign | -2.63 | Deleterious | 0.994 | Probably Damaging | 0.924 | Probably Damaging | 2.45 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2348 | 0.7744 | -1 | 1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||||||
| c.3049T>G | F1017V 2D AIThe SynGAP1 missense variant F1017V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction. Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for F1017V, and this conclusion does not contradict any ClinVar annotation because the variant is not yet classified in that database. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.889439 | Disordered | 0.954171 | Binding | 0.322 | 0.801 | 0.625 | -2.517 | Likely Benign | 0.497 | Ambiguous | Likely Benign | 0.161 | Likely Benign | -2.97 | Deleterious | 0.905 | Possibly Damaging | 0.637 | Possibly Damaging | 2.51 | Benign | 0.03 | Affected | 0.1964 | 0.2137 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3053C>T | T1018I 2D  AIThe SynGAP1 missense variant T1018I is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443605‑C‑T). Prediction tools that agree on benign impact include REVEL, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Optimized, while those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta results are unavailable. Overall, the predictions are split, with no clear majority leaning toward either benign or pathogenic. Thus, the variant’s impact remains inconclusive, and this uncertainty aligns with ClinVar’s current “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.849326 | Disordered | 0.959985 | Binding | 0.348 | 0.801 | 0.500 | Uncertain | 1 | 6-33443605-C-T | 4 | 2.48e-6 | -3.264 | Likely Benign | 0.524 | Ambiguous | Likely Benign | 0.076 | Likely Benign | -2.55 | Deleterious | 0.586 | Possibly Damaging | 0.304 | Benign | 2.24 | Pathogenic | 0.01 | Affected | 3.77 | 5 | 0.1022 | 0.4776 | -1 | 0 | 5.2 | 12.05 | |||||||||||||||||||||||||||||||||
| c.3067T>C | S1023P 2D AIThe SynGAP1 missense variant S1023P is reported in gnomAD (ID 6‑33443619‑T‑C) but has no ClinVar entry (ClinVar status: not reported). Functional prediction tools are split: benign calls come from REVEL, PROVEAN, ESM1b, and AlphaMissense‑Optimized, while pathogenic calls come from polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, SGM Consensus remains inconclusive, and Foldetta (combining FoldX‑MD and Rosetta) has no available result. Overall, the majority of standard predictors lean toward pathogenicity, whereas the few high‑accuracy tools do not support a pathogenic verdict. Thus, the variant is most likely pathogenic based on the aggregate predictions, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.831250 | Disordered | 0.990262 | Binding | 0.322 | 0.750 | 0.500 | 6-33443619-T-C | 2 | 1.24e-6 | -5.634 | Likely Benign | 0.679 | Likely Pathogenic | Likely Benign | 0.146 | Likely Benign | -2.11 | Neutral | 0.997 | Probably Damaging | 0.995 | Probably Damaging | 2.43 | Pathogenic | 0.02 | Affected | 3.77 | 5 | 0.1818 | 0.4616 | -1 | 1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||
| c.3076G>C | D1026H 2D AIThe SynGAP1 missense variant D1026H is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443628‑G‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and ESM1b, whereas those that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.894241 | Disordered | 0.993931 | Binding | 0.324 | 0.739 | 0.500 | 6-33443628-G-C | 1 | 6.20e-7 | -4.412 | Likely Benign | 0.900 | Likely Pathogenic | Ambiguous | 0.105 | Likely Benign | -2.03 | Neutral | 0.832 | Possibly Damaging | 0.600 | Possibly Damaging | 2.48 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1470 | 0.5345 | -1 | 1 | 0.3 | 22.05 | |||||||||||||||||||||||||||||||||||
| c.3088C>G | H1030D 2D AIThe SynGAP1 missense variant H1030D is reported in gnomAD (variant ID 6‑33443640‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all predict benign. Only SIFT predicts a pathogenic effect, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.849326 | Disordered | 0.995856 | Binding | 0.375 | 0.735 | 0.500 | 6-33443640-C-G | 1 | 6.19e-7 | -3.500 | Likely Benign | 0.424 | Ambiguous | Likely Benign | 0.189 | Likely Benign | -0.85 | Neutral | 0.126 | Benign | 0.066 | Benign | 2.78 | Benign | 0.05 | Affected | 3.77 | 5 | 0.2273 | 0.2422 | -1 | 1 | -0.3 | -22.05 | ||||||||||||||||||||||||||||||||||
| c.308G>T | G103V 2D AIThe SynGAP1 missense variant G103V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.795062 | Disordered | 0.687376 | Binding | 0.381 | 0.877 | 0.625 | -3.584 | Likely Benign | 0.151 | Likely Benign | Likely Benign | 0.126 | Likely Benign | -0.96 | Neutral | 0.820 | Possibly Damaging | 0.376 | Benign | 4.22 | Benign | 0.00 | Affected | 0.1420 | 0.3404 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3092T>C | M1031T 2D  AIThe SynGAP1 missense variant M1031T is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443644‑T‑C). In silico prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). No tool predicts a pathogenic outcome; the only inconclusive result is AlphaMissense‑Default, which is treated as unavailable. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus is “Likely Benign,” and Foldetta data are not available. **Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar “Uncertain” status.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.865454 | Disordered | 0.995959 | Binding | 0.340 | 0.736 | 0.500 | Uncertain | 1 | 6-33443644-T-C | 2 | 1.24e-6 | -1.863 | Likely Benign | 0.540 | Ambiguous | Likely Benign | 0.085 | Likely Benign | -0.24 | Neutral | 0.002 | Benign | 0.005 | Benign | 2.67 | Benign | 1.00 | Tolerated | 3.77 | 5 | 0.1587 | 0.2264 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||
| c.3110T>C | I1037T 2D  AIThe SynGAP1 missense variant I1037T is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33443662‑T‑C). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect are AlphaMissense‑Default and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains Likely Benign; Foldetta results are unavailable. Overall, the majority of predictions (seven benign vs. two pathogenic) indicate that the variant is most likely benign, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.939629 | Disordered | 0.986140 | Binding | 0.309 | 0.774 | 0.625 | 6-33443662-T-C | 1 | 6.21e-7 | -2.565 | Likely Benign | 0.973 | Likely Pathogenic | Likely Pathogenic | 0.066 | Likely Benign | 0.40 | Neutral | 0.292 | Benign | 0.110 | Benign | 2.79 | Benign | 0.34 | Tolerated | 3.77 | 5 | 0.1071 | 0.2175 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||
| c.3110T>G | I1037S 2D AIThe SynGAP1 missense variant I1037S is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), which classifies the variant as Likely Benign. Only AlphaMissense‑Default predicts a pathogenic outcome, while AlphaMissense‑Optimized is uncertain and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta) has no available result for this variant. Based on the overall consensus of the majority of tools, the variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.939629 | Disordered | 0.986140 | Binding | 0.309 | 0.774 | 0.625 | -2.247 | Likely Benign | 0.935 | Likely Pathogenic | Ambiguous | 0.120 | Likely Benign | 0.43 | Neutral | 0.032 | Benign | 0.017 | Benign | 2.83 | Benign | 0.27 | Tolerated | 0.2634 | 0.1110 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3116T>C | I1039T 2D AIThe SynGAP1 missense variant I1039T is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443668‑T‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a likely benign outcome. Only AlphaMissense‑Default predicts a pathogenic effect. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while a Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.979204 | Binding | 0.292 | 0.806 | 0.625 | Uncertain | 2 | 6-33443668-T-C | 12 | 7.43e-6 | -2.465 | Likely Benign | 0.645 | Likely Pathogenic | Likely Benign | 0.193 | Likely Benign | 0.45 | Neutral | 0.004 | Benign | 0.008 | Benign | 2.75 | Benign | 0.10 | Tolerated | 3.77 | 5 | 0.1248 | 0.2293 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||
| c.3116T>G | I1039S 2D AIThe SynGAP1 missense variant I1039S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM all classify the substitution as benign. In contrast, SIFT and AlphaMissense‑Default predict a pathogenic impact. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely benign outcome. No Foldetta stability assessment is available. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.969315 | Disordered | 0.979204 | Binding | 0.292 | 0.806 | 0.625 | -1.688 | Likely Benign | 0.829 | Likely Pathogenic | Ambiguous | 0.171 | Likely Benign | -0.20 | Neutral | 0.032 | Benign | 0.008 | Benign | 2.76 | Benign | 0.03 | Affected | 0.2917 | 0.1294 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3119G>T | G1040V 2D AIThe SynGAP1 missense variant G1040V is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443671‑G‑T). Prediction tools that agree on a benign effect are ESM1b and AlphaMissense‑Optimized; those that predict a pathogenic effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta results are unavailable. Overall, the majority of predictions indicate a pathogenic impact, and this is not in conflict with the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.964893 | Disordered | 0.973805 | Binding | 0.332 | 0.816 | 0.625 | Uncertain | 1 | 6-33443671-G-T | 4 | 2.48e-6 | -3.453 | Likely Benign | 0.645 | Likely Pathogenic | Likely Benign | 0.774 | Likely Pathogenic | -2.89 | Deleterious | 0.827 | Possibly Damaging | 0.456 | Possibly Damaging | -0.74 | Pathogenic | 0.01 | Affected | 3.77 | 5 | 0.1239 | 0.4213 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||
| c.3128G>C | R1043T 2D  AIThe SynGAP1 missense variant R1043T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978672 | Disordered | 0.954069 | Binding | 0.299 | 0.853 | 0.625 | -3.928 | Likely Benign | 0.298 | Likely Benign | Likely Benign | 0.463 | Likely Benign | -1.77 | Neutral | 0.001 | Benign | 0.003 | Benign | 5.39 | Benign | 0.00 | Affected | 0.1975 | 0.5513 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3129G>C | R1043S 2D  AIThe SynGAP1 missense variant R1043S is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools cluster into two groups: benign predictions include PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from REVEL and SIFT. AlphaMissense‑Default is uncertain, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign effect. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized scores benign, SGM‑Consensus is likely benign, and Foldetta results are unavailable. Taken together, the majority of evidence points to a benign impact for R1043S. This conclusion is consistent with the absence of a ClinVar assertion, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978672 | Disordered | 0.954069 | Binding | 0.299 | 0.853 | 0.625 | -3.223 | Likely Benign | 0.457 | Ambiguous | Likely Benign | 0.509 | Likely Pathogenic | -2.10 | Neutral | 0.036 | Benign | 0.018 | Benign | 5.42 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2727 | 0.4628 | -1 | 0 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||
| c.3129G>T | R1043S 2D AIThe SynGAP1 missense variant R1043S is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443681‑G‑T). Prediction tools that agree on a benign effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are REVEL and SIFT. AlphaMissense‑Default remains uncertain, and no Foldetta stability result is available. High‑accuracy assessments: AlphaMissense‑Optimized classifies the variant as benign; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome; Foldetta data are missing, so it does not contribute to the evaluation. Based on the collective predictions, the variant is most likely benign, which is consistent with its ClinVar “Uncertain” classification and does not contradict the available evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978672 | Disordered | 0.954069 | Binding | 0.299 | 0.853 | 0.625 | Uncertain | 1 | 6-33443681-G-T | 2 | 1.24e-6 | -3.223 | Likely Benign | 0.457 | Ambiguous | Likely Benign | 0.509 | Likely Pathogenic | -2.10 | Neutral | 0.036 | Benign | 0.018 | Benign | 5.42 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2727 | 0.4628 | -1 | 0 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||
| c.3130C>A | P1044T 2D  AIThe SynGAP1 missense variant P1044T is catalogued in gnomAD (ID 6‑33443682‑C‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.979741 | Disordered | 0.952126 | Binding | 0.331 | 0.855 | 0.750 | 6-33443682-C-A | 1 | 6.20e-7 | -4.605 | Likely Benign | 0.075 | Likely Benign | Likely Benign | 0.372 | Likely Benign | -1.02 | Neutral | 0.126 | Benign | 0.096 | Benign | 5.53 | Benign | 0.17 | Tolerated | 3.77 | 5 | 0.1681 | 0.6860 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||
| c.3136C>G | P1046A 2D  AIThe SynGAP1 missense variant P1046A is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443688‑C‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; a Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.970265 | Disordered | 0.942366 | Binding | 0.364 | 0.898 | 0.750 | Uncertain | 1 | 6-33443688-C-G | 1 | 6.20e-7 | -3.246 | Likely Benign | 0.048 | Likely Benign | Likely Benign | 0.041 | Likely Benign | -1.67 | Neutral | 0.001 | Benign | 0.008 | Benign | 2.39 | Pathogenic | 0.29 | Tolerated | 3.77 | 5 | 0.2976 | 0.5358 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||
| c.313T>C | S105P 2D AIThe SynGAP1 missense variant S105P is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only two tools—polyPhen‑2 HumDiv and SIFT—predict a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as likely benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates likely benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence points to a benign effect, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.669201 | Binding | 0.364 | 0.870 | 0.625 | Uncertain | 1 | -3.631 | Likely Benign | 0.166 | Likely Benign | Likely Benign | 0.204 | Likely Benign | 0.03 | Neutral | 0.808 | Possibly Damaging | 0.212 | Benign | 4.00 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2236 | 0.4584 | -1 | 1 | -0.8 | 10.04 | |||||||||||||||||||||||||||||||||||
| c.3143G>T | G1048V 2D AIThe SynGAP1 missense variant G1048V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.977651 | Disordered | 0.923876 | Binding | 0.346 | 0.916 | 0.750 | -6.108 | Likely Benign | 0.079 | Likely Benign | Likely Benign | 0.520 | Likely Pathogenic | -0.59 | Neutral | 0.958 | Probably Damaging | 0.787 | Possibly Damaging | 2.54 | Benign | 0.11 | Tolerated | 0.1312 | 0.3688 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3145C>T | P1049S 2D AIThe SynGAP1 missense variant P1049S is reported in gnomAD (variant ID 6‑33443697‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. No Foldetta stability data are available, so it does not influence the assessment. Overall, the majority of predictions, including the high‑accuracy tools, indicate that P1049S is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978316 | Disordered | 0.917915 | Binding | 0.428 | 0.920 | 0.750 | 6-33443697-C-T | 2 | 1.24e-6 | -2.351 | Likely Benign | 0.053 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -1.53 | Neutral | 0.519 | Possibly Damaging | 0.303 | Benign | 2.76 | Benign | 0.04 | Affected | 3.77 | 5 | 0.3144 | 0.5083 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3149G>T | G1050V 2D AIThe SynGAP1 missense variant G1050V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; the Foldetta protein‑folding stability analysis is unavailable for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently contains no classification for G1050V. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.987317 | Disordered | 0.906802 | Binding | 0.370 | 0.928 | 0.875 | -6.450 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.066 | Likely Benign | -0.83 | Neutral | 0.126 | Benign | 0.096 | Benign | 2.49 | Pathogenic | 0.13 | Tolerated | 0.1260 | 0.3684 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3152G>A | G1051D 2D AISynGAP1 missense variant G1051D is listed in ClinVar as Benign and is present in gnomAD (variant ID 6‑33443704‑G‑A). Prediction tools that classify the variant as benign include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict pathogenicity are polyPhen‑2 HumDiv, ESM1b, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign versus two pathogenic votes), and Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a benign effect, consistent with the ClinVar annotation and not contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.987317 | Disordered | 0.900141 | Binding | 0.358 | 0.936 | 0.875 | Benign | 1 | 6-33443704-G-A | 2 | 1.24e-6 | -9.379 | Likely Pathogenic | 0.311 | Likely Benign | Likely Benign | 0.445 | Likely Benign | -0.31 | Neutral | 0.761 | Possibly Damaging | 0.239 | Benign | -0.74 | Pathogenic | 0.39 | Tolerated | 3.77 | 5 | 0.1872 | 0.2235 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||
| c.3155G>T | G1052V 2D AIThe SynGAP1 missense variant G1052V is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, PROVEAN, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate benign, while the high‑accuracy AlphaMissense‑Optimized score is benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Benign. In contrast, polyPhen‑2 HumDiv and HumVar both predict pathogenic, and ESM1b remains uncertain. No Foldetta stability assessment is available, so it does not influence the overall interpretation. Overall, the majority of evidence points to a benign effect, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.984420 | Disordered | 0.892068 | Binding | 0.367 | 0.938 | 0.875 | -7.717 | In-Between | 0.094 | Likely Benign | Likely Benign | 0.452 | Likely Benign | -0.12 | Neutral | 0.901 | Possibly Damaging | 0.619 | Possibly Damaging | 3.90 | Benign | 0.19 | Tolerated | 0.1329 | 0.3499 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3157A>C | S1053R 2D AIThe SynGAP1 missense variant S1053R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign, while the majority‑vote SGM‑Consensus also reports a likely benign outcome. In contrast, the two polyPhen‑2 implementations (HumDiv and HumVar) predict a pathogenic impact. The AlphaMissense‑Default score is uncertain, and no Foldetta stability assessment is available. High‑accuracy analyses reinforce the benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely benign, with no contradictory Foldetta data. Overall, the preponderance of evidence points to a benign effect for S1053R, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.981594 | Disordered | 0.885608 | Binding | 0.399 | 0.944 | 0.875 | -6.421 | Likely Benign | 0.359 | Ambiguous | Likely Benign | 0.225 | Likely Benign | 0.43 | Neutral | 0.969 | Probably Damaging | 0.581 | Possibly Damaging | 5.33 | Benign | 0.59 | Tolerated | 3.77 | 5 | 0.1321 | 0.3820 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3157A>T | S1053C 2D 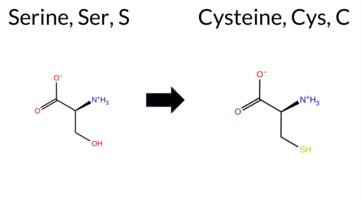 AIThe SynGAP1 missense variant S1053C is catalogued in gnomAD (ID 6‑33443709‑A‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). In contrast, polyPhen‑2 (HumDiv and HumVar) predict a pathogenic impact. The ESM1b score is uncertain, providing no clear direction. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant. Overall, the majority of reliable predictors classify S1053C as benign, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.981594 | Disordered | 0.885608 | Binding | 0.399 | 0.944 | 0.875 | 6-33443709-A-T | -7.574 | In-Between | 0.095 | Likely Benign | Likely Benign | 0.220 | Likely Benign | -0.61 | Neutral | 0.977 | Probably Damaging | 0.777 | Possibly Damaging | 5.30 | Benign | 0.11 | Tolerated | 3.77 | 5 | 0.1675 | 0.5895 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||
| c.3158G>T | S1053I 2D AIThe SynGAP1 missense variant S1053I is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence indicates that S1053I is most likely benign, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.981594 | Disordered | 0.885608 | Binding | 0.399 | 0.944 | 0.875 | -6.572 | Likely Benign | 0.177 | Likely Benign | Likely Benign | 0.250 | Likely Benign | -0.46 | Neutral | 0.925 | Possibly Damaging | 0.413 | Benign | 5.32 | Benign | 0.10 | Tolerated | 0.1536 | 0.4780 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3159C>A | S1053R 2D AIThe SynGAP1 missense variant S1053R is reported in gnomAD (variant ID 6‑33443711‑C‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. Overall, the majority of high‑accuracy predictors and consensus analyses indicate a benign impact. Thus, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.981594 | Disordered | 0.885608 | Binding | 0.399 | 0.944 | 0.875 | 6-33443711-C-A | 1 | 1.10e-6 | -6.421 | Likely Benign | 0.359 | Ambiguous | Likely Benign | 0.303 | Likely Benign | 0.43 | Neutral | 0.969 | Probably Damaging | 0.581 | Possibly Damaging | 5.33 | Benign | 0.59 | Tolerated | 3.77 | 5 | 0.1321 | 0.3820 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||
| c.3159C>G | S1053R 2D AIThe SynGAP1 missense variant S1053R is reported in gnomAD (ID 6‑33443711‑C‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain, and the high‑accuracy consensus methods give a benign verdict: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports “Likely Benign.” Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. The predictions do not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.981594 | Disordered | 0.885608 | Binding | 0.399 | 0.944 | 0.875 | 6-33443711-C-G | -6.421 | Likely Benign | 0.359 | Ambiguous | Likely Benign | 0.304 | Likely Benign | 0.43 | Neutral | 0.969 | Probably Damaging | 0.581 | Possibly Damaging | 5.33 | Benign | 0.59 | Tolerated | 3.77 | 5 | 0.1321 | 0.3820 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.3161G>T | G1054V 2D AIThe SynGAP1 missense variant G1054V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv indicates a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely benign. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts benign and the SGM‑Consensus also reports likely benign; Foldetta results are unavailable. Overall, the consensus of the available predictions points to a benign impact, and this is consistent with the lack of ClinVar evidence; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.983019 | Disordered | 0.878015 | Binding | 0.389 | 0.936 | 0.875 | -6.994 | Likely Benign | 0.110 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -0.22 | Neutral | 0.818 | Possibly Damaging | 0.221 | Benign | 4.01 | Benign | 0.18 | Tolerated | 0.1578 | 0.3694 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3164G>T | G1055V 2D AIThe SynGAP1 missense variant G1055V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while ESM1b remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.982235 | Disordered | 0.872113 | Binding | 0.379 | 0.935 | 0.875 | -7.434 | In-Between | 0.114 | Likely Benign | Likely Benign | 0.399 | Likely Benign | 0.26 | Neutral | 0.818 | Possibly Damaging | 0.222 | Benign | 3.28 | Benign | 0.17 | Tolerated | 0.1399 | 0.3694 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3167G>T | G1056V 2D AIThe SynGAP1 missense variant G1056V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence (7 benign vs 2 pathogenic) supports a benign classification. This consensus does not contradict ClinVar status, which has no entry for this variant. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.988291 | Disordered | 0.868632 | Binding | 0.402 | 0.935 | 0.875 | -8.130 | Likely Pathogenic | 0.097 | Likely Benign | Likely Benign | 0.448 | Likely Benign | -0.24 | Neutral | 0.292 | Benign | 0.110 | Benign | 1.83 | Pathogenic | 0.08 | Tolerated | 0.1488 | 0.3507 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3169A>C | S1057R 2D AIThe SynGAP1 missense variant S1057R is catalogued in gnomAD (ID 6‑33443721‑A‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.869507 | Binding | 0.413 | 0.927 | 0.875 | 6-33443721-A-C | -6.648 | Likely Benign | 0.379 | Ambiguous | Likely Benign | 0.221 | Likely Benign | -0.24 | Neutral | 0.677 | Possibly Damaging | 0.168 | Benign | 5.30 | Benign | 0.21 | Tolerated | 3.77 | 5 | 0.1584 | 0.3620 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.3169A>T | S1057C 2D AIThe SynGAP1 missense variant S1057C is reported in gnomAD (ID 6‑33443721‑A‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Two tools, polyPhen‑2 HumDiv and polyPhen‑2 HumVar, predict a pathogenic impact, while ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar classification (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.869507 | Binding | 0.413 | 0.927 | 0.875 | 6-33443721-A-T | -7.529 | In-Between | 0.100 | Likely Benign | Likely Benign | 0.258 | Likely Benign | -0.64 | Neutral | 0.977 | Probably Damaging | 0.683 | Possibly Damaging | 5.23 | Benign | 0.10 | Tolerated | 3.77 | 5 | 0.1856 | 0.6106 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||
| c.3170G>T | S1057I 2D AIThe SynGAP1 missense variant S1057I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to the variant being most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.869507 | Binding | 0.413 | 0.927 | 0.875 | -6.887 | Likely Benign | 0.186 | Likely Benign | Likely Benign | 0.259 | Likely Benign | -0.87 | Neutral | 0.925 | Possibly Damaging | 0.238 | Benign | 5.24 | Benign | 0.07 | Tolerated | 0.1802 | 0.4980 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3171C>A | S1057R 2D AIThe SynGAP1 missense variant S1057R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Only polyPhen‑2 HumDiv flags it as pathogenic, while AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.869507 | Binding | 0.413 | 0.927 | 0.875 | -6.648 | Likely Benign | 0.379 | Ambiguous | Likely Benign | 0.272 | Likely Benign | -0.24 | Neutral | 0.677 | Possibly Damaging | 0.168 | Benign | 5.30 | Benign | 0.21 | Tolerated | 3.77 | 5 | 0.1584 | 0.3620 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3171C>G | S1057R 2D AIThe SynGAP1 missense variant S1057R is catalogued in gnomAD (ID 6‑33443723‑C‑G) but has no ClinVar submission. Functional prediction tools largely converge on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all report benign or tolerated. Only polyPhen‑2 HumDiv flags it as pathogenic, while AlphaMissense‑Default remains uncertain. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus confirms a benign likelihood; Foldetta data are unavailable, so no stability evidence is provided. Taken together, the preponderance of evidence indicates the variant is most likely benign, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.988291 | Disordered | 0.869507 | Binding | 0.413 | 0.927 | 0.875 | 6-33443723-C-G | -6.648 | Likely Benign | 0.379 | Ambiguous | Likely Benign | 0.272 | Likely Benign | -0.24 | Neutral | 0.677 | Possibly Damaging | 0.168 | Benign | 5.30 | Benign | 0.21 | Tolerated | 3.77 | 5 | 0.1584 | 0.3620 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.3173G>A | G1058D 2D AIThe SynGAP1 missense variant G1058D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443725‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and ESM1b, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.980739 | Disordered | 0.885724 | Binding | 0.407 | 0.929 | 0.875 | 6-33443725-G-A | 1 | 6.21e-7 | -10.344 | Likely Pathogenic | 0.391 | Ambiguous | Likely Benign | 0.177 | Likely Benign | -0.33 | Neutral | 0.077 | Benign | 0.042 | Benign | 5.20 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1889 | 0.2435 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||
| c.3176G>T | G1059V 2D AIThe SynGAP1 missense variant G1059V is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in silico predictors indicates a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score benign, while the majority‑vote SGM‑Consensus also classifies it as likely benign. Only SIFT predicts a pathogenic outcome, and ESM1b remains uncertain. High‑accuracy tools corroborate the benign prediction: AlphaMissense‑Optimized is benign and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely benign; Foldetta results are not available. Overall, the preponderance of evidence supports a benign classification for G1059V, and this assessment does not conflict with ClinVar, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.983019 | Disordered | 0.898939 | Binding | 0.399 | 0.926 | 0.875 | -7.242 | In-Between | 0.106 | Likely Benign | Likely Benign | 0.478 | Likely Benign | -0.82 | Neutral | 0.259 | Benign | 0.066 | Benign | 2.54 | Benign | 0.00 | Affected | 0.1462 | 0.3494 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3179G>T | G1060V 2D AIThe SynGAP1 missense variant G1060V is listed in ClinVar as benign (ClinVar ID 1345112.0) and is observed in gnomAD (6‑33443731‑G‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Two tools, polyPhen‑2 HumDiv and HumVar, predict a pathogenic effect. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as likely benign, and AlphaMissense‑Optimized also reports a benign outcome. No Foldetta stability assessment is available for this variant. Overall, the majority of evidence points to a benign impact, aligning with the ClinVar designation and not contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.979242 | Disordered | 0.913048 | Binding | 0.407 | 0.928 | 0.875 | Benign | 1 | 6-33443731-G-T | 1 | 6.22e-7 | -6.966 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 0.369 | Likely Benign | -0.73 | Neutral | 0.986 | Probably Damaging | 0.728 | Possibly Damaging | 2.63 | Benign | 0.33 | Tolerated | 4.32 | 2 | 0.1453 | 0.3494 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||
| c.317G>C | R106T 2D AIThe SynGAP1 missense variant R106T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. Separately, the high‑accuracy consensus (SGM‑Consensus) classifies the variant as Likely Benign, while AlphaMissense‑Optimized remains uncertain and Foldetta data are missing. Based on the overall pattern of predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.675549 | Disordered | 0.663409 | Binding | 0.345 | 0.862 | 0.875 | -4.197 | Likely Benign | 0.950 | Likely Pathogenic | Ambiguous | 0.229 | Likely Benign | -2.30 | Neutral | 0.004 | Benign | 0.002 | Benign | 3.67 | Benign | 0.00 | Affected | 0.1941 | 0.4742 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3182G>T | G1061V 2D AIThe SynGAP1 missense variant G1061V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign. Foldetta results are unavailable. Overall, the majority of evidence indicates that G1061V is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.978672 | Disordered | 0.926729 | Binding | 0.394 | 0.923 | 0.875 | -6.709 | Likely Benign | 0.106 | Likely Benign | Likely Benign | 0.307 | Likely Benign | -1.41 | Neutral | 0.224 | Benign | 0.066 | Benign | 3.98 | Benign | 0.00 | Affected | 0.1431 | 0.3684 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3185G>T | G1062V 2D AIThe SynGAP1 missense variant G1062V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.976962 | Disordered | 0.936972 | Binding | 0.368 | 0.917 | 0.875 | -6.598 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 0.377 | Likely Benign | -0.78 | Neutral | 0.259 | Benign | 0.066 | Benign | 4.12 | Benign | 0.01 | Affected | 0.1441 | 0.3694 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3188G>T | G1063V 2D AIThe SynGAP1 missense variant G1063V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority of the high‑accuracy tools) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and high‑accuracy tools indicates that G1063V is most likely benign, with no ClinVar status to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.975134 | Disordered | 0.945784 | Binding | 0.394 | 0.913 | 0.875 | -6.228 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.045 | Likely Benign | -0.82 | Neutral | 0.004 | Benign | 0.002 | Benign | 4.29 | Benign | 0.03 | Affected | 0.1434 | 0.3707 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3209G>C | R1070T 2D AIThe SynGAP1 missense variant R1070T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain” and the SGM‑Consensus as “Likely Benign”; Foldetta results are not available. Overall, the balance of evidence leans toward a benign impact, with no ClinVar entry to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.982693 | Binding | 0.297 | 0.906 | 0.875 | -5.093 | Likely Benign | 0.860 | Likely Pathogenic | Ambiguous | 0.144 | Likely Benign | -2.35 | Neutral | 0.948 | Possibly Damaging | 0.507 | Possibly Damaging | 3.78 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1634 | 0.4727 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||
| c.3209_3210delinsCA | R1070T 2D AIThe SynGAP1 missense variant R1070T is listed in ClinVar (ID 2759838.0) with an “Uncertain” clinical significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include PROVEAN, ESM1b, FATHMM, and the SGM‑Consensus (which aggregates these three benign calls with the pathogenic AlphaMissense‑Default to yield a Likely Benign verdict). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized returns an uncertain result. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Benign, and no Foldetta stability data is available. Overall, the balance of evidence leans toward a benign impact, which is consistent with the ClinVar “Uncertain” status and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.982693 | Binding | 0.297 | 0.906 | 0.875 | Uncertain | 1 | -5.093 | Likely Benign | 0.860 | Likely Pathogenic | Ambiguous | -2.35 | Neutral | 0.948 | Possibly Damaging | 0.507 | Possibly Damaging | 3.78 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1634 | 0.4727 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||
| c.320G>C | R107T 2D AIThe SynGAP1 missense variant R107T is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (2 pathogenic vs. 2 benign) and Foldetta results are unavailable. Overall, the majority of predictions (five benign vs. four pathogenic) lean toward a benign interpretation. Thus, the variant is most likely benign based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.666105 | Disordered | 0.663448 | Binding | 0.331 | 0.863 | 0.875 | -2.902 | Likely Benign | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.191 | Likely Benign | -2.63 | Deleterious | 0.421 | Benign | 0.050 | Benign | 2.98 | Benign | 0.00 | Affected | 0.1549 | 0.4761 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3217T>C | S1073P 2D AIThe SynGAP1 missense variant S1073P is reported in gnomAD (variant ID 6‑33443769‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign effect. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for S1073P, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.916840 | Disordered | 0.985818 | Binding | 0.313 | 0.905 | 0.750 | 6-33443769-T-C | 1 | 6.23e-7 | -4.520 | Likely Benign | 0.338 | Likely Benign | Likely Benign | 0.082 | Likely Benign | -0.76 | Neutral | 0.006 | Benign | 0.008 | Benign | 3.85 | Benign | 0.01 | Affected | 3.77 | 5 | 0.2243 | 0.5900 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.3218C>G | S1073C 2D AISynGAP1 missense variant S1073C is recorded in gnomAD (ID 6‑33443770‑C‑G) but has no ClinVar submission. Functional prediction tools show mixed results: benign calls come from REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic calls are made by polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The AlphaMissense‑Default score is uncertain. A consensus from the SGM framework (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because the votes are split. High‑accuracy methods give a benign prediction from AlphaMissense‑Optimized; the SGM Consensus remains ambiguous, and Foldetta stability analysis is unavailable. Consequently, the evidence does not strongly favor either outcome. The variant is most likely of uncertain significance, with no contradiction to ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.916840 | Disordered | 0.985818 | Binding | 0.313 | 0.905 | 0.750 | 6-33443770-C-G | 1 | 6.23e-7 | -8.862 | Likely Pathogenic | 0.461 | Ambiguous | Likely Benign | 0.137 | Likely Benign | -1.52 | Neutral | 0.997 | Probably Damaging | 0.840 | Possibly Damaging | 3.81 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1343 | 0.6088 | -1 | 0 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||
| c.3221A>C | Q1074P 2D  AIThe SynGAP1 missense variant Q1074P is listed in gnomAD (ID 6‑33443773‑A‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single outlier. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.930790 | Disordered | 0.987006 | Binding | 0.339 | 0.897 | 0.750 | 6-33443773-A-C | 1 | 6.23e-7 | -4.259 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.188 | Likely Benign | 0.52 | Neutral | 0.925 | Possibly Damaging | 0.432 | Benign | 2.66 | Benign | 0.13 | Tolerated | 3.77 | 5 | 0.2046 | 0.5784 | -1 | 0 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||
| c.3230C>G | T1077R 2D  AIThe SynGAP1 missense variant T1077R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), which collectively classify the variant as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default predict a pathogenic impact. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and no Foldetta stability assessment is available. Overall, the balance of evidence leans toward a benign interpretation, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.988141 | Binding | 0.329 | 0.892 | 0.750 | -4.109 | Likely Benign | 0.890 | Likely Pathogenic | Ambiguous | 0.121 | Likely Benign | -1.01 | Neutral | 0.970 | Probably Damaging | 0.728 | Possibly Damaging | 4.18 | Benign | 0.03 | Affected | 0.1028 | 0.3491 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3230C>T | T1077I 2D AIThe SynGAP1 missense variant T1077I is listed in gnomAD (ID 6‑33443782‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions (REVEL, PROVEAN, SIFT, ESM1b, FATHMM) and pathogenic predictions (PolyPhen‑2 HumDiv, PolyPhen‑2 HumVar, AlphaMissense‑Default). The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to a likely benign verdict (3 benign vs. 1 pathogenic). High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized returns an uncertain result, while Foldetta (combining FoldX‑MD and Rosetta outputs) is not available for this residue. Taken together, the majority of evidence points toward a benign effect. This conclusion is consistent with the absence of a ClinVar pathogenic classification, and there is no contradiction with existing database annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.903857 | Disordered | 0.988141 | Binding | 0.329 | 0.892 | 0.750 | 6-33443782-C-T | 1 | 6.25e-7 | -4.710 | Likely Benign | 0.919 | Likely Pathogenic | Ambiguous | 0.155 | Likely Benign | -1.11 | Neutral | 0.970 | Probably Damaging | 0.787 | Possibly Damaging | 4.19 | Benign | 0.33 | Tolerated | 3.77 | 5 | 0.1135 | 0.5870 | -1 | 0 | 5.2 | 12.05 | ||||||||||||||||||||||||||||||||||
| c.3232G>T | V1078F 2D AIThe SynGAP1 missense variant V1078F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect for V1078F, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.986989 | Binding | 0.294 | 0.898 | 0.750 | -3.768 | Likely Benign | 0.570 | Likely Pathogenic | Likely Benign | 0.142 | Likely Benign | -0.97 | Neutral | 0.977 | Probably Damaging | 0.722 | Possibly Damaging | 3.87 | Benign | 0.02 | Affected | 0.0677 | 0.4561 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3233T>G | V1078G 2D AIThe SynGAP1 missense variant V1078G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign (3 benign vs 1 pathogenic). Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence indicates a benign impact. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.882776 | Disordered | 0.986989 | Binding | 0.294 | 0.898 | 0.750 | -3.270 | Likely Benign | 0.699 | Likely Pathogenic | Likely Benign | 0.168 | Likely Benign | -0.54 | Neutral | 0.157 | Benign | 0.292 | Benign | 3.85 | Benign | 0.00 | Affected | 0.2041 | 0.2693 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3236G>T | S1079I 2D AIThe SynGAP1 missense variant S1079I is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.919029 | Disordered | 0.983887 | Binding | 0.307 | 0.900 | 0.750 | -4.732 | Likely Benign | 0.688 | Likely Pathogenic | Likely Benign | 0.093 | Likely Benign | -2.86 | Deleterious | 0.078 | Benign | 0.025 | Benign | 3.83 | Benign | 0.00 | Affected | 0.0921 | 0.4775 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3239C>A | A1080E 2D  AIThe SynGAP1 missense variant A1080E is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33443791‑C‑A). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM. Tools that predict a pathogenic outcome are polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. High‑accuracy evidence therefore points to a benign or uncertain impact: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus is benign, and Foldetta data are missing. Overall, the balance of predictions leans toward a benign effect, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.981457 | Binding | 0.303 | 0.900 | 0.750 | 6-33443791-C-A | -3.672 | Likely Benign | 0.855 | Likely Pathogenic | Ambiguous | 0.090 | Likely Benign | -1.50 | Neutral | 0.901 | Possibly Damaging | 0.540 | Possibly Damaging | 4.00 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1394 | 0.2437 | -1 | 0 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||
| c.3248A>C | K1083T 2D  AIThe SynGAP1 missense variant K1083T is reported in gnomAD (ID 6‑33443800‑A‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are PolyPhen‑2 HumDiv, PolyPhen‑2 HumVar, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.837511 | Disordered | 0.978906 | Binding | 0.302 | 0.893 | 1.000 | 6-33443800-A-C | 2 | 1.26e-6 | -2.870 | Likely Benign | 0.690 | Likely Pathogenic | Likely Benign | 0.233 | Likely Benign | -0.76 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 4.05 | Benign | 0.31 | Tolerated | 3.77 | 5 | 0.2192 | 0.4150 | -1 | 0 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||||
| c.3250C>A | P1084T 2D AIThe SynGAP1 missense variant P1084T is reported in ClinVar as “Not submitted” and is present in gnomAD (ID 6‑33443802‑C‑A). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it as pathogenic, but this is the sole discordant call. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign; Foldetta results are not available. Overall, the preponderance of evidence indicates the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently contains no pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.842060 | Disordered | 0.979020 | Binding | 0.348 | 0.889 | 1.000 | 6-33443802-C-A | -4.665 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.127 | Likely Benign | -2.14 | Neutral | 0.025 | Benign | 0.012 | Benign | 4.11 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1546 | 0.6793 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3250C>G | P1084A 2D AIThe SynGAP1 missense variant P1084A is listed in ClinVar (ID 2827308.0) with an “Uncertain” status and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which itself is “Likely Benign”). In contrast, PROVEAN and polyPhen‑2 HumDiv predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; a Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.842060 | Disordered | 0.979020 | Binding | 0.348 | 0.889 | 1.000 | Uncertain | 1 | -3.928 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -2.54 | Deleterious | 0.649 | Possibly Damaging | 0.157 | Benign | 4.05 | Benign | 0.35 | Tolerated | 3.77 | 5 | 0.3164 | 0.5784 | -1 | 1 | 3.4 | -26.04 | |||||||||||||||||||||||||||||||||||
| c.3250C>T | P1084S 2D AIThe SynGAP1 missense variant P1084S is reported in gnomAD (variant ID 6‑33443802‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 HumDiv and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.842060 | Disordered | 0.979020 | Binding | 0.348 | 0.889 | 1.000 | 6-33443802-C-T | 1 | 6.31e-7 | -3.987 | Likely Benign | 0.119 | Likely Benign | Likely Benign | 0.086 | Likely Benign | -2.24 | Neutral | 0.481 | Possibly Damaging | 0.157 | Benign | 4.03 | Benign | 0.03 | Affected | 3.77 | 5 | 0.3102 | 0.6215 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3256C>A | P1086T 2D AIThe SynGAP1 missense variant P1086T is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443808‑C‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta results are unavailable. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic impact. The variant is most likely pathogenic based on the available computational evidence, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.849326 | Disordered | 0.977190 | Binding | 0.393 | 0.885 | 1.000 | 6-33443808-C-A | -4.181 | Likely Benign | 0.568 | Likely Pathogenic | Likely Benign | 0.229 | Likely Benign | -2.97 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.75 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1343 | 0.5848 | -1 | 0 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||
| c.3256C>T | P1086S 2D AIThe SynGAP1 missense variant P1086S is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443808‑C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 pathogenic vs. 2 benign). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. four benign) lean toward a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.849326 | Disordered | 0.977190 | Binding | 0.393 | 0.885 | 1.000 | 6-33443808-C-T | -3.165 | Likely Benign | 0.604 | Likely Pathogenic | Likely Benign | 0.212 | Likely Benign | -3.04 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.78 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3067 | 0.4894 | -1 | 1 | 0.8 | -10.04 | |||||||||||||||||||||||||||||||||||||
| c.3257C>A | P1086Q 2D  AIThe SynGAP1 missense variant P1086Q is not reported in ClinVar (ClinVar status: None) but is present in gnomAD (ID 6‑33443809‑C‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two pathogenic vs. two benign votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of predictions (five pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.849326 | Disordered | 0.977190 | Binding | 0.393 | 0.885 | 1.000 | 6-33443809-C-A | -4.668 | Likely Benign | 0.652 | Likely Pathogenic | Likely Benign | 0.185 | Likely Benign | -2.92 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.77 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1285 | 0.4784 | -1 | 0 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||
| c.3259T>C | S1087P 2D AIThe SynGAP1 missense variant S1087P is reported in gnomAD (ID 6‑33443811‑T‑C) and has no ClinVar entry. All available in silico predictors classify it as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Based on the unanimous benign predictions and lack of ClinVar pathogenic annotation, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.974805 | Binding | 0.357 | 0.891 | 1.000 | 6-33443811-T-C | 1 | 6.34e-7 | -2.946 | Likely Benign | 0.133 | Likely Benign | Likely Benign | 0.135 | Likely Benign | -1.92 | Neutral | 0.006 | Benign | 0.008 | Benign | 2.56 | Benign | 0.12 | Tolerated | 3.77 | 5 | 0.1803 | 0.5700 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||||
| c.3260C>G | S1087C 2D AIThe SynGAP1 missense variant S1087C is catalogued in gnomAD (ID 6‑33443812‑C‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact, while ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus also as likely benign; the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.912647 | Disordered | 0.974805 | Binding | 0.357 | 0.891 | 1.000 | 6-33443812-C-G | 1 | 6.34e-7 | -7.369 | In-Between | 0.194 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -2.22 | Neutral | 0.997 | Probably Damaging | 0.840 | Possibly Damaging | 2.55 | Benign | 0.05 | Affected | 3.77 | 5 | 0.0979 | 0.6118 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||
| c.3262A>C | S1088R 2D AIThe SynGAP1 missense variant S1088R is not reported in ClinVar and has no gnomAD entry. Prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, and FATHMM, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves as Likely Benign, reflecting the majority of benign calls. High‑accuracy assessments further support this: AlphaMissense‑Optimized labels the variant as Pathogenic, but the SGM‑Consensus (majority vote) remains Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the balance of evidence leans toward a benign effect; this conclusion does not conflict with ClinVar, which contains no entry for S1088R. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.975261 | Binding | 0.336 | 0.889 | 1.000 | -4.588 | Likely Benign | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.209 | Likely Benign | -1.96 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.72 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1190 | 0.4502 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3264C>A | S1088R 2D AIThe SynGAP1 missense variant S1088R is not reported in ClinVar and is present in gnomAD (ID 6‑33443816‑C‑A). Prediction tools that classify the variant as benign include REVEL, PROVEAN, ESM1b, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized predict it to be pathogenic. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains benign; Foldetta results are unavailable. Overall, the balance of evidence—five pathogenic versus four benign predictions—suggests the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for S1088R. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.975261 | Binding | 0.336 | 0.889 | 1.000 | 6-33443816-C-A | -4.588 | Likely Benign | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.181 | Likely Benign | -1.96 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.72 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1190 | 0.4502 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.3264C>G | S1088R 2D AIThe SynGAP1 missense variant S1088R has no ClinVar record and is not listed in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic, whereas the SGM‑Consensus remains Benign; Foldetta results are unavailable. Overall, the balance of evidence slightly favors a pathogenic interpretation, with no conflict with ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.975261 | Binding | 0.336 | 0.889 | 1.000 | -4.588 | Likely Benign | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.181 | Likely Benign | -1.96 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.72 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1190 | 0.4502 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3266G>T | G1089V 2D AIThe SynGAP1 missense variant G1089V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta results are unavailable. Overall, the majority of evidence points to a pathogenic impact for G1089V. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.891961 | Disordered | 0.976771 | Binding | 0.366 | 0.890 | 1.000 | -4.809 | Likely Benign | 0.527 | Ambiguous | Likely Benign | 0.182 | Likely Benign | -2.81 | Deleterious | 0.984 | Probably Damaging | 0.722 | Possibly Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.1326 | 0.4413 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.326G>T | S109I 2D AIThe SynGAP1 missense variant S109I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie, and Foldetta data are unavailable. Overall, the majority of evidence (five benign vs. three pathogenic predictions) points toward a benign impact. This conclusion does not contradict ClinVar, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.669335 | Binding | 0.328 | 0.864 | 0.750 | -5.195 | Likely Benign | 0.927 | Likely Pathogenic | Ambiguous | 0.200 | Likely Benign | -2.56 | Deleterious | 0.267 | Benign | 0.039 | Benign | 3.47 | Benign | 0.00 | Affected | 0.0910 | 0.4930 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3283C>T | P1095S 2D AIThe SynGAP1 missense variant P1095S is reported in gnomAD (ID 6‑33443835‑C‑T) and has no ClinVar entry. All evaluated in‑silico predictors classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Based on the unanimous benign predictions and the absence of any pathogenic signal, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.922952 | Disordered | 0.979251 | Binding | 0.387 | 0.870 | 1.000 | 6-33443835-C-T | 7 | 4.51e-6 | -3.819 | Likely Benign | 0.182 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -0.64 | Neutral | 0.207 | Benign | 0.072 | Benign | 2.80 | Benign | 1.00 | Tolerated | 3.77 | 5 | 0.3150 | 0.5963 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3287A>C | E1096A 2D  AIThe SynGAP1 missense variant E1096A is listed in ClinVar (ID 2579889.0) with an uncertain significance annotation and is not reported in gnomAD. Consensus from multiple in‑silico predictors shows a predominance of benign calls: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only polyPhen‑2 HumDiv assigns a pathogenic label, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates likely benign; Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the aggregate evidence points to a benign effect, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.926919 | Disordered | 0.976475 | Binding | 0.308 | 0.858 | 1.000 | Uncertain | 1 | -4.504 | Likely Benign | 0.510 | Ambiguous | Likely Benign | 0.164 | Likely Benign | -1.37 | Neutral | 0.626 | Possibly Damaging | 0.184 | Benign | 2.77 | Benign | 0.16 | Tolerated | 3.77 | 5 | 0.3805 | 0.7569 | -1 | 0 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||
| c.328G>T | V110F 2D AIThe SynGAP1 missense variant V110F is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, SGM‑Consensus is Likely Benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.622677 | Disordered | 0.665934 | Binding | 0.347 | 0.860 | 0.750 | -4.872 | Likely Benign | 0.397 | Ambiguous | Likely Benign | 0.042 | Likely Benign | -1.63 | Neutral | 0.006 | Benign | 0.003 | Benign | 4.13 | Benign | 0.01 | Affected | 0.0794 | 0.4027 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3290C>A | P1097Q 2D AIThe SynGAP1 missense variant P1097Q is reported in gnomAD (ID 6‑33443842‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict pathogenicity. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.974957 | Binding | 0.384 | 0.858 | 1.000 | 6-33443842-C-A | -4.765 | Likely Benign | 0.198 | Likely Benign | Likely Benign | 0.092 | Likely Benign | -1.10 | Neutral | 0.918 | Possibly Damaging | 0.604 | Possibly Damaging | 2.58 | Benign | 0.12 | Tolerated | 3.77 | 5 | 0.1613 | 0.5136 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.329T>G | V110G 2D AIThe SynGAP1 missense variant V110G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.622677 | Disordered | 0.665934 | Binding | 0.347 | 0.860 | 0.750 | -4.012 | Likely Benign | 0.724 | Likely Pathogenic | Likely Benign | 0.162 | Likely Benign | -2.87 | Deleterious | 0.377 | Benign | 0.928 | Probably Damaging | 4.06 | Benign | 0.00 | Affected | 0.2302 | 0.2671 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3301C>T | P1101S 2D AIThe SynGAP1 missense variant P1101S is reported in gnomAD (variant ID 6‑33443853‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant call. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is Likely Benign; Foldetta results are not available. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.874069 | Disordered | 0.968967 | Binding | 0.457 | 0.861 | 0.875 | 6-33443853-C-T | 1 | 6.52e-7 | -3.845 | Likely Benign | 0.080 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -1.31 | Neutral | 0.626 | Possibly Damaging | 0.255 | Benign | 4.25 | Benign | 0.07 | Tolerated | 3.77 | 5 | 0.3265 | 0.5400 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3304G>C | A1102P 2D  AIThe SynGAP1 missense variant A1102P is listed in ClinVar (ID 2789225.0) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.915074 | Disordered | 0.962659 | Binding | 0.388 | 0.859 | 0.875 | Uncertain | 1 | -5.120 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.118 | Likely Benign | -0.97 | Neutral | 0.000 | Benign | 0.002 | Benign | 2.26 | Pathogenic | 0.13 | Tolerated | 3.77 | 5 | 0.1978 | 0.5919 | -1 | 1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||
| c.3310C>A | P1104T 2D AIThe SynGAP1 missense variant P1104T is reported in gnomAD (variant ID 6‑33443862‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 (HumDiv and HumVar) predict it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. No Foldetta stability analysis is available, so it does not influence the conclusion. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar classification (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.936162 | Disordered | 0.954801 | Binding | 0.440 | 0.863 | 0.875 | 6-33443862-C-A | -3.995 | Likely Benign | 0.070 | Likely Benign | Likely Benign | 0.094 | Likely Benign | -0.14 | Neutral | 0.770 | Possibly Damaging | 0.481 | Possibly Damaging | 2.76 | Benign | 0.09 | Tolerated | 3.77 | 5 | 0.1549 | 0.6700 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3310C>T | P1104S 2D AIThe SynGAP1 missense variant P1104S is listed in ClinVar (ID 2912797.0) as Benign and is present in gnomAD (variant ID 6‑33443862‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Benign, and AlphaMissense‑Optimized also reports Benign. Foldetta results are not available. Overall, the majority of computational evidence supports a benign classification, which is consistent with the ClinVar status. Thus, the variant is most likely benign and does not contradict the ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.936162 | Disordered | 0.954801 | Binding | 0.440 | 0.863 | 0.875 | Benign | 1 | 6-33443862-C-T | 1 | 6.54e-7 | -2.330 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -0.30 | Neutral | 0.770 | Possibly Damaging | 0.404 | Benign | 2.77 | Benign | 0.10 | Tolerated | 3.77 | 5 | 0.3271 | 0.5746 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||
| c.3311C>A | P1104Q 2D AIThe SynGAP1 missense variant P1104Q is reported in gnomAD (ID 6‑33443863‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates a benign likelihood. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is assigned). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.936162 | Disordered | 0.954801 | Binding | 0.440 | 0.863 | 0.875 | 6-33443863-C-A | -3.161 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -0.64 | Neutral | 0.986 | Probably Damaging | 0.825 | Possibly Damaging | 2.68 | Benign | 0.06 | Tolerated | 3.77 | 5 | 0.1498 | 0.5063 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3332A>C | K1111T 2D AIThe SynGAP1 missense variant K1111T is listed in ClinVar (ID 4746140) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33443884‑A‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. No tool in the dataset predicts pathogenicity. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Overall, the collective predictions point to a benign effect, which does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.910643 | Disordered | 0.921455 | Binding | 0.300 | 0.902 | 0.875 | Uncertain | 1 | 6-33443884-A-C | 2 | 1.32e-6 | -4.037 | Likely Benign | 0.519 | Ambiguous | Likely Benign | 0.080 | Likely Benign | -0.90 | Neutral | 0.292 | Benign | 0.110 | Benign | 2.64 | Benign | 0.28 | Tolerated | 4.32 | 2 | 0.2250 | 0.4425 | -1 | 0 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||||
| c.3338G>A | G1113D 2D AIThe SynGAP1 missense variant G1113D is listed in ClinVar with an uncertain significance (ClinVar ID 2766136.0) and is present in gnomAD (variant ID 6‑33443890‑G‑A). Functional prediction tools that agree on a benign outcome include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM—all of which classify the substitution as benign. AlphaMissense‑Optimized also predicts a benign effect, whereas AlphaMissense‑Default remains uncertain. No tool predicts a pathogenic effect. The high‑accuracy consensus methods reinforce this benign assessment: the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign,” and AlphaMissense‑Optimized also indicates benign. Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the collective evidence strongly supports a benign impact, and this conclusion does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.905695 | Disordered | 0.900456 | Binding | 0.327 | 0.910 | 0.875 | Uncertain | 1 | 6-33443890-G-A | -4.638 | Likely Benign | 0.354 | Ambiguous | Likely Benign | 0.061 | Likely Benign | -0.72 | Neutral | 0.029 | Benign | 0.017 | Benign | 2.58 | Benign | 0.34 | Tolerated | 4.32 | 2 | 0.1872 | 0.2452 | -1 | 1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||
| c.3338G>T | G1113V 2D AIThe SynGAP1 missense variant G1113V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. The variant is most likely benign, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.905695 | Disordered | 0.900456 | Binding | 0.327 | 0.910 | 0.875 | -5.708 | Likely Benign | 0.093 | Likely Benign | Likely Benign | 0.088 | Likely Benign | -1.98 | Neutral | 0.827 | Possibly Damaging | 0.456 | Possibly Damaging | 2.53 | Benign | 0.11 | Tolerated | 0.1333 | 0.4224 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3340A>T | S1114C 2D AIThe SynGAP1 missense variant S1114C is reported in gnomAD (ID 6‑33443892‑A‑T) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.895196 | Binding | 0.295 | 0.908 | 0.875 | 6-33443892-A-T | -8.600 | Likely Pathogenic | 0.091 | Likely Benign | Likely Benign | 0.038 | Likely Benign | -1.77 | Neutral | 0.938 | Possibly Damaging | 0.552 | Possibly Damaging | 2.63 | Benign | 0.01 | Affected | 4.32 | 2 | 0.1101 | 0.5675 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||
| c.3350G>A | G1117D 2D AIThe SynGAP1 missense variant G1117D is catalogued in gnomAD (6‑33443902‑G‑A) but has no ClinVar entry. In silico predictors that agree on benign impact include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic effect, while ESM1b and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it contains two benign and two uncertain calls, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar classification (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.882776 | Disordered | 0.853192 | Binding | 0.323 | 0.914 | 0.750 | 6-33443902-G-A | 1 | 6.61e-7 | -7.594 | In-Between | 0.358 | Ambiguous | Likely Benign | 0.315 | Likely Benign | -0.38 | Neutral | 0.666 | Possibly Damaging | 0.355 | Benign | 4.57 | Benign | 0.24 | Tolerated | 4.32 | 2 | 0.1920 | 0.2635 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||
| c.3350G>T | G1117V 2D AIThe SynGAP1 missense variant G1117V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while ESM1b is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.853192 | Binding | 0.323 | 0.914 | 0.750 | -7.251 | In-Between | 0.083 | Likely Benign | Likely Benign | 0.284 | Likely Benign | -1.32 | Neutral | 0.011 | Benign | 0.014 | Benign | 4.57 | Benign | 0.04 | Affected | 0.1312 | 0.3680 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3352A>C | S1118R 2D AIThe SynGAP1 missense variant S1118R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that S1118R is most likely benign, and this conclusion is consistent with the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.828802 | Binding | 0.310 | 0.919 | 0.750 | -2.670 | Likely Benign | 0.553 | Ambiguous | Likely Benign | 0.175 | Likely Benign | -0.74 | Neutral | 0.034 | Benign | 0.023 | Benign | 5.17 | Benign | 0.05 | Affected | 4.32 | 2 | 0.1314 | 0.3431 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3352A>T | S1118C 2D AIThe SynGAP1 missense variant S1118C is listed in gnomAD (ID 6‑33443904‑A‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT, while ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.828802 | Binding | 0.310 | 0.919 | 0.750 | 6-33443904-A-T | -7.402 | In-Between | 0.096 | Likely Benign | Likely Benign | 0.311 | Likely Benign | -1.05 | Neutral | 0.997 | Probably Damaging | 0.889 | Possibly Damaging | 5.15 | Benign | 0.01 | Affected | 4.32 | 2 | 0.1648 | 0.5695 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||
| c.3353G>T | S1118I 2D AIThe SynGAP1 missense variant S1118I is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.828802 | Binding | 0.310 | 0.919 | 0.750 | -5.710 | Likely Benign | 0.154 | Likely Benign | Likely Benign | 0.309 | Likely Benign | -1.09 | Neutral | 0.990 | Probably Damaging | 0.798 | Possibly Damaging | 5.16 | Benign | 0.01 | Affected | 0.1486 | 0.4580 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3354C>A | S1118R 2D AIThe SynGAP1 missense variant S1118R (ClinVar ID 2656489.0) is listed as ClinVar status Uncertain and is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only SIFT predicts a pathogenic effect, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar Uncertain designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.828802 | Binding | 0.310 | 0.919 | 0.750 | Uncertain | 1 | -2.670 | Likely Benign | 0.553 | Ambiguous | Likely Benign | 0.166 | Likely Benign | -0.74 | Neutral | 0.034 | Benign | 0.023 | Benign | 5.17 | Benign | 0.05 | Affected | 4.32 | 2 | 0.1314 | 0.3431 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||
| c.3354C>G | S1118R 2D AIThe SynGAP1 missense variant S1118R is listed in gnomAD (ID 6‑33443906‑C‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.828802 | Binding | 0.310 | 0.919 | 0.750 | 6-33443906-C-G | -2.670 | Likely Benign | 0.553 | Ambiguous | Likely Benign | 0.165 | Likely Benign | -0.74 | Neutral | 0.034 | Benign | 0.023 | Benign | 5.17 | Benign | 0.05 | Affected | 4.32 | 2 | 0.1314 | 0.3431 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.3356G>T | G1119V 2D AIThe SynGAP1 missense variant G1119V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools, polyPhen‑2 HumDiv and polyPhen‑2 HumVar, predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.818538 | Binding | 0.339 | 0.928 | 0.875 | -6.428 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.352 | Likely Benign | -0.94 | Neutral | 0.918 | Possibly Damaging | 0.604 | Possibly Damaging | 3.91 | Benign | 0.31 | Tolerated | 0.1493 | 0.3707 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3359G>A | G1120D 2D AIThe SynGAP1 missense variant G1120D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33443911‑G‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and ESM1b. AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) resolves to a benign majority (2 benign vs. 1 pathogenic). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.871313 | Disordered | 0.804931 | Binding | 0.335 | 0.925 | 0.875 | 6-33443911-G-A | 4 | 2.65e-6 | -9.244 | Likely Pathogenic | 0.378 | Ambiguous | Likely Benign | 0.351 | Likely Benign | -0.82 | Neutral | 0.666 | Possibly Damaging | 0.355 | Benign | 3.60 | Benign | 0.19 | Tolerated | 3.77 | 5 | 0.1659 | 0.1835 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||
| c.335G>T | G112V 2D AIThe SynGAP1 missense variant G112V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.728858 | Disordered | 0.640153 | Binding | 0.332 | 0.867 | 0.750 | -2.411 | Likely Benign | 0.230 | Likely Benign | Likely Benign | 0.159 | Likely Benign | -3.39 | Deleterious | 0.421 | Benign | 0.108 | Benign | 3.96 | Benign | 0.00 | Affected | 0.1225 | 0.4218 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3361A>C | S1121R 2D AIThe SynGAP1 missense variant S1121R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign, reflecting the majority of benign predictions. High‑accuracy assessments show AlphaMissense‑Optimized as benign, and the SGM‑Consensus (majority vote) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact for S1121R, and this conclusion is consistent with the lack of ClinVar reporting. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.810024 | Binding | 0.365 | 0.935 | 0.875 | -6.945 | Likely Benign | 0.597 | Likely Pathogenic | Likely Benign | 0.142 | Likely Benign | -0.34 | Neutral | 0.016 | Benign | 0.015 | Benign | 5.45 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1346 | 0.3431 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3361A>T | S1121C 2D AIThe SynGAP1 missense variant S1121C is listed in gnomAD (ID 6‑33443913‑A‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while the Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.810024 | Binding | 0.365 | 0.935 | 0.875 | 6-33443913-A-T | -7.431 | In-Between | 0.094 | Likely Benign | Likely Benign | 0.354 | Likely Benign | -0.99 | Neutral | 0.994 | Probably Damaging | 0.840 | Possibly Damaging | 5.43 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1705 | 0.5691 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||||
| c.3362G>T | S1121I 2D AIThe SynGAP1 missense variant S1121I is listed in ClinVar (ID 4768628) with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the ClinVar “Uncertain” designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.810024 | Binding | 0.365 | 0.935 | 0.875 | Uncertain | 1 | -6.215 | Likely Benign | 0.147 | Likely Benign | Likely Benign | 0.455 | Likely Benign | -0.96 | Neutral | 0.875 | Possibly Damaging | 0.559 | Possibly Damaging | 5.44 | Benign | 0.00 | Affected | 0.1504 | 0.4776 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||
| c.3363C>A | S1121R 2D AIThe SynGAP1 missense variant S1121R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Benign, and Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.810024 | Binding | 0.365 | 0.935 | 0.875 | -6.945 | Likely Benign | 0.597 | Likely Pathogenic | Likely Benign | 0.100 | Likely Benign | -0.34 | Neutral | 0.016 | Benign | 0.015 | Benign | 5.45 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1346 | 0.3431 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3363C>G | S1121R 2D AIThe SynGAP1 missense variant S1121R is catalogued in gnomAD (ID 6‑33443915‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all report benign. Only SIFT and AlphaMissense‑Default predict pathogenicity, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.810024 | Binding | 0.365 | 0.935 | 0.875 | 6-33443915-C-G | -6.945 | Likely Benign | 0.597 | Likely Pathogenic | Likely Benign | 0.101 | Likely Benign | -0.34 | Neutral | 0.016 | Benign | 0.015 | Benign | 5.45 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1346 | 0.3431 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||||
| c.3365G>T | G1122V 2D AIThe SynGAP1 missense variant G1122V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly classify the variant as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) reports likely benign; Foldetta results are unavailable. Consequently, the variant is most likely benign, and this assessment is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.896620 | Disordered | 0.814918 | Binding | 0.357 | 0.932 | 0.875 | -6.398 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.271 | Likely Benign | -1.18 | Neutral | 0.059 | Benign | 0.025 | Benign | 4.49 | Benign | 0.10 | Tolerated | 0.1381 | 0.3694 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3368G>T | G1123V 2D AIThe SynGAP1 missense variant G1123V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; ESM1b is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective predictions strongly suggest that G1123V is most likely benign, and this conclusion is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.837511 | Disordered | 0.827246 | Binding | 0.346 | 0.934 | 0.875 | -7.129 | In-Between | 0.091 | Likely Benign | Likely Benign | 0.333 | Likely Benign | -1.03 | Neutral | 0.292 | Benign | 0.157 | Benign | 4.35 | Benign | 0.29 | Tolerated | 0.1362 | 0.3694 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3371G>T | G1124V 2D AIThe SynGAP1 missense variant G1124V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta results are unavailable, so they do not influence the assessment. Overall, the consensus of available predictions indicates that G1124V is most likely benign, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.837511 | Disordered | 0.833401 | Binding | 0.341 | 0.931 | 0.875 | -6.980 | Likely Benign | 0.094 | Likely Benign | Likely Benign | 0.315 | Likely Benign | -0.96 | Neutral | 0.586 | Possibly Damaging | 0.172 | Benign | 4.75 | Benign | 0.00 | Affected | 0.1299 | 0.3888 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3377G>T | G1126V 2D AIThe SynGAP1 missense variant G1126V is listed in ClinVar with an uncertain significance and is present in the gnomAD database. Consensus from multiple in‑silico predictors indicates a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score all classify the change as benign. Only the SIFT algorithm predicts a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized reports a benign effect, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely benign, while Foldetta’s protein‑folding stability analysis is unavailable. Overall, the preponderance of evidence points to a benign variant, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.894241 | Disordered | 0.837209 | Binding | 0.345 | 0.918 | 0.875 | Uncertain | 1 | 6-33443929-G-T | -6.536 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.357 | Likely Benign | -1.20 | Neutral | 0.009 | Benign | 0.008 | Benign | 4.76 | Benign | 0.03 | Affected | 3.77 | 5 | 0.1366 | 0.3884 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||
| c.3380G>T | G1127V 2D AIThe SynGAP1 missense variant G1127V is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33443932‑G‑T). All available in silico predictors classify the change as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool reports a pathogenic prediction. Grouping by agreement, the benign‑predicting tools comprise the entire set, while no pathogenic predictions exist. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates “Likely Benign.” Foldetta results are unavailable. Overall, the variant is most likely benign, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.852422 | Binding | 0.344 | 0.915 | 0.875 | Uncertain | 1 | 6-33443932-G-T | 1 | 6.69e-7 | -6.097 | Likely Benign | 0.094 | Likely Benign | Likely Benign | 0.230 | Likely Benign | -1.01 | Neutral | 0.004 | Benign | 0.005 | Benign | 4.81 | Benign | 0.17 | Tolerated | 4.32 | 4 | 0.1281 | 0.3669 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||
| c.338G>T | G113V 2D AIThe SynGAP1 missense variant G113V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.639486 | Binding | 0.350 | 0.870 | 0.750 | -3.752 | Likely Benign | 0.188 | Likely Benign | Likely Benign | 0.124 | Likely Benign | -1.77 | Neutral | 0.838 | Possibly Damaging | 0.145 | Benign | 4.18 | Benign | 0.05 | Affected | 0.1316 | 0.4002 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3391C>T | P1131S 2D AIThe SynGAP1 missense variant P1131S is reported in gnomAD (variant ID 6‑33443943‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign, and this is not contradictory to ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.855155 | Binding | 0.360 | 0.899 | 0.750 | 6-33443943-C-T | 1 | 6.72e-7 | -4.089 | Likely Benign | 0.246 | Likely Benign | Likely Benign | 0.209 | Likely Benign | -2.62 | Deleterious | 0.025 | Benign | 0.015 | Benign | 5.54 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2936 | 0.5936 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3398T>C | I1133T 2D AIThe SynGAP1 missense variant I1133T has no ClinVar entry and is present in gnomAD (ID 6‑33443950‑T‑C). Consensus among most tools is benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized all predict a benign effect, while AlphaMissense‑Default remains uncertain and no tool predicts pathogenicity. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized reports benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Benign,” and Foldetta’s protein‑folding stability analysis is unavailable. Overall, the evidence strongly supports a benign classification, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.832785 | Binding | 0.316 | 0.892 | 0.750 | 6-33443950-T-C | -4.522 | Likely Benign | 0.563 | Ambiguous | Likely Benign | 0.275 | Likely Benign | -0.30 | Neutral | 0.026 | Benign | 0.030 | Benign | 5.50 | Benign | 0.18 | Tolerated | 4.32 | 3 | 0.1007 | 0.1975 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||||
| c.3398T>G | I1133S 2D AIThe SynGAP1 missense variant I1133S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign. Foldetta results are unavailable, so they do not influence the assessment. Overall, the collective evidence strongly supports a benign impact for I1133S, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.832785 | Binding | 0.316 | 0.892 | 0.750 | -4.034 | Likely Benign | 0.492 | Ambiguous | Likely Benign | 0.186 | Likely Benign | -0.60 | Neutral | 0.007 | Benign | 0.016 | Benign | 5.70 | Benign | 0.08 | Tolerated | 0.2762 | 0.1110 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3413C>G | S1138C 2D AIThe SynGAP1 missense variant S1138C is catalogued in gnomAD (ID 6‑33444448‑C‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the preponderance of evidence points to a benign impact. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists for S1138C. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.738250 | Binding | 0.346 | 0.869 | 1.000 | 6-33444448-C-G | 1 | 6.20e-7 | -7.850 | In-Between | 0.117 | Likely Benign | Likely Benign | 0.425 | Likely Benign | -2.48 | Neutral | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 5.40 | Benign | 0.04 | Affected | 4.32 | 4 | 0.1472 | 0.6396 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||
| c.3419C>G | T1140R 2D AIThe SynGAP1 missense variant T1140R is listed in gnomAD (ID 6‑33444454‑C‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.926919 | Disordered | 0.708094 | Binding | 0.293 | 0.854 | 1.000 | 6-33444454-C-G | 1 | 6.20e-7 | -4.245 | Likely Benign | 0.682 | Likely Pathogenic | Likely Benign | 0.073 | Likely Benign | -1.69 | Neutral | 0.761 | Possibly Damaging | 0.398 | Benign | 2.61 | Benign | 0.15 | Tolerated | 4.32 | 4 | 0.0997 | 0.2442 | -1 | -1 | -3.8 | 55.08 | ||||||||||||||||||||||||||||||||||
| c.3424T>C | S1142P 2D AIThe SynGAP1 missense variant S1142P is listed in ClinVar (ID 2747352.0) as Benign and is present in gnomAD (variant ID 6‑33444459‑T‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, aligning with the ClinVar classification and not contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.719935 | Binding | 0.276 | 0.844 | 1.000 | Likely Benign | 1 | 6-33444459-T-C | 1 | 6.20e-7 | -2.713 | Likely Benign | 0.222 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -2.19 | Neutral | 0.918 | Possibly Damaging | 0.761 | Possibly Damaging | 2.64 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2080 | 0.5618 | -1 | 1 | -0.8 | 10.04 | ||||||||||||||||||||||||||||||||
| c.3428C>G | T1143R 2D AIThe SynGAP1 missense variant T1143R is not reported in ClinVar and is absent from gnomAD, indicating no known population frequency data. Functional prediction tools cluster into two groups: benign predictions include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from AlphaMissense‑Default, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.722918 | Binding | 0.275 | 0.837 | 1.000 | -2.882 | Likely Benign | 0.782 | Likely Pathogenic | Likely Benign | 0.175 | Likely Benign | -2.31 | Neutral | 0.996 | Probably Damaging | 0.951 | Probably Damaging | 2.73 | Benign | 0.09 | Tolerated | 0.1087 | 0.3060 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3436C>T | P1146S 2D AIThe SynGAP1 missense variant P1146S is reported in ClinVar as “Not submitted” and is present in gnomAD (variant ID 6-33444471-C‑T). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of a ClinVar pathogenic classification. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.919029 | Disordered | 0.732173 | Binding | 0.415 | 0.837 | 1.000 | 6-33444471-C-T | 2 | 1.24e-6 | -3.045 | Likely Benign | 0.318 | Likely Benign | Likely Benign | 0.470 | Likely Benign | -3.66 | Deleterious | 0.945 | Possibly Damaging | 0.760 | Possibly Damaging | 5.52 | Benign | 0.00 | Affected | 4.32 | 4 | 0.3408 | 0.5186 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3440C>G | T1147R 2D AIThe SynGAP1 missense variant T1147R is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools cluster around a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized all indicate benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized reports a benign effect, and the SGM‑Consensus likewise indicates Likely Benign; Foldetta results are unavailable. Taken together, the preponderance of evidence points to a benign impact for T1147R, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.831250 | Disordered | 0.746520 | Binding | 0.345 | 0.839 | 0.875 | -3.234 | Likely Benign | 0.534 | Ambiguous | Likely Benign | 0.442 | Likely Benign | -2.44 | Neutral | 0.411 | Benign | 0.139 | Benign | 5.46 | Benign | 0.01 | Affected | 0.1014 | 0.2742 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3443T>C | M1148T 2D AIThe SynGAP1 missense variant M1148T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.774279 | Binding | 0.343 | 0.835 | 0.875 | -0.972 | Likely Benign | 0.159 | Likely Benign | Likely Benign | 0.151 | Likely Benign | -1.50 | Neutral | 0.001 | Benign | 0.002 | Benign | 2.56 | Benign | 0.00 | Affected | 0.2063 | 0.2214 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.3452C>G | S1151C 2D AIThe SynGAP1 missense variant S1151C is reported in gnomAD (ID 6‑33444487‑C‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Two tools (polyPhen‑2 HumDiv and HumVar) predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.741537 | Disordered | 0.805072 | Binding | 0.394 | 0.839 | 0.625 | 6-33444487-C-G | 1 | 6.20e-7 | -6.778 | Likely Benign | 0.225 | Likely Benign | Likely Benign | 0.181 | Likely Benign | -0.86 | Neutral | 0.999 | Probably Damaging | 0.944 | Probably Damaging | 2.67 | Benign | 0.07 | Tolerated | 3.77 | 5 | 0.1056 | 0.5260 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||||||
| c.3461C>G | T1154R 2D AIThe SynGAP1 missense variant T1154R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic. No Foldetta stability prediction is available for this variant. Overall, the majority of computational evidence indicates that T1154R is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.685117 | Disordered | 0.838654 | Binding | 0.382 | 0.851 | 0.625 | -3.464 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.300 | Likely Benign | -4.05 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 1.73 | Pathogenic | 0.00 | Affected | 0.0903 | 0.2101 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3464T>G | V1155G 2D AIThe SynGAP1 missense variant V1155G is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods give mixed results: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (benign), FATHMM (benign), and PROVEAN (benign), reports a benign outcome; Foldetta’s stability assessment is unavailable. Overall, the majority of conventional tools lean toward pathogenicity, whereas the high‑accuracy consensus leans benign, leaving the functional impact uncertain. The variant is most likely pathogenic based on the prevailing predictions, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.750527 | Disordered | 0.855718 | Binding | 0.335 | 0.857 | 0.500 | -3.018 | Likely Benign | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.209 | Likely Benign | -1.91 | Neutral | 0.997 | Probably Damaging | 0.999 | Probably Damaging | 2.59 | Benign | 0.05 | Affected | 0.2211 | 0.2374 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3472G>T | V1158F 2D AIThe SynGAP1 missense variant V1158F is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and ESM1b, while the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is classified as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports the variant as Likely Pathogenic. Foldetta results are unavailable. Overall, the majority of evidence indicates that V1158F is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar annotation exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.599170 | Disordered | 0.877504 | Binding | 0.369 | 0.847 | 0.250 | -3.888 | Likely Benign | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.264 | Likely Benign | -2.66 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.33 | Pathogenic | 0.02 | Affected | 0.0792 | 0.3577 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3473T>G | V1158G 2D AIThe SynGAP1 missense variant V1158G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic outcome. High‑accuracy assessments reinforce this pattern: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields a pathogenic verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that V1158G is most likely pathogenic, and this conclusion does not contradict the current ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.599170 | Disordered | 0.877504 | Binding | 0.369 | 0.847 | 0.250 | -3.058 | Likely Benign | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.433 | Likely Benign | -4.62 | Deleterious | 0.997 | Probably Damaging | 0.999 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.1870 | 0.3289 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3482T>C | M1161T 2D AIThe SynGAP1 missense variant M1161T is listed in gnomAD (ID 6‑33444517‑T‑C) but has no ClinVar entry. Functional prediction tools fall into two groups: benign predictions come from REVEL, SIFT, and ESM1b, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Pathogenic. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.580690 | Disordered | 0.864109 | Binding | 0.372 | 0.830 | 0.375 | 6-33444517-T-C | 1 | 6.20e-7 | -2.620 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.148 | Likely Benign | -2.75 | Deleterious | 0.968 | Probably Damaging | 0.954 | Probably Damaging | 2.19 | Pathogenic | 0.27 | Tolerated | 3.77 | 5 | 0.2218 | 0.1786 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||
| c.349A>C | S117R 2D AIThe SynGAP1 missense variant S117R has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, whereas the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome; Foldetta results are unavailable. Overall, the majority of predictions lean toward a benign impact, and this does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.672422 | Binding | 0.357 | 0.877 | 0.625 | -4.187 | Likely Benign | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.276 | Likely Benign | -1.81 | Neutral | 0.845 | Possibly Damaging | 0.326 | Benign | 3.70 | Benign | 0.01 | Affected | 3.61 | 5 | 0.1040 | 0.3770 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.3503T>G | I1168S 2D AISynGAP1 missense variant I1168S has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, whereas the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome; Foldetta results are unavailable. Overall, the evidence is split, with an equal number of benign and pathogenic calls, and the high‑accuracy predictions are contradictory. Therefore, the variant is not conclusively predicted to be benign or pathogenic, and there is no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.763262 | Binding | 0.423 | 0.796 | 0.500 | -2.212 | Likely Benign | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.442 | Likely Benign | -1.46 | Neutral | 0.998 | Probably Damaging | 0.958 | Probably Damaging | 5.67 | Benign | 0.01 | Affected | 0.3247 | 0.1540 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3509G>T | S1170I 2D AIThe SynGAP1 missense variant S1170I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote) also predicts benign, and the Foldetta protein‑folding stability analysis is unavailable. Taken together, the preponderance of high‑confidence predictions indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.622677 | Disordered | 0.719138 | Binding | 0.417 | 0.767 | 0.500 | -3.813 | Likely Benign | 0.635 | Likely Pathogenic | Likely Benign | 0.600 | Likely Pathogenic | -2.16 | Neutral | 0.998 | Probably Damaging | 0.990 | Probably Damaging | 5.29 | Benign | 0.01 | Affected | 0.0754 | 0.5329 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||
| c.350G>T | S117I 2D AIThe SynGAP1 missense variant S117I is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus (SGM‑Consensus) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.672422 | Binding | 0.357 | 0.877 | 0.625 | -5.483 | Likely Benign | 0.559 | Ambiguous | Likely Benign | 0.137 | Likely Benign | -2.33 | Neutral | 0.971 | Probably Damaging | 0.598 | Possibly Damaging | 3.70 | Benign | 0.00 | Affected | 0.1060 | 0.5048 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3518T>G | I1173S 2D AIThe SynGAP1 missense variant I1173S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, while those that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for I1173S, and this conclusion does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.501700 | Disordered | 0.653145 | Binding | 0.521 | 0.756 | 0.375 | -2.416 | Likely Benign | 0.557 | Ambiguous | Likely Benign | 0.455 | Likely Benign | -1.18 | Neutral | 0.625 | Possibly Damaging | 0.265 | Benign | 5.45 | Benign | 0.02 | Affected | 0.2796 | 0.0512 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.351C>A | S117R 2D AIThe SynGAP1 missense variant S117R has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification (3 benign vs. 1 pathogenic votes). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, whereas the SGM‑Consensus (majority vote) remains benign; Foldetta results are unavailable. Overall, the majority of tools (six benign vs. four pathogenic) and the consensus evidence lean toward a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.672422 | Binding | 0.357 | 0.877 | 0.625 | -4.187 | Likely Benign | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.144 | Likely Benign | -1.81 | Neutral | 0.845 | Possibly Damaging | 0.326 | Benign | 3.70 | Benign | 0.01 | Affected | 3.61 | 5 | 0.1040 | 0.3770 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.351C>G | S117R 2D AIThe SynGAP1 missense variant S117R is listed in ClinVar with no submitted interpretation and is present in gnomAD (ID 6‑33432216‑C‑G). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, while the SGM‑Consensus remains likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign effect, and this is consistent with the absence of a pathogenic ClinVar annotation. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.661982 | Disordered | 0.672422 | Binding | 0.357 | 0.877 | 0.625 | 6-33432216-C-G | 1 | 6.20e-7 | -4.187 | Likely Benign | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.144 | Likely Benign | -1.81 | Neutral | 0.845 | Possibly Damaging | 0.326 | Benign | 3.70 | Benign | 0.01 | Affected | 3.61 | 5 | 0.1040 | 0.3770 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||
| c.353T>C | M118T 2D AIThe SynGAP1 missense variant M118T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome; AlphaMissense‑Default is uncertain. The high‑accuracy consensus (SGM‑Consensus) also indicates a likely benign classification, and AlphaMissense‑Optimized independently predicts benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the evidence from multiple independent predictors and the high‑accuracy consensus strongly supports a benign impact. This conclusion is consistent with the lack of any ClinVar pathogenic annotation, so there is no contradiction with existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.694846 | Disordered | 0.676867 | Binding | 0.330 | 0.883 | 0.500 | -2.468 | Likely Benign | 0.454 | Ambiguous | Likely Benign | 0.217 | Likely Benign | -2.41 | Neutral | 0.396 | Benign | 0.067 | Benign | 3.86 | Benign | 0.08 | Tolerated | 0.2207 | 0.2237 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.3560T>C | M1187T 2D AIThe SynGAP1 missense variant M1187T is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” outcome. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as benign, and Foldetta results are unavailable. Overall, the majority of individual predictors (seven) indicate pathogenicity, whereas only three suggest benignity. Consequently, the variant is most likely pathogenic based on the available computational evidence, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.562014 | Disordered | 0.499801 | Uncertain | 0.597 | 0.636 | 0.625 | -3.502 | Likely Benign | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.597 | Likely Pathogenic | -1.75 | Neutral | 0.968 | Probably Damaging | 0.954 | Probably Damaging | 5.63 | Benign | 0.04 | Affected | 0.2378 | 0.1757 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||||||
| c.3569G>T | S1190I 2D AIThe SynGAP1 missense change S1190I is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM. Those that predict a pathogenic impact are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized model classifies the variant as pathogenic, whereas the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Benign. Foldetta stability analysis is unavailable. Overall, the balance of evidence favors a pathogenic interpretation, and this assessment does not contradict any ClinVar annotation because no ClinVar record exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.575842 | Disordered | 0.455760 | Uncertain | 0.742 | 0.624 | 0.625 | -4.842 | Likely Benign | 0.958 | Likely Pathogenic | Likely Pathogenic | 0.371 | Likely Benign | -1.89 | Neutral | 0.997 | Probably Damaging | 0.996 | Probably Damaging | 5.26 | Benign | 0.04 | Affected | 0.0881 | 0.4175 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3578A>G | D1193G 2D AIThe SynGAP1 D1193G missense variant is catalogued in gnomAD (ID 6‑33444613‑A‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority of the four high‑accuracy tools) also favors benign. Foldetta results are unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.585406 | Disordered | 0.433390 | Uncertain | 0.807 | 0.600 | 0.375 | 6-33444613-A-G | 1 | 6.21e-7 | -6.506 | Likely Benign | 0.765 | Likely Pathogenic | Likely Benign | 0.480 | Likely Benign | -2.27 | Neutral | 0.924 | Possibly Damaging | 0.652 | Possibly Damaging | 5.42 | Benign | 0.00 | Affected | 4.32 | 4 | 0.2695 | 0.4765 | -1 | 1 | 3.1 | -58.04 | |||||||||||||||||||||||||||||||||
| c.3581G>C | R1194T 2D AIThe SynGAP1 missense variant R1194T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default predict a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels it as likely benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the predictions are split, with four tools supporting benign and four supporting pathogenic, and the high‑accuracy consensus is conflicting. Based on the available evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.685117 | Disordered | 0.425297 | Uncertain | 0.796 | 0.602 | 0.375 | -4.875 | Likely Benign | 0.973 | Likely Pathogenic | Likely Pathogenic | 0.402 | Likely Benign | -1.96 | Neutral | 0.991 | Probably Damaging | 0.991 | Probably Damaging | 5.50 | Benign | 0.02 | Affected | 0.1866 | 0.3706 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||
| c.3584T>G | V1195G 2D AIThe SynGAP1 missense variant V1195G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a pathogenic effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. Tools that agree on a benign effect are ESM1b and FATHMM. AlphaMissense‑Optimized is uncertain, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive because it yields a 2‑to‑2 split. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, six of the eight evaluated tools predict pathogenicity while only two predict benign, and no high‑accuracy consensus or folding‑stability evidence contradicts this. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not conflict with the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.604312 | Disordered | 0.434133 | Uncertain | 0.842 | 0.603 | 0.250 | -5.463 | Likely Benign | 0.881 | Likely Pathogenic | Ambiguous | 0.586 | Likely Pathogenic | -2.81 | Deleterious | 0.998 | Probably Damaging | 1.000 | Probably Damaging | 5.55 | Benign | 0.01 | Affected | 0.2005 | 0.2100 | -1 | -3 | -4.6 | -42.08 | |||||||||||||||||||||||||||||||||||||||
| c.359G>T | G120V 2D AIThe SynGAP1 missense variant G120V is not reported in ClinVar and is absent from gnomAD. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign.” Foldetta results are unavailable. Overall, the consensus of all available predictions points to a benign impact, and this conclusion is not contradicted by any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.659993 | Binding | 0.359 | 0.887 | 0.750 | -4.571 | Likely Benign | 0.107 | Likely Benign | Likely Benign | 0.036 | Likely Benign | -0.68 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.28 | Benign | 0.19 | Tolerated | 0.1321 | 0.3883 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.35G>T | S12I 2D AIThe SynGAP1 missense variant S12I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools—polyPhen‑2 HumDiv and SIFT—suggest a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.480142 | Structured | 0.490599 | Uncertain | 0.355 | 0.916 | 0.500 | -4.838 | Likely Benign | 0.299 | Likely Benign | Likely Benign | 0.121 | Likely Benign | 0.05 | Neutral | 0.659 | Possibly Damaging | 0.072 | Benign | 4.09 | Benign | 0.00 | Affected | 0.0964 | 0.6082 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3605T>G | I1202S 2D AIThe SynGAP1 missense variant I1202S is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are not available. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.529623 | Disordered | 0.510422 | Binding | 0.874 | 0.593 | 0.250 | -11.877 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.431 | Likely Benign | -4.68 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.3021 | 0.0640 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3632T>C | M1211T 2D AIThe SynGAP1 missense variant M1211T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those predicting pathogenicity are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a benign impact, and this conclusion does not contradict any ClinVar status, as none is assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.585406 | Disordered | 0.578388 | Binding | 0.876 | 0.565 | 0.500 | -3.885 | Likely Benign | 0.669 | Likely Pathogenic | Likely Benign | 0.448 | Likely Benign | -1.99 | Neutral | 0.987 | Probably Damaging | 0.968 | Probably Damaging | 5.54 | Benign | 0.02 | Affected | 0.1987 | 0.1614 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||||||
| c.364C>T | P122S 2D AIThe SynGAP1 missense variant P122S is catalogued in gnomAD (ID 6‑33432229‑C‑T) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status remains unavailable. Overall, the computational evidence overwhelmingly supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.618285 | Disordered | 0.672358 | Binding | 0.400 | 0.887 | 0.750 | 6-33432229-C-T | 1 | 6.20e-7 | -3.389 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -1.13 | Neutral | 0.036 | Benign | 0.025 | Benign | 4.22 | Benign | 0.31 | Tolerated | 3.61 | 5 | 0.4009 | 0.4759 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3665G>C | R1222T 2D AIThe SynGAP1 missense variant R1222T is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and SIFT, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—consistently predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Pathogenic. Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the preponderance of evidence indicates that R1222T is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.703578 | Disordered | 0.423869 | Uncertain | 0.895 | 0.541 | 0.250 | -11.164 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.407 | Likely Benign | -4.39 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.48 | Pathogenic | 0.13 | Tolerated | 0.1550 | 0.2851 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||
| c.3692G>T | S1231I 2D AIThe SynGAP1 missense variant S1231I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, and AlphaMissense‑Optimized, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains likely pathogenic; Foldetta results are unavailable. Overall, the balance of evidence favors a pathogenic classification for S1231I, and this conclusion does not contradict any existing ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.490133 | Structured | 0.519419 | Binding | 0.876 | 0.544 | 0.250 | -9.360 | Likely Pathogenic | 0.712 | Likely Pathogenic | Likely Benign | 0.203 | Likely Benign | -3.24 | Deleterious | 0.966 | Probably Damaging | 0.690 | Possibly Damaging | 2.64 | Benign | 0.04 | Affected | 0.0701 | 0.4550 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3698T>G | I1233S 2D AIThe SynGAP1 missense variant I1233S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and FATHMM, whereas the majority of other predictors—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized—classify the change as pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic verdict. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus likewise indicates likely pathogenic. Foldetta, a protein‑folding stability method that combines FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a pathogenic effect, and this conclusion does not conflict with the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.525368 | Disordered | 0.564054 | Binding | 0.881 | 0.531 | 0.125 | -8.066 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.184 | Likely Benign | -3.60 | Deleterious | 0.946 | Possibly Damaging | 0.673 | Possibly Damaging | 2.53 | Benign | 0.00 | Affected | 0.2848 | 0.0910 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3704T>C | M1235T 2D AIThe SynGAP1 missense variant M1235T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of tools, including the high‑accuracy methods, points to a benign impact. This prediction does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.690604 | Disordered | 0.577958 | Binding | 0.872 | 0.532 | 0.125 | -5.381 | Likely Benign | 0.149 | Likely Benign | Likely Benign | 0.060 | Likely Benign | -2.39 | Neutral | 0.425 | Benign | 0.158 | Benign | 2.66 | Benign | 0.01 | Affected | 0.2079 | 0.1814 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||||||
| c.3731G>T | S1244I 2D AIThe SynGAP1 missense variant S1244I is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus as likely pathogenic; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.648219 | Disordered | 0.411055 | Uncertain | 0.833 | 0.549 | 0.500 | -13.073 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 0.284 | Likely Benign | -4.39 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.08 | Pathogenic | 0.02 | Affected | 0.0780 | 0.4684 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3740G>C | R1247T 2D AIThe SynGAP1 missense variant R1247T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score, which is labeled Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic. No Foldetta stability result is available. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not conflict with ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.736850 | Disordered | 0.374141 | Uncertain | 0.875 | 0.557 | 0.625 | -6.302 | Likely Benign | 0.598 | Likely Pathogenic | Likely Benign | 0.206 | Likely Benign | -4.79 | Deleterious | 0.980 | Probably Damaging | 0.783 | Possibly Damaging | 1.71 | Pathogenic | 0.00 | Affected | 0.1543 | 0.2403 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||
| c.3746G>C | R1249T 2D AIThe SynGAP1 missense variant R1249T is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL and AlphaMissense‑Optimized, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score. The SGM‑Consensus, a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic effect. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM‑Consensus remains pathogenic; Foldetta results are unavailable. Overall, the preponderance of evidence (10 pathogenic vs. 2 benign predictions) points to a pathogenic impact for R1249T. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.745909 | Disordered | 0.366265 | Uncertain | 0.874 | 0.556 | 0.875 | -8.014 | Likely Pathogenic | 0.712 | Likely Pathogenic | Likely Benign | 0.185 | Likely Benign | -4.59 | Deleterious | 0.990 | Probably Damaging | 0.903 | Possibly Damaging | 1.71 | Pathogenic | 0.00 | Affected | 0.1646 | 0.2458 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||
| c.376T>G | F126V 2D AIThe SynGAP1 missense variant F126V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as Likely Benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact for F126V, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.712056 | Binding | 0.316 | 0.874 | 0.500 | -1.584 | Likely Benign | 0.635 | Likely Pathogenic | Likely Benign | 0.086 | Likely Benign | -2.47 | Neutral | 0.160 | Benign | 0.045 | Benign | 3.98 | Benign | 0.00 | Affected | 0.2496 | 0.2646 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.3776T>G | I1259S 2D AIThe SynGAP1 missense variant I1259S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled “Likely Pathogenic.” No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.494003 | Structured | 0.576405 | Binding | 0.885 | 0.574 | 0.250 | -12.269 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.426 | Likely Benign | -2.76 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 2.53 | Benign | 0.01 | Affected | 0.3004 | 0.1110 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3782G>T | S1261I 2D AIThe SynGAP1 missense variant S1261I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and AlphaMissense‑Optimized, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score indicates a benign effect, whereas the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as likely pathogenic. No Foldetta stability assessment is available for this residue. Overall, the preponderance of evidence from standard and high‑accuracy predictors points to a pathogenic impact for S1261I. This conclusion is not contradicted by ClinVar status, which currently contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.501700 | Disordered | 0.671500 | Binding | 0.889 | 0.574 | 0.250 | -8.835 | Likely Pathogenic | 0.761 | Likely Pathogenic | Likely Benign | 0.244 | Likely Benign | -4.20 | Deleterious | 0.996 | Probably Damaging | 0.898 | Possibly Damaging | 2.21 | Pathogenic | 0.02 | Affected | 0.0631 | 0.4410 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3785T>G | I1262S 2D AIThe SynGAP1 missense variant I1262S is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM‑Consensus also reports it as likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion is not contradicted by ClinVar status, which currently has no entry for I1262S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.497853 | Structured | 0.707863 | Binding | 0.886 | 0.576 | 0.125 | -15.167 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.483 | Likely Benign | -4.98 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.2943 | 0.1110 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3788T>G | I1263S 2D AIThe SynGAP1 missense variant I1263S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only REVEL, whereas the remaining tools—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic, and the SGM‑Consensus is labeled likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.425610 | Structured | 0.740957 | Binding | 0.867 | 0.574 | 0.000 | -8.074 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.493 | Likely Benign | -4.97 | Deleterious | 0.984 | Probably Damaging | 0.825 | Possibly Damaging | 1.80 | Pathogenic | 0.00 | Affected | 0.3242 | 0.0910 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||
| c.3791G>T | G1264V 2D AIThe SynGAP1 missense variant G1264V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | Coiled-coil | 0.429200 | Structured | 0.762385 | Binding | 0.897 | 0.579 | 0.000 | -6.116 | Likely Benign | 0.488 | Ambiguous | Likely Benign | 0.136 | Likely Benign | -2.33 | Neutral | 0.966 | Probably Damaging | 0.617 | Possibly Damaging | 2.74 | Benign | 0.15 | Tolerated | 0.0972 | 0.3891 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||
| c.3794G>C | R1265T 2D AIThe SynGAP1 missense variant R1265T is reported in ClinVar as Pathogenic (ClinVar ID 522047.0) and is not found in gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic. No tool in the dataset predicts a benign outcome. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a Likely Pathogenic verdict. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of all available predictions supports a pathogenic classification, which is consistent with the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.414856 | Structured | 0.782497 | Binding | 0.887 | 0.592 | 0.000 | Likely Pathogenic | 1 | -10.129 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.529 | Likely Pathogenic | -4.97 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 2.29 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1947 | 0.4618 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||
| c.3800T>C | M1267T 2D AIThe SynGAP1 missense variant M1267T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and ESM1b, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) likewise indicates likely pathogenicity. No Foldetta stability prediction is available for this variant. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | Coiled-coil | 0.429200 | Structured | 0.812047 | Binding | 0.847 | 0.614 | 0.000 | -5.315 | Likely Benign | 0.958 | Likely Pathogenic | Likely Pathogenic | 0.270 | Likely Benign | -5.00 | Deleterious | 0.982 | Probably Damaging | 0.672 | Possibly Damaging | 2.32 | Pathogenic | 0.00 | Affected | 0.2036 | 0.1934 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||||||
| c.3806T>G | V1269G 2D AIThe SynGAP1 missense variant V1269G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include only REVEL, whereas all other evaluated algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic outcome. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is pathogenic, and the SGM Consensus is pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Coiled-coil | 0.433034 | Structured | 0.787464 | Binding | 0.843 | 0.647 | 0.125 | -9.927 | Likely Pathogenic | 0.969 | Likely Pathogenic | Likely Pathogenic | 0.420 | Likely Benign | -5.91 | Deleterious | 0.995 | Probably Damaging | 0.999 | Probably Damaging | 2.10 | Pathogenic | 0.00 | Affected | 0.2180 | 0.2557 | -1 | -3 | -4.6 | -42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3820C>A | R1274S 2D AIThe SynGAP1 missense variant R1274S is not reported in ClinVar (status: “None”) but is present in gnomAD (ID 6‑33447868‑C‑A). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are limited: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward a pathogenic impact. Thus, the variant is most likely pathogenic based on current computational evidence, and this assessment does not contradict any ClinVar status because no ClinVar claim exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.613573 | Disordered | 0.779985 | Binding | 0.746 | 0.688 | 0.625 | 6-33447868-C-A | -3.149 | Likely Benign | 0.830 | Likely Pathogenic | Ambiguous | 0.139 | Likely Benign | -3.19 | Deleterious | 0.997 | Probably Damaging | 0.993 | Probably Damaging | 2.52 | Benign | 0.01 | Affected | 3.77 | 5 | 0.3056 | 0.1830 | -1 | 0 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||
| c.3829C>G | H1277D 2D AIThe SynGAP1 missense variant H1277D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33447877‑C‑G). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), and ESM1b, while those that predict a pathogenic effect are PROVEAN, SIFT, and FATHMM. AlphaMissense‑Default is uncertain, whereas AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a pathogenic outcome (two pathogenic, one benign, one uncertain). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑accuracy and consensus predictions indicate a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.775545 | Disordered | 0.805725 | Binding | 0.562 | 0.718 | 0.750 | 6-33447877-C-G | -4.632 | Likely Benign | 0.537 | Ambiguous | Likely Benign | 0.172 | Likely Benign | -6.38 | Deleterious | 0.411 | Benign | 0.091 | Benign | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2389 | 0.1266 | -1 | 1 | -0.3 | -22.05 | |||||||||||||||||||||||||||||||||||||
| c.382C>A | P128T 2D AIThe SynGAP1 missense variant P128T is listed in ClinVar with an “Uncertain” status (ClinVar ID 2801315.0) and is present in gnomAD (ID 6‑33432247‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and polyPhen‑2 HumVar. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote) as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.713069 | Binding | 0.376 | 0.878 | 0.625 | Uncertain | 1 | 6-33432247-C-A | 1 | 6.20e-7 | -4.217 | Likely Benign | 0.267 | Likely Benign | Likely Benign | 0.075 | Likely Benign | -0.96 | Neutral | 0.952 | Possibly Damaging | 0.500 | Possibly Damaging | 4.19 | Benign | 0.35 | Tolerated | 3.74 | 4 | 0.2023 | 0.4053 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||
| c.382C>T | P128S 2D AIThe SynGAP1 missense variant P128S is reported in gnomAD (variant ID 6‑33432247‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant prediction. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.713069 | Binding | 0.376 | 0.878 | 0.625 | 6-33432247-C-T | 1 | 6.20e-7 | -3.234 | Likely Benign | 0.268 | Likely Benign | Likely Benign | 0.084 | Likely Benign | -1.35 | Neutral | 0.734 | Possibly Damaging | 0.243 | Benign | 4.20 | Benign | 0.24 | Tolerated | 3.74 | 4 | 0.3893 | 0.3697 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3832C>A | P1278T 2D AIThe SynGAP1 missense variant P1278T is catalogued in gnomAD (ID 6‑33447880‑C‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status remains unavailable. Overall, the computational evidence overwhelmingly supports a benign effect, and this assessment does not contradict any ClinVar classification. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.806955 | Binding | 0.532 | 0.722 | 0.750 | 6-33447880-C-A | -5.505 | Likely Benign | 0.095 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -0.60 | Neutral | 0.059 | Benign | 0.026 | Benign | 2.73 | Benign | 0.11 | Tolerated | 3.77 | 5 | 0.1825 | 0.4053 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3839T>C | M1280T 2D AIThe SynGAP1 missense variant M1280T is catalogued in gnomAD (6‑33447887‑T‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate a benign or likely benign outcome. In contrast, SIFT and FATHMM predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are not available. Overall, the majority of evidence supports a benign classification, and this is consistent with the absence of a ClinVar pathogenic designation. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.822030 | Binding | 0.510 | 0.726 | 0.875 | 6-33447887-T-C | 3 | 1.93e-6 | -2.305 | Likely Benign | 0.112 | Likely Benign | Likely Benign | 0.158 | Likely Benign | -2.26 | Neutral | 0.369 | Benign | 0.120 | Benign | 2.34 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1814 | 0.2031 | -1 | -1 | -2.6 | -30.09 | ||||||||||||||||||||||||||||||||||
| c.3847C>A | P1283T 2D AIThe SynGAP1 missense variant P1283T is catalogued in gnomAD (ID 6‑33447895‑C‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Therefore, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.819686 | Binding | 0.484 | 0.732 | 0.875 | 6-33447895-C-A | -4.781 | Likely Benign | 0.064 | Likely Benign | Likely Benign | 0.071 | Likely Benign | -0.45 | Neutral | 0.451 | Benign | 0.193 | Benign | 2.76 | Benign | 0.11 | Tolerated | 3.77 | 5 | 0.1269 | 0.3784 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3847C>T | P1283S 2D AIThe SynGAP1 missense variant P1283S is catalogued in gnomAD (ID 6‑33447895‑C‑T) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the computational evidence overwhelmingly supports a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Therefore, the variant is most likely benign, and this assessment does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.819686 | Binding | 0.484 | 0.732 | 0.875 | 6-33447895-C-T | -4.289 | Likely Benign | 0.072 | Likely Benign | Likely Benign | 0.040 | Likely Benign | 0.06 | Neutral | 0.292 | Benign | 0.110 | Benign | 2.86 | Benign | 0.30 | Tolerated | 3.77 | 5 | 0.2837 | 0.3719 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||
| c.3848C>A | P1283Q 2D AIThe SynGAP1 missense variant P1283Q is reported in gnomAD (variant ID 6‑33447896‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta’s protein‑folding stability analysis is unavailable. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.762850 | Disordered | 0.819686 | Binding | 0.484 | 0.732 | 0.875 | 6-33447896-C-A | -4.028 | Likely Benign | 0.085 | Likely Benign | Likely Benign | 0.102 | Likely Benign | -1.11 | Neutral | 0.991 | Probably Damaging | 0.567 | Possibly Damaging | 2.72 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1195 | 0.2926 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3853C>A | P1285T 2D AIThe SynGAP1 missense variant P1285T is catalogued in gnomAD (ID 6‑33447901‑C‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as tolerated or benign. No tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a Likely Benign verdict. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its status is unavailable. Overall, the computational evidence overwhelmingly supports a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.821643 | Binding | 0.557 | 0.759 | 0.750 | 6-33447901-C-A | -4.230 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.047 | Likely Benign | -0.35 | Neutral | 0.451 | Benign | 0.193 | Benign | 4.33 | Benign | 0.40 | Tolerated | 4.32 | 2 | 0.1496 | 0.3556 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3853C>T | P1285S 2D AIThe SynGAP1 missense variant P1285S is reported in gnomAD (variant ID 6‑33447901‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant call. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of tools and the high‑accuracy predictions point to a benign impact, and this is not in conflict with ClinVar, which currently has no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.821643 | Binding | 0.557 | 0.759 | 0.750 | 6-33447901-C-T | 1 | 6.44e-7 | -3.875 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.035 | Likely Benign | -1.27 | Neutral | 0.802 | Possibly Damaging | 0.249 | Benign | 4.34 | Benign | 0.29 | Tolerated | 4.32 | 2 | 0.2912 | 0.3319 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3854C>A | P1285Q 2D AIThe SynGAP1 missense variant P1285Q is reported in gnomAD (variant ID 6‑33447902‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, polyPhen‑2 (HumDiv and HumVar) predict it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also likely benign. No Foldetta stability analysis is available, so it does not influence the overall assessment. Based on the preponderance of evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.834292 | Disordered | 0.821643 | Binding | 0.557 | 0.759 | 0.750 | 6-33447902-C-A | -4.073 | Likely Benign | 0.100 | Likely Benign | Likely Benign | 0.072 | Likely Benign | -1.67 | Neutral | 0.977 | Probably Damaging | 0.632 | Possibly Damaging | 4.22 | Benign | 0.13 | Tolerated | 4.32 | 2 | 0.1476 | 0.3025 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3859C>A | P1287T 2D AIThe SynGAP1 missense variant P1287T is listed in ClinVar with an “Uncertain” status and is present in gnomAD (ID 6‑33447907‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.813701 | Binding | 0.538 | 0.777 | 0.750 | Uncertain | 1 | 6-33447907-C-A | -3.940 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.044 | Likely Benign | -0.22 | Neutral | 0.126 | Benign | 0.041 | Benign | 2.78 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1189 | 0.3995 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||
| c.3859C>T | P1287S 2D AIThe SynGAP1 missense variant P1287S is catalogued in gnomAD (ID 6‑33447907‑C‑T) but has no ClinVar entry. Across the spectrum of in‑silico predictors, every tool listed—REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently scores the variant as benign. No pathogenic predictions are reported. Grouping by agreement, the benign‑predicting tools comprise the entire set, while the pathogenic group is empty. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is assigned). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.813701 | Binding | 0.538 | 0.777 | 0.750 | 6-33447907-C-T | -3.258 | Likely Benign | 0.090 | Likely Benign | Likely Benign | 0.055 | Likely Benign | 0.70 | Neutral | 0.004 | Benign | 0.004 | Benign | 2.96 | Benign | 0.96 | Tolerated | 3.77 | 5 | 0.2820 | 0.3566 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||
| c.3869G>C | R1290T 2D AIThe SynGAP1 missense variant R1290T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a Likely Benign classification. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Based on the majority of predictions and the high‑accuracy consensus, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.784345 | Disordered | 0.844138 | Binding | 0.567 | 0.795 | 0.625 | -4.044 | Likely Benign | 0.311 | Likely Benign | Likely Benign | 0.111 | Likely Benign | -3.50 | Deleterious | 0.625 | Possibly Damaging | 0.266 | Benign | 2.64 | Benign | 0.01 | Affected | 0.1607 | 0.3359 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3880G>C | A1294P 2D AIThe SynGAP1 missense variant A1294P is catalogued in gnomAD (6‑33447928‑G‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—yields a 2‑to‑2 split and is therefore inconclusive; Foldetta results are not available. Overall, the balance of evidence leans toward a pathogenic effect, though the single high‑accuracy benign call and the inconclusive consensus temper certainty. This assessment does not conflict with ClinVar, which currently contains no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.784345 | Disordered | 0.895011 | Binding | 0.565 | 0.806 | 0.625 | 6-33447928-G-C | 1 | 6.45e-7 | -2.688 | Likely Benign | 0.220 | Likely Benign | Likely Benign | 0.367 | Likely Benign | -3.79 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 2.14 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.1682 | 0.3207 | -1 | 1 | -3.4 | 26.04 | |||||||||||||||||||||||||||||||||||
| c.3890G>C | R1297T 2D AIThe SynGAP1 missense variant R1297T is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Functional prediction tools cluster into two groups: seven tools (REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a benign effect, while two tools (PROVEAN, FATHMM) predict pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (2 benign vs 2 pathogenic), and Foldetta results are unavailable. Overall, the preponderance of evidence points to a benign impact. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.859585 | Disordered | 0.895222 | Binding | 0.511 | 0.817 | 0.625 | -3.941 | Likely Benign | 0.287 | Likely Benign | Likely Benign | 0.151 | Likely Benign | -2.95 | Deleterious | 0.224 | Benign | 0.171 | Benign | 2.49 | Pathogenic | 0.08 | Tolerated | 0.1634 | 0.2972 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3898C>G | P1300A 2D AIThe SynGAP1 missense variant P1300A is reported in gnomAD (ID 6‑33451772‑C‑G) and has no ClinVar entry. Across the evaluated in‑silico tools, every predictor classifies the substitution as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return benign scores. No tool predicts pathogenicity, so the pathogenic‑prediction group is empty. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized indicates a benign effect, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a likely benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not available for this variant. Based on the unanimous benign predictions and the absence of any pathogenic signal, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.885826 | Binding | 0.400 | 0.834 | 0.875 | 6-33451772-C-G | 2 | 1.24e-6 | -3.802 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.065 | Likely Benign | 0.55 | Neutral | 0.008 | Benign | 0.006 | Benign | 3.23 | Benign | 1.00 | Tolerated | 3.77 | 5 | 0.3342 | 0.3744 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||
| c.3898C>T | P1300S 2D AIThe SynGAP1 missense variant P1300S is reported in gnomAD (variant ID 6‑33451772‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only polyPhen‑2 HumDiv flags it as pathogenic, creating a single discordant call. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods indicates that P1300S is most likely benign, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.788093 | Disordered | 0.885826 | Binding | 0.400 | 0.834 | 0.875 | 6-33451772-C-T | 1 | 6.20e-7 | -3.476 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.036 | Likely Benign | -0.29 | Neutral | 0.481 | Possibly Damaging | 0.157 | Benign | 2.89 | Benign | 0.27 | Tolerated | 3.77 | 5 | 0.3242 | 0.4139 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.38T>C | I13T 2D AIThe SynGAP1 missense variant I13T is listed in gnomAD (ID 6‑33420302‑T‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign and the SGM‑Consensus is “Likely Benign.” No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | 6-33420302-T-C | -2.856 | Likely Benign | 0.385 | Ambiguous | Likely Benign | 0.132 | Likely Benign | -0.02 | Neutral | 0.024 | Benign | 0.003 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1401 | 0.1778 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||||||||||||||
| c.38T>G | I13S 2D AIThe SynGAP1 missense variant I13S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus methods also support a benign classification: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for I13S, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.486429 | Structured | 0.482657 | Uncertain | 0.318 | 0.916 | 0.375 | -1.756 | Likely Benign | 0.187 | Likely Benign | Likely Benign | 0.264 | Likely Benign | 0.20 | Neutral | 0.024 | Benign | 0.002 | Benign | 4.06 | Benign | 0.00 | Affected | 0.3292 | 0.1682 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.3901C>A | P1301T 2D AIThe SynGAP1 missense variant P1301T is reported in gnomAD (ID 6‑33451775‑C‑A) and has no ClinVar entry. All evaluated in silico predictors classify it as benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this benign prediction: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are unavailable. Based on the consensus of all available predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.728858 | Disordered | 0.885064 | Binding | 0.447 | 0.841 | 0.875 | 6-33451775-C-A | 1 | 6.20e-7 | -4.945 | Likely Benign | 0.075 | Likely Benign | Likely Benign | 0.090 | Likely Benign | 0.34 | Neutral | 0.000 | Benign | 0.001 | Benign | 2.87 | Benign | 0.47 | Tolerated | 3.77 | 5 | 0.1255 | 0.3312 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||
| c.3901C>G | P1301A 2D AIThe SynGAP1 missense variant P1301A is catalogued in gnomAD (ID 6‑33451775‑C‑G) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the substitution as tolerated or benign. No tool in the dataset predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as likely benign. Foldetta, a protein‑folding stability predictor, was not available for this residue. Overall, the consensus of all available predictions is benign, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.728858 | Disordered | 0.885064 | Binding | 0.447 | 0.841 | 0.875 | 6-33451775-C-G | 1 | 6.20e-7 | -4.290 | Likely Benign | 0.055 | Likely Benign | Likely Benign | 0.088 | Likely Benign | 0.66 | Neutral | 0.003 | Benign | 0.012 | Benign | 3.05 | Benign | 0.88 | Tolerated | 3.77 | 5 | 0.2850 | 0.3087 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||
| c.3908G>T | G1303V 2D AIThe SynGAP1 missense variant G1303V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect, and this is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.871313 | Disordered | 0.886612 | Binding | 0.429 | 0.854 | 0.875 | -3.513 | Likely Benign | 0.162 | Likely Benign | Likely Benign | 0.147 | Likely Benign | -2.44 | Neutral | 0.990 | Probably Damaging | 0.846 | Possibly Damaging | 2.79 | Benign | 0.02 | Affected | 0.1511 | 0.3508 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3910C>T | P1304S 2D AIThe SynGAP1 missense variant P1304S is reported in gnomAD (ID 6‑33451784‑C‑T) but has no ClinVar entry. Functional prediction tools uniformly classify the substitution as benign: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a benign effect. No tool in the dataset indicates pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.885302 | Disordered | 0.886417 | Binding | 0.475 | 0.866 | 0.875 | 6-33451784-C-T | 1 | 6.20e-7 | -4.103 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.039 | Likely Benign | 0.36 | Neutral | 0.059 | Benign | 0.041 | Benign | 3.03 | Benign | 0.58 | Tolerated | 3.77 | 5 | 0.2900 | 0.4442 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3914C>G | T1305R 2D AIThe SynGAP1 missense variant T1305R is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic prediction comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates a benign outcome. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.791621 | Disordered | 0.894658 | Binding | 0.390 | 0.873 | 0.875 | -2.945 | Likely Benign | 0.194 | Likely Benign | Likely Benign | 0.220 | Likely Benign | -1.33 | Neutral | 0.027 | Benign | 0.114 | Benign | 2.75 | Benign | 0.01 | Affected | 0.1115 | 0.3214 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3919C>G | P1307A 2D AIThe SynGAP1 missense variant P1307A is catalogued in gnomAD (ID 6‑33451793‑C‑G) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the substitution as tolerated or benign. No tool in the dataset predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized reports a benign outcome, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as likely benign. Foldetta, a protein‑folding stability predictor, was not available for this variant. Overall, the consensus of all available predictions is benign, and this is consistent with the absence of a ClinVar pathogenic classification. Based on the consensus of all predictions, the variant is most likely benign, and this does not contradict ClinVar status, which has no pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.798249 | Disordered | 0.913511 | Binding | 0.491 | 0.901 | 0.875 | 6-33451793-C-G | 1 | 6.20e-7 | -4.042 | Likely Benign | 0.078 | Likely Benign | Likely Benign | 0.064 | Likely Benign | 0.49 | Neutral | 0.003 | Benign | 0.006 | Benign | 3.11 | Benign | 1.00 | Tolerated | 3.77 | 5 | 0.3747 | 0.4451 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||
| c.3926T>G | V1309G 2D AIThe SynGAP1 missense variant V1309G is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic), and Foldetta results are unavailable. Overall, the majority of evidence points toward a benign impact, and this conclusion does not contradict any ClinVar annotation because the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.712013 | Disordered | 0.948596 | Binding | 0.402 | 0.907 | 0.750 | -2.618 | Likely Benign | 0.160 | Likely Benign | Likely Benign | 0.326 | Likely Benign | -3.40 | Deleterious | 0.391 | Benign | 0.767 | Possibly Damaging | 2.40 | Pathogenic | 0.00 | Affected | 0.1901 | 0.2144 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3929C>G | T1310R 2D AIThe SynGAP1 missense variant T1310R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as tolerated or benign. Only SIFT predicts a damaging outcome, labeling it pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign status. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign; Foldetta results are unavailable. Consequently, the aggregate evidence indicates that T1310R is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.959076 | Binding | 0.398 | 0.904 | 0.750 | -3.632 | Likely Benign | 0.226 | Likely Benign | Likely Benign | 0.231 | Likely Benign | -0.59 | Neutral | 0.001 | Benign | 0.006 | Benign | 2.78 | Benign | 0.03 | Affected | 0.0844 | 0.2501 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.3929C>T | T1310M 2D 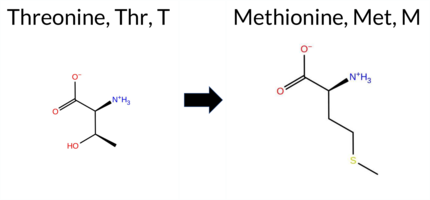 AIThe SynGAP1 missense variant T1310M is listed in ClinVar (ID 2160201.0) as Benign and is present in gnomAD (gnomAD ID 6‑33451803‑C‑T). All evaluated in‑silico predictors report a benign outcome: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the computational evidence overwhelmingly supports a benign classification, which aligns with the ClinVar status and does not contradict it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.812494 | Disordered | 0.959076 | Binding | 0.398 | 0.904 | 0.750 | Benign | 1 | 6-33451803-C-T | 17 | 1.05e-5 | -4.822 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.069 | Likely Benign | 2.19 | Neutral | 0.021 | Benign | 0.005 | Benign | 2.98 | Benign | 0.93 | Tolerated | 3.77 | 5 | 0.1144 | 0.5397 | -1 | -1 | 2.6 | 30.09 | ||||||||||||||||||||||||||||||||
| c.392G>T | G131V 2D AIThe SynGAP1 missense variant G131V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a tie and is therefore unavailable, and Foldetta (combining FoldX‑MD and Rosetta outputs) has no result for this variant. Overall, the balance of evidence favors a pathogenic classification, and this assessment does not contradict any existing ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.429200 | Structured | 0.724779 | Binding | 0.302 | 0.891 | 0.250 | -6.248 | Likely Benign | 0.795 | Likely Pathogenic | Ambiguous | 0.199 | Likely Benign | -4.25 | Deleterious | 0.983 | Probably Damaging | 0.559 | Possibly Damaging | 3.92 | Benign | 0.00 | Affected | 0.1168 | 0.4175 | -1 | -3 | 4.6 | 42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3940C>A | P1314T 2D AIThe SynGAP1 missense variant P1314T is present in gnomAD (ID 6‑33451814‑C‑A) but has no ClinVar entry. Across a broad panel of in silico predictors, every tool reports a benign effect. Benign predictions come from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool in the set predicts pathogenicity, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized returns a benign prediction, and the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status remains unavailable. Overall, the computational evidence overwhelmingly supports a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Thus, the variant is most likely benign, and this is consistent with the lack of a ClinVar pathogenic designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.971592 | Binding | 0.467 | 0.903 | 0.750 | 6-33451814-C-A | -4.083 | Likely Benign | 0.096 | Likely Benign | Likely Benign | 0.066 | Likely Benign | 1.26 | Neutral | 0.061 | Benign | 0.045 | Benign | 4.32 | Benign | 0.97 | Tolerated | 3.77 | 5 | 0.1791 | 0.5626 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3940C>G | P1314A 2D AIThe SynGAP1 missense variant P1314A is catalogued in gnomAD (ID 6‑33451814‑C‑G) but has no ClinVar entry. Across the spectrum of in silico predictors, every tool listed—REVEL, PROVEAN, PolyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—returns a benign or likely benign verdict. No pathogenic predictions appear in the dataset, so the pathogenic group is empty. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized scores benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely benign. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Taken together, the evidence overwhelmingly supports a benign classification, and this conclusion does not conflict with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.971592 | Binding | 0.467 | 0.903 | 0.750 | 6-33451814-C-G | 1 | 6.22e-7 | -3.540 | Likely Benign | 0.061 | Likely Benign | Likely Benign | 0.043 | Likely Benign | -0.37 | Neutral | 0.001 | Benign | 0.002 | Benign | 4.23 | Benign | 0.93 | Tolerated | 3.77 | 5 | 0.3502 | 0.4441 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||
| c.3941C>A | P1314Q 2D AIThe SynGAP1 missense variant P1314Q is listed in gnomAD (ID 6‑33451815‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign. No Foldetta stability data are available, so folding‑stability evidence is unavailable. Overall, the preponderance of computational evidence indicates that P1314Q is most likely benign, and this conclusion is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.882776 | Disordered | 0.971592 | Binding | 0.467 | 0.903 | 0.750 | 6-33451815-C-A | -4.222 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 0.039 | Likely Benign | -0.55 | Neutral | 0.618 | Possibly Damaging | 0.333 | Benign | 4.24 | Benign | 0.04 | Affected | 3.77 | 5 | 0.1612 | 0.4588 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.394T>G | F132V 2D AIThe SynGAP1 missense variant F132V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that agree on a pathogenic effect are PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of high‑confidence predictions indicate a pathogenic effect, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.450668 | Structured | 0.727897 | Binding | 0.345 | 0.892 | 0.250 | -7.637 | In-Between | 0.979 | Likely Pathogenic | Likely Pathogenic | 0.328 | Likely Benign | -3.86 | Deleterious | 0.084 | Benign | 0.034 | Benign | 3.38 | Benign | 0.00 | Affected | 0.2349 | 0.1725 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.3950G>A | G1317D 2D AIThe SynGAP1 missense variant G1317D is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (ID 6‑33451824‑G‑A). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and SIFT. AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign prediction (2 benign vs. 1 pathogenic, with one uncertain). Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict ClinVar, which has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | 6-33451824-G-A | -4.860 | Likely Benign | 0.520 | Ambiguous | Likely Benign | 0.081 | Likely Benign | -3.54 | Deleterious | 0.588 | Possibly Damaging | 0.212 | Benign | 4.03 | Benign | 0.00 | Affected | 3.77 | 5 | 0.2115 | 0.2859 | -1 | 1 | -3.1 | 58.04 | |||||||||||||||||||||||||||||||||||||
| c.3950G>T | G1317V 2D AIThe SynGAP1 missense variant G1317V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. In contrast, PROVEAN and SIFT predict a pathogenic impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with the absence of a ClinVar assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.908098 | Disordered | 0.971158 | Binding | 0.385 | 0.879 | 0.750 | -4.604 | Likely Benign | 0.286 | Likely Benign | Likely Benign | 0.050 | Likely Benign | -2.99 | Deleterious | 0.004 | Benign | 0.004 | Benign | 4.05 | Benign | 0.00 | Affected | 0.1132 | 0.3635 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.3955G>C | A1319P 2D AIThe SynGAP1 missense variant A1319P is reported in ClinVar as “Not submitted” and is present in gnomAD (ID 6‑33451829‑G‑C). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. The SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the ClinVar status, which currently contains no pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.842060 | Disordered | 0.960481 | Binding | 0.454 | 0.851 | 0.750 | 6-33451829-G-C | 3 | 1.95e-6 | -2.783 | Likely Benign | 0.057 | Likely Benign | Likely Benign | 0.104 | Likely Benign | -0.81 | Neutral | 0.992 | Probably Damaging | 0.878 | Possibly Damaging | 4.06 | Benign | 0.03 | Affected | 3.77 | 5 | 0.2360 | 0.5715 | -1 | 1 | -3.4 | 26.04 | ||||||||||||||||||||||||||||||||||
| c.3958C>A | P1320T 2D AIThe SynGAP1 missense variant P1320T is reported in gnomAD (variant ID 6‑33451832‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: the majority (REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a benign effect, while a minority (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT) predict pathogenicity. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.899122 | Disordered | 0.946297 | Binding | 0.510 | 0.833 | 0.750 | 6-33451832-C-A | 1 | 6.42e-7 | -5.355 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.091 | Likely Benign | -0.86 | Neutral | 0.994 | Probably Damaging | 0.981 | Probably Damaging | 4.21 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1787 | 0.6392 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||
| c.3961C>A | P1321T 2D AIThe SynGAP1 missense variant P1321T is catalogued in gnomAD (ID 6‑33451835‑C‑A) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all score the variant as benign. No tool predicts pathogenicity. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta, a protein‑folding stability method, has no available result for this variant. Overall, the evidence strongly supports a benign classification, and this conclusion is consistent with the absence of a ClinVar pathogenic designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.933505 | Binding | 0.463 | 0.828 | 0.875 | 6-33451835-C-A | -4.946 | Likely Benign | 0.074 | Likely Benign | Likely Benign | 0.058 | Likely Benign | 1.07 | Neutral | 0.115 | Benign | 0.012 | Benign | 4.25 | Benign | 1.00 | Tolerated | 3.77 | 5 | 0.2016 | 0.5901 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3961C>G | P1321A 2D AIThe SynGAP1 missense variant P1321A is listed in ClinVar with an uncertain significance (ClinVar ID 4697583) and is present in gnomAD (ID 6‑33451835‑C‑G). All evaluated in‑silico predictors classify the substitution as benign: REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “likely benign,” and the Foldetta stability analysis is unavailable. Overall, the computational evidence indicates that the variant is most likely benign, and this conclusion does not contradict the current ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.827927 | Disordered | 0.933505 | Binding | 0.463 | 0.828 | 0.875 | Uncertain | 1 | 6-33451835-C-G | -4.411 | Likely Benign | 0.051 | Likely Benign | Likely Benign | 0.058 | Likely Benign | -0.33 | Neutral | 0.001 | Benign | 0.000 | Benign | 4.29 | Benign | 0.42 | Tolerated | 3.77 | 5 | 0.3615 | 0.4716 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||
| c.3967C>A | P1323T 2D AIThe SynGAP1 missense variant P1323T is reported in gnomAD (ID 6‑33451841‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as tolerated or benign. Only two algorithms—polyPhen‑2 HumDiv and SIFT—suggest a damaging impact. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus also indicates benign. Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not conflict with any ClinVar classification, as none is present. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451841-C-A | -6.296 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.064 | Likely Benign | -0.37 | Neutral | 0.588 | Possibly Damaging | 0.227 | Benign | 3.95 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1988 | 0.5161 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3967C>G | P1323A 2D AIThe SynGAP1 missense variant P1323A is catalogued in gnomAD (ID 6‑33451841‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes, while the single pathogenic signal comes from SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign classification. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote) is likely benign; Foldetta results are not available. Overall, the consensus of available predictions points to a benign impact for P1323A, and this conclusion is not contradicted by ClinVar status, which is currently absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451841-C-G | -4.802 | Likely Benign | 0.053 | Likely Benign | Likely Benign | 0.040 | Likely Benign | -0.73 | Neutral | 0.011 | Benign | 0.017 | Benign | 3.92 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3743 | 0.3976 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||||
| c.3968C>A | P1323Q 2D AIThe SynGAP1 missense variant P1323Q is reported in gnomAD (ID 6‑33451842‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. In contrast, polyPhen‑2 HumDiv and SIFT predict pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence indicates that P1323Q is most likely benign, and this conclusion is not contradicted by any ClinVar classification (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.901269 | Disordered | 0.907659 | Binding | 0.489 | 0.814 | 0.875 | 6-33451842-C-A | -6.134 | Likely Benign | 0.088 | Likely Benign | Likely Benign | 0.033 | Likely Benign | -0.85 | Neutral | 0.712 | Possibly Damaging | 0.328 | Benign | 3.82 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1791 | 0.4657 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3970C>A | P1324T 2D AIThe SynGAP1 missense variant P1324T is listed in gnomAD (ID 6‑33451844‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions come from polyPhen‑2 HumDiv and SIFT. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the consensus of the available predictions points to a benign effect, and this conclusion does not contradict any ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | 6-33451844-C-A | -5.289 | Likely Benign | 0.077 | Likely Benign | Likely Benign | 0.072 | Likely Benign | 0.95 | Neutral | 0.588 | Possibly Damaging | 0.175 | Benign | 4.43 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1863 | 0.4761 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3971C>A | P1324Q 2D AIThe SynGAP1 missense variant P1324Q is reported in gnomAD (ID 6‑33451845‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign status. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the consensus of the majority of tools, including the high‑accuracy predictors, points to a benign effect. This prediction is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.891961 | Disordered | 0.899181 | Binding | 0.432 | 0.793 | 0.875 | 6-33451845-C-A | -5.577 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 0.028 | Likely Benign | -0.69 | Neutral | 0.027 | Benign | 0.010 | Benign | 4.27 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1744 | 0.4736 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||||||
| c.3973C>A | P1325T 2D AIThe SynGAP1 missense variant P1325T is reported in gnomAD (ID 6‑33451847‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign effect for P1325T, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451847-C-A | -5.880 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.135 | Likely Benign | -0.14 | Neutral | 0.000 | Benign | 0.001 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1814 | 0.5150 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3973C>G | P1325A 2D AIThe SynGAP1 missense variant P1325A is reported in gnomAD (ID 6‑33451847‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign. Only SIFT predicts a pathogenic effect. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy methods confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign impact for P1325A, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451847-C-G | -3.786 | Likely Benign | 0.058 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -0.95 | Neutral | 0.019 | Benign | 0.004 | Benign | 4.10 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3650 | 0.4016 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||||
| c.3973C>T | P1325S 2D AIThe SynGAP1 missense variant P1325S is reported in gnomAD (ID 6‑33451847‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.876521 | Disordered | 0.893621 | Binding | 0.439 | 0.791 | 0.875 | 6-33451847-C-T | 1 | 6.43e-7 | -5.273 | Likely Benign | 0.083 | Likely Benign | Likely Benign | 0.053 | Likely Benign | 0.29 | Neutral | 0.010 | Benign | 0.007 | Benign | 4.10 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3657 | 0.4360 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.3976C>A | P1326T 2D AIThe SynGAP1 missense variant P1326T is reported in gnomAD (ID 6‑33451850‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports a benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority of the same four high‑accuracy tools) is benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that P1326T is most likely benign, and this assessment does not contradict any ClinVar classification because no ClinVar variant is listed. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | 6-33451850-C-A | -5.755 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.135 | Likely Benign | -0.26 | Neutral | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 3.71 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1955 | 0.5517 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3976C>T | P1326S 2D AIThe SynGAP1 missense variant P1326S is listed in ClinVar with no submitted interpretation and is present in gnomAD (ID 6‑33451850‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence—including the high‑accuracy tools—supports a benign classification. This conclusion does not contradict the ClinVar status, which currently has no pathogenic assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | 6-33451850-C-T | -5.221 | Likely Benign | 0.121 | Likely Benign | Likely Benign | 0.107 | Likely Benign | -0.35 | Neutral | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 3.74 | Benign | 0.00 | Affected | 3.77 | 5 | 0.3722 | 0.4563 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||
| c.3977C>A | P1326Q 2D AIThe SynGAP1 missense variant P1326Q is listed in ClinVar (ID 2806103.0) with an “Uncertain” status and is present in gnomAD (variant ID 6‑33451851‑C‑A). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar “Uncertain” classification, which remains inconclusive. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.948786 | Disordered | 0.887377 | Binding | 0.393 | 0.782 | 0.875 | Uncertain | 1 | 6-33451851-C-A | 1 | 6.40e-7 | -5.422 | Likely Benign | 0.128 | Likely Benign | Likely Benign | 0.138 | Likely Benign | -0.86 | Neutral | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.62 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1769 | 0.4457 | -1 | 0 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||||
| c.3979C>A | P1327T 2D AIThe SynGAP1 missense variant P1327T is reported in gnomAD (ID 6‑33451853‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only the two polyPhen‑2 implementations (HumDiv and HumVar) predict a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign; Foldetta results are not available. Taken together, the majority of evidence points to a benign impact. There is no ClinVar classification to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.896620 | Disordered | 0.900145 | Binding | 0.369 | 0.777 | 0.875 | 6-33451853-C-A | -5.263 | Likely Benign | 0.150 | Likely Benign | Likely Benign | 0.144 | Likely Benign | 1.20 | Neutral | 0.994 | Probably Damaging | 0.908 | Possibly Damaging | 4.51 | Benign | 1.00 | Tolerated | 3.77 | 5 | 0.1386 | 0.4779 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.3979C>G | P1327A 2D AIThe SynGAP1 missense variant P1327A is reported in gnomAD (ID 6‑33451853‑C‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports it as likely benign. In contrast, polyPhen‑2 (HumDiv and HumVar) predict the variant to be pathogenic. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this is consistent with the absence of a pathogenic ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.896620 | Disordered | 0.900145 | Binding | 0.369 | 0.777 | 0.875 | 6-33451853-C-G | 2 | 1.28e-6 | -4.661 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 0.072 | Likely Benign | -0.37 | Neutral | 0.980 | Probably Damaging | 0.811 | Possibly Damaging | 4.24 | Benign | 0.42 | Tolerated | 3.77 | 5 | 0.3152 | 0.3645 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||
| c.3989A>C | Q1330P 2D AIThe SynGAP1 missense variant Q1330P is listed in gnomAD (ID 6‑33451863‑A‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, and FATHMM; pathogenic predictions come from polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments are mixed: AlphaMissense‑Optimized yields an “Uncertain” result, SGM‑Consensus confirms a benign leaning, and Foldetta (combining FoldX‑MD and Rosetta outputs) is not available for this variant. Overall, the balance of evidence favors a benign effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.943310 | Disordered | 0.931969 | Binding | 0.369 | 0.752 | 0.875 | 6-33451863-A-C | -3.087 | Likely Benign | 0.814 | Likely Pathogenic | Ambiguous | 0.076 | Likely Benign | -2.45 | Neutral | 0.898 | Possibly Damaging | 0.441 | Benign | 3.91 | Benign | 0.03 | Affected | 3.77 | 5 | 0.2266 | 0.4983 | -1 | 0 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||||
| c.3992T>G | I1331S 2D AIThe SynGAP1 I1331S variant is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that agree on a pathogenic effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenicity, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta results are unavailable. Overall, the majority of evidence (six pathogenic vs. three benign predictions) indicates that the variant is most likely pathogenic. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.921076 | Disordered | 0.941705 | Binding | 0.359 | 0.752 | 0.875 | -3.014 | Likely Benign | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.230 | Likely Benign | -3.40 | Deleterious | 0.984 | Probably Damaging | 0.969 | Probably Damaging | 3.30 | Benign | 0.00 | Affected | 0.2929 | 0.0836 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3995C>A | T1332K 2D 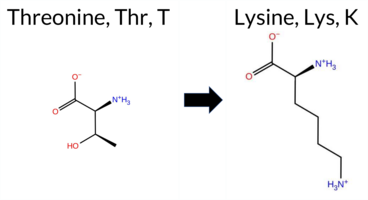 AIThe SynGAP1 missense variant T1332K is catalogued in gnomAD (ID 6‑33451869‑C‑A) but has no ClinVar submission. Functional prediction tools cluster into two groups: benign predictions from REVEL, ESM1b, and FATHMM; pathogenic predictions from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. A high‑accuracy consensus (SGM) derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN yields a 2‑to‑2 split, leaving the consensus inconclusive. No Foldetta stability assessment is available. Overall, the majority of evidence (five pathogenic versus three benign) points to a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | 6-33451869-C-A | -3.264 | Likely Benign | 0.935 | Likely Pathogenic | Ambiguous | 0.247 | Likely Benign | -3.48 | Deleterious | 0.998 | Probably Damaging | 0.989 | Probably Damaging | 2.96 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1424 | 0.4376 | -1 | 0 | -3.2 | 27.07 | |||||||||||||||||||||||||||||||||||||
| c.3995C>G | T1332R 2D AIThe SynGAP1 missense variant T1332R is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM, whereas a majority of tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default) predict a pathogenic impact. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑vs‑2 tie and therefore unavailable; Foldetta results are not provided. Overall, the balance of evidence (five pathogenic versus three benign predictions) indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | -3.354 | Likely Benign | 0.878 | Likely Pathogenic | Ambiguous | 0.271 | Likely Benign | -3.59 | Deleterious | 0.998 | Probably Damaging | 0.993 | Probably Damaging | 2.96 | Benign | 0.00 | Affected | 0.1173 | 0.3867 | -1 | -1 | -3.8 | 55.08 | ||||||||||||||||||||||||||||||||||||||||
| c.3995C>T | T1332M 2D AISynGAP1 missense variant T1332M is listed as Benign in ClinVar (ID 794425) and is present in gnomAD (6‑33451869‑C‑T). Functional prediction tools show mixed results: benign calls come from REVEL, ESM1b, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized returned an uncertain result, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie, and no Foldetta stability data are available. Overall, the majority of evidence points toward a pathogenic effect, which contradicts the ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.915074 | Disordered | 0.948427 | Binding | 0.442 | 0.754 | 0.875 | Likely Benign | 1 | 6-33451869-C-T | 20 | 1.86e-5 | -4.107 | Likely Benign | 0.948 | Likely Pathogenic | Ambiguous | 0.252 | Likely Benign | -3.63 | Deleterious | 1.000 | Probably Damaging | 0.991 | Probably Damaging | 2.95 | Benign | 0.00 | Affected | 3.77 | 5 | 0.1551 | 0.6513 | -1 | -1 | 2.6 | 30.09 | |||||||||||||||||||||||||||||||||
| c.4004G>A | G1335D 2D AIThe SynGAP1 missense variant G1335D is reported in ClinVar as “not listed” and is present in gnomAD (variant ID 6‑33451878‑G‑A). Functional prediction tools that agree on a benign effect are REVEL and ESM1b, whereas the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta results are not available for this variant. Overall, the preponderance of evidence from multiple prediction algorithms and the SGM‑Consensus suggests that G1335D is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | 6-33451878-G-A | -5.687 | Likely Benign | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.404 | Likely Benign | -4.42 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 3.77 | 5 | 0.2120 | 0.3702 | -1 | 1 | -3.1 | 58.04 | ||||||||||||||||||||||||||||||||||||
| c.4004G>T | G1335V 2D AIThe SynGAP1 missense variant G1335V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL and ESM1b, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a pathogenic impact, and this assessment does not conflict with the ClinVar status, which currently has no entry for G1335V. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.891961 | Disordered | 0.967705 | Binding | 0.323 | 0.724 | 0.750 | -4.982 | Likely Benign | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.394 | Likely Benign | -5.65 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.02 | Pathogenic | 0.00 | Affected | 0.1124 | 0.4195 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||||||||||||
| c.4009T>G | F1337V 2D AIThe SynGAP1 missense variant F1337V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive (two pathogenic vs two benign), and Foldetta results are unavailable. Overall, the majority of evidence (six pathogenic vs three benign) points to a pathogenic impact. The variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.823549 | Disordered | 0.979265 | Binding | 0.388 | 0.712 | 0.625 | -2.425 | Likely Benign | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.335 | Likely Benign | -3.94 | Deleterious | 0.947 | Possibly Damaging | 0.932 | Probably Damaging | 2.76 | Benign | 0.00 | Affected | 0.2470 | 0.2902 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.4018A>C | T1340P 2D 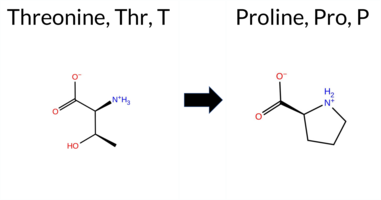 AIThe SynGAP1 missense variant T1340P is catalogued in gnomAD (ID 6‑33451892‑A‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes, while only SIFT predicts a pathogenic impact. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is Likely Benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that T1340P is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.852992 | Disordered | 0.977899 | Binding | 0.444 | 0.697 | 0.750 | 6-33451892-A-C | -2.681 | Likely Benign | 0.102 | Likely Benign | Likely Benign | 0.190 | Likely Benign | -1.81 | Neutral | 0.334 | Benign | 0.099 | Benign | 4.08 | Benign | 0.01 | Affected | 3.77 | 5 | 0.2268 | 0.4983 | -1 | 0 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||||||||
| c.4019C>T | T1340I 2D AIThe SynGAP1 missense variant T1340I is not reported in ClinVar (ClinVar status: “None”) but is present in gnomAD (ID 6‑33451893‑C‑T). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are PROVEAN and SIFT. AlphaMissense‑Default is uncertain, while AlphaMissense‑Optimized predicts benign. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to benign (two benign versus one pathogenic, with the uncertain result treated as unavailable). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the high‑accuracy predictions (AlphaMissense‑Optimized, SGM Consensus) both indicate a benign impact, and no evidence contradicts this assessment with the ClinVar status. Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.852992 | Disordered | 0.977899 | Binding | 0.444 | 0.697 | 0.750 | 6-33451893-C-T | -3.476 | Likely Benign | 0.402 | Ambiguous | Likely Benign | 0.089 | Likely Benign | -2.57 | Deleterious | 0.334 | Benign | 0.099 | Benign | 4.08 | Benign | 0.01 | Affected | 3.77 | 5 | 0.1163 | 0.5848 | -1 | 0 | 5.2 | 12.05 | |||||||||||||||||||||||||||||||||||||
| c.401G>T | S134I 2D AIThe SynGAP1 missense variant S134I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic and the SGM‑Consensus as Likely Pathogenic; Foldetta results are unavailable. Based on the collective predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.447574 | Structured | 0.695837 | Binding | 0.333 | 0.898 | 0.250 | -8.860 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.230 | Likely Benign | -3.27 | Deleterious | 0.041 | Benign | 0.031 | Benign | 3.81 | Benign | 0.00 | Affected | 0.0683 | 0.5191 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.4025A>G | D1342G 2D AIThe SynGAP1 missense variant D1342G is reported in gnomAD (variant ID 6‑33451899‑A‑G) but has no ClinVar entry. In silico prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. No tool predicts pathogenicity. The high‑accuracy consensus methods corroborate this: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Benign.” Foldetta results are not available. Consequently, the variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.921076 | Disordered | 0.981682 | Binding | 0.316 | 0.678 | 0.875 | 6-33451899-A-G | 1 | 7.29e-7 | -3.227 | Likely Benign | 0.129 | Likely Benign | Likely Benign | 0.021 | Likely Benign | -0.89 | Neutral | 0.225 | Benign | 0.045 | Benign | 4.05 | Benign | 0.09 | Tolerated | 4.32 | 4 | 0.3633 | 0.5601 | -1 | 1 | 3.1 | -58.04 | ||||||||||||||||||||||||||||||||||
| c.40C>A | P14T 2D AIThe SynGAP1 missense variant P14T is reported in gnomAD (ID 6‑33420304‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also reports likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for P14T, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.433034 | Structured | 0.471596 | Uncertain | 0.399 | 0.909 | 0.375 | 6-33420304-C-A | -3.261 | Likely Benign | 0.150 | Likely Benign | Likely Benign | 0.078 | Likely Benign | -0.27 | Neutral | 0.212 | Benign | 0.014 | Benign | 4.20 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1912 | 0.6658 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.416G>T | S139I 2D AIThe SynGAP1 missense variant S139I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; pathogenic predictions come from PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy tools points to a pathogenic effect. This conclusion is consistent with the absence of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.553315 | Disordered | 0.600637 | Binding | 0.353 | 0.900 | 0.250 | -9.228 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 0.203 | Likely Benign | -3.36 | Deleterious | 0.380 | Benign | 0.102 | Benign | 4.09 | Benign | 0.03 | Affected | 0.0948 | 0.4796 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.422T>G | I141S 2D AIThe SynGAP1 missense variant I141S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus also as Likely Pathogenic; the Foldetta protein‑folding stability analysis is unavailable. Overall, the majority of evidence points toward a pathogenic impact for I141S, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.465241 | Structured | 0.577021 | Binding | 0.367 | 0.877 | 0.500 | -11.874 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.267 | Likely Benign | -3.28 | Deleterious | 0.567 | Possibly Damaging | 0.249 | Benign | 3.56 | Benign | 0.00 | Affected | 0.2885 | 0.0840 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||||||||||||
| c.431C>G | T144R 2D AIThe SynGAP1 missense variant T144R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign calls come from REVEL, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic effect. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the aggregate predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | -13.331 | Likely Pathogenic | 0.922 | Likely Pathogenic | Ambiguous | 0.177 | Likely Benign | -2.83 | Deleterious | 0.609 | Possibly Damaging | 0.150 | Benign | 3.75 | Benign | 0.00 | Affected | 0.1188 | 0.2823 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||||||||||||
| c.431C>T | T144M 2D AIThe SynGAP1 missense variant T144M is listed in ClinVar with an “Uncertain” status (ClinVar ID 2231966.0) and is present in the gnomAD database (gnomAD ID 6‑33432728‑C‑T). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM. Those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Pathogenic.” High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain,” SGM‑Consensus as “Likely Pathogenic,” and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of computational predictions lean toward a pathogenic impact, and this assessment does not contradict the ClinVar designation of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.666105 | Disordered | 0.524000 | Binding | 0.335 | 0.838 | 0.625 | Uncertain | 2 | 6-33432728-C-T | 2 | 1.30e-6 | -11.228 | Likely Pathogenic | 0.922 | Likely Pathogenic | Ambiguous | 0.118 | Likely Benign | -3.16 | Deleterious | 0.913 | Possibly Damaging | 0.333 | Benign | 3.73 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1504 | 0.6510 | -1 | -1 | 2.6 | 30.09 | ||||||||||||||||||||||||||||||||
| c.442C>G | P148A 2D AIThe SynGAP1 missense variant P148A is catalogued in gnomAD (ID 6‑33432739‑C‑G) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Benign” because three of the four contributing tools predict benign. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect. Thus, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.653063 | Disordered | 0.500109 | Binding | 0.372 | 0.837 | 0.625 | 6-33432739-C-G | 1 | 6.33e-7 | -6.890 | Likely Benign | 0.729 | Likely Pathogenic | Likely Benign | 0.117 | Likely Benign | -2.31 | Neutral | 0.999 | Probably Damaging | 0.991 | Probably Damaging | 4.00 | Benign | 0.03 | Affected | 3.61 | 5 | 0.3134 | 0.4123 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||||||||||||
| c.442C>T | P148S 2D AIThe SynGAP1 missense variant P148S is not reported in ClinVar (ClinVar status: not listed) and is present in gnomAD (ID 6‑33432739‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Benign, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) is unavailable for this variant. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.653063 | Disordered | 0.500109 | Binding | 0.372 | 0.837 | 0.625 | 6-33432739-C-T | 1 | 6.33e-7 | -3.258 | Likely Benign | 0.874 | Likely Pathogenic | Ambiguous | 0.102 | Likely Benign | -1.81 | Neutral | 1.000 | Probably Damaging | 0.994 | Probably Damaging | 4.05 | Benign | 0.39 | Tolerated | 3.61 | 5 | 0.3143 | 0.4597 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.44C>A | A15E 2D AIThe SynGAP1 missense variant A15E is reported in gnomAD (ID 6‑33420308‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is provided). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.436924 | Structured | 0.466055 | Uncertain | 0.330 | 0.912 | 0.375 | 6-33420308-C-A | -3.423 | Likely Benign | 0.277 | Likely Benign | Likely Benign | 0.169 | Likely Benign | 0.46 | Neutral | 0.406 | Benign | 0.040 | Benign | 4.13 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1563 | 0.1908 | -1 | 0 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||||||||
| c.451G>C | D151H 2D AIThe SynGAP1 missense variant D151H is listed in ClinVar with an “Uncertain” status and is present in gnomAD (variant ID 6‑33432748‑G‑C). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict a pathogenic outcome. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized classifies the variant as pathogenic, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as “Likely Pathogenic.” No Foldetta stability result is available for this variant. Overall, the majority of computational evidence indicates that D151H is most likely pathogenic, which does not contradict the current ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.529623 | Disordered | 0.503277 | Binding | 0.342 | 0.841 | 0.625 | Uncertain | 1 | 6-33432748-G-C | 2 | 1.26e-6 | -11.747 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.335 | Likely Benign | -3.90 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 3.86 | Benign | 0.00 | Affected | 3.61 | 5 | 0.1543 | 0.8419 | -1 | 1 | 0.3 | 22.05 | ||||||||||||||||||||||||||||||||
| c.461G>T | S154I 2D AIThe SynGAP1 missense variant S154I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two benign, two pathogenic) and therefore unavailable as evidence. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, was not provided and is treated as unavailable. High‑accuracy predictions therefore indicate a benign outcome (AlphaMissense‑Optimized) with no decisive evidence from SGM Consensus or Foldetta. Overall, the variant is most likely benign, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.508330 | Binding | 0.284 | 0.795 | 0.500 | -10.061 | Likely Pathogenic | 0.665 | Likely Pathogenic | Likely Benign | 0.105 | Likely Benign | -2.24 | Neutral | 0.990 | Probably Damaging | 0.797 | Possibly Damaging | 4.03 | Benign | 0.07 | Tolerated | 0.0953 | 0.5123 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.464G>T | S155I 2D AIThe SynGAP1 missense variant S155I is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL and FATHMM, whereas the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict it to be pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized scores the variant as pathogenic, the SGM‑Consensus indicates a likely pathogenic status, and the Foldetta stability analysis is unavailable. Taken together, the majority of evidence points to a pathogenic impact for S155I. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.497853 | Structured | 0.515359 | Binding | 0.292 | 0.787 | 0.500 | -11.298 | Likely Pathogenic | 0.972 | Likely Pathogenic | Likely Pathogenic | 0.264 | Likely Benign | -2.70 | Deleterious | 0.995 | Probably Damaging | 0.986 | Probably Damaging | 3.81 | Benign | 0.00 | Affected | 0.0764 | 0.5711 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.466T>G | F156V 2D AIThe SynGAP1 missense variant F156V is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of algorithms—PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—classify the change as pathogenic. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is uncertain, but the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely pathogenic status. Foldetta, a protein‑folding stability predictor that integrates FoldX‑MD and Rosetta outputs, was not available for this variant. Overall, the preponderance of evidence points to a pathogenic effect for F156V, and this conclusion does not conflict with the current ClinVar annotation, which is absent. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.476583 | Structured | 0.521964 | Binding | 0.284 | 0.785 | 0.500 | -11.945 | Likely Pathogenic | 0.950 | Likely Pathogenic | Ambiguous | 0.246 | Likely Benign | -2.91 | Deleterious | 0.981 | Probably Damaging | 0.954 | Probably Damaging | 4.03 | Benign | 0.00 | Affected | 0.2177 | 0.2166 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||||||||||||
| c.476T>G | I159S 2D AIThe SynGAP1 missense variant I159S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM. Tools that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized tool rates the variant as uncertain, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑vs‑2 split, and Foldetta results are unavailable. Overall, the majority of available predictions (five pathogenic vs. three benign) lean toward pathogenicity. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.454136 | Structured | 0.529953 | Binding | 0.278 | 0.731 | 0.125 | -13.684 | Likely Pathogenic | 0.905 | Likely Pathogenic | Ambiguous | 0.241 | Likely Benign | -1.70 | Neutral | 0.995 | Probably Damaging | 0.979 | Probably Damaging | 3.85 | Benign | 0.00 | Affected | 0.2854 | 0.0712 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.47T>C | M16T 2D AIThe SynGAP1 missense variant M16T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores the variant as benign and the SGM‑Consensus classifies it as likely benign. No Foldetta stability analysis is available for this residue, so folding‑stability evidence is lacking. Overall, the majority of computational evidence indicates that M16T is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.440853 | Structured | 0.459925 | Uncertain | 0.346 | 0.908 | 0.375 | -2.060 | Likely Benign | 0.515 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -0.46 | Neutral | 0.006 | Benign | 0.001 | Benign | 4.23 | Benign | 0.00 | Affected | 0.2385 | 0.2545 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.487T>G | F163V 2D AIThe SynGAP1 missense variant F163V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, and FATHMM, whereas polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default all predict a pathogenic impact. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: AlphaMissense‑Optimized remains uncertain; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and therefore unavailable; Foldetta data are not provided. Overall, the majority of available predictions (five pathogenic vs. three benign) indicate that the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.433034 | Structured | 0.513928 | Binding | 0.325 | 0.686 | 0.375 | -12.580 | Likely Pathogenic | 0.927 | Likely Pathogenic | Ambiguous | 0.236 | Likely Benign | -2.06 | Neutral | 0.981 | Probably Damaging | 0.954 | Probably Damaging | 4.13 | Benign | 0.02 | Affected | 0.2214 | 0.2547 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.494G>T | S165I 2D AISynGAP1 missense variant S165I has no ClinVar record and is absent from gnomAD. Individual prediction tools cluster into two groups: benign predictions come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus remains Likely Pathogenic; Foldetta stability analysis is unavailable. Overall, the majority of single‑tool predictions favor a benign effect, whereas the consensus score suggests pathogenicity. Consequently, the variant is most likely benign according to the bulk of evidence, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.509123 | Binding | 0.324 | 0.644 | 0.250 | -11.304 | Likely Pathogenic | 0.774 | Likely Pathogenic | Likely Benign | 0.259 | Likely Benign | -2.53 | Deleterious | 0.084 | Benign | 0.031 | Benign | 3.96 | Benign | 0.00 | Affected | 0.0943 | 0.5027 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.4A>C | S2R 2D AIThe SynGAP1 missense variant S2R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.543646 | Binding | 0.382 | 0.922 | 0.750 | -3.684 | Likely Benign | 0.426 | Ambiguous | Likely Benign | 0.062 | Likely Benign | -0.44 | Neutral | 0.117 | Benign | 0.008 | Benign | 4.05 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0996 | 0.4503 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.521T>C | M174T 2D AIThe SynGAP1 missense variant M174T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, and the SGM‑Consensus (majority vote) also indicates Likely Pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.661982 | Disordered | 0.485854 | Uncertain | 0.373 | 0.620 | 0.375 | -9.174 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.286 | Likely Benign | -3.12 | Deleterious | 0.244 | Benign | 0.049 | Benign | 4.08 | Benign | 0.01 | Affected | 0.1949 | 0.2334 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||||||
| c.524A>C | Q175P 2D AIThe SynGAP1 missense variant Q175P is reported in gnomAD (ID 6‑33435166‑A‑C) but has no ClinVar entry. Functional prediction tools uniformly indicate a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM all score the substitution as benign. The consensus from the SGM framework, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also classifies the variant as likely benign. High‑accuracy predictors corroborate this assessment: AlphaMissense‑Optimized predicts a benign outcome, and the SGM‑Consensus (majority vote) confirms a likely benign status. No high‑accuracy folding‑stability analysis (Foldetta) is available for this residue, so stability effects remain unassessed. Overall, the collective evidence points to a benign impact for Q175P, and this conclusion is consistent with the absence of a pathogenic ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.653063 | Disordered | 0.474689 | Uncertain | 0.367 | 0.618 | 0.375 | 6-33435166-A-C | 1 | 6.20e-7 | -6.995 | Likely Benign | 0.432 | Ambiguous | Likely Benign | 0.211 | Likely Benign | 1.55 | Neutral | 0.396 | Benign | 0.188 | Benign | 4.20 | Benign | 1.00 | Tolerated | 3.61 | 5 | 0.2075 | 0.4584 | -1 | 0 | 1.9 | -31.01 | ||||||||||||||||||||||||||||||||||
| c.527G>T | S176I 2D AIThe SynGAP1 missense variant S176I is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments are inconclusive: the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a 2‑vs‑2 split, and Foldetta results are not available. Overall, the majority of evidence (six benign predictions versus two pathogenic) supports a benign classification. This conclusion does not contradict ClinVar, as the variant has no ClinVar entry. Thus, the variant is most likely benign based on current predictive data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.562014 | Disordered | 0.466016 | Uncertain | 0.380 | 0.597 | 0.375 | -10.247 | Likely Pathogenic | 0.903 | Likely Pathogenic | Ambiguous | 0.152 | Likely Benign | -2.03 | Neutral | 0.002 | Benign | 0.003 | Benign | 4.04 | Benign | 0.06 | Tolerated | 0.0790 | 0.5290 | -1 | -2 | 5.3 | 26.08 | ||||||||||||||||||||||||||||||||||||||||
| c.529T>G | F177V 2D AIThe SynGAP1 missense variant F177V is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 vs 2), and Foldetta results are unavailable. Overall, the majority of standard predictors favor a benign outcome, but the optimized AlphaMissense model and ESM1b suggest potential pathogenicity. Thus, the variant is most likely benign based on the collective evidence, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.505461 | Disordered | 0.461817 | Uncertain | 0.357 | 0.598 | 0.500 | -11.582 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | 0.188 | Likely Benign | -1.26 | Neutral | 0.028 | Benign | 0.009 | Benign | 4.18 | Benign | 0.06 | Tolerated | 0.2454 | 0.3526 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||||||
| c.55G>C | A19P 2D AIThe SynGAP1 missense variant A19P is reported in gnomAD (variant ID 6‑33420319‑G‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while only SIFT predicts it as pathogenic. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are not available. Overall, the consensus of the available predictions points to a benign impact, and this is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.443393 | Uncertain | 0.378 | 0.906 | 0.500 | 6-33420319-G-C | -3.579 | Likely Benign | 0.184 | Likely Benign | Likely Benign | 0.033 | Likely Benign | 0.09 | Neutral | 0.001 | Benign | 0.000 | Benign | 4.07 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2609 | 0.6018 | -1 | 1 | -3.4 | 26.04 | ||||||||||||||||||||||||||||||||||||
| c.563G>T | S188I 2D AIThe SynGAP1 missense variant S188I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect are REVEL and FATHMM, while the remaining tools—PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—predict a pathogenic or likely pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic and the SGM‑Consensus also pathogenic; Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for S188I, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.490133 | Structured | 0.428502 | Uncertain | 0.298 | 0.603 | 0.500 | -12.133 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | 0.205 | Likely Benign | -3.98 | Deleterious | 0.995 | Probably Damaging | 0.880 | Possibly Damaging | 3.90 | Benign | 0.00 | Affected | 0.0878 | 0.6335 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.569G>T | S190I 2D AIThe SynGAP1 missense variant S190I is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumVar, and FATHMM, whereas those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. AlphaMissense‑Optimized returns an Uncertain result. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is unavailable for this variant. Overall, the majority of evidence points to a pathogenic impact, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.422041 | Structured | 0.428613 | Uncertain | 0.338 | 0.615 | 0.250 | -9.868 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.316 | Likely Benign | -3.39 | Deleterious | 0.845 | Possibly Damaging | 0.368 | Benign | 4.03 | Benign | 0.03 | Affected | 0.0769 | 0.5963 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.572G>T | S191I 2D AIThe SynGAP1 missense variant S191I is not reported in ClinVar and has no entries in gnomAD, indicating it is not catalogued in these databases. Functional prediction tools show a split opinion: benign predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), and FATHMM, while pathogenic predictions arise from PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. When the predictions are grouped by consensus, four tools favor benign and four favor pathogenic. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized returns an uncertain result, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels the variant as Likely Pathogenic. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Taken together, the majority of evidence, including the SGM Consensus, points to a pathogenic effect, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.429200 | Structured | 0.428475 | Uncertain | 0.322 | 0.615 | 0.125 | -11.271 | Likely Pathogenic | 0.927 | Likely Pathogenic | Ambiguous | 0.283 | Likely Benign | -4.51 | Deleterious | 0.421 | Benign | 0.086 | Benign | 3.76 | Benign | 0.00 | Affected | 0.0954 | 0.6842 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||||||||||||
| c.58C>A | P20T 2D AIThe SynGAP1 missense variant P20T is reported in gnomAD (ID 6‑33420322‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates a likely benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT each predict pathogenic. High‑accuracy assessments reinforce the benign view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is likely benign. No Foldetta stability prediction is available, so it does not influence the assessment. Overall, the preponderance of evidence, including the high‑accuracy tools, points to the variant being most likely benign, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.442804 | Uncertain | 0.448 | 0.899 | 0.500 | 6-33420322-C-A | -3.258 | Likely Benign | 0.223 | Likely Benign | Likely Benign | 0.065 | Likely Benign | -0.50 | Neutral | 0.909 | Possibly Damaging | 0.641 | Possibly Damaging | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1781 | 0.6640 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||||||||
| c.58C>T | P20S 2D AIThe SynGAP1 missense variant P20S is reported in gnomAD (ID 6‑33420322‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from the four benign‑oriented tools). Pathogenic predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a benign effect for P20S, and this conclusion is not contradicted by any ClinVar classification (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.442804 | Uncertain | 0.448 | 0.899 | 0.500 | 6-33420322-C-T | -3.054 | Likely Benign | 0.153 | Likely Benign | Likely Benign | 0.076 | Likely Benign | -0.25 | Neutral | 0.909 | Possibly Damaging | 0.641 | Possibly Damaging | 4.30 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3732 | 0.5920 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||||
| c.614T>G | I205S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I205S missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, AlphaMissense‑Optimized, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and ESM1b. Tools with uncertain or inconclusive results (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of available predictions lean toward a benign impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.264545 | Structured | 0.409933 | Uncertain | 0.821 | 0.414 | 0.125 | -8.694 | Likely Pathogenic | 0.518 | Ambiguous | Likely Benign | 1.17 | Ambiguous | 0.2 | 0.95 | Ambiguous | 1.06 | Ambiguous | 0.88 | Ambiguous | 0.178 | Likely Benign | -2.73 | Deleterious | 0.838 | Possibly Damaging | 0.368 | Benign | 4.12 | Benign | 0.24 | Tolerated | 0.2346 | 0.0800 | -1 | -2 | -5.3 | -26.08 | ||||||||||||||||||||||||||||||
| c.617T>G | I206S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I206S missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FATHMM, and polyPhen‑2 HumVar. All other evaluated algorithms (SGM‑Consensus, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, is pathogenic. Based on the preponderance of evidence from these tools, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.298791 | Structured | 0.405123 | Uncertain | 0.863 | 0.391 | 0.125 | -13.711 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 3.73 | Destabilizing | 0.3 | 3.89 | Destabilizing | 3.81 | Destabilizing | 1.74 | Destabilizing | 0.251 | Likely Benign | -4.86 | Deleterious | 0.838 | Possibly Damaging | 0.368 | Benign | 3.63 | Benign | 0.00 | Affected | 0.2189 | 0.0728 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||
| c.61T>G | F21V 2D AIThe SynGAP1 missense variant F21V is listed in gnomAD (ID 6‑33420325‑T‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.534167 | Disordered | 0.443284 | Uncertain | 0.369 | 0.897 | 0.500 | 6-33420325-T-G | -2.823 | Likely Benign | 0.563 | Ambiguous | Likely Benign | 0.224 | Likely Benign | 0.64 | Neutral | 0.462 | Possibly Damaging | 0.307 | Benign | 4.44 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2536 | 0.2497 | -1 | -1 | 1.4 | -48.04 | ||||||||||||||||||||||||||||||||||||
| c.626T>G | V209G 2D AIThe SynGAP1 missense variant V209G is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools (FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a pathogenic classification; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a destabilizing, pathogenic effect. Taken together, the overwhelming majority of computational evidence indicates that V209G is likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.247041 | Structured | 0.397624 | Uncertain | 0.874 | 0.331 | 0.125 | -13.763 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 3.05 | Destabilizing | 0.5 | 3.99 | Destabilizing | 3.52 | Destabilizing | 1.27 | Destabilizing | 0.390 | Likely Benign | -5.49 | Deleterious | 0.829 | Possibly Damaging | 0.995 | Probably Damaging | 3.65 | Benign | 0.04 | Affected | 0.1600 | 0.2035 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.632G>T | S211I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 S211I missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, and FATHMM; pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, while Foldetta, which integrates FoldX‑MD and Rosetta outputs, is inconclusive. No evidence from FoldX or Rosetta is available. Overall, the preponderance of evidence indicates the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.209395 | Structured | 0.389893 | Uncertain | 0.846 | 0.300 | 0.125 | -17.090 | Likely Pathogenic | 0.973 | Likely Pathogenic | Likely Pathogenic | 1.06 | Ambiguous | 1.1 | 1.05 | Ambiguous | 1.06 | Ambiguous | 0.12 | Likely Benign | 0.251 | Likely Benign | -4.94 | Deleterious | 0.995 | Probably Damaging | 0.767 | Possibly Damaging | 3.90 | Benign | 0.01 | Affected | 0.0909 | 0.5220 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||
| c.638T>G | I213S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I213S missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while all other evaluated algorithms (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) classify the change as pathogenic. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. No prediction or stability result is missing or inconclusive. Based on the consensus of these tools, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.158265 | Structured | 0.372201 | Uncertain | 0.850 | 0.295 | 0.125 | -12.858 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 2.00 | Destabilizing | 1.1 | 2.95 | Destabilizing | 2.48 | Destabilizing | 1.53 | Destabilizing | 0.879 | Likely Pathogenic | -4.88 | Deleterious | 0.995 | Probably Damaging | 0.829 | Possibly Damaging | 5.83 | Benign | 0.00 | Affected | 0.2290 | 0.0800 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||
| c.644G>T | G215V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G215V is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while the remaining twelve tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) all classify the variant as pathogenic. premPS is uncertain and does not influence the consensus. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. No prediction or stability result is missing or inconclusive. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.155435 | Structured | 0.382818 | Uncertain | 0.791 | 0.291 | 0.000 | -12.960 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 3.69 | Destabilizing | 0.2 | 5.41 | Destabilizing | 4.55 | Destabilizing | 0.53 | Ambiguous | 0.884 | Likely Pathogenic | -7.66 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 5.56 | Benign | 0.00 | Affected | 0.1087 | 0.4777 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||
| c.652T>G | F218V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F218V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM; pathogenic predictions arise from REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is labeled “Likely Pathogenic.” High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus agrees on a likely pathogenic outcome, and Foldetta (combining FoldX‑MD and Rosetta outputs) also indicates pathogenicity. No predictions or stability results are missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion aligns with the absence of a ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.281712 | Structured | 0.408725 | Uncertain | 0.848 | 0.272 | 0.000 | -10.081 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 4.18 | Destabilizing | 0.2 | 6.35 | Destabilizing | 5.27 | Destabilizing | 1.15 | Destabilizing | 0.691 | Likely Pathogenic | -4.14 | Deleterious | 0.300 | Benign | 0.066 | Benign | 5.81 | Benign | 0.08 | Tolerated | 0.2173 | 0.2258 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.658T>G | F220V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F220V is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The remaining tools—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic effect. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is pathogenic. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.219301 | Structured | 0.429422 | Uncertain | 0.898 | 0.295 | 0.000 | -11.599 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 3.83 | Destabilizing | 0.1 | 4.11 | Destabilizing | 3.97 | Destabilizing | 1.56 | Destabilizing | 0.911 | Likely Pathogenic | -5.81 | Deleterious | 0.075 | Benign | 0.015 | Benign | 4.04 | Benign | 0.01 | Affected | 0.2351 | 0.3315 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.65G>C | R22T 2D AIThe SynGAP1 missense variant R22T is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools show a split: benign calls from REVEL, PROVEAN, polyPhen2_HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic calls come from polyPhen2_HumDiv and SIFT. The majority‑vote SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely benign effect. High‑accuracy predictors further support a benign interpretation: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus itself is benign; Foldetta predictions are not available. Given the preponderance of benign predictions and the lack of pathogenic evidence, the variant is most likely benign. This assessment aligns with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.441505 | Uncertain | 0.377 | 0.891 | 0.500 | -4.079 | Likely Benign | 0.510 | Ambiguous | Likely Benign | 0.146 | Likely Benign | -0.21 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.23 | Benign | 0.00 | Affected | 0.2140 | 0.5599 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||||
| c.665T>G | V222G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V222G resides in the PH domain. ClinVar has no entry for this variant, and it is absent from gnomAD. Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining eleven tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) all predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a pathogenic verdict; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. Thus, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict the ClinVar status, which currently contains no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.116183 | Structured | 0.402706 | Uncertain | 0.885 | 0.310 | 0.125 | -14.857 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 4.54 | Destabilizing | 0.4 | 5.25 | Destabilizing | 4.90 | Destabilizing | 2.05 | Destabilizing | 0.962 | Likely Pathogenic | -6.05 | Deleterious | 0.987 | Probably Damaging | 0.998 | Probably Damaging | 5.21 | Benign | 0.00 | Affected | 0.1678 | 0.1996 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.668C>G | T223R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T223R has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a mixed profile: benign calls come from FoldX, Rosetta, Foldetta, polyPhen‑2 HumVar, SIFT, and FATHMM, while pathogenic calls arise from REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. Two tools report uncertainty: premPS and AlphaMissense‑Optimized. High‑accuracy consensus (SGM Consensus) favors pathogenicity (3/4 votes pathogenic), whereas Foldetta predicts benign stability. Because the majority of individual predictors lean benign and the high‑accuracy consensus is split, the overall assessment remains inconclusive. The variant is most likely benign, but the presence of several pathogenic predictions and the SGM Consensus result indicates that pathogenicity cannot be ruled out. This conclusion does not contradict ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.070400 | Structured | 0.382605 | Uncertain | 0.867 | 0.316 | 0.125 | -12.079 | Likely Pathogenic | 0.794 | Likely Pathogenic | Ambiguous | -0.36 | Likely Benign | 0.1 | -0.27 | Likely Benign | -0.32 | Likely Benign | 0.75 | Ambiguous | 0.827 | Likely Pathogenic | -4.62 | Deleterious | 0.561 | Possibly Damaging | 0.178 | Benign | 5.73 | Benign | 0.06 | Tolerated | 0.0708 | 0.2407 | -1 | -1 | -3.8 | 55.08 | |||||||||||||||||||||||||||||
| c.66A>C | R22S 2D AIThe SynGAP1 missense variant R22S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include SGM‑Consensus (Likely Benign), REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also as benign; a Foldetta stability prediction is unavailable. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.441505 | Uncertain | 0.377 | 0.891 | 0.500 | -3.419 | Likely Benign | 0.602 | Likely Pathogenic | Likely Benign | 0.190 | Likely Benign | 0.01 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3252 | 0.5187 | -1 | 0 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||
| c.66A>T | R22S 2D AIThe SynGAP1 missense variant R22S is listed in gnomAD (ID 6‑33420330‑A‑T) but has no ClinVar entry. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant, so its stability impact is unavailable. Overall, the majority of evidence points to a benign effect, and this is not in conflict with ClinVar, which has no classification for R22S. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.441505 | Uncertain | 0.377 | 0.891 | 0.500 | 6-33420330-A-T | -3.419 | Likely Benign | 0.602 | Likely Pathogenic | Likely Benign | 0.190 | Likely Benign | 0.01 | Neutral | 0.462 | Possibly Damaging | 0.227 | Benign | 4.28 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3252 | 0.5187 | -1 | 0 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||||
| c.680G>T | G227V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G227V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while the remaining twelve tools—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The high‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. No prediction or stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.106997 | Structured | 0.329995 | Uncertain | 0.800 | 0.329 | 0.250 | -9.329 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 3.57 | Destabilizing | 0.4 | 6.24 | Destabilizing | 4.91 | Destabilizing | 0.78 | Ambiguous | 0.839 | Likely Pathogenic | -7.58 | Deleterious | 0.952 | Possibly Damaging | 0.521 | Possibly Damaging | 5.67 | Benign | 0.01 | Affected | 0.1130 | 0.4703 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||
| c.683C>G | T228R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T228R missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Foldetta, and FATHMM. Those that predict a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain predictions come from Rosetta, premPS, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a pathogenic impact. This conclusion is not contradicted by any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.104810 | Structured | 0.321733 | Uncertain | 0.829 | 0.316 | 0.125 | -7.218 | In-Between | 0.965 | Likely Pathogenic | Likely Pathogenic | -0.23 | Likely Benign | 0.1 | 0.69 | Ambiguous | 0.23 | Likely Benign | 0.67 | Ambiguous | 0.693 | Likely Pathogenic | -3.40 | Deleterious | 0.952 | Possibly Damaging | 0.694 | Possibly Damaging | 5.60 | Benign | 0.01 | Affected | 0.1108 | 0.3405 | -1 | -1 | -3.8 | 55.08 | ||||||||||||||||||||||||||||||
| c.691T>G | F231V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F231V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are polyPhen‑2 HumVar and FATHMM. All other evaluated algorithms—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely pathogenic status; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also indicates pathogenicity. Taken together, the evidence overwhelmingly points to a pathogenic effect for F231V, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.366687 | Structured | 0.306467 | Uncertain | 0.895 | 0.300 | 0.000 | -13.201 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 2.16 | Destabilizing | 0.3 | 2.30 | Destabilizing | 2.23 | Destabilizing | 1.13 | Destabilizing | 0.910 | Likely Pathogenic | -5.90 | Deleterious | 0.759 | Possibly Damaging | 0.328 | Benign | 5.72 | Benign | 0.00 | Affected | 0.2212 | 0.3030 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.6C>A | S2R 2D AIThe SynGAP1 missense variant S2R is present in gnomAD (ID 6‑33420270‑C‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus score all report benign or likely benign. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also favors benign. Foldetta, a protein‑folding stability predictor, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.543646 | Binding | 0.382 | 0.922 | 0.750 | 6-33420270-C-A | 1 | 6.52e-7 | -3.684 | Likely Benign | 0.426 | Ambiguous | Likely Benign | 0.070 | Likely Benign | -0.44 | Neutral | 0.117 | Benign | 0.008 | Benign | 4.05 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0996 | 0.4503 | -1 | 0 | -3.7 | 69.11 | ||||||||||||||||||||||||||||||||||
| c.6C>G | S2R 2D AIThe SynGAP1 missense variant S2R is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome, while AlphaMissense‑Default remains uncertain. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus also indicates Likely Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, did not provide a result for this variant, so its status is unavailable. Overall, the preponderance of evidence from multiple prediction tools and high‑accuracy methods indicates that the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.671169 | Disordered | 0.543646 | Binding | 0.382 | 0.922 | 0.750 | -3.684 | Likely Benign | 0.426 | Ambiguous | Likely Benign | 0.065 | Likely Benign | -0.44 | Neutral | 0.117 | Benign | 0.008 | Benign | 4.05 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0996 | 0.4503 | -1 | 0 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||||||||
| c.707C>A | A236E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A236E is not reported in ClinVar and is present in gnomAD (ID 6‑33435558‑C‑A). Functional prediction tools show a split assessment: benign calls come from FATHMM, Rosetta, and the protein‑folding stability method Foldetta; pathogenic calls come from REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score. When grouped by agreement, the benign‑predicting tools (FATHMM, Rosetta, Foldetta) represent one consensus, while the pathogenic‑predicting tools (REVEL, premPS, PROVEAN, polyPhen‑2, SIFT, ESM1b, AlphaMissense‑Default, SGM‑Consensus) form the opposing consensus. High‑accuracy methods give mixed results: AlphaMissense‑Optimized is uncertain; SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is likely pathogenic; Foldetta, combining FoldX‑MD and Rosetta outputs, is benign. Overall, the majority of predictions lean toward pathogenicity, and this does not contradict the ClinVar status, which has no reported classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.185198 | Structured | 0.329926 | Uncertain | 0.775 | 0.330 | 0.000 | 6-33435558-C-A | 1 | 6.20e-7 | -10.844 | Likely Pathogenic | 0.835 | Likely Pathogenic | Ambiguous | -0.75 | Ambiguous | 0.2 | 0.28 | Likely Benign | -0.24 | Likely Benign | 1.08 | Destabilizing | 0.844 | Likely Pathogenic | -4.24 | Deleterious | 0.998 | Probably Damaging | 0.900 | Possibly Damaging | 6.06 | Benign | 0.02 | Affected | 3.40 | 14 | 0.1075 | 0.1970 | -1 | 0 | -5.3 | 58.04 | ||||||||||||||||||||||||
| c.716G>C | R239T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R239T is recorded in gnomAD (ID 6‑33435567‑G‑C) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; pathogenic predictions arise from REVEL, Rosetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). FoldX reports an uncertain effect and is therefore not considered. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenicity. Consequently, the variant is most likely pathogenic according to the available computational evidence, and this assessment does not contradict any ClinVar classification because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.170161 | Structured | 0.336504 | Uncertain | 0.854 | 0.319 | 0.000 | 6-33435567-G-C | 1 | 6.19e-7 | -14.792 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 1.96 | Ambiguous | 0.3 | 2.44 | Destabilizing | 2.20 | Destabilizing | 1.21 | Destabilizing | 0.869 | Likely Pathogenic | -5.35 | Deleterious | 0.259 | Benign | 0.064 | Benign | 5.66 | Benign | 0.01 | Affected | 3.40 | 14 | 0.1805 | 0.4414 | -1 | -1 | 3.8 | -55.08 | ||||||||||||||||||||||||
| c.71T>G | V24G 2D AIThe SynGAP1 missense variant V24G is not reported in ClinVar (ClinVar status: not present) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is benign, and Foldetta results are unavailable. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.541878 | Disordered | 0.438970 | Uncertain | 0.382 | 0.890 | 0.500 | -2.673 | Likely Benign | 0.256 | Likely Benign | Likely Benign | 0.178 | Likely Benign | -1.67 | Neutral | 0.026 | Benign | 0.049 | Benign | 3.77 | Benign | 0.00 | Affected | 0.2081 | 0.3105 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||||||||||||
| c.728T>G | I243S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I243S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that assess pathogenicity largely agree: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a pathogenic effect. Only FATHMM predicts a benign outcome. High‑accuracy methods reinforce the pathogenic signal: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) is pathogenic. No predictions are missing or inconclusive. Based on the consensus of these tools, the variant is most likely pathogenic, and this assessment does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | PH | 0.363090 | Structured | 0.344471 | Uncertain | 0.842 | 0.347 | 0.000 | -14.097 | Likely Pathogenic | 0.975 | Likely Pathogenic | Likely Pathogenic | 2.52 | Destabilizing | 0.2 | 2.22 | Destabilizing | 2.37 | Destabilizing | 1.71 | Destabilizing | 0.802 | Likely Pathogenic | -3.55 | Deleterious | 0.995 | Probably Damaging | 0.795 | Possibly Damaging | 5.52 | Benign | 0.00 | Affected | 0.2658 | 0.0600 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||
| c.749T>G | V250G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V250G missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools largely agree on a deleterious effect: pathogenic calls come from REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (both HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default, while only FATHMM predicts a benign outcome. Uncertain results are reported by FoldX and AlphaMissense‑Optimized. High‑accuracy assessments reinforce the pathogenic signal: the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, AlphaMissense‑Optimized remains uncertain, and Foldetta is pathogenic. Taken together, the overwhelming majority of evidence indicates a pathogenic effect. This conclusion is consistent with the absence of a ClinVar classification; there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | PH | 0.447574 | Structured | 0.244075 | Uncertain | 0.778 | 0.324 | 0.125 | -11.255 | Likely Pathogenic | 0.917 | Likely Pathogenic | Ambiguous | 1.65 | Ambiguous | 0.3 | 3.18 | Destabilizing | 2.42 | Destabilizing | 2.08 | Destabilizing | 0.900 | Likely Pathogenic | -5.90 | Deleterious | 0.879 | Possibly Damaging | 0.997 | Probably Damaging | 5.75 | Benign | 0.00 | Affected | 0.1884 | 0.1996 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.770G>T | S257I 2D AIThe SynGAP1 missense variant S257I is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include premPS, SIFT, FATHMM, AlphaMissense‑Optimized, and Foldetta. Those that predict a pathogenic effect comprise SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicating likely pathogenic, and Foldetta predicting a benign outcome. FoldX and Rosetta results are uncertain and therefore not considered. Overall, the balance of evidence leans toward a pathogenic interpretation, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.384043 | Structured | 0.258293 | Uncertain | 0.847 | 0.272 | 0.250 | -12.126 | Likely Pathogenic | 0.595 | Likely Pathogenic | Likely Benign | 0.78 | Ambiguous | 1.0 | -1.17 | Ambiguous | -0.20 | Likely Benign | 0.30 | Likely Benign | 0.739 | Likely Pathogenic | -2.97 | Deleterious | 0.998 | Probably Damaging | 0.991 | Probably Damaging | 5.81 | Benign | 0.07 | Tolerated | 0.0681 | 0.5209 | -1 | -2 | 5.3 | 26.08 | |||||||||||||||||||||||||||||
| c.779T>G | V260G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V260G missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include FATHMM and AlphaMissense‑Optimized, while the majority of tools (REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) predict a pathogenic impact. FoldX, Rosetta, and Foldetta are uncertain and therefore not considered evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, whereas the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—leans pathogenic (3 pathogenic vs. 1 benign). Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for V260G, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.254060 | Structured | 0.382651 | Uncertain | 0.888 | 0.259 | 0.250 | -9.300 | Likely Pathogenic | 0.644 | Likely Pathogenic | Likely Benign | 1.00 | Ambiguous | 0.3 | 1.86 | Ambiguous | 1.43 | Ambiguous | 1.40 | Destabilizing | 0.817 | Likely Pathogenic | -4.20 | Deleterious | 0.991 | Probably Damaging | 0.999 | Probably Damaging | 5.76 | Benign | 0.00 | Affected | 0.1844 | 0.1949 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.788T>G | V263G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V263G missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are FATHMM and AlphaMissense‑Optimized; those that agree on a pathogenic effect are REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Rosetta and Foldetta give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic. Foldetta’s stability prediction is uncertain. Overall, the preponderance of evidence points to a pathogenic impact for V263G, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.268042 | Structured | 0.356141 | Uncertain | 0.918 | 0.257 | 0.000 | -10.388 | Likely Pathogenic | 0.669 | Likely Pathogenic | Likely Benign | 2.27 | Destabilizing | 0.2 | 1.63 | Ambiguous | 1.95 | Ambiguous | 1.88 | Destabilizing | 0.820 | Likely Pathogenic | -4.59 | Deleterious | 0.991 | Probably Damaging | 0.999 | Probably Damaging | 6.07 | Benign | 0.01 | Affected | 0.1790 | 0.1868 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.794A>C | K265T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K265T missense variant is reported in gnomAD (ID 6‑33437699‑A‑C) but has no ClinVar entry. Prediction tools that classify the variant as benign include REVEL, SIFT, and Rosetta. Those that predict pathogenicity are SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. Four tools give uncertain results: FoldX, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.209395 | Structured | 0.309758 | Uncertain | 0.936 | 0.275 | 0.000 | 6-33437699-A-C | 1 | 6.20e-7 | -9.425 | Likely Pathogenic | 0.839 | Likely Pathogenic | Ambiguous | 0.99 | Ambiguous | 0.1 | 0.37 | Likely Benign | 0.68 | Ambiguous | 0.83 | Ambiguous | 0.441 | Likely Benign | -3.75 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.91 | Pathogenic | 0.07 | Tolerated | 3.38 | 18 | 0.2042 | 0.3178 | -1 | 0 | 3.2 | -27.07 | ||||||||||||||||||||||||
| c.79C>T | P27S 2D AIThe SynGAP1 missense variant P27S is reported in gnomAD (ID 6‑33423488‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign, and the SGM‑Consensus (derived from the majority of these high‑accuracy predictors) also reports a likely benign outcome. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT predict a pathogenic impact. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign. Foldetta stability analysis is unavailable. Overall, the preponderance of evidence indicates that P27S is most likely benign, and this assessment does not contradict any ClinVar classification, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.517562 | Disordered | 0.437871 | Uncertain | 0.430 | 0.881 | 0.375 | 6-33423488-C-T | 1 | 6.20e-7 | -2.891 | Likely Benign | 0.098 | Likely Benign | Likely Benign | 0.063 | Likely Benign | -2.01 | Neutral | 0.909 | Possibly Damaging | 0.901 | Possibly Damaging | 3.91 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3916 | 0.5443 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||||||||||||
| c.803T>G | I268S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I268S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that assess pathogenicity all agree that the variant is deleterious: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify it as pathogenic. No tool predicts a benign effect. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, indicates a destabilizing, pathogenic effect. All available predictions are concordant and supportive. Based on these computational assessments, the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.216401 | Structured | 0.314336 | Uncertain | 0.951 | 0.264 | 0.000 | -13.032 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 4.93 | Destabilizing | 0.1 | 4.54 | Destabilizing | 4.74 | Destabilizing | 2.10 | Destabilizing | 0.841 | Likely Pathogenic | -5.34 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 1.53 | Pathogenic | 0.00 | Affected | 0.2037 | 0.0530 | -1 | -2 | -5.3 | -26.08 | |||||||||||||||||||||||||||||
| c.806T>C | I269T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I269T is not reported in ClinVar (no ClinVar entry) but is present in gnomAD (variant ID 6‑33437711‑T‑C). Among general in‑silico predictors, only SIFT classifies the change as benign, whereas the remaining tools that provide a definitive call (REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic effect. High‑accuracy assessments give a more nuanced view: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely pathogenic outcome; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, also reports a pathogenic effect. Based on the preponderance of pathogenic predictions and the high‑accuracy consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.216401 | Structured | 0.343787 | Uncertain | 0.937 | 0.244 | 0.125 | 6-33437711-T-C | 2 | 1.24e-6 | -9.376 | Likely Pathogenic | 0.887 | Likely Pathogenic | Ambiguous | 1.97 | Ambiguous | 0.1 | 2.10 | Destabilizing | 2.04 | Destabilizing | 1.38 | Destabilizing | 0.727 | Likely Pathogenic | -3.70 | Deleterious | 0.997 | Probably Damaging | 0.994 | Probably Damaging | 1.72 | Pathogenic | 0.09 | Tolerated | 3.38 | 19 | 0.0833 | 0.0808 | -1 | 0 | -5.2 | -12.05 | ||||||||||||||||||||||||
| c.823C>A | P275T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant P275T is reported in gnomAD (ID 6‑33437728‑C‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, AlphaMissense‑Default, and AlphaMissense‑Optimized; pathogenic predictions arise from FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, whereas the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain and therefore not considered evidence. No other tools provide conclusive results. Overall, the majority of predictions, including the SGM‑Consensus, indicate a pathogenic effect, and this assessment does not contradict any ClinVar classification because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.059222 | Structured | 0.353469 | Uncertain | 0.811 | 0.208 | 0.250 | 6-33437728-C-A | 3 | 1.86e-6 | -8.708 | Likely Pathogenic | 0.309 | Likely Benign | Likely Benign | 2.44 | Destabilizing | 0.3 | 1.15 | Ambiguous | 1.80 | Ambiguous | 0.69 | Ambiguous | 0.425 | Likely Benign | -5.38 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.75 | Pathogenic | 0.01 | Affected | 3.38 | 19 | 0.1808 | 0.4000 | -1 | 0 | 0.9 | 3.99 | ||||||||||||||||||||||||
| c.823C>G | P275A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P275A is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33437728‑C‑G). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments are less decisive: AlphaMissense‑Optimized reports a benign outcome, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic), and Foldetta is uncertain. Consequently, the overall evidence leans toward a benign interpretation, with no ClinVar record to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.059222 | Structured | 0.353469 | Uncertain | 0.811 | 0.208 | 0.250 | 6-33437728-C-G | 1 | 6.20e-7 | -6.137 | Likely Benign | 0.133 | Likely Benign | Likely Benign | 1.87 | Ambiguous | 0.2 | 1.11 | Ambiguous | 1.49 | Ambiguous | 0.50 | Likely Benign | 0.410 | Likely Benign | -4.95 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.79 | Pathogenic | 0.32 | Tolerated | 3.38 | 19 | 0.3475 | 0.3243 | -1 | 1 | 3.4 | -26.04 | |||||||||||||||||||||||||
| c.823C>T | P275S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P275S is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33437728‑C‑T). Prediction tools that agree on a benign effect include REVEL, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The remaining tools—Rosetta, Foldetta, premPS, and ESM1b—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact for P275S, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.059222 | Structured | 0.353469 | Uncertain | 0.811 | 0.208 | 0.250 | 6-33437728-C-T | 1 | 6.20e-7 | -7.886 | In-Between | 0.312 | Likely Benign | Likely Benign | 2.11 | Destabilizing | 0.3 | 1.28 | Ambiguous | 1.70 | Ambiguous | 0.77 | Ambiguous | 0.388 | Likely Benign | -5.24 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.78 | Pathogenic | 0.03 | Affected | 3.38 | 19 | 0.3489 | 0.3339 | -1 | 1 | 0.8 | -10.04 | |||||||||||||||||||||||||
| c.826C>G | P276A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P276A is reported in gnomAD (variant ID 6‑33437731‑C‑G) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The high‑accuracy consensus methods give a benign signal: AlphaMissense‑Optimized is benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign,” while Foldetta (combining FoldX‑MD and Rosetta outputs) is inconclusive. Overall, the majority of evidence points to a benign effect, and this assessment does not contradict any ClinVar status (none is available). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.037156 | Structured | 0.338937 | Uncertain | 0.724 | 0.230 | 0.250 | 6-33437731-C-G | 5 | 3.10e-6 | -3.414 | Likely Benign | 0.058 | Likely Benign | Likely Benign | 1.42 | Ambiguous | 0.1 | 1.01 | Ambiguous | 1.22 | Ambiguous | 0.50 | Likely Benign | 0.187 | Likely Benign | -2.31 | Neutral | 0.044 | Benign | 0.030 | Benign | 1.98 | Pathogenic | 0.57 | Tolerated | 3.38 | 19 | 0.3149 | 0.3669 | -1 | 1 | 3.4 | -26.04 | ||||||||||||||||||||||||
| c.848A>C | E283A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E283A is listed in ClinVar with an “Uncertain” status and is present in the gnomAD database (ID 6‑33437753‑A‑C). Prediction tools that assess pathogenicity uniformly favor a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report pathogenic or likely pathogenic. No tool in the dataset predicts a benign outcome; the remaining methods (FoldX, Rosetta, premPS, Foldetta) are inconclusive. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Pathogenic,” while Foldetta’s stability prediction is uncertain. Overall, the evidence strongly indicates that E283A is most likely pathogenic, which is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.098513 | Structured | 0.358602 | Uncertain | 0.950 | 0.249 | 0.000 | Uncertain | 1 | 6-33437753-A-C | 2 | 1.24e-6 | -12.547 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 1.26 | Ambiguous | 0.1 | 1.19 | Ambiguous | 1.23 | Ambiguous | 0.53 | Ambiguous | 0.529 | Likely Pathogenic | -5.52 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.67 | Pathogenic | 0.01 | Affected | 3.38 | 19 | 0.4104 | 0.5807 | -1 | 0 | 5.3 | -58.04 | ||||||||||||||||||||||
| c.866T>C | M289T 2D AIThe SynGAP1 missense variant M289T is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that classify the variant as benign include REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized scores the variant as benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is labeled likely benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts a benign effect. Taken together, the majority of evidence indicates that M289T is most likely benign, and this conclusion does not contradict the current ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.127496 | Structured | 0.403499 | Uncertain | 0.886 | 0.276 | 0.000 | Uncertain | 1 | -4.668 | Likely Benign | 0.238 | Likely Benign | Likely Benign | 0.73 | Ambiguous | 0.1 | 0.17 | Likely Benign | 0.45 | Likely Benign | -0.01 | Likely Benign | 0.222 | Likely Benign | -0.47 | Neutral | 0.801 | Possibly Damaging | 0.315 | Benign | 1.83 | Pathogenic | 0.57 | Tolerated | 0.1657 | 0.1534 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||
| c.86T>C | M29T 2D AIThe SynGAP1 missense variant M29T is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that M29T is most likely benign, and this conclusion does not contradict the current ClinVar “Uncertain” designation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.541878 | Disordered | 0.438540 | Uncertain | 0.341 | 0.883 | 0.250 | Uncertain | 1 | -2.167 | Likely Benign | 0.122 | Likely Benign | Likely Benign | 0.199 | Likely Benign | -0.37 | Neutral | 0.018 | Benign | 0.184 | Benign | 4.33 | Benign | 0.00 | Affected | 4.32 | 1 | 0.2633 | 0.2716 | -1 | -1 | -2.6 | -30.09 | |||||||||||||||||||||||||||||||||||
| c.884C>T | T295I 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant T295I is reported in gnomAD (ID 6‑33437789‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two consensus groups: benign predictions come from FoldX and Foldetta, while pathogenic predictions are supported by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain results are reported by AlphaMissense‑Optimized, Rosetta, and premPS and are treated as inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (integrating FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points toward a pathogenic effect, with only a minority of tools indicating benign or uncertain outcomes. This prediction does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.401658 | Structured | 0.295548 | Uncertain | 0.881 | 0.288 | 0.125 | 6-33437789-C-T | 4 | 2.48e-6 | -9.330 | Likely Pathogenic | 0.892 | Likely Pathogenic | Ambiguous | 0.21 | Likely Benign | 0.2 | 0.55 | Ambiguous | 0.38 | Likely Benign | 0.58 | Ambiguous | 0.607 | Likely Pathogenic | -4.87 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.88 | Pathogenic | 0.04 | Affected | 3.38 | 23 | 0.1025 | 0.5599 | -1 | 0 | 5.2 | 12.05 | ||||||||||||||||||||||||
| c.892C>T | P298S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P298S is listed in ClinVar as Benign (ClinVar ID 2965590.0) and is present in gnomAD (ID 6‑33437797‑C‑T). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM‑Consensus as Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Uncertain. No evidence from FoldX, Rosetta, or premPS is available to support either outcome. Overall, the majority of predictions support a benign impact, aligning with the ClinVar designation. Thus, the variant is most likely benign and does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.328603 | Structured | 0.268765 | Uncertain | 0.860 | 0.283 | 0.500 | Benign | 1 | 6-33437797-C-T | 5 | 3.10e-6 | -6.342 | Likely Benign | 0.144 | Likely Benign | Likely Benign | 1.38 | Ambiguous | 0.2 | 1.41 | Ambiguous | 1.40 | Ambiguous | 0.58 | Ambiguous | 0.189 | Likely Benign | -1.20 | Neutral | 0.991 | Probably Damaging | 0.898 | Possibly Damaging | 2.03 | Pathogenic | 0.85 | Tolerated | 3.39 | 20 | 0.3678 | 0.5855 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||
| c.899C>G | S300C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S300C is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33437804‑C‑G). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. Rosetta and Foldetta give uncertain results and are therefore not considered evidence for either side. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, SGM‑Consensus as Likely Benign, and Foldetta as Uncertain. Overall, the majority of reliable predictions indicate a benign impact, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.356642 | Structured | 0.256848 | Uncertain | 0.742 | 0.280 | 0.375 | 6-33437804-C-G | -6.749 | Likely Benign | 0.108 | Likely Benign | Likely Benign | 0.31 | Likely Benign | 0.2 | 1.45 | Ambiguous | 0.88 | Ambiguous | 0.34 | Likely Benign | 0.129 | Likely Benign | -2.45 | Neutral | 0.975 | Probably Damaging | 0.815 | Possibly Damaging | 1.55 | Pathogenic | 0.01 | Affected | 3.47 | 19 | 0.1005 | 0.6493 | -1 | 0 | 3.3 | 16.06 | ||||||||||||||||||||||||||
| c.8G>C | R3T 2D AIThe SynGAP1 missense variant R3T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.550331 | Binding | 0.358 | 0.920 | 0.875 | -3.693 | Likely Benign | 0.285 | Likely Benign | Likely Benign | 0.053 | Likely Benign | -0.55 | Neutral | 0.208 | Benign | 0.018 | Benign | 4.01 | Benign | 0.00 | Affected | 0.1809 | 0.5145 | -1 | -1 | 3.8 | -55.08 | |||||||||||||||||||||||||||||||||||||||
| c.908G>T | G303V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G303V is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in silico tools cluster around a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign) all indicate a tolerated change. Pathogenic signals arise only from SIFT and FoldX, while Rosetta and Foldetta are inconclusive. High‑accuracy assessments reinforce the benign trend: AlphaMissense‑Optimized predicts benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields Likely Benign, and Foldetta remains uncertain. Overall, the preponderance of evidence points to a benign effect, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.450668 | Structured | 0.271087 | Uncertain | 0.630 | 0.254 | 0.250 | -3.046 | Likely Benign | 0.198 | Likely Benign | Likely Benign | 3.06 | Destabilizing | 0.6 | -0.72 | Ambiguous | 1.17 | Ambiguous | 0.14 | Likely Benign | 0.071 | Likely Benign | -1.57 | Neutral | 0.011 | Benign | 0.017 | Benign | 3.96 | Benign | 0.01 | Affected | 0.1002 | 0.4577 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||
| c.914C>T | T305I 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant T305I is listed in ClinVar with an uncertain significance and is present in gnomAD (ID 6‑33437819‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, FoldX, premPS, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Foldetta. Tools that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split. Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports a benign effect. Overall, the balance of evidence from high‑confidence predictors leans toward a benign impact, and this assessment does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.359901 | Structured | 0.299706 | Uncertain | 0.872 | 0.274 | 0.125 | Uncertain | 1 | 6-33437819-C-T | 1 | 6.20e-7 | -5.222 | Likely Benign | 0.305 | Likely Benign | Likely Benign | 0.37 | Likely Benign | 0.2 | 0.58 | Ambiguous | 0.48 | Likely Benign | 0.25 | Likely Benign | 0.224 | Likely Benign | -2.90 | Deleterious | 0.997 | Probably Damaging | 0.929 | Probably Damaging | 1.70 | Pathogenic | 0.04 | Affected | 3.40 | 20 | 0.0939 | 0.5693 | -1 | 0 | 5.2 | 12.05 | |||||||||||||||||||||||
| c.916G>T | V306F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V306F is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL, premPS, and AlphaMissense‑Optimized, whereas the remaining tools (SGM‑Consensus, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) labeling it likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicting a pathogenic effect. Overall, the majority of evidence supports a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.363090 | Structured | 0.315026 | Uncertain | 0.896 | 0.287 | 0.125 | -9.311 | Likely Pathogenic | 0.700 | Likely Pathogenic | Likely Benign | 13.59 | Destabilizing | 1.4 | 6.99 | Destabilizing | 10.29 | Destabilizing | 0.23 | Likely Benign | 0.336 | Likely Benign | -3.28 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.74 | Pathogenic | 0.01 | Affected | 0.0535 | 0.3318 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.917T>G | V306G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V306G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that assess pathogenicity unanimously classify the variant as pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. No tool predicts a benign effect. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. All available evidence points to a pathogenic effect. Therefore, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.315026 | Uncertain | 0.896 | 0.287 | 0.125 | -12.313 | Likely Pathogenic | 0.795 | Likely Pathogenic | Ambiguous | 4.39 | Destabilizing | 0.2 | 5.89 | Destabilizing | 5.14 | Destabilizing | 2.46 | Destabilizing | 0.529 | Likely Pathogenic | -5.48 | Deleterious | 0.998 | Probably Damaging | 1.000 | Probably Damaging | 1.78 | Pathogenic | 0.00 | Affected | 0.1874 | 0.2035 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.919T>G | F307V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F307V is not reported in ClinVar and has no entries in gnomAD. Prediction tools that indicate a benign effect include Rosetta and premPS, whereas the majority of other in silico predictors (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) and the consensus score SGM‑Consensus all classify the change as pathogenic or likely pathogenic. High‑accuracy methods give a pathogenic call from AlphaMissense‑Optimized, a likely pathogenic result from the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and an inconclusive result from Foldetta (combining FoldX‑MD and Rosetta). No prediction is missing or inconclusive enough to alter the overall assessment. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.298791 | Structured | 0.327302 | Uncertain | 0.900 | 0.315 | 0.125 | -12.262 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.91 | Ambiguous | 0.1 | 0.10 | Likely Benign | 0.51 | Ambiguous | 0.36 | Likely Benign | 0.605 | Likely Pathogenic | -6.43 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 2.29 | Pathogenic | 0.05 | Affected | 0.2523 | 0.3393 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.926G>T | G309V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G309V is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while premPS remains uncertain. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports pathogenic. Consequently, the variant is most likely pathogenic, and this prediction is consistent with the absence of a ClinVar entry (no contradiction). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.275179 | Structured | 0.338439 | Uncertain | 0.882 | 0.342 | 0.125 | -13.595 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 4.55 | Destabilizing | 0.5 | 3.61 | Destabilizing | 4.08 | Destabilizing | 0.76 | Ambiguous | 0.531 | Likely Pathogenic | -8.27 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.67 | Pathogenic | 0.00 | Affected | 0.1256 | 0.3910 | -1 | -3 | 4.6 | 42.08 | |||||||||||||||||||||||||||||
| c.934T>G | F312V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F312V is not reported in ClinVar and is absent from gnomAD. All available in‑silico predictors classify the change as pathogenic: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts pathogenic. Based on the unanimous computational evidence, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.225814 | Structured | 0.361163 | Uncertain | 0.915 | 0.317 | 0.000 | -12.206 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 2.13 | Destabilizing | 0.1 | 3.46 | Destabilizing | 2.80 | Destabilizing | 1.80 | Destabilizing | 0.877 | Likely Pathogenic | -6.43 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 1.25 | Pathogenic | 0.00 | Affected | 0.1998 | 0.2810 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.940T>G | F314V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F314V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). All available in silico predictors classify the variant as pathogenic: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect, so the benign group is empty. High‑accuracy methods—AlphaMissense‑Optimized, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta—each return a pathogenic prediction. No predictions or stability results are missing or inconclusive. Based on the unanimous pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.173081 | Structured | 0.374049 | Uncertain | 0.900 | 0.271 | 0.125 | -8.907 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 3.31 | Destabilizing | 0.4 | 2.54 | Destabilizing | 2.93 | Destabilizing | 1.44 | Destabilizing | 0.594 | Likely Pathogenic | -5.90 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 1.26 | Pathogenic | 0.00 | Affected | 0.2054 | 0.2075 | -1 | -1 | 1.4 | -48.04 | |||||||||||||||||||||||||||||
| c.952C>T | P318S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P318S is present in gnomAD (variant ID 6‑33437857‑C‑T) but has no ClinVar entry. Functional prediction tools uniformly indicate a deleterious effect. Pathogenic predictions come from SGM‑Consensus, REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain predictions come from Rosetta and Foldetta. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic, SGM‑Consensus is likely pathogenic, and Foldetta remains uncertain. Taken together, the overwhelming majority of evidence supports a pathogenic classification, and this conclusion is consistent with the absence of a ClinVar record rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.111485 | Structured | 0.400936 | Uncertain | 0.858 | 0.234 | 0.000 | 6-33437857-C-T | 1 | 6.19e-7 | -9.954 | Likely Pathogenic | 0.956 | Likely Pathogenic | Likely Pathogenic | 2.22 | Destabilizing | 0.1 | 1.71 | Ambiguous | 1.97 | Ambiguous | 1.00 | Destabilizing | 0.626 | Likely Pathogenic | -7.05 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.87 | Pathogenic | 0.03 | Affected | 3.38 | 23 | 0.3692 | 0.5653 | -1 | 1 | 0.8 | -10.04 | ||||||||||||||||||||||||
| c.955G>C | A319P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A319P is catalogued in gnomAD (ID 6‑33437860‑G‑C) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Predictions that are inconclusive are Rosetta, ESM1b, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign effect, and this is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.179055 | Structured | 0.410405 | Uncertain | 0.879 | 0.254 | 0.125 | 6-33437860-G-C | 3 | 1.86e-6 | -7.213 | In-Between | 0.109 | Likely Benign | Likely Benign | -0.02 | Likely Benign | 0.9 | -1.18 | Ambiguous | -0.60 | Ambiguous | 0.22 | Likely Benign | 0.286 | Likely Benign | 0.11 | Neutral | 0.999 | Probably Damaging | 0.977 | Probably Damaging | 1.97 | Pathogenic | 1.00 | Tolerated | 3.38 | 23 | 0.1839 | 0.5067 | -1 | 1 | -3.4 | 26.04 | |||||||||||||||||||||||||
| c.958G>T | V320F 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant V320F is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on benign impact include REVEL, premPS, and SIFT, whereas tools that agree on pathogenic impact include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as inconclusive, SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as inconclusive. Because the majority of available predictions and the SGM‑Consensus favor pathogenicity, the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for V320F. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.185198 | Structured | 0.419626 | Uncertain | 0.905 | 0.266 | 0.125 | -9.958 | Likely Pathogenic | 0.877 | Likely Pathogenic | Ambiguous | 1.49 | Ambiguous | 1.4 | 1.55 | Ambiguous | 1.52 | Ambiguous | 0.44 | Likely Benign | 0.237 | Likely Benign | -3.26 | Deleterious | 0.994 | Probably Damaging | 0.944 | Probably Damaging | 1.79 | Pathogenic | 0.06 | Tolerated | 0.0473 | 0.3064 | -1 | -1 | -1.4 | 48.04 | |||||||||||||||||||||||||||||
| c.959T>G | V320G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V320G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools cluster into two agreement groups: the single benign prediction comes from REVEL, while the pathogenic group includes FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Two tools give uncertain results: Rosetta and AlphaMissense‑Optimized. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is uncertain; the SGM Consensus—derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—returns pathogenic; and Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, also predicts pathogenic. Overall, the preponderance of evidence indicates that V320G is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.185198 | Structured | 0.419626 | Uncertain | 0.905 | 0.266 | 0.125 | -9.043 | Likely Pathogenic | 0.816 | Likely Pathogenic | Ambiguous | 2.15 | Destabilizing | 1.1 | 1.87 | Ambiguous | 2.01 | Destabilizing | 1.48 | Destabilizing | 0.438 | Likely Benign | -5.74 | Deleterious | 0.958 | Probably Damaging | 0.999 | Probably Damaging | 1.89 | Pathogenic | 0.02 | Affected | 0.1688 | 0.1949 | -1 | -3 | -4.6 | -42.08 | ||||||||||||||||||||||||||||||
| c.95C>T | T32I 2D AIThe SynGAP1 missense variant T32I is reported in gnomAD (ID 6‑33423504‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also reports likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign impact for T32I, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.707965 | Disordered | 0.437154 | Uncertain | 0.349 | 0.879 | 0.375 | 6-33423504-C-T | 1 | 6.20e-7 | -3.689 | Likely Benign | 0.213 | Likely Benign | Likely Benign | 0.024 | Likely Benign | -0.57 | Neutral | 0.049 | Benign | 0.026 | Benign | 4.26 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1081 | 0.6403 | -1 | 0 | 5.2 | 12.05 | ||||||||||||||||||||||||||||||||||
| c.9G>C | R3S 2D AIThe SynGAP1 missense variant R3S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta results are not available for this variant. Overall, the majority of evidence points to a benign effect. The prediction is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.550331 | Binding | 0.358 | 0.920 | 0.875 | -2.296 | Likely Benign | 0.310 | Likely Benign | Likely Benign | 0.111 | Likely Benign | -0.53 | Neutral | 0.115 | Benign | 0.013 | Benign | 4.02 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3148 | 0.4732 | -1 | 0 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||
| c.9G>T | R3S 2D AIThe SynGAP1 missense variant R3S is reported in gnomAD (ID 6‑33420273‑G‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus itself is benign; Foldetta results are not available. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign effect. This conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.538167 | Disordered | 0.550331 | Binding | 0.358 | 0.920 | 0.875 | 6-33420273-G-T | 1 | 6.50e-7 | -2.296 | Likely Benign | 0.310 | Likely Benign | Likely Benign | 0.111 | Likely Benign | -0.53 | Neutral | 0.115 | Benign | 0.013 | Benign | 4.02 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3148 | 0.4732 | -1 | 0 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||||||
| c.1000A>G | K334E 2D 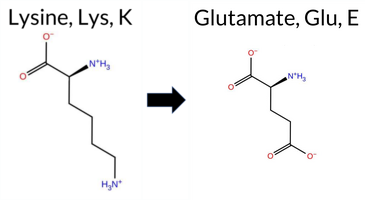 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K334E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, and premPS. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Pathogenic, SGM‑Consensus as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Benign. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.377384 | Structured | 0.325972 | Uncertain | 0.544 | 0.414 | 0.500 | -12.770 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.30 | Likely Benign | 0.2 | -0.07 | Likely Benign | 0.12 | Likely Benign | 0.42 | Likely Benign | 0.372 | Likely Benign | -3.67 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 1.74 | Pathogenic | 0.02 | Affected | 0.3929 | 0.0882 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1001A>C | K334T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K334T is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, and premPS. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Predictions that are inconclusive or uncertain are FoldX, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show that the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts a likely pathogenic outcome, AlphaMissense‑Optimized is uncertain, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also uncertain. Overall, the majority of evidence points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.377384 | Structured | 0.325972 | Uncertain | 0.544 | 0.414 | 0.500 | -8.313 | Likely Pathogenic | 0.943 | Likely Pathogenic | Ambiguous | 0.78 | Ambiguous | 0.3 | 0.21 | Likely Benign | 0.50 | Ambiguous | 0.17 | Likely Benign | 0.320 | Likely Benign | -5.51 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.78 | Pathogenic | 0.02 | Affected | 0.2062 | 0.2978 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1001A>T | K334M 2D 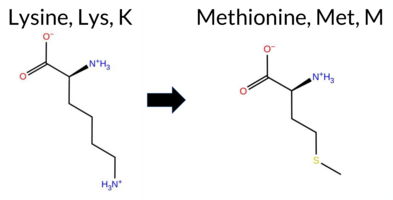 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K334M is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include REVEL, FoldX, and premPS. In contrast, the majority of tools predict a pathogenic outcome: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as deleterious. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports the variant as Likely Pathogenic. Foldetta and Rosetta provide uncertain results. Focusing on high‑accuracy methods, AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus confirms a Likely Pathogenic status, and Foldetta remains inconclusive. Overall, the preponderance of evidence points to a pathogenic effect for K334M, and this assessment does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.377384 | Structured | 0.325972 | Uncertain | 0.544 | 0.414 | 0.500 | -10.530 | Likely Pathogenic | 0.973 | Likely Pathogenic | Likely Pathogenic | 0.44 | Likely Benign | 0.0 | 0.56 | Ambiguous | 0.50 | Ambiguous | 0.14 | Likely Benign | 0.323 | Likely Benign | -5.51 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.77 | Pathogenic | 0.01 | Affected | 0.1027 | 0.3690 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1003C>A | R335S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R335S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from REVEL, premPS, and SIFT; pathogenic predictions from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is labeled Likely Pathogenic, while Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is inconclusive. No evidence from FoldX or Rosetta alone is available. Based on the preponderance of pathogenic predictions and the high‑accuracy tools, the variant is most likely pathogenic, which is consistent with the absence of ClinVar reporting and gnomAD data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.305330 | Structured | 0.331028 | Uncertain | 0.483 | 0.428 | 0.500 | -9.286 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 0.68 | Ambiguous | 0.1 | 0.72 | Ambiguous | 0.70 | Ambiguous | 0.14 | Likely Benign | 0.184 | Likely Benign | -3.30 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.89 | Pathogenic | 0.11 | Tolerated | 0.2598 | 0.4005 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||
| c.1004G>C | R335P 2D 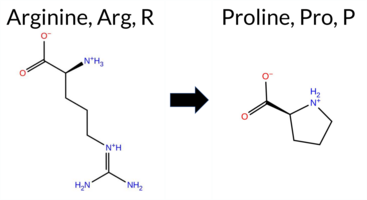 3DClick to see structure in 3D Viewer AISynGAP1 missense variant R335P has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, and SIFT, whereas pathogenic predictions are reported by FoldX, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments give a more focused view: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts a pathogenic effect. Taken together, the majority of evidence points to a pathogenic impact for R335P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.305330 | Structured | 0.331028 | Uncertain | 0.483 | 0.428 | 0.500 | -13.952 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 3.23 | Destabilizing | 0.8 | 5.73 | Destabilizing | 4.48 | Destabilizing | 0.43 | Likely Benign | 0.250 | Likely Benign | -4.22 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.69 | Pathogenic | 0.09 | Tolerated | 0.1866 | 0.4497 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1006A>G | K336E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K336E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, and polyPhen‑2 HumVar. Tools that predict a pathogenic effect comprise SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points toward a pathogenic impact for K336E, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.318242 | Structured | 0.338219 | Uncertain | 0.396 | 0.428 | 0.500 | -16.091 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | -0.28 | Likely Benign | 0.0 | 0.19 | Likely Benign | -0.05 | Likely Benign | 0.35 | Likely Benign | 0.236 | Likely Benign | -3.43 | Deleterious | 0.625 | Possibly Damaging | 0.192 | Benign | 1.60 | Pathogenic | 0.01 | Affected | 0.4003 | 0.1082 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1007A>C | K336T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K336T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, and polyPhen‑2 HumVar. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy assessment shows AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points toward a pathogenic impact for K336T, and this conclusion does not contradict any ClinVar annotation, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.318242 | Structured | 0.338219 | Uncertain | 0.396 | 0.428 | 0.500 | -13.468 | Likely Pathogenic | 0.971 | Likely Pathogenic | Likely Pathogenic | 0.33 | Likely Benign | 0.1 | -0.08 | Likely Benign | 0.13 | Likely Benign | 0.15 | Likely Benign | 0.212 | Likely Benign | -4.94 | Deleterious | 0.891 | Possibly Damaging | 0.315 | Benign | 1.58 | Pathogenic | 0.01 | Affected | 0.2014 | 0.4048 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1007A>T | K336M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K336M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions (REVEL, FoldX, premPS) and pathogenic predictions (SIFT, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized). The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is labeled “Likely Pathogenic.” Stability‑based assessments are inconclusive: Foldetta is uncertain, and Rosetta is also uncertain. High‑accuracy tools specifically indicate pathogenicity: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus confirms pathogenic, while Foldetta remains uncertain. Based on the overall pattern of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.318242 | Structured | 0.338219 | Uncertain | 0.396 | 0.428 | 0.500 | -15.395 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 0.34 | Likely Benign | 0.1 | 0.82 | Ambiguous | 0.58 | Ambiguous | -0.23 | Likely Benign | 0.301 | Likely Benign | -5.07 | Deleterious | 0.989 | Probably Damaging | 0.832 | Possibly Damaging | 1.53 | Pathogenic | 0.00 | Affected | 0.1185 | 0.4760 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1009A>G | K337E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K337E missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, and Foldetta. Those that predict a pathogenic impact comprise PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). The premPS score is uncertain and does not influence the overall assessment. High‑accuracy analyses show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as Likely Pathogenic, and Foldetta as benign. Based on the majority of predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.321458 | Structured | 0.348540 | Uncertain | 0.449 | 0.438 | 0.500 | -13.673 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | -0.05 | Likely Benign | 0.1 | 0.49 | Likely Benign | 0.22 | Likely Benign | 0.52 | Ambiguous | 0.316 | Likely Benign | -3.48 | Deleterious | 0.997 | Probably Damaging | 0.992 | Probably Damaging | 1.76 | Pathogenic | 0.02 | Affected | 0.3102 | 0.1039 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.100T>C | Y34H 2D 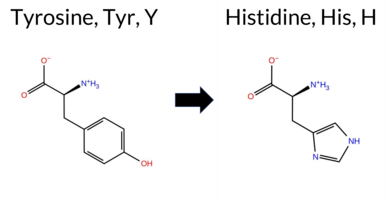 AIThe SynGAP1 missense variant Y34H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for Y34H, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.435847 | Uncertain | 0.303 | 0.855 | 0.375 | -2.929 | Likely Benign | 0.315 | Likely Benign | Likely Benign | 0.102 | Likely Benign | -0.53 | Neutral | 0.824 | Possibly Damaging | 0.775 | Possibly Damaging | 4.15 | Benign | 0.00 | Affected | 0.2630 | 0.1062 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||||||||||||
| c.1010A>C | K337T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K337T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two consensus groups: benign predictions come from REVEL, FoldX, and premPS, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Three tools report uncertainty: Rosetta, Foldetta, and AlphaMissense‑Optimized. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Pathogenic. In the high‑accuracy subset, AlphaMissense‑Optimized remains uncertain, SGM‑Consensus is Likely Pathogenic, and Foldetta is uncertain. Taken together, the majority of evidence points toward a deleterious effect. Therefore, K337T is most likely pathogenic, and this assessment does not conflict with the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.321458 | Structured | 0.348540 | Uncertain | 0.449 | 0.438 | 0.500 | -10.896 | Likely Pathogenic | 0.953 | Likely Pathogenic | Ambiguous | 0.45 | Likely Benign | 0.2 | 1.33 | Ambiguous | 0.89 | Ambiguous | 0.25 | Likely Benign | 0.338 | Likely Benign | -5.32 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.70 | Pathogenic | 0.01 | Affected | 0.1741 | 0.3354 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1010A>T | K337M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K337M missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that classify it as benign include REVEL, FoldX, premPS, and the protein‑folding stability method Foldetta. In contrast, the majority of in‑silico predictors flag it as pathogenic: SIFT, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Pathogenic” verdict. For high‑accuracy assessment, AlphaMissense‑Optimized remains pathogenic, the SGM‑Consensus also indicates likely pathogenic, whereas Foldetta predicts benign stability. No prediction is inconclusive; Rosetta is uncertain but not counted as evidence. Overall, the preponderance of evidence points to a pathogenic effect, and this assessment does not contradict any ClinVar classification because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.321458 | Structured | 0.348540 | Uncertain | 0.449 | 0.438 | 0.500 | -13.406 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 0.28 | Likely Benign | 0.1 | 0.61 | Ambiguous | 0.45 | Likely Benign | -0.24 | Likely Benign | 0.345 | Likely Benign | -5.32 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.66 | Pathogenic | 0.00 | Affected | 0.0862 | 0.3871 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1013A>C | D338A 2D 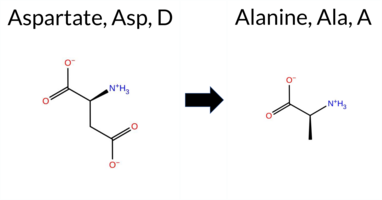 3DClick to see structure in 3D Viewer AISynGAP1 D338A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, polyPhen‑2 HumVar, and SIFT, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Optimized) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.335645 | Structured | 0.363354 | Uncertain | 0.460 | 0.438 | 0.375 | -10.639 | Likely Pathogenic | 0.902 | Likely Pathogenic | Ambiguous | 1.22 | Ambiguous | 0.3 | 1.11 | Ambiguous | 1.17 | Ambiguous | 0.16 | Likely Benign | 0.479 | Likely Benign | -5.74 | Deleterious | 0.625 | Possibly Damaging | 0.192 | Benign | 1.73 | Pathogenic | 0.11 | Tolerated | 0.3830 | 0.5988 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||
| c.1015A>G | K339E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K339E missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, and premPS. Tools that predict a pathogenic effect include PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus score, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is “Likely Pathogenic.” High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not contradict any ClinVar annotation because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.447574 | Structured | 0.384153 | Uncertain | 0.535 | 0.465 | 0.250 | -14.284 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 0.13 | Likely Benign | 0.1 | -0.03 | Likely Benign | 0.05 | Likely Benign | 0.40 | Likely Benign | 0.482 | Likely Benign | -3.00 | Deleterious | 0.939 | Possibly Damaging | 0.670 | Possibly Damaging | 1.92 | Pathogenic | 0.03 | Affected | 0.3421 | 0.0882 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1016A>C | K339T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K339T missense variant is not reported in ClinVar and is absent from gnomAD. Consensus predictions from multiple in‑silico tools are split: benign calls come from FoldX, Rosetta, Foldetta, premPS, and SIFT, whereas pathogenic calls are made by SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments further highlight this discordance: the SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic effect, whereas Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, predicts a benign impact. AlphaMissense‑Optimized returned an uncertain result and is treated as unavailable. Overall, the majority of robust predictors lean toward pathogenicity, and this conclusion does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.447574 | Structured | 0.384153 | Uncertain | 0.535 | 0.465 | 0.250 | -10.061 | Likely Pathogenic | 0.910 | Likely Pathogenic | Ambiguous | 0.32 | Likely Benign | 0.0 | 0.10 | Likely Benign | 0.21 | Likely Benign | -0.04 | Likely Benign | 0.512 | Likely Pathogenic | -4.73 | Deleterious | 0.991 | Probably Damaging | 0.795 | Possibly Damaging | 1.94 | Pathogenic | 0.10 | Tolerated | 0.1900 | 0.3020 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1016A>T | K339M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K339M missense variant is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include FoldX and premPS, whereas the majority of tools predict a pathogenic impact: SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools with uncertain or inconclusive results are Rosetta and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for K339M. This conclusion is not contradicted by ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.447574 | Structured | 0.384153 | Uncertain | 0.535 | 0.465 | 0.250 | -13.387 | Likely Pathogenic | 0.976 | Likely Pathogenic | Likely Pathogenic | 0.23 | Likely Benign | 0.0 | 0.88 | Ambiguous | 0.56 | Ambiguous | -0.37 | Likely Benign | 0.575 | Likely Pathogenic | -4.95 | Deleterious | 0.999 | Probably Damaging | 0.964 | Probably Damaging | 1.92 | Pathogenic | 0.01 | Affected | 0.0967 | 0.3541 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1018G>A | A340T 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant A340T is reported in gnomAD (ID 6‑33437923‑G‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus all classify the change as benign or likely benign. Only two tools predict pathogenicity—polyPhen‑2 HumDiv and FATHMM—while stability‑based methods (FoldX, Rosetta, premPS, Foldetta) return uncertain or inconclusive results. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized scores benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely benign, and Foldetta provides no definitive stability change. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.390993 | Structured | 0.410781 | Uncertain | 0.558 | 0.485 | 0.250 | 6-33437923-G-A | -3.286 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.84 | Ambiguous | 0.2 | 0.96 | Ambiguous | 0.90 | Ambiguous | -0.54 | Ambiguous | 0.105 | Likely Benign | 0.62 | Neutral | 0.454 | Possibly Damaging | 0.192 | Benign | 1.93 | Pathogenic | 0.47 | Tolerated | 3.42 | 13 | 0.1740 | 0.7297 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||||
| c.1019C>A | A340E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A340E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Default. Predictions that are inconclusive or unavailable are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports a likely pathogenic outcome, while the protein‑folding stability method Foldetta is uncertain. AlphaMissense‑Optimized also yields an uncertain result. Overall, the majority of evidence points toward a pathogenic impact, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.390993 | Structured | 0.410781 | Uncertain | 0.558 | 0.485 | 0.250 | -8.225 | Likely Pathogenic | 0.803 | Likely Pathogenic | Ambiguous | 0.63 | Ambiguous | 0.4 | 1.55 | Ambiguous | 1.09 | Ambiguous | 0.06 | Likely Benign | 0.138 | Likely Benign | -0.33 | Neutral | 0.625 | Possibly Damaging | 0.252 | Benign | 1.91 | Pathogenic | 0.39 | Tolerated | 0.1418 | 0.2160 | 0 | -1 | -5.3 | 58.04 | |||||||||||||||||||||||||||||
| c.1019C>T | A340V 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A340V variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta (combining FoldX‑MD and Rosetta stability outputs) is uncertain and therefore unavailable for interpretation. Overall, the majority of evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.390993 | Structured | 0.410781 | Uncertain | 0.558 | 0.485 | 0.250 | -6.427 | Likely Benign | 0.174 | Likely Benign | Likely Benign | 0.69 | Ambiguous | 0.3 | 0.32 | Likely Benign | 0.51 | Ambiguous | 0.40 | Likely Benign | 0.102 | Likely Benign | -1.81 | Neutral | 0.801 | Possibly Damaging | 0.315 | Benign | 2.09 | Pathogenic | 0.57 | Tolerated | 0.1197 | 0.5780 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.101A>G | Y34C 2D  AIThe SynGAP1 missense variant Y34C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized reports Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for Y34C, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.529623 | Disordered | 0.435847 | Uncertain | 0.303 | 0.855 | 0.375 | -3.730 | Likely Benign | 0.171 | Likely Benign | Likely Benign | 0.196 | Likely Benign | 0.87 | Neutral | 0.943 | Possibly Damaging | 0.941 | Probably Damaging | 4.32 | Benign | 0.00 | Affected | 0.3010 | 0.2719 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||||||||||||
| c.1025A>G | Y342C 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Y342C is listed in ClinVar as Benign (ClinVar ID 1213078.0) and is observed in gnomAD (ID 6‑33437930‑A‑G). Across general prediction tools, benign calls are made by REVEL and AlphaMissense‑Optimized, whereas pathogenic calls are made by FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). Uncertain results are reported by premPS and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized predicting Benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicting Pathogenic. Overall, the majority of predictions support a pathogenic effect, contradicting the ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.366687 | Structured | 0.408200 | Uncertain | 0.866 | 0.487 | 0.250 | Benign/Likely benign | 2 | 6-33437930-A-G | 21 | 1.30e-5 | -7.596 | In-Between | 0.682 | Likely Pathogenic | Likely Benign | 2.48 | Destabilizing | 0.1 | 2.73 | Destabilizing | 2.61 | Destabilizing | 0.92 | Ambiguous | 0.404 | Likely Benign | -6.67 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.72 | Pathogenic | 0.02 | Affected | 3.37 | 25 | 0.2836 | 0.2870 | 0 | -2 | 3.8 | -60.04 | 242.4 | 62.8 | 0.1 | 0.0 | -0.1 | 0.2 | Potentially Pathogenic | The phenol ring of Tyr342, located at the end of an anti-parallel β sheet strand (res. Gly341-Pro349), faces outward in the C2 domain. This phenol ring contributes to a triple tyrosine stack (Tyr342, Tyr328, and Tyr281) that links together three anti-parallel β sheet strands. Additionally, it shields Gly344 from the solvent, reducing its exposure and providing stability for the β-sandwich. This motif also contributes to a twist formation in the β sheet.In the variant simulations, the Cys342 side chain cannot participate in the stack formation. Instead, its thiol group forms a hydrogen bond with the backbone carbonyl group of Leu327. Although these changes in surface interactions could weaken the characteristic twist that strengthens the β sheet fold, no major structural effects are observed in the variant simulations. The residue swap could also affect the SynGAP-membrane association; however, this phenomenon cannot be addressed using solvent-only simulations. Notably, the thiol group of cysteine is not a particularly strong hydrogen-bonding partner, which could mitigate the negative effects of the residue swap. | ||||||||||||||
| c.1028T>C | V343A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V343A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and AlphaMissense‑Optimized, whereas the remaining tools—SGM‑Consensus, FoldX (uncertain), Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely pathogenic effect, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenicity. Overall, the preponderance of evidence points to a pathogenic effect for V343A, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.291804 | Structured | 0.383911 | Uncertain | 0.882 | 0.497 | 0.250 | -8.088 | Likely Pathogenic | 0.588 | Likely Pathogenic | Likely Benign | 1.66 | Ambiguous | 0.1 | 2.33 | Destabilizing | 2.00 | Destabilizing | 1.69 | Destabilizing | 0.218 | Likely Benign | -3.15 | Deleterious | 0.826 | Possibly Damaging | 0.551 | Possibly Damaging | 1.63 | Pathogenic | 0.01 | Affected | 0.3137 | 0.3301 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1037T>C | V346A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V346A is reported in gnomAD (6‑33437942‑T‑C) but has no ClinVar entry. Prediction tools that agree on a benign effect are limited to REVEL, whereas the remaining tools (FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) uniformly predict a pathogenic impact; AlphaMissense‑Optimized is uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is inconclusive, the SGM Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a destabilizing, pathogenic effect. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.260850 | Structured | 0.350921 | Uncertain | 0.949 | 0.461 | 0.000 | 6-33437942-T-C | 1 | 6.20e-7 | -8.556 | Likely Pathogenic | 0.856 | Likely Pathogenic | Ambiguous | 2.72 | Destabilizing | 0.2 | 2.73 | Destabilizing | 2.73 | Destabilizing | 1.92 | Destabilizing | 0.477 | Likely Benign | -3.68 | Deleterious | 0.994 | Probably Damaging | 0.970 | Probably Damaging | 1.70 | Pathogenic | 0.01 | Affected | 3.37 | 25 | 0.3341 | 0.2967 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||
| c.1039A>C | T347P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T347P variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and FATHMM. Three tools (FoldX, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.268042 | Structured | 0.349915 | Uncertain | 0.951 | 0.434 | 0.000 | -9.323 | Likely Pathogenic | 0.417 | Ambiguous | Likely Benign | 0.55 | Ambiguous | 0.0 | 0.28 | Likely Benign | 0.42 | Likely Benign | 0.65 | Ambiguous | 0.236 | Likely Benign | -2.41 | Neutral | 0.627 | Possibly Damaging | 0.139 | Benign | 1.63 | Pathogenic | 0.12 | Tolerated | 0.2000 | 0.5350 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.1039A>G | T347A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T347A is catalogued in gnomAD (ID 6‑33437944‑A‑G) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated. Only FATHMM predicts a pathogenic outcome, while Foldetta, premPS, and Rosetta are inconclusive. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign, SGM‑Consensus is Likely Benign, and Foldetta remains uncertain. Overall, the consensus of the majority of tools indicates the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.268042 | Structured | 0.349915 | Uncertain | 0.951 | 0.434 | 0.000 | 6-33437944-A-G | 9 | 5.58e-6 | -5.858 | Likely Benign | 0.086 | Likely Benign | Likely Benign | 0.34 | Likely Benign | 0.1 | 0.70 | Ambiguous | 0.52 | Ambiguous | 0.70 | Ambiguous | 0.093 | Likely Benign | -1.52 | Neutral | 0.031 | Benign | 0.016 | Benign | 1.68 | Pathogenic | 0.42 | Tolerated | 3.37 | 25 | 0.4032 | 0.4615 | 0 | 1 | 2.5 | -30.03 | ||||||||||||||||||||||||
| c.1040C>A | T347N 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T347N is listed in ClinVar with an uncertain significance (ClinVar ID 3672484.0) and is present in the gnomAD database (gnomAD ID 6‑33437945‑C‑A). Prediction tools that uniformly indicate a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome. The SGM‑Consensus, which aggregates the majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, resolves to “Likely Benign” (3 benign vs. 1 pathogenic). High‑accuracy assessments are consistent: AlphaMissense‑Optimized is benign, the SGM‑Consensus is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. Overall, the collective evidence points to a benign effect, aligning with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.268042 | Structured | 0.349915 | Uncertain | 0.951 | 0.434 | 0.000 | Uncertain | 1 | 6-33437945-C-A | 9 | 5.58e-6 | -5.545 | Likely Benign | 0.165 | Likely Benign | Likely Benign | 0.41 | Likely Benign | 0.1 | 0.46 | Likely Benign | 0.44 | Likely Benign | -0.06 | Likely Benign | 0.059 | Likely Benign | 1.96 | Neutral | 0.001 | Benign | 0.001 | Benign | 1.67 | Pathogenic | 0.60 | Tolerated | 3.37 | 25 | 0.1070 | 0.4359 | 0 | 0 | -2.8 | 13.00 | ||||||||||||||||||||||
| c.1040C>T | T347I 2D 3DClick to see structure in 3D Viewer AISynGAP1 T347I is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that provide definitive calls cluster into two groups: benign predictions come from REVEL, premPS, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized; pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, ESM1b, and FATHMM. Tools with inconclusive outputs (FoldX, Rosetta, Foldetta, AlphaMissense‑Default) are treated as unavailable. High‑accuracy assessments further split the evidence: AlphaMissense‑Optimized predicts benign, while the SGM consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—predicts pathogenic; Foldetta remains uncertain. Consequently, the variant’s pathogenicity is ambiguous, with an equal number of strong benign and pathogenic calls and no consensus from the most reliable methods. The variant is therefore most likely uncertain, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.268042 | Structured | 0.349915 | Uncertain | 0.951 | 0.434 | 0.000 | -10.148 | Likely Pathogenic | 0.387 | Ambiguous | Likely Benign | -0.79 | Ambiguous | 0.1 | -0.94 | Ambiguous | -0.87 | Ambiguous | 0.29 | Likely Benign | 0.079 | Likely Benign | -3.12 | Deleterious | 0.627 | Possibly Damaging | 0.139 | Benign | 1.70 | Pathogenic | 0.08 | Tolerated | 0.0885 | 0.6298 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.1043T>C | V348A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V348A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign predictions come from REVEL, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized, whereas pathogenic predictions are reported by SGM‑Consensus, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments further highlight the discrepancy: AlphaMissense‑Optimized predicts a benign effect, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) classifies the variant as pathogenic. Taken together, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict ClinVar status, which currently has no entry for V348A. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.170161 | Structured | 0.346556 | Uncertain | 0.951 | 0.414 | 0.000 | -9.993 | Likely Pathogenic | 0.767 | Likely Pathogenic | Likely Benign | 2.46 | Destabilizing | 0.1 | 3.13 | Destabilizing | 2.80 | Destabilizing | 2.23 | Destabilizing | 0.175 | Likely Benign | -3.42 | Deleterious | 0.622 | Possibly Damaging | 0.152 | Benign | 1.59 | Pathogenic | 0.14 | Tolerated | 0.3223 | 0.2437 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1045C>A | P349T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P349T is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include REVEL, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and the SGM‑Consensus score (which is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools with inconclusive results are AlphaMissense‑Default, FoldX, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as pathogenic, and Foldetta as uncertain. Overall, the majority of predictions lean toward pathogenicity, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for this variant. Thus, the variant is most likely pathogenic based on the available computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.167087 | Structured | 0.348607 | Uncertain | 0.947 | 0.396 | 0.000 | -9.736 | Likely Pathogenic | 0.420 | Ambiguous | Likely Benign | 1.37 | Ambiguous | 0.1 | 2.56 | Destabilizing | 1.97 | Ambiguous | 0.76 | Ambiguous | 0.246 | Likely Benign | -6.12 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 1.57 | Pathogenic | 0.07 | Tolerated | 0.1615 | 0.6238 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||
| c.1046C>A | P349Q 2D 3DClick to see structure in 3D Viewer AISynGAP1 P349Q is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, and AlphaMissense‑Optimized, while pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Default, Foldetta) give uncertain or inconclusive results. High‑accuracy assessments further clarify the variant’s likely effect: AlphaMissense‑Optimized predicts a benign outcome; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a pathogenic outcome; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, remains uncertain. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion does not conflict with the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.167087 | Structured | 0.348607 | Uncertain | 0.947 | 0.396 | 0.000 | -10.545 | Likely Pathogenic | 0.508 | Ambiguous | Likely Benign | 0.98 | Ambiguous | 0.1 | 1.87 | Ambiguous | 1.43 | Ambiguous | 0.91 | Ambiguous | 0.345 | Likely Benign | -6.40 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 1.54 | Pathogenic | 0.06 | Tolerated | 0.1480 | 0.4766 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||
| c.1046C>G | P349R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P349R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only REVEL, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and Rosetta. Uncertain or inconclusive results come from FoldX, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show that AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Pathogenic, and Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for P349R. This conclusion is not contradicted by ClinVar status, as the variant has no ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.167087 | Structured | 0.348607 | Uncertain | 0.947 | 0.396 | 0.000 | -14.001 | Likely Pathogenic | 0.791 | Likely Pathogenic | Ambiguous | 0.99 | Ambiguous | 0.1 | 2.15 | Destabilizing | 1.57 | Ambiguous | 0.93 | Ambiguous | 0.335 | Likely Benign | -7.22 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 1.55 | Pathogenic | 0.01 | Affected | 0.1395 | 0.3009 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1049T>C | V350A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant V350A is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), premPS, PROVEAN, ESM1b, and FATHMM. Stability‑based methods FoldX, Rosetta, and Foldetta give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as pathogenic, and Foldetta as unavailable. Overall, the predictions are split, with a slight edge toward pathogenicity from the consensus and high‑accuracy tools. Therefore, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.179055 | Structured | 0.353227 | Uncertain | 0.931 | 0.371 | 0.000 | -8.323 | Likely Pathogenic | 0.280 | Likely Benign | Likely Benign | 1.42 | Ambiguous | 0.0 | 1.97 | Ambiguous | 1.70 | Ambiguous | 2.07 | Destabilizing | 0.096 | Likely Benign | -2.73 | Deleterious | 0.435 | Benign | 0.115 | Benign | 1.64 | Pathogenic | 0.55 | Tolerated | 0.3207 | 0.2967 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.104T>C | V35A 2D AIThe SynGAP1 missense variant V35A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 HumDiv, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. Only two tools, polyPhen‑2 HumVar and SIFT, predict pathogenic. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. High‑accuracy assessments reinforce this: AlphaMissense‑Optimized is benign and the SGM‑Consensus is likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact, and this is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.613573 | Disordered | 0.434838 | Uncertain | 0.360 | 0.851 | 0.375 | -3.552 | Likely Benign | 0.120 | Likely Benign | Likely Benign | 0.098 | Likely Benign | -0.05 | Neutral | 0.267 | Benign | 0.481 | Possibly Damaging | 4.25 | Benign | 0.00 | Affected | 0.2340 | 0.2144 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||||||||||||
| c.1052C>A | A351D 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A351D missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, AlphaMissense‑Optimized, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and FATHMM; AlphaMissense‑Default and ESM1b are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leaning toward pathogenic due to two pathogenic and two uncertain calls. Overall, the majority of tools predict a benign outcome, and this does not contradict the lack of ClinVar annotation. Thus, based on the available predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.216401 | Structured | 0.362025 | Uncertain | 0.925 | 0.342 | 0.000 | -7.917 | In-Between | 0.543 | Ambiguous | Likely Benign | -0.02 | Likely Benign | 0.0 | -0.12 | Likely Benign | -0.07 | Likely Benign | 0.49 | Likely Benign | 0.114 | Likely Benign | -3.61 | Deleterious | 0.842 | Possibly Damaging | 0.321 | Benign | 1.77 | Pathogenic | 0.11 | Tolerated | 0.1535 | 0.1386 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1052C>T | A351V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A351V is not reported in ClinVar and is absent from gnomAD. Computational predictions cluster into two groups: benign (REVEL, FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Default, AlphaMissense‑Optimized, polyPhen2_HumVar) and pathogenic (PROVEAN, polyPhen2_HumDiv, SIFT, ESM1b, FATHMM, SGM‑Consensus). High‑accuracy tools give a mixed signal: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the balance of evidence leans toward a benign effect, but the presence of several pathogenic predictions introduces uncertainty. The variant is most likely benign based on the current computational data, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.216401 | Structured | 0.362025 | Uncertain | 0.925 | 0.342 | 0.000 | -9.002 | Likely Pathogenic | 0.124 | Likely Benign | Likely Benign | 0.09 | Likely Benign | 0.0 | 0.19 | Likely Benign | 0.14 | Likely Benign | 0.29 | Likely Benign | 0.052 | Likely Benign | -2.84 | Deleterious | 0.915 | Possibly Damaging | 0.321 | Benign | 1.66 | Pathogenic | 0.03 | Affected | 0.1140 | 0.6565 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1054A>C | T352P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant T352P has no ClinVar entry and is not reported in gnomAD. Prediction tools that converge on a benign outcome include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, FATHMM, and Rosetta. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, the SGM‑Consensus as Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Uncertain. Overall, the balance of evidence favors a benign interpretation; this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.137348 | Structured | 0.367886 | Uncertain | 0.926 | 0.329 | 0.000 | -3.562 | Likely Benign | 0.138 | Likely Benign | Likely Benign | 1.04 | Ambiguous | 0.1 | 2.57 | Destabilizing | 1.81 | Ambiguous | 0.51 | Ambiguous | 0.159 | Likely Benign | -2.31 | Neutral | 0.627 | Possibly Damaging | 0.196 | Benign | 1.72 | Pathogenic | 0.20 | Tolerated | 0.2244 | 0.5968 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1055C>A | T352N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T352N is listed in ClinVar as Benign (ClinVar ID 590151.0) and is present in the gnomAD database (gnomAD ID 6‑33437960‑C‑A). Across the broad panel of in‑silico predictors, 13 tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) uniformly report a benign effect, whereas only FATHMM predicts pathogenicity. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is also benign. No predictions or stability analyses are missing or inconclusive. Overall, the computational evidence strongly supports a benign classification, consistent with the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.137348 | Structured | 0.367886 | Uncertain | 0.926 | 0.329 | 0.000 | Likely Benign | 1 | 6-33437960-C-A | 2 | 1.24e-6 | -4.817 | Likely Benign | 0.117 | Likely Benign | Likely Benign | 0.20 | Likely Benign | 0.0 | -0.04 | Likely Benign | 0.08 | Likely Benign | 0.45 | Likely Benign | 0.027 | Likely Benign | -0.92 | Neutral | 0.255 | Benign | 0.057 | Benign | 1.75 | Pathogenic | 0.19 | Tolerated | 3.37 | 25 | 0.1311 | 0.4358 | 0 | 0 | -2.8 | 13.00 | 208.4 | -14.5 | -0.2 | 0.1 | -0.1 | 0.0 | X | Potentially Benign | Thr352 is located in a short α helical section within a loop connecting two β strands (res. Gly341-Pro349, res. Thr359-Pro364) originating from two different anti-parallel β sheets of the C2 domain. In the WT simulations, the side chain hydroxyl and backbone amide groups of Thr354 form hydrogen bonds with the backbone carbonyl group of Pro349 at the end of the preceding β strand. This arrangement likely stabilizes the α helical section and aids in folding, keeping the short secondary structure element intact in the variant simulations. However, the carboxamide group of the Asn352 side chain does not form hydrogen bonds with the backbone carbonyl group of Pro349. Instead, it packs against the cyclic ring and forms hydrogen bonds with the phenol group of the Tyr363 side chain in the other β strand. | |||||||||||||
| c.1055C>T | T352I 2D AISynGAP1 T352I is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL, Rosetta, premPS, SIFT, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen‑2 HumVar. Those that predict pathogenicity are SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, ESM1b, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus majority vote (AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) as benign. With seven benign versus five pathogenic predictions and two high‑accuracy benign versus one pathogenic, the evidence leans toward a benign effect. This conclusion does not contradict ClinVar status, as no ClinVar entry exists for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.137348 | Structured | 0.367886 | Uncertain | 0.926 | 0.329 | 0.000 | -8.023 | Likely Pathogenic | 0.321 | Likely Benign | Likely Benign | -0.54 | Ambiguous | 0.7 | 0.43 | Likely Benign | -0.06 | Likely Benign | 0.09 | Likely Benign | 0.099 | Likely Benign | -3.02 | Deleterious | 0.627 | Possibly Damaging | 0.196 | Benign | 1.67 | Pathogenic | 0.14 | Tolerated | 0.1009 | 0.6484 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.1061C>A | A354D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A354D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and SIFT. Those that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. Four tools (Foldetta, premPS, ESM1b, and Rosetta) return uncertain results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors a pathogenic interpretation. Foldetta’s stability prediction is uncertain. Overall, the majority of consensus tools lean toward a benign effect, and there is no ClinVar annotation to contradict this assessment. Thus, the variant is most likely benign based on the available predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.381329 | Uncertain | 0.882 | 0.335 | 0.125 | -7.914 | In-Between | 0.725 | Likely Pathogenic | Likely Benign | 0.22 | Likely Benign | 0.1 | 1.10 | Ambiguous | 0.66 | Ambiguous | 0.57 | Ambiguous | 0.087 | Likely Benign | -1.49 | Neutral | 0.255 | Benign | 0.053 | Benign | 1.75 | Pathogenic | 0.07 | Tolerated | 0.1759 | 0.1345 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1061C>T | A354V 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A354V is not reported in ClinVar and is present in the gnomAD database (variant ID 6‑33437966‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are SIFT and FATHMM. The high‑accuracy consensus methods support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is also benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a benign outcome. Predictions from FoldX and Rosetta are inconclusive and are treated as unavailable. Overall, the majority of evidence indicates that A354V is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.203355 | Structured | 0.381329 | Uncertain | 0.882 | 0.335 | 0.125 | 6-33437966-C-T | 1 | 6.26e-7 | -6.223 | Likely Benign | 0.122 | Likely Benign | Likely Benign | 0.65 | Ambiguous | 0.1 | -1.02 | Ambiguous | -0.19 | Likely Benign | 0.18 | Likely Benign | 0.027 | Likely Benign | -1.11 | Neutral | 0.146 | Benign | 0.038 | Benign | 1.81 | Pathogenic | 0.05 | Affected | 3.38 | 24 | 0.1429 | 0.6388 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||
| c.1066C>A | R356S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant R356S is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL and SIFT, whereas the remaining tools—premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized—predict it to be pathogenic. The SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments further support this: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus is pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, is inconclusive. Taken together, the preponderance of evidence points to a pathogenic effect for R356S, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.219301 | Structured | 0.395028 | Uncertain | 0.802 | 0.373 | 0.250 | -13.059 | Likely Pathogenic | 0.965 | Likely Pathogenic | Likely Pathogenic | 1.00 | Ambiguous | 0.1 | 0.79 | Ambiguous | 0.90 | Ambiguous | 1.04 | Destabilizing | 0.292 | Likely Benign | -5.28 | Deleterious | 0.993 | Probably Damaging | 0.982 | Probably Damaging | 1.84 | Pathogenic | 0.07 | Tolerated | 0.2891 | 0.4491 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||
| c.1067G>A | R356H 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant R356H is recorded in ClinVar as benign (ClinVar ID 2984966.0) and is present in the gnomAD database (6‑33437972‑G‑A). Prediction tools that indicate a benign effect include REVEL, Rosetta, Foldetta, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default, with the SGM‑Consensus also labeling it likely pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of predictions support a pathogenic impact, which contradicts the ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.219301 | Structured | 0.395028 | Uncertain | 0.802 | 0.373 | 0.250 | Likely Benign | 1 | 6-33437972-G-A | 9 | 5.66e-6 | -11.453 | Likely Pathogenic | 0.614 | Likely Pathogenic | Likely Benign | 0.59 | Ambiguous | 0.1 | -0.27 | Likely Benign | 0.16 | Likely Benign | 1.17 | Destabilizing | 0.314 | Likely Benign | -4.43 | Deleterious | 0.999 | Probably Damaging | 0.987 | Probably Damaging | 1.70 | Pathogenic | 0.01 | Affected | 3.39 | 22 | 0.3493 | 0.2206 | 0 | 2 | 1.3 | -19.05 | ||||||||||||||||||||||
| c.1067G>C | R356P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 R356P missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FoldX, whereas a majority of tools (premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic impact. Predictions that are inconclusive are AlphaMissense‑Optimized, Rosetta, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for R356P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.219301 | Structured | 0.395028 | Uncertain | 0.802 | 0.373 | 0.250 | -12.956 | Likely Pathogenic | 0.919 | Likely Pathogenic | Ambiguous | 0.48 | Likely Benign | 0.1 | 0.72 | Ambiguous | 0.60 | Ambiguous | 1.05 | Destabilizing | 0.451 | Likely Benign | -6.23 | Deleterious | 0.998 | Probably Damaging | 0.992 | Probably Damaging | 1.72 | Pathogenic | 0.02 | Affected | 0.2213 | 0.5033 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.106C>T | H36Y 2D  AIThe SynGAP1 missense variant H36Y is listed in ClinVar with an uncertain significance (ClinVar ID 2089635.0) and is present in the gnomAD database (gnomAD ID 6‑33423515‑C‑T). Functional prediction tools largely agree that the substitution is benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all report a benign effect. Only SIFT predicts a pathogenic outcome. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled Likely Benign. No Foldetta stability prediction is available. Overall, the computational evidence overwhelmingly supports a benign classification, which is consistent with the ClinVar designation of uncertain significance rather than a pathogenic claim. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.433974 | Uncertain | 0.334 | 0.834 | 0.375 | Uncertain | 1 | 6-33423515-C-T | 2 | 1.24e-6 | -3.461 | Likely Benign | 0.139 | Likely Benign | Likely Benign | 0.023 | Likely Benign | -1.03 | Neutral | 0.219 | Benign | 0.066 | Benign | 4.16 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1265 | 0.5024 | 0 | 2 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||||
| c.1070A>C | H357P 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 H357P missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are Rosetta, PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Two tools give uncertain results: Foldetta (protein‑folding stability) and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign, while Foldetta remains uncertain. Overall, the majority of evidence points to a benign effect. Thus, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.203355 | Structured | 0.399052 | Uncertain | 0.861 | 0.413 | 0.250 | -7.953 | In-Between | 0.267 | Likely Benign | Likely Benign | 0.17 | Likely Benign | 0.9 | 3.52 | Destabilizing | 1.85 | Ambiguous | -0.06 | Likely Benign | 0.197 | Likely Benign | -2.70 | Deleterious | 0.936 | Possibly Damaging | 0.469 | Possibly Damaging | 4.18 | Benign | 0.17 | Tolerated | 0.2433 | 0.4468 | 0 | -2 | 1.6 | -40.02 | ||||||||||||||||||||||||||||||
| c.1075A>C | T359P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T359P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and FATHMM. Uncertain or inconclusive results come from FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.281712 | Structured | 0.414952 | Uncertain | 0.939 | 0.480 | 0.250 | -6.884 | Likely Benign | 0.319 | Likely Benign | Likely Benign | 0.97 | Ambiguous | 0.1 | 1.27 | Ambiguous | 1.12 | Ambiguous | 0.68 | Ambiguous | 0.248 | Likely Benign | -2.36 | Neutral | 0.627 | Possibly Damaging | 0.091 | Benign | 1.78 | Pathogenic | 0.14 | Tolerated | 0.2352 | 0.5611 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1076C>A | T359K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T359K missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM. The remaining tools—FoldX, Rosetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of predictions (seven benign vs. two pathogenic) support a benign classification, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.281712 | Structured | 0.414952 | Uncertain | 0.939 | 0.480 | 0.250 | -10.822 | Likely Pathogenic | 0.517 | Ambiguous | Likely Benign | -0.65 | Ambiguous | 0.1 | 0.66 | Ambiguous | 0.01 | Likely Benign | 0.75 | Ambiguous | 0.163 | Likely Benign | -2.23 | Neutral | 0.255 | Benign | 0.045 | Benign | 1.79 | Pathogenic | 0.24 | Tolerated | 0.1618 | 0.3483 | 0 | -1 | -3.2 | 27.07 | ||||||||||||||||||||||||||||||
| c.1079A>C | E360A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E360A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only premPS. All other evaluated tools—SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—predict a pathogenic impact. High‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic effect, and Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Because the majority of consensus tools predict pathogenicity and no ClinVar entry contradicts this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.250310 | Structured | 0.421183 | Uncertain | 0.955 | 0.498 | 0.250 | -13.229 | Likely Pathogenic | 0.939 | Likely Pathogenic | Ambiguous | 1.37 | Ambiguous | 0.1 | 1.72 | Ambiguous | 1.55 | Ambiguous | 0.39 | Likely Benign | 0.545 | Likely Pathogenic | -5.52 | Deleterious | 0.997 | Probably Damaging | 0.980 | Probably Damaging | 1.63 | Pathogenic | 0.01 | Affected | 0.4295 | 0.8243 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1079A>G | E360G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E360G is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: premPS is the only tool that predicts a benign outcome, whereas all other evaluated algorithms—including REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the substitution as pathogenic. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized returns a pathogenic prediction; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a pathogenic effect. Taken together, the overwhelming majority of evidence points to a pathogenic impact for E360G, and this conclusion is consistent with the absence of a ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.250310 | Structured | 0.421183 | Uncertain | 0.955 | 0.498 | 0.250 | -13.972 | Likely Pathogenic | 0.971 | Likely Pathogenic | Likely Pathogenic | 2.55 | Destabilizing | 0.1 | 2.99 | Destabilizing | 2.77 | Destabilizing | 0.31 | Likely Benign | 0.569 | Likely Pathogenic | -6.43 | Deleterious | 0.999 | Probably Damaging | 0.986 | Probably Damaging | 1.68 | Pathogenic | 0.04 | Affected | 0.2993 | 0.6935 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.107A>C | H36P 2D AIThe SynGAP1 missense variant H36P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments show AlphaMissense‑Optimized as Benign and the SGM‑Consensus as Likely Benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.637480 | Disordered | 0.433974 | Uncertain | 0.334 | 0.834 | 0.375 | -2.285 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 0.171 | Likely Benign | -1.23 | Neutral | 0.182 | Benign | 0.046 | Benign | 4.16 | Benign | 0.00 | Affected | 0.2097 | 0.4723 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||||||||||||
| c.1087T>C | Y363H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y363H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized, while the majority of other in silico predictors (Rosetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, FATHMM, AlphaMissense‑Default, Foldetta) indicate a pathogenic impact; FoldX and ESM1b are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the preponderance of evidence points to a pathogenic effect for Y363H, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.321458 | Structured | 0.435392 | Uncertain | 0.954 | 0.586 | 0.125 | -7.003 | In-Between | 0.747 | Likely Pathogenic | Likely Benign | 1.75 | Ambiguous | 0.1 | 2.40 | Destabilizing | 2.08 | Destabilizing | 1.04 | Destabilizing | 0.419 | Likely Benign | -4.38 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 1.56 | Pathogenic | 0.01 | Affected | 0.3267 | 0.2021 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.1090C>A | P364T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P364T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Four tools (FoldX, Foldetta, premPS, ESM1b) return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evaluated tools (five benign versus four pathogenic) lean toward a benign classification, and this assessment does not contradict any ClinVar annotation because no ClinVar claim exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.390993 | Structured | 0.439474 | Uncertain | 0.942 | 0.590 | 0.250 | -7.951 | In-Between | 0.230 | Likely Benign | Likely Benign | 1.18 | Ambiguous | 0.6 | 0.47 | Likely Benign | 0.83 | Ambiguous | 0.53 | Ambiguous | 0.342 | Likely Benign | -5.60 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 1.60 | Pathogenic | 0.09 | Tolerated | 0.1660 | 0.5726 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1091C>G | P364R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P364R missense variant is not reported in ClinVar (status: None) and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect include SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain or inconclusive results come from FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments: AlphaMissense‑Optimized predicts benign; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic; Foldetta’s stability analysis is uncertain. Overall, the majority of evaluated tools (seven pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.390993 | Structured | 0.439474 | Uncertain | 0.942 | 0.590 | 0.250 | -12.652 | Likely Pathogenic | 0.715 | Likely Pathogenic | Likely Benign | 0.95 | Ambiguous | 0.7 | 0.88 | Ambiguous | 0.92 | Ambiguous | 0.73 | Ambiguous | 0.416 | Likely Benign | -6.76 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 1.57 | Pathogenic | 0.12 | Tolerated | 0.1520 | 0.3977 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1094T>C | V365A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V365A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions are made by REVEL and polyPhen‑2 HumVar, whereas the remaining tools (SGM‑Consensus, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic effect. High‑accuracy assessments are consistent with a deleterious outcome: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) reports a pathogenic effect. Taken together, the majority of evidence supports a pathogenic classification for V365A, and this conclusion does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.414856 | Structured | 0.441505 | Uncertain | 0.923 | 0.608 | 0.250 | -8.954 | Likely Pathogenic | 0.867 | Likely Pathogenic | Ambiguous | 2.61 | Destabilizing | 0.1 | 2.62 | Destabilizing | 2.62 | Destabilizing | 2.10 | Destabilizing | 0.297 | Likely Benign | -3.18 | Deleterious | 0.622 | Possibly Damaging | 0.235 | Benign | 1.67 | Pathogenic | 0.01 | Affected | 0.2682 | 0.2962 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1096A>C | T366P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T366P is not reported in ClinVar and is absent from gnomAD. Consensus from multiple in‑silico predictors leans toward a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign) all classify the substitution as tolerated. In contrast, polyPhen‑2 HumDiv, FATHMM, Rosetta, and the Foldetta stability analysis predict a damaging or pathogenic outcome. FoldX reports an uncertain effect and is therefore not considered evidence. High‑accuracy tools give mixed results: AlphaMissense‑Optimized and the SGM‑Consensus both indicate benign, whereas Foldetta predicts pathogenic. Overall, the majority of predictors (8 benign vs. 4 pathogenic) support a benign classification, and this is consistent with the lack of ClinVar evidence. Thus, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.441902 | Uncertain | 0.897 | 0.642 | 0.250 | -6.483 | Likely Benign | 0.226 | Likely Benign | Likely Benign | 1.75 | Ambiguous | 0.5 | 3.10 | Destabilizing | 2.43 | Destabilizing | 0.47 | Likely Benign | 0.150 | Likely Benign | -2.49 | Neutral | 0.627 | Possibly Damaging | 0.139 | Benign | 1.70 | Pathogenic | 0.24 | Tolerated | 0.2250 | 0.6251 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1097C>A | T366N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense change T366N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) all indicate a benign or likely benign outcome. Only two tools—polyPhen‑2 HumDiv and FATHMM—suggest a pathogenic effect, while premPS remains uncertain. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also reports likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) classifies the variant as benign. Taken together, the majority of evidence supports a benign impact and is consistent with the absence of any ClinVar pathogenic annotation. Therefore, the variant is most likely benign and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.414856 | Structured | 0.441902 | Uncertain | 0.897 | 0.642 | 0.250 | -5.694 | Likely Benign | 0.176 | Likely Benign | Likely Benign | 0.06 | Likely Benign | 0.0 | 0.36 | Likely Benign | 0.21 | Likely Benign | 0.92 | Ambiguous | 0.038 | Likely Benign | -1.72 | Neutral | 0.454 | Possibly Damaging | 0.038 | Benign | 1.72 | Pathogenic | 0.34 | Tolerated | 0.1595 | 0.4934 | 0 | 0 | -2.8 | 13.00 | |||||||||||||||||||||||||||||
| c.1102C>A | P368T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P368T missense variant is not listed in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM. FoldX, Rosetta, and Foldetta report uncertain or inconclusive stability changes and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie (2 benign, 2 pathogenic) and thus inconclusive, and Foldetta remains uncertain. Overall, the predictions are evenly split between benign and pathogenic, providing no definitive classification. The variant’s status does not contradict ClinVar, which has no entry for it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -5.308 | Likely Benign | 0.284 | Likely Benign | Likely Benign | 1.95 | Ambiguous | 0.6 | 1.61 | Ambiguous | 1.78 | Ambiguous | 0.45 | Likely Benign | 0.188 | Likely Benign | -5.43 | Deleterious | 0.941 | Possibly Damaging | 0.527 | Possibly Damaging | 1.72 | Pathogenic | 0.01 | Affected | 0.1983 | 0.6155 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1103C>A | P368Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P368Q missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, ESM1b, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Remaining tools (AlphaMissense‑Default, FoldX, Rosetta, Foldetta, premPS) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of definitive predictions (five pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -6.019 | Likely Benign | 0.413 | Ambiguous | Likely Benign | 1.56 | Ambiguous | 0.8 | 1.71 | Ambiguous | 1.64 | Ambiguous | 0.71 | Ambiguous | 0.205 | Likely Benign | -5.10 | Deleterious | 0.991 | Probably Damaging | 0.881 | Possibly Damaging | 1.71 | Pathogenic | 0.04 | Affected | 0.1694 | 0.5283 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||
| c.1103C>G | P368R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P368R missense variant is not reported in ClinVar (status: None) and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect include SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain or inconclusive results come from FoldX, Rosetta, Foldetta, and premPS. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts benign, whereas the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts likely pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, is uncertain. Overall, the majority of evidence—including the SGM‑Consensus and several individual high‑accuracy tools—points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.363090 | Structured | 0.439989 | Uncertain | 0.580 | 0.677 | 0.250 | -9.564 | Likely Pathogenic | 0.736 | Likely Pathogenic | Likely Benign | 1.57 | Ambiguous | 1.0 | 1.54 | Ambiguous | 1.56 | Ambiguous | 0.58 | Ambiguous | 0.263 | Likely Benign | -6.07 | Deleterious | 0.991 | Probably Damaging | 0.881 | Possibly Damaging | 1.78 | Pathogenic | 0.00 | Affected | 0.1439 | 0.3922 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1105A>C | T369P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T369P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). Only FATHMM predicts a pathogenic outcome, while FoldX and Foldetta are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as Likely Benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact for T369P, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | -2.743 | Likely Benign | 0.066 | Likely Benign | Likely Benign | 1.20 | Ambiguous | 2.1 | 0.18 | Likely Benign | 0.69 | Ambiguous | 0.17 | Likely Benign | 0.138 | Likely Benign | -2.09 | Neutral | 0.396 | Benign | 0.142 | Benign | 1.83 | Pathogenic | 0.16 | Tolerated | 0.2589 | 0.6046 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1106C>A | T369K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T369K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FATHMM and AlphaMissense‑Default. Foldetta and Rosetta give uncertain results, which are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (two benign vs two pathogenic). Foldetta’s stability prediction is uncertain. Overall, the majority of evidence (nine benign vs two pathogenic) supports a benign classification. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | -5.884 | Likely Benign | 0.616 | Likely Pathogenic | Likely Benign | -0.10 | Likely Benign | 0.1 | 1.13 | Ambiguous | 0.52 | Ambiguous | 0.27 | Likely Benign | 0.102 | Likely Benign | -1.87 | Neutral | 0.118 | Benign | 0.054 | Benign | 1.84 | Pathogenic | 0.34 | Tolerated | 0.1387 | 0.3855 | 0 | -1 | -3.2 | 27.07 | ||||||||||||||||||||||||||||||
| c.1106C>T | T369I 2D AIThe SynGAP1 missense variant T369I is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. Stability‑based methods (FoldX, Rosetta, Foldetta) are inconclusive, providing no definitive evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as likely benign, and Foldetta as uncertain. Overall, the majority of evidence points to a benign impact; there is no conflict with ClinVar status, which contains no entry for this variant. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.468512 | Structured | 0.437011 | Uncertain | 0.417 | 0.707 | 0.500 | -6.759 | Likely Benign | 0.289 | Likely Benign | Likely Benign | 0.60 | Ambiguous | 0.8 | 1.41 | Ambiguous | 1.01 | Ambiguous | -0.08 | Likely Benign | 0.078 | Likely Benign | -2.37 | Neutral | 0.396 | Benign | 0.142 | Benign | 1.72 | Pathogenic | 0.13 | Tolerated | 0.1106 | 0.7207 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.110C>G | S37C 2D AIThe SynGAP1 missense variant S37C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.642678 | Disordered | 0.433492 | Uncertain | 0.317 | 0.806 | 0.500 | -4.304 | Likely Benign | 0.141 | Likely Benign | Likely Benign | 0.116 | Likely Benign | -1.18 | Neutral | 0.880 | Possibly Damaging | 0.923 | Probably Damaging | 3.89 | Benign | 0.00 | Affected | 0.1623 | 0.5775 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.1111A>T | S371C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S371C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv and SIFT. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Based on the overall consensus of the majority of evidence, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.444081 | Structured | 0.432086 | Uncertain | 0.294 | 0.746 | 0.375 | -6.330 | Likely Benign | 0.099 | Likely Benign | Likely Benign | 0.19 | Likely Benign | 0.2 | -0.34 | Likely Benign | -0.08 | Likely Benign | 0.23 | Likely Benign | 0.450 | Likely Benign | -1.41 | Neutral | 0.875 | Possibly Damaging | 0.359 | Benign | 4.61 | Benign | 0.02 | Affected | 0.1786 | 0.6580 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||
| c.1115G>A | G372E 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G372E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that agree on a pathogenic effect are REVEL, Rosetta, Foldetta, FATHMM, and AlphaMissense‑Default; FoldX is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, while Foldetta predicts pathogenic. The SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to a 2‑to‑2 split and is treated as unavailable. Overall, the majority of evidence (seven benign vs. five pathogenic) supports a benign classification. This conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.433034 | Structured | 0.430335 | Uncertain | 0.322 | 0.774 | 0.375 | -6.682 | Likely Benign | 0.604 | Likely Pathogenic | Likely Benign | 1.58 | Ambiguous | 0.4 | 2.91 | Destabilizing | 2.25 | Destabilizing | 0.34 | Likely Benign | 0.566 | Likely Pathogenic | -0.81 | Neutral | 0.001 | Benign | 0.001 | Benign | -0.74 | Pathogenic | 0.08 | Tolerated | 0.1606 | 0.4103 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1118G>A | G373E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G373E is listed in ClinVar with an Uncertain significance and is not present in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, Foldetta, SIFT, and AlphaMissense‑Default. Predictions from Rosetta and ESM1b are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as pathogenic. Overall, the majority of evidence points to a benign impact, which does not contradict the ClinVar status of Uncertain. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.529623 | Disordered | 0.429267 | Uncertain | 0.295 | 0.799 | 0.625 | Uncertain | 1 | -7.281 | In-Between | 0.569 | Likely Pathogenic | Likely Benign | 4.13 | Destabilizing | 3.2 | 0.52 | Ambiguous | 2.33 | Destabilizing | -0.02 | Likely Benign | 0.420 | Likely Benign | -0.69 | Neutral | 0.001 | Benign | 0.000 | Benign | 3.90 | Benign | 0.01 | Affected | 0.1572 | 0.4309 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||
| c.1124G>A | G375E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G375E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumDiv, FATHMM, and AlphaMissense‑Default; ESM1b remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) favors pathogenic, and Foldetta also predicts pathogenic. Overall, the majority of tools, including the high‑accuracy ones, indicate a pathogenic impact. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.604312 | Disordered | 0.428340 | Uncertain | 0.301 | 0.836 | 0.625 | -7.780 | In-Between | 0.600 | Likely Pathogenic | Likely Benign | 2.89 | Destabilizing | 1.4 | 9.47 | Destabilizing | 6.18 | Destabilizing | 0.45 | Likely Benign | 0.545 | Likely Pathogenic | -1.07 | Neutral | 0.845 | Possibly Damaging | 0.244 | Benign | 1.32 | Pathogenic | 0.09 | Tolerated | 0.1619 | 0.4299 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1126G>A | G376S 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant G376S is reported in gnomAD (ID 6‑33438031‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Benign). Pathogenic predictions arise from FoldX, polyPhen‑2 (HumDiv and HumVar), and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates benign, while Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. Overall, the majority of evidence points to a benign effect; this conclusion is not contradicted by ClinVar, which contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.680603 | Disordered | 0.428979 | Uncertain | 0.326 | 0.869 | 0.625 | 6-33438031-G-A | 1 | 6.21e-7 | -4.913 | Likely Benign | 0.087 | Likely Benign | Likely Benign | 2.30 | Destabilizing | 0.5 | -0.45 | Likely Benign | 0.93 | Ambiguous | 0.32 | Likely Benign | 0.471 | Likely Benign | -0.73 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.33 | Pathogenic | 0.22 | Tolerated | 4.32 | 12 | 0.2831 | 0.4801 | 0 | 1 | -0.4 | 30.03 | ||||||||||||||||||||||||
| c.112C>A | P38T 2D AIThe SynGAP1 missense variant P38T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.433285 | Uncertain | 0.344 | 0.791 | 0.375 | -3.248 | Likely Benign | 0.116 | Likely Benign | Likely Benign | 0.114 | Likely Benign | -1.91 | Neutral | 0.909 | Possibly Damaging | 0.901 | Possibly Damaging | 4.06 | Benign | 0.00 | Affected | 0.1993 | 0.6717 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.1130T>A | M377K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M377K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are Rosetta and Foldetta. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely benign outcome, while premPS remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Thus, the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.675549 | Disordered | 0.431183 | Uncertain | 0.324 | 0.884 | 0.625 | -3.718 | Likely Benign | 0.226 | Likely Benign | Likely Benign | 0.20 | Likely Benign | 0.2 | 4.44 | Destabilizing | 2.32 | Destabilizing | 0.62 | Ambiguous | 0.440 | Likely Benign | -0.26 | Neutral | 0.002 | Benign | 0.003 | Benign | 5.46 | Benign | 0.07 | Tolerated | 0.2423 | 0.1671 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1139G>A | G380E 2D AIThe SynGAP1 missense variant G380E has no ClinVar record and is not reported in gnomAD. Prediction tools that agree on a benign effect include Rosetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Foldetta, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy methods give mixed results: AlphaMissense‑Optimized classifies the variant as benign, Foldetta predicts a pathogenic impact on protein stability, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split. Overall, the majority of tools (7 benign vs. 6 pathogenic) lean toward a benign interpretation, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Thus, based on current computational predictions, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.432982 | Uncertain | 0.316 | 0.939 | 0.750 | -9.334 | Likely Pathogenic | 0.617 | Likely Pathogenic | Likely Benign | 6.25 | Destabilizing | 5.1 | -0.28 | Likely Benign | 2.99 | Destabilizing | 0.24 | Likely Benign | 0.582 | Likely Pathogenic | -0.86 | Neutral | 0.056 | Benign | 0.010 | Benign | 2.53 | Benign | 0.03 | Affected | 0.1585 | 0.3741 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.113C>A | P38Q 2D AIThe SynGAP1 missense variant P38Q is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P38Q, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.433285 | Uncertain | 0.344 | 0.791 | 0.375 | -3.520 | Likely Benign | 0.183 | Likely Benign | Likely Benign | 0.129 | Likely Benign | -2.16 | Neutral | 0.989 | Probably Damaging | 0.975 | Probably Damaging | 4.00 | Benign | 0.00 | Affected | 0.1737 | 0.5521 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||||||||||||
| c.113C>G | P38R 2D AIThe SynGAP1 missense variant P38R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized returns Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability prediction is available. Overall, the majority of evidence points to a benign effect for P38R, and this conclusion is not contradicted by any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.497853 | Structured | 0.433285 | Uncertain | 0.344 | 0.791 | 0.375 | -1.799 | Likely Benign | 0.314 | Likely Benign | Likely Benign | 0.168 | Likely Benign | -2.29 | Neutral | 0.989 | Probably Damaging | 0.975 | Probably Damaging | 4.00 | Benign | 0.00 | Affected | 0.1620 | 0.4060 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.1142G>A | G381E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G381E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, Foldetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Two tools give uncertain results: Rosetta and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. With seven pathogenic versus four benign predictions and two high‑accuracy tools supporting pathogenicity, the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.724957 | Disordered | 0.431692 | Uncertain | 0.301 | 0.951 | 0.750 | -9.360 | Likely Pathogenic | 0.540 | Ambiguous | Likely Benign | 5.52 | Destabilizing | 1.1 | 0.53 | Ambiguous | 3.03 | Destabilizing | 0.24 | Likely Benign | 0.588 | Likely Pathogenic | -0.71 | Neutral | 0.985 | Probably Damaging | 0.720 | Possibly Damaging | 1.32 | Pathogenic | 0.11 | Tolerated | 0.1554 | 0.3735 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1142G>C | G381A 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G381A is reported in gnomAD (variant ID 6-33438047‑G‑C) but has no ClinVar entry. Functional prediction tools fall into two groups: benign predictions come from premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen‑2 HumVar; pathogenic predictions come from REVEL, FoldX, Rosetta, polyPhen‑2 HumDiv, FATHMM, and the SGM‑Consensus score. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely benign, while Foldetta (combining FoldX‑MD and Rosetta stability outputs) indicates a pathogenic effect. No prediction or stability result is missing or inconclusive. Overall, the majority of tools suggest a benign effect, and the high‑accuracy consensus leans toward benign, though Foldetta’s pathogenic signal introduces uncertainty. The variant is most likely benign based on the collective predictions, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.724957 | Disordered | 0.431692 | Uncertain | 0.301 | 0.951 | 0.750 | 6-33438047-G-C | 1 | 6.23e-7 | -6.266 | Likely Benign | 0.103 | Likely Benign | Likely Benign | 3.97 | Destabilizing | 0.7 | 2.05 | Destabilizing | 3.01 | Destabilizing | 0.06 | Likely Benign | 0.507 | Likely Pathogenic | -0.63 | Neutral | 0.718 | Possibly Damaging | 0.332 | Benign | 1.33 | Pathogenic | 0.52 | Tolerated | 4.32 | 9 | 0.3809 | 0.4770 | 0 | 1 | 2.2 | 14.03 | ||||||||||||||||||||||||
| c.1148G>A | G383E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G383E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and FATHMM. Tools that predict a pathogenic effect are FoldX, Rosetta, SIFT, ESM1b, AlphaMissense‑Default, and the protein‑folding stability method Foldetta. High‑accuracy assessments give a benign result from AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default (pathogenic), ESM1b (pathogenic), FATHMM (benign), and PROVEAN (benign), is inconclusive (tie). Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a pathogenic effect. Overall, the majority of predictions (seven pathogenic vs. six benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.728858 | Disordered | 0.429104 | Uncertain | 0.296 | 0.949 | 0.750 | -9.535 | Likely Pathogenic | 0.570 | Likely Pathogenic | Likely Benign | 4.09 | Destabilizing | 2.4 | 2.62 | Destabilizing | 3.36 | Destabilizing | 0.39 | Likely Benign | 0.308 | Likely Benign | -0.78 | Neutral | 0.000 | Benign | 0.002 | Benign | 4.10 | Benign | 0.01 | Affected | 0.1582 | 0.3741 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.1159G>A | G387S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 G387S missense variant is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (variant ID 6‑33438064‑G‑A). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are FoldX and FATHMM; Rosetta is uncertain. The high‑accuracy consensus (SGM‑Consensus) is derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN and is therefore likely benign. AlphaMissense‑Optimized itself predicts benign. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts a pathogenic effect. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar classification (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.642678 | Disordered | 0.422910 | Uncertain | 0.293 | 0.861 | 0.750 | 6-33438064-G-A | -4.674 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 2.37 | Destabilizing | 0.5 | 1.94 | Ambiguous | 2.16 | Destabilizing | 0.12 | Likely Benign | 0.359 | Likely Benign | -0.12 | Neutral | 0.000 | Benign | 0.002 | Benign | 1.33 | Pathogenic | 0.13 | Tolerated | 4.32 | 3 | 0.3051 | 0.4724 | 0 | 1 | -0.4 | 30.03 | ||||||||||||||||||||||||||
| c.115T>C | Y39H 2D AIThe SynGAP1 missense variant Y39H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.425610 | Structured | 0.432876 | Uncertain | 0.343 | 0.787 | 0.375 | -3.166 | Likely Benign | 0.267 | Likely Benign | Likely Benign | 0.083 | Likely Benign | -1.08 | Neutral | 0.824 | Possibly Damaging | 0.775 | Possibly Damaging | 4.02 | Benign | 0.00 | Affected | 0.2569 | 0.0843 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||||||||||||
| c.1160G>C | G387A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G387A is reported in gnomAD (variant ID 6‑33438065‑G‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only FATHMM predicts it as pathogenic. High‑accuracy assessments reinforce the benign consensus: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Benign”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is inconclusive (treated as unavailable). No other tools provide decisive evidence. Based on the collective predictions, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.642678 | Disordered | 0.422910 | Uncertain | 0.293 | 0.861 | 0.750 | 6-33438065-G-C | 3 | 1.87e-6 | -6.313 | Likely Benign | 0.104 | Likely Benign | Likely Benign | 1.95 | Ambiguous | 0.1 | 1.68 | Ambiguous | 1.82 | Ambiguous | -0.02 | Likely Benign | 0.300 | Likely Benign | -0.29 | Neutral | 0.007 | Benign | 0.010 | Benign | 1.33 | Pathogenic | 0.06 | Tolerated | 4.32 | 3 | 0.4149 | 0.4576 | 0 | 1 | 2.2 | 14.03 | ||||||||||||||||||||||||
| c.1162G>A | G388S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant G388S is catalogued in gnomAD (ID 6‑33438067‑G‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that converge on a pathogenic interpretation are FoldX, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. The high‑accuracy AlphaMissense‑Optimized score is benign, while the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also reports a likely benign outcome. In contrast, the protein‑folding stability predictor Foldetta, which integrates FoldX‑MD and Rosetta outputs, indicates a pathogenic effect. No prediction is available from Rosetta alone, and the SGM‑Consensus result aligns with the benign consensus. Overall, the majority of evidence points to a benign impact, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.736850 | Disordered | 0.420316 | Uncertain | 0.319 | 0.827 | 0.750 | 6-33438067-G-A | 1 | 6.21e-7 | -5.036 | Likely Benign | 0.089 | Likely Benign | Likely Benign | 3.42 | Destabilizing | 3.5 | 1.69 | Ambiguous | 2.56 | Destabilizing | 0.18 | Likely Benign | 0.446 | Likely Benign | -0.52 | Neutral | 0.980 | Probably Damaging | 0.968 | Probably Damaging | 1.33 | Pathogenic | 0.05 | Affected | 4.32 | 3 | 0.2901 | 0.4985 | 0 | 1 | -0.4 | 30.03 | ||||||||||||||||||||||||
| c.1169G>A | G390E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G390E is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that classify the variant as benign include premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Those that predict pathogenicity are REVEL, FoldX, Rosetta, Foldetta, SIFT, FATHMM, and AlphaMissense‑Default. A high‑accuracy assessment shows AlphaMissense‑Optimized as benign, the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of predictions support a pathogenic effect, and this aligns with the ClinVar designation of uncertain significance rather than contradicting it. Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.626927 | Disordered | 0.413274 | Uncertain | 0.304 | 0.763 | 0.875 | Uncertain | 1 | -7.913 | In-Between | 0.646 | Likely Pathogenic | Likely Benign | 2.61 | Destabilizing | 0.9 | 4.28 | Destabilizing | 3.45 | Destabilizing | 0.47 | Likely Benign | 0.575 | Likely Pathogenic | -0.87 | Neutral | 0.276 | Benign | 0.045 | Benign | 1.32 | Pathogenic | 0.05 | Affected | 4.32 | 8 | 0.1595 | 0.4309 | 0 | -2 | -3.1 | 72.06 | 241.5 | -108.4 | 0.6 | 0.5 | -0.1 | 0.1 | Uncertain | Gly390 is located in the Gly-rich Ω loop (res. Pro364-Pro398) between two anti-parallel β sheet strands (res. Thr359-Pro364 and res. Ala399-Ile411). The Ω loop is assumed to directly interact with the membrane, and it is observed to move arbitrarily throughout the WT solvent simulations. This loop potentially plays a crucial role in the SynGAP-membrane complex association, stability, and dynamics. However, this aspect cannot be fully addressed through solvent simulations alone.Ω loops are known to play significant roles in protein functions that require flexibility, and so they are rich in glycine residues, prolines, and to a lesser extent, small hydrophilic residues to ensure maximum flexibility. Thus, the variant’s Glu390 may not be as well tolerated in the Ω loop. Additionally, the carboxylate group of Glu390 occasionally forms H-bonds with other loop residues in the variant simulations. The interaction between the acidic carboxylate side chain and the acidic membrane lipids may further influence the SynGAP-membrane complex. However, since the effects on the Gly-rich Ω loop dynamics can only be well studied through the SynGAP-membrane complex, no definite conclusions can be drawn. | ||||||||||||||||||
| c.116A>G | Y39C 2D AIThe SynGAP1 missense variant Y39C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available, so it does not influence the overall assessment. Overall, the majority of evidence points to the variant being most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.425610 | Structured | 0.432876 | Uncertain | 0.343 | 0.787 | 0.375 | -3.109 | Likely Benign | 0.123 | Likely Benign | Likely Benign | 0.203 | Likely Benign | -2.00 | Neutral | 0.943 | Possibly Damaging | 0.941 | Probably Damaging | 3.96 | Benign | 0.00 | Affected | 0.3007 | 0.2849 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||||||||||||
| c.1174A>G | K392E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and the SGM‑Consensus score. Tools that predict a pathogenic effect are REVEL, SIFT, and AlphaMissense‑Default. The high‑accuracy AlphaMissense‑Optimized score is uncertain, while the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) and Foldetta (combining FoldX‑MD and Rosetta) both indicate a benign outcome. Overall, the majority of evidence supports a benign classification, and this is consistent with the lack of ClinVar annotation; there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -4.392 | Likely Benign | 0.850 | Likely Pathogenic | Ambiguous | 0.09 | Likely Benign | 0.0 | -0.04 | Likely Benign | 0.03 | Likely Benign | 0.28 | Likely Benign | 0.529 | Likely Pathogenic | -1.92 | Neutral | 0.276 | Benign | 0.083 | Benign | 4.60 | Benign | 0.02 | Affected | 0.4812 | 0.1916 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1175A>C | K392T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K392T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Rosetta, Foldetta, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are REVEL, PROVEAN, and SIFT. Two tools—FoldX and AlphaMissense‑Default—return uncertain results and are treated as unavailable. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) resolves to benign; and Foldetta, a protein‑folding stability method, also predicts benign. No ClinVar entry exists to contradict these predictions. Based on the collective evidence, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -3.224 | Likely Benign | 0.481 | Ambiguous | Likely Benign | 0.52 | Ambiguous | 0.1 | -0.29 | Likely Benign | 0.12 | Likely Benign | 0.05 | Likely Benign | 0.512 | Likely Pathogenic | -3.14 | Deleterious | 0.276 | Benign | 0.045 | Benign | 4.60 | Benign | 0.02 | Affected | 0.2807 | 0.4053 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||
| c.1175A>T | K392M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K392M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, ESM1b, and FATHMM, whereas those that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Default. The remaining tools—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—return uncertain or inconclusive results. High‑accuracy methods give no definitive signal: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable, and Foldetta is uncertain. Overall, the majority of available predictions (six pathogenic vs. three benign) lean toward a pathogenic impact. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.541878 | Disordered | 0.405672 | Uncertain | 0.319 | 0.702 | 0.750 | -3.856 | Likely Benign | 0.788 | Likely Pathogenic | Ambiguous | 0.52 | Ambiguous | 0.1 | 0.67 | Ambiguous | 0.60 | Ambiguous | -0.09 | Likely Benign | 0.665 | Likely Pathogenic | -3.24 | Deleterious | 0.952 | Possibly Damaging | 0.496 | Possibly Damaging | 4.59 | Benign | 0.00 | Affected | 0.1799 | 0.4941 | 0 | -1 | 5.8 | 3.02 | ||||||||||||||||||||||||||||||
| c.1181A>C | K394T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K394T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Those that predict a pathogenic effect are PROVEAN, SIFT, and AlphaMissense‑Default. Predictions that are uncertain or inconclusive are FoldX, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (a majority vote among AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is inconclusive due to a 2‑to‑2 split, and Foldetta is also inconclusive. Overall, the balance of evidence (five benign versus three pathogenic predictions) suggests the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.505461 | Disordered | 0.399336 | Uncertain | 0.387 | 0.634 | 0.625 | -6.487 | Likely Benign | 0.599 | Likely Pathogenic | Likely Benign | 0.50 | Ambiguous | 0.1 | 2.46 | Destabilizing | 1.48 | Ambiguous | 0.57 | Ambiguous | 0.482 | Likely Benign | -3.35 | Deleterious | 0.247 | Benign | 0.166 | Benign | 4.61 | Benign | 0.01 | Affected | 0.2727 | 0.4453 | 0 | -1 | 3.2 | -27.07 | ||||||||||||||||||||||||||||||
| c.1184G>A | G395E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G395E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Only AlphaMissense‑Default predicts a pathogenic outcome, while FoldX and Rosetta give uncertain results. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (majority vote) predicts benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts benign. Taken together, the majority of evidence points to a benign effect. There is no ClinVar entry to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.513880 | Disordered | 0.396199 | Uncertain | 0.474 | 0.601 | 0.500 | -6.350 | Likely Benign | 0.594 | Likely Pathogenic | Likely Benign | 1.60 | Ambiguous | 0.6 | -0.88 | Ambiguous | 0.36 | Likely Benign | 0.30 | Likely Benign | 0.310 | Likely Benign | -1.81 | Neutral | 0.037 | Benign | 0.010 | Benign | 4.25 | Benign | 0.12 | Tolerated | 0.1328 | 0.4089 | 0 | -2 | -3.1 | 72.06 | |||||||||||||||||||||||||||||
| c.1189T>A | C397S 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a benign effect: REVEL, FoldX, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, PROVEAN, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as benign, while Rosetta remains uncertain. When predictions are grouped, the benign category contains all available evidence and the pathogenic category is empty. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports a benign effect. Consequently, the variant is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -2.324 | Likely Benign | 0.178 | Likely Benign | Likely Benign | 0.37 | Likely Benign | 0.1 | 0.59 | Ambiguous | 0.48 | Likely Benign | 0.43 | Likely Benign | 0.174 | Likely Benign | -0.01 | Neutral | 0.276 | Benign | 0.066 | Benign | 4.68 | Benign | 0.53 | Tolerated | 0.5221 | 0.2474 | Weaken | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||
| c.1190G>A | C397Y 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are FoldX, Rosetta, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and the combined Foldetta method. Uncertain results come from AlphaMissense‑Default and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta as pathogenic. Overall, the majority of tools and the high‑accuracy consensus lean toward a benign interpretation, and this does not contradict any ClinVar classification (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -7.213 | In-Between | 0.397 | Ambiguous | Likely Benign | 2.01 | Destabilizing | 2.3 | 8.64 | Destabilizing | 5.33 | Destabilizing | 0.12 | Likely Benign | 0.455 | Likely Benign | -1.82 | Neutral | 0.952 | Possibly Damaging | 0.497 | Possibly Damaging | 4.64 | Benign | 0.07 | Tolerated | 0.1299 | 0.5084 | 0 | -2 | -3.8 | 60.04 | ||||||||||||||||||||||||||||||
| c.1190G>C | C397S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C397S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). In silico prediction tools that assess pathogenicity largely agree on a benign outcome: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict benign. No tool predicts pathogenicity; Rosetta’s assessment is uncertain and therefore treated as unavailable. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is benign. Consequently, the variant is most likely benign based on the collective predictions, and this conclusion does not contradict any ClinVar status (none is assigned). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.429200 | Structured | 0.395774 | Uncertain | 0.778 | 0.551 | 0.250 | -2.324 | Likely Benign | 0.178 | Likely Benign | Likely Benign | 0.37 | Likely Benign | 0.1 | 0.59 | Ambiguous | 0.48 | Likely Benign | 0.43 | Likely Benign | 0.253 | Likely Benign | -0.01 | Neutral | 0.276 | Benign | 0.066 | Benign | 4.68 | Benign | 0.53 | Tolerated | 0.5221 | 0.2474 | Weaken | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||
| c.1192C>A | P398T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant P398T is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, and SIFT. The remaining tools (Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain results. High‑accuracy methods give a benign consensus: AlphaMissense‑Optimized is benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is benign, and Foldetta is uncertain. Overall, the majority of high‑confidence predictions lean toward a benign impact, although several other predictors indicate pathogenicity. There is no conflict with ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | -6.670 | Likely Benign | 0.536 | Ambiguous | Likely Benign | 2.11 | Destabilizing | 0.4 | 1.57 | Ambiguous | 1.84 | Ambiguous | 0.78 | Ambiguous | 0.608 | Likely Pathogenic | -5.70 | Deleterious | 0.816 | Possibly Damaging | 0.307 | Benign | 5.51 | Benign | 0.01 | Affected | 0.1671 | 0.6607 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1193C>A | P398Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P398Q has no ClinVar record (ClinVar ID None) and is not reported in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM, while the majority of tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, AlphaMissense‑Default) predict a pathogenic impact. Two tools report uncertainty: ESM1b and AlphaMissense‑Optimized. High‑accuracy assessments further support pathogenicity: the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a pathogenic verdict, and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. No prediction or stability result is missing or inconclusive. Consequently, the variant is most likely pathogenic based on the collective evidence, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | -7.402 | In-Between | 0.792 | Likely Pathogenic | Ambiguous | 2.13 | Destabilizing | 0.1 | 2.01 | Destabilizing | 2.07 | Destabilizing | 1.01 | Destabilizing | 0.719 | Likely Pathogenic | -5.57 | Deleterious | 0.996 | Probably Damaging | 0.724 | Possibly Damaging | 5.48 | Benign | 0.00 | Affected | 0.1522 | 0.5176 | 0 | -1 | -1.9 | 31.01 | ||||||||||||||||||||||||||||||
| c.1193C>G | P398R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P398R variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that classify it as benign include only FATHMM. All other evaluated predictors—SGM‑Consensus, REVEL, FoldX, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—indicate a pathogenic effect, while Rosetta, premPS, and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Based on the preponderance of pathogenic predictions and the high‑accuracy consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.436924 | Structured | 0.401041 | Uncertain | 0.891 | 0.525 | 0.250 | -9.575 | Likely Pathogenic | 0.889 | Likely Pathogenic | Ambiguous | 3.01 | Destabilizing | 0.5 | 1.35 | Ambiguous | 2.18 | Destabilizing | 0.98 | Ambiguous | 0.755 | Likely Pathogenic | -6.55 | Deleterious | 0.988 | Probably Damaging | 0.724 | Possibly Damaging | 5.49 | Benign | 0.00 | Affected | 0.1479 | 0.3472 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1196C>A | A399D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A399D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Those that agree on a pathogenic effect are REVEL, FoldX, SIFT, ESM1b, and AlphaMissense‑Default. The remaining tools—Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized—return uncertain or inconclusive results. High‑accuracy methods give no definitive verdict: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable, and Foldetta is uncertain. Overall, the majority of available predictions (five pathogenic vs. four benign) lean toward pathogenicity. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.394753 | Structured | 0.407674 | Uncertain | 0.939 | 0.490 | 0.125 | -10.441 | Likely Pathogenic | 0.943 | Likely Pathogenic | Ambiguous | 2.45 | Destabilizing | 0.2 | 1.17 | Ambiguous | 1.81 | Ambiguous | 0.91 | Ambiguous | 0.525 | Likely Pathogenic | -1.84 | Neutral | 0.421 | Benign | 0.054 | Benign | 5.38 | Benign | 0.05 | Affected | 0.1533 | 0.1760 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1196C>T | A399V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A399V is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). No tool predicts a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, while Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Overall, the evidence overwhelmingly supports a benign classification, and this is consistent with the lack of ClinVar reporting. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | C2 | 0.394753 | Structured | 0.407674 | Uncertain | 0.939 | 0.490 | 0.125 | -4.943 | Likely Benign | 0.170 | Likely Benign | Likely Benign | 0.49 | Likely Benign | 0.2 | 0.56 | Ambiguous | 0.53 | Ambiguous | 0.22 | Likely Benign | 0.305 | Likely Benign | -1.18 | Neutral | 0.421 | Benign | 0.073 | Benign | 5.37 | Benign | 0.24 | Tolerated | 0.1145 | 0.6807 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1199T>C | V400A 2D 3DClick to see structure in 3D Viewer AISynGAP1 V400A is not reported in ClinVar and is absent from gnomAD. Consensus from standard predictors shows a split: benign calls from REVEL, polyPhen‑2 (HumDiv and HumVar) and FATHMM, while pathogenic calls come from FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT and AlphaMissense‑Default. Two high‑accuracy tools give a clear signal: AlphaMissense‑Optimized is uncertain, but the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta also predicts pathogenic. With most evidence pointing to deleterious effects, the variant is most likely pathogenic, and this assessment does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.398279 | Structured | 0.415488 | Uncertain | 0.951 | 0.451 | 0.000 | -7.564 | In-Between | 0.871 | Likely Pathogenic | Ambiguous | 3.14 | Destabilizing | 0.1 | 3.12 | Destabilizing | 3.13 | Destabilizing | 2.29 | Destabilizing | 0.479 | Likely Benign | -3.12 | Deleterious | 0.435 | Benign | 0.049 | Benign | 5.32 | Benign | 0.01 | Affected | 0.3291 | 0.2767 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||||||||
| c.119A>C | D40A 2D AIThe SynGAP1 missense variant D40A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign, while the single pathogenic call comes from SIFT. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign consensus. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus likewise indicates likely benign; Foldetta results are not available. Taken together, the majority of evidence supports a benign interpretation, and this is consistent with the absence of any ClinVar pathogenic annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.384043 | Structured | 0.432002 | Uncertain | 0.319 | 0.769 | 0.375 | -3.630 | Likely Benign | 0.339 | Likely Benign | Likely Benign | 0.122 | Likely Benign | -1.22 | Neutral | 0.006 | Benign | 0.023 | Benign | 4.05 | Benign | 0.00 | Affected | 0.4299 | 0.7993 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||||
| c.11C>G | S4C 2D AIThe SynGAP1 missense variant S4C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for S4C, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.608892 | Disordered | 0.547364 | Binding | 0.390 | 0.924 | 0.750 | -5.210 | Likely Benign | 0.124 | Likely Benign | Likely Benign | 0.106 | Likely Benign | 0.41 | Neutral | 0.880 | Possibly Damaging | 0.700 | Possibly Damaging | 4.11 | Benign | 0.00 | Affected | 0.0976 | 0.6129 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||||||||||
| c.1202G>C | R401P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R401P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while the remaining twelve tools—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. The high‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. No predictions are missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.314870 | Structured | 0.424277 | Uncertain | 0.961 | 0.419 | 0.000 | -14.090 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 9.69 | Destabilizing | 0.3 | 8.07 | Destabilizing | 8.88 | Destabilizing | 0.77 | Ambiguous | 0.892 | Likely Pathogenic | -6.45 | Deleterious | 0.999 | Probably Damaging | 0.995 | Probably Damaging | 5.42 | Benign | 0.00 | Affected | 0.2265 | 0.4064 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1207A>G | K403E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K403E missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Predictions from FoldX, Rosetta, Foldetta, and premPS are uncertain and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact for K403E, and this conclusion does not contradict the ClinVar status, which currently has no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.275179 | Structured | 0.424920 | Uncertain | 0.960 | 0.372 | 0.000 | -15.279 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 1.01 | Ambiguous | 0.1 | 0.79 | Ambiguous | 0.90 | Ambiguous | 0.70 | Ambiguous | 0.445 | Likely Benign | -3.62 | Deleterious | 0.998 | Probably Damaging | 0.981 | Probably Damaging | 3.79 | Benign | 0.04 | Affected | 0.3650 | 0.1715 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1208A>C | K403T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K403T missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are limited to FATHMM, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: SGM‑Consensus (Likely Pathogenic), REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts pathogenic; SGM‑Consensus predicts likely pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is uncertain. Other stability predictors (FoldX, Rosetta, premPS) are also uncertain. Overall, the consensus of the majority of tools indicates a pathogenic effect. This conclusion is not contradicted by ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.275179 | Structured | 0.424920 | Uncertain | 0.960 | 0.372 | 0.000 | -13.135 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 1.41 | Ambiguous | 0.1 | 0.59 | Ambiguous | 1.00 | Ambiguous | 0.67 | Ambiguous | 0.522 | Likely Pathogenic | -5.43 | Deleterious | 0.999 | Probably Damaging | 1.000 | Probably Damaging | 3.73 | Benign | 0.01 | Affected | 0.1861 | 0.4230 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1211C>A | A404E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A404E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. In contrast, the majority of tools predict a pathogenic impact: FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate pathogenicity, and the SGM‑Consensus score is “Likely Pathogenic.” High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic; the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.232838 | Structured | 0.415505 | Uncertain | 0.965 | 0.355 | 0.000 | -13.639 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 2.16 | Destabilizing | 0.1 | 3.04 | Destabilizing | 2.60 | Destabilizing | 1.51 | Destabilizing | 0.163 | Likely Benign | -2.77 | Deleterious | 0.179 | Benign | 0.033 | Benign | 4.02 | Benign | 0.02 | Affected | 0.1423 | 0.2383 | 0 | -1 | -5.3 | 58.04 | |||||||||||||||||||||||||||||
| c.1211C>T | A404V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A404V missense variant is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438116‑C‑T). Prediction tools that agree on a benign effect include REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and ESM1b. Four tools (FoldX, Rosetta, AlphaMissense‑Default, Foldetta) give uncertain or inconclusive results. High‑accuracy methods give mixed evidence: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta remains uncertain. Overall, the majority of predictions lean toward a benign impact, though a key consensus method suggests pathogenicity, leaving the variant’s effect unresolved. **The variant is most likely benign based on the prevailing predictions, and this assessment does not contradict ClinVar status, as no ClinVar classification exists.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.232838 | Structured | 0.415505 | Uncertain | 0.965 | 0.355 | 0.000 | 6-33438116-C-T | 1 | 6.20e-7 | -8.219 | Likely Pathogenic | 0.343 | Ambiguous | Likely Benign | 0.56 | Ambiguous | 0.3 | -1.55 | Ambiguous | -0.50 | Ambiguous | -0.05 | Likely Benign | 0.118 | Likely Benign | -2.96 | Deleterious | 0.345 | Benign | 0.018 | Benign | 4.20 | Benign | 0.47 | Tolerated | 3.38 | 28 | 0.1288 | 0.6842 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||
| c.1213C>A | R405S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R405S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: benign predictions come from REVEL and FATHMM, whereas pathogenic predictions are made by FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts Pathogenic; the SGM‑Consensus itself is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts Pathogenic. Taken together, the overwhelming majority of evidence indicates that R405S is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.250310 | Structured | 0.404888 | Uncertain | 0.949 | 0.315 | 0.000 | -11.326 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 2.66 | Destabilizing | 0.5 | 2.08 | Destabilizing | 2.37 | Destabilizing | 1.22 | Destabilizing | 0.405 | Likely Benign | -5.40 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.67 | Benign | 0.05 | Affected | 0.2829 | 0.4646 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||
| c.1216T>C | Y406H 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Y406H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of other in silico predictors—FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—consistently classify the substitution as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely pathogenic outcome, and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a pathogenic effect. Taken together, the preponderance of evidence indicates that Y406H is most likely pathogenic, and this conclusion does not conflict with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.179055 | Structured | 0.393707 | Uncertain | 0.946 | 0.291 | 0.000 | -9.015 | Likely Pathogenic | 0.858 | Likely Pathogenic | Ambiguous | 2.13 | Destabilizing | 0.1 | 2.15 | Destabilizing | 2.14 | Destabilizing | 1.03 | Destabilizing | 0.239 | Likely Benign | -4.21 | Deleterious | 0.997 | Probably Damaging | 0.966 | Probably Damaging | 3.80 | Benign | 0.03 | Affected | 0.2666 | 0.0811 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.1217A>G | Y406C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y406C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: FoldX, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome, and the Foldetta stability assessment (combining FoldX‑MD and Rosetta) predicts a destabilizing, pathogenic effect. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while both SGM‑Consensus and Foldetta are pathogenic. Overall, the preponderance of evidence points to a pathogenic effect for Y406C, and this conclusion does not contradict the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.179055 | Structured | 0.393707 | Uncertain | 0.946 | 0.291 | 0.000 | -9.954 | Likely Pathogenic | 0.810 | Likely Pathogenic | Ambiguous | 2.47 | Destabilizing | 0.2 | 3.22 | Destabilizing | 2.85 | Destabilizing | 1.27 | Destabilizing | 0.229 | Likely Benign | -6.72 | Deleterious | 0.998 | Probably Damaging | 0.851 | Possibly Damaging | 3.78 | Benign | 0.01 | Affected | 0.3026 | 0.2861 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||
| c.121C>A | R41S 2D AIThe SynGAP1 missense variant R41S is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default is uncertain, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign; Foldetta results are unavailable. Overall, the majority of evidence points to a benign impact for R41S, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.311707 | Structured | 0.431757 | Uncertain | 0.344 | 0.765 | 0.375 | -3.367 | Likely Benign | 0.393 | Ambiguous | Likely Benign | 0.130 | Likely Benign | -0.22 | Neutral | 0.686 | Possibly Damaging | 0.735 | Possibly Damaging | 4.24 | Benign | 0.00 | Affected | 0.3190 | 0.5013 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||||||||||||
| c.1220A>C | Q407P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q407P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact; premPS is uncertain and is not counted. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. Based on the consensus of these predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.109221 | Structured | 0.382522 | Uncertain | 0.916 | 0.271 | 0.000 | -13.578 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 3.04 | Destabilizing | 0.5 | 2.07 | Destabilizing | 2.56 | Destabilizing | 0.88 | Ambiguous | 0.515 | Likely Pathogenic | -5.40 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.88 | Benign | 0.02 | Affected | 0.1786 | 0.4352 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||||
| c.1221G>C | Q407H 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q407H (C2 domain) has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict pathogenicity: premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain; the SGM‑Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, is uncertain. No evidence from FoldX, Rosetta, or Foldetta supports a benign outcome. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.109221 | Structured | 0.382522 | Uncertain | 0.916 | 0.271 | 0.000 | -10.526 | Likely Pathogenic | 0.830 | Likely Pathogenic | Ambiguous | 0.59 | Ambiguous | 0.0 | 0.61 | Ambiguous | 0.60 | Ambiguous | 1.10 | Destabilizing | 0.206 | Likely Benign | -4.51 | Deleterious | 0.982 | Probably Damaging | 0.947 | Probably Damaging | 3.88 | Benign | 0.01 | Affected | 3.38 | 28 | 0.1205 | 0.3154 | 0 | 3 | 0.3 | 9.01 | |||||||||||||||||||||||||||
| c.1221G>T | Q407H 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q407H is listed in ClinVar with an uncertain significance (ClinVar ID 2772184.0) and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and FATHMM, while pathogenic predictions are made by premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Predictions marked as uncertain include FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for Q407H. This conclusion does not conflict with the ClinVar designation of uncertain significance, which remains unresolved pending further evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.109221 | Structured | 0.382522 | Uncertain | 0.916 | 0.271 | 0.000 | Uncertain | 1 | -10.526 | Likely Pathogenic | 0.830 | Likely Pathogenic | Ambiguous | 0.59 | Ambiguous | 0.0 | 0.61 | Ambiguous | 0.60 | Ambiguous | 1.10 | Destabilizing | 0.206 | Likely Benign | -4.51 | Deleterious | 0.982 | Probably Damaging | 0.947 | Probably Damaging | 3.88 | Benign | 0.01 | Affected | 3.38 | 28 | 0.1205 | 0.3154 | 0 | 3 | 0.3 | 9.01 | |||||||||||||||||||||||||
| c.1222A>C | T408P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T408P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. FoldX, premPS, and Foldetta are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a tie and thus unavailable; Foldetta remains uncertain. Overall, the majority of available predictions (six pathogenic vs. four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | C2 | 0.161087 | Structured | 0.370935 | Uncertain | 0.907 | 0.239 | 0.000 | -10.384 | Likely Pathogenic | 0.230 | Likely Benign | Likely Benign | 1.08 | Ambiguous | 0.3 | 2.27 | Destabilizing | 1.68 | Ambiguous | 0.73 | Ambiguous | 0.323 | Likely Benign | -4.19 | Deleterious | 0.998 | Probably Damaging | 0.963 | Probably Damaging | 4.07 | Benign | 0.05 | Affected | 0.1985 | 0.5779 | 0 | -1 | -0.9 | -3.99 | ||||||||||||||||||||||||||||||
| c.1223C>A | T408K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T408K is not listed in ClinVar (ClinVar ID None) and has no reported allele in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, SIFT, FATHMM, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default; premPS remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labeling it likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicting a benign effect. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.161087 | Structured | 0.370935 | Uncertain | 0.907 | 0.239 | 0.000 | -11.841 | Likely Pathogenic | 0.756 | Likely Pathogenic | Likely Benign | -0.44 | Likely Benign | 0.2 | 0.14 | Likely Benign | -0.15 | Likely Benign | 0.88 | Ambiguous | 0.131 | Likely Benign | -3.79 | Deleterious | 0.851 | Possibly Damaging | 0.163 | Benign | 4.17 | Benign | 0.15 | Tolerated | 0.1205 | 0.3384 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||
| c.1226T>A | M409K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M409K has no ClinVar record and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, Rosetta, SIFT, and FATHMM, while pathogenic calls arise from premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default. Uncertain results are reported by FoldX, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized is inconclusive; the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, predicts pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is also inconclusive. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion does not conflict with the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | C2 | 0.150080 | Structured | 0.360643 | Uncertain | 0.884 | 0.219 | 0.000 | -13.618 | Likely Pathogenic | 0.927 | Likely Pathogenic | Ambiguous | 0.93 | Ambiguous | 0.3 | 0.29 | Likely Benign | 0.61 | Ambiguous | 1.45 | Destabilizing | 0.490 | Likely Benign | -4.26 | Deleterious | 0.769 | Possibly Damaging | 0.750 | Possibly Damaging | 4.18 | Benign | 0.40 | Tolerated | 0.1318 | 0.0656 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1228A>C | S410R 2D 3DClick to see structure in 3D Viewer AIClinVar contains no record for the SynGAP1 S410R variant, and it is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, Foldetta predicts a benign impact, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Taken together, the majority of evidence points to a benign outcome. This conclusion does not contradict ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -8.203 | Likely Pathogenic | 0.941 | Likely Pathogenic | Ambiguous | -0.67 | Ambiguous | 0.2 | 0.56 | Ambiguous | -0.06 | Likely Benign | 0.62 | Ambiguous | 0.242 | Likely Benign | -2.47 | Neutral | 0.871 | Possibly Damaging | 0.298 | Benign | 4.20 | Benign | 0.40 | Tolerated | 0.0955 | 0.3927 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||
| c.1228A>T | S410C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S410C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict it as pathogenic are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar; ESM1b is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or folding stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -7.552 | In-Between | 0.144 | Likely Benign | Likely Benign | -0.24 | Likely Benign | 0.1 | 0.31 | Likely Benign | 0.04 | Likely Benign | 0.22 | Likely Benign | 0.230 | Likely Benign | -3.10 | Deleterious | 0.993 | Probably Damaging | 0.536 | Possibly Damaging | 4.12 | Benign | 0.11 | Tolerated | 0.1041 | 0.6543 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||||
| c.122G>A | R41H 2D AIThe SynGAP1 missense variant R41H is reported in gnomAD (variant ID 6-33423531‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: the majority (REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a benign effect, while a minority (polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT) predict pathogenicity. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized classifies the variant as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports it as Likely Benign. No Foldetta stability data are available, so it does not influence the overall assessment. Based on the preponderance of evidence from both general and high‑accuracy predictors, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.311707 | Structured | 0.431757 | Uncertain | 0.344 | 0.765 | 0.375 | 6-33423531-G-A | 6 | 3.72e-6 | -4.425 | Likely Benign | 0.106 | Likely Benign | Likely Benign | 0.059 | Likely Benign | -0.74 | Neutral | 0.976 | Probably Damaging | 0.848 | Possibly Damaging | 4.17 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3388 | 0.3065 | 0 | 2 | 1.3 | -19.05 | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||||||||||||||||||
| c.122G>C | R41P 2D AIThe SynGAP1 missense variant R41P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for R41P, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.311707 | Structured | 0.431757 | Uncertain | 0.344 | 0.765 | 0.375 | -3.863 | Likely Benign | 0.307 | Likely Benign | Likely Benign | 0.128 | Likely Benign | -0.70 | Neutral | 0.841 | Possibly Damaging | 0.809 | Possibly Damaging | 4.14 | Benign | 0.00 | Affected | 0.2244 | 0.5735 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.1230C>A | S410R 2D 3DClick to see structure in 3D Viewer AIClinVar contains no record for the SynGAP1 S410R variant, and it is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, Foldetta predicts a benign impact, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Taken together, the majority of evidence points to a benign outcome. This conclusion does not contradict ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -8.203 | Likely Pathogenic | 0.941 | Likely Pathogenic | Ambiguous | -0.67 | Ambiguous | 0.2 | 0.56 | Ambiguous | -0.06 | Likely Benign | 0.62 | Ambiguous | 0.148 | Likely Benign | -2.47 | Neutral | 0.871 | Possibly Damaging | 0.298 | Benign | 4.20 | Benign | 0.40 | Tolerated | 0.0955 | 0.3927 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||
| c.1230C>G | S410R 2D 3DClick to see structure in 3D Viewer AIClinVar contains no record for the SynGAP1 S410R variant, and it is not present in gnomAD. Prediction tools that agree on a benign effect include REVEL, Foldetta, PROVEAN, polyPhen‑2 HumVar, SIFT, and FATHMM. Those that predict a pathogenic effect are polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The remaining tools (FoldX, Rosetta, premPS, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, Foldetta predicts a benign impact, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive. Taken together, the majority of evidence points to a benign outcome. This conclusion does not contradict ClinVar, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.098513 | Structured | 0.349627 | Uncertain | 0.908 | 0.206 | 0.000 | -8.203 | Likely Pathogenic | 0.941 | Likely Pathogenic | Ambiguous | -0.67 | Ambiguous | 0.2 | 0.56 | Ambiguous | -0.06 | Likely Benign | 0.62 | Ambiguous | 0.146 | Likely Benign | -2.47 | Neutral | 0.871 | Possibly Damaging | 0.298 | Benign | 4.20 | Benign | 0.40 | Tolerated | 0.0955 | 0.3927 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||||
| c.1232T>C | I411T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 I411T missense variant is not reported in ClinVar (status: None) and has no entries in gnomAD. Prediction tools that agree on a benign effect are limited to FATHMM, while the majority of tools predict a pathogenic impact: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Rosetta’s assessment is uncertain. High‑accuracy methods give consistent pathogenic predictions: AlphaMissense‑Optimized is Pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) is Pathogenic. No prediction or stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.116183 | Structured | 0.339366 | Uncertain | 0.927 | 0.198 | 0.000 | -11.258 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 2.86 | Destabilizing | 0.0 | 1.75 | Ambiguous | 2.31 | Destabilizing | 1.86 | Destabilizing | 0.579 | Likely Pathogenic | -4.54 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.31 | Benign | 0.00 | Affected | 0.0903 | 0.1389 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1237C>A | P413T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P413T missense variant is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are limited to FATHMM, while the remaining tools—SGM‑Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and Foldetta—consistently predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this view: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a Likely Pathogenic verdict; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenicity. No predictions are inconclusive or missing. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.332472 | Uncertain | 0.927 | 0.201 | 0.000 | -12.851 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 3.22 | Destabilizing | 0.2 | 1.87 | Ambiguous | 2.55 | Destabilizing | 0.93 | Ambiguous | 0.554 | Likely Pathogenic | -7.37 | Deleterious | 1.000 | Probably Damaging | 0.993 | Probably Damaging | 3.18 | Benign | 0.01 | Affected | 0.1663 | 0.4670 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||
| c.1238C>A | P413H 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P413H is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include only FATHMM, while the remaining tools—SGM‑Consensus, REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic or likely pathogenic impact; Rosetta is uncertain. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No predictions are missing or inconclusive. Based on the overwhelming agreement among pathogenic predictors and the high‑accuracy tools, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.332472 | Uncertain | 0.927 | 0.201 | 0.000 | -13.376 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 4.89 | Destabilizing | 1.0 | 1.85 | Ambiguous | 3.37 | Destabilizing | 1.07 | Destabilizing | 0.546 | Likely Pathogenic | -8.29 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.16 | Benign | 0.00 | Affected | 0.1745 | 0.3480 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.1238C>G | P413R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P413R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining tools—SGM‑Consensus, REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. The remaining methods, Foldetta and Rosetta, yield uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.332472 | Uncertain | 0.927 | 0.201 | 0.000 | -15.523 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 2.40 | Destabilizing | 0.2 | 1.34 | Ambiguous | 1.87 | Ambiguous | 1.07 | Destabilizing | 0.562 | Likely Pathogenic | -8.29 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.32 | Benign | 0.01 | Affected | 0.1550 | 0.2199 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1241T>A | M414K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M414K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are limited to FATHMM, whereas the remaining tools—SGM‑Consensus, REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. FoldX and Foldetta provide uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta is unavailable. Based on the overwhelming agreement among the majority of prediction tools and the high‑accuracy methods, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | -13.537 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 1.18 | Ambiguous | 0.1 | 2.39 | Destabilizing | 1.79 | Ambiguous | 1.50 | Destabilizing | 0.503 | Likely Pathogenic | -5.03 | Deleterious | 0.942 | Possibly Damaging | 0.860 | Possibly Damaging | 3.40 | Benign | 0.04 | Affected | 0.1299 | 0.0888 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1241T>G | M414R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M414R has no ClinVar entry and is not reported in gnomAD. Functional prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while REVEL, Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Pathogenic.” High‑accuracy assessments further support a damaging outcome: AlphaMissense‑Optimized is pathogenic, SGM‑Consensus remains likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is inconclusive. Overall, the preponderance of evidence indicates that M414R is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.081712 | Structured | 0.329108 | Uncertain | 0.914 | 0.217 | 0.000 | -13.404 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 1.17 | Ambiguous | 0.1 | 2.58 | Destabilizing | 1.88 | Ambiguous | 1.45 | Destabilizing | 0.525 | Likely Pathogenic | -5.20 | Deleterious | 0.972 | Probably Damaging | 0.895 | Possibly Damaging | 3.38 | Benign | 0.03 | Affected | 0.1509 | 0.1037 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||
| c.1243G>A | E415K 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E415K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The high‑accuracy consensus shows AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Based on the overall balance of predictions—particularly the two high‑accuracy pathogenic calls versus one benign—the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.100716 | Structured | 0.330366 | Uncertain | 0.915 | 0.236 | 0.000 | -11.433 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | -0.07 | Likely Benign | 0.3 | -0.13 | Likely Benign | -0.10 | Likely Benign | 0.46 | Likely Benign | 0.326 | Likely Benign | -3.69 | Deleterious | 0.998 | Probably Damaging | 0.975 | Probably Damaging | 3.22 | Benign | 0.03 | Affected | 0.2174 | 0.3860 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1244A>C | E415A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E415A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, Rosetta, Foldetta, and FATHMM, whereas pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain results are given by FoldX and premPS. High‑accuracy assessments indicate that AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic, while Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts benign. Overall, the majority of tools lean toward pathogenicity, and the high‑accuracy predictions support this view. Therefore, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.100716 | Structured | 0.330366 | Uncertain | 0.915 | 0.236 | 0.000 | -11.743 | Likely Pathogenic | 0.981 | Likely Pathogenic | Likely Pathogenic | 0.61 | Ambiguous | 0.2 | 0.05 | Likely Benign | 0.33 | Likely Benign | 0.53 | Ambiguous | 0.435 | Likely Benign | -5.56 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 3.19 | Benign | 0.03 | Affected | 0.2909 | 0.3432 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1244A>G | E415G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E415G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and FATHMM. Tools that predict a pathogenic effect comprise PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Stability‑based methods (FoldX, Rosetta, premPS) and Foldetta are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as Likely Pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact for E415G. This prediction is consistent with the lack of ClinVar annotation and does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.100716 | Structured | 0.330366 | Uncertain | 0.915 | 0.236 | 0.000 | -12.244 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 1.16 | Ambiguous | 0.2 | 0.88 | Ambiguous | 1.02 | Ambiguous | 0.65 | Ambiguous | 0.432 | Likely Benign | -6.45 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.20 | Benign | 0.01 | Affected | 0.2518 | 0.3158 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1249T>C | Y417H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y417H is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include only FATHMM, whereas all other evaluated algorithms—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic or likely pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. No predictions are missing or inconclusive. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar entry exists for Y417H. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.179055 | Structured | 0.346865 | Uncertain | 0.958 | 0.250 | 0.000 | -11.447 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 3.22 | Destabilizing | 0.1 | 2.80 | Destabilizing | 3.01 | Destabilizing | 1.54 | Destabilizing | 0.513 | Likely Pathogenic | -4.77 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.97 | Benign | 0.00 | Affected | 0.2596 | 0.0668 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.124C>A | P42T 2D AIThe SynGAP1 missense variant P42T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P42T, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.308712 | Structured | 0.431487 | Uncertain | 0.420 | 0.771 | 0.375 | -3.626 | Likely Benign | 0.101 | Likely Benign | Likely Benign | 0.109 | Likely Benign | -0.97 | Neutral | 0.909 | Possibly Damaging | 0.901 | Possibly Damaging | 4.29 | Benign | 0.00 | Affected | 0.1834 | 0.4896 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.1250A>G | Y417C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y417C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining 13 tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact. High‑accuracy methods further support this: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely pathogenic effect; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also classifies the variant as pathogenic. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.179055 | Structured | 0.346865 | Uncertain | 0.958 | 0.250 | 0.000 | -12.021 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 4.03 | Destabilizing | 0.1 | 3.81 | Destabilizing | 3.92 | Destabilizing | 2.52 | Destabilizing | 0.597 | Likely Pathogenic | -8.59 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.95 | Benign | 0.00 | Affected | 0.2960 | 0.2005 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||
| c.1252A>G | K418E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K418E is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, and FATHMM. Tools that predict a pathogenic effect comprise SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. FoldX, Rosetta, and Foldetta are uncertain and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as uncertain. Overall, the majority of evaluated tools (seven pathogenic vs four benign) indicate that the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists for K418E. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.104810 | Structured | 0.360134 | Uncertain | 0.948 | 0.263 | 0.000 | -12.443 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.63 | Ambiguous | 0.0 | 0.80 | Ambiguous | 0.72 | Ambiguous | 0.47 | Likely Benign | 0.367 | Likely Benign | -3.42 | Deleterious | 0.999 | Probably Damaging | 0.991 | Probably Damaging | 3.53 | Benign | 0.07 | Tolerated | 0.3578 | 0.0471 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1253A>C | K418T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K418T variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that indicate a benign effect include REVEL, FoldX, FATHMM, premPS, and Foldetta. Those that predict a pathogenic effect comprise SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic, while Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of tools (10 pathogenic vs. 5 benign) support a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar record exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.104810 | Structured | 0.360134 | Uncertain | 0.948 | 0.263 | 0.000 | -11.994 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.39 | Likely Benign | 0.1 | 0.55 | Ambiguous | 0.47 | Likely Benign | 0.41 | Likely Benign | 0.392 | Likely Benign | -5.36 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.37 | Benign | 0.04 | Affected | 0.1975 | 0.1894 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1255G>A | E419K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E419K missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, and FATHMM. Tools that agree on a pathogenic effect include SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Foldetta and Rosetta give uncertain results and are not counted in either group. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus predicts likely pathogenic, and Foldetta remains uncertain. Overall, the majority of evidence points to a pathogenic impact for E419K. This conclusion is not contradicted by ClinVar, which contains no entry for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.371949 | Uncertain | 0.961 | 0.261 | 0.000 | -12.257 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.01 | Likely Benign | 0.1 | 1.30 | Ambiguous | 0.66 | Ambiguous | -0.03 | Likely Benign | 0.399 | Likely Benign | -3.75 | Deleterious | 0.998 | Probably Damaging | 0.975 | Probably Damaging | 3.36 | Benign | 0.07 | Tolerated | 0.2759 | 0.6899 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1256A>C | E419A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E419A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, and FATHMM, whereas a majority of tools predict a pathogenic impact: SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta as uncertain. Overall, the balance of evidence favors a pathogenic classification; this conclusion does not contradict ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.371949 | Uncertain | 0.961 | 0.261 | 0.000 | -10.951 | Likely Pathogenic | 0.944 | Likely Pathogenic | Ambiguous | 0.56 | Ambiguous | 0.1 | 0.94 | Ambiguous | 0.75 | Ambiguous | 0.28 | Likely Benign | 0.398 | Likely Benign | -5.60 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 3.32 | Benign | 0.03 | Affected | 0.4051 | 0.6705 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1256A>G | E419G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E419G is listed in ClinVar with an uncertain significance (ClinVar ID 2004834.0) and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL and FATHMM, while pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as likely pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus confirms likely pathogenic, and the Foldetta stability analysis is inconclusive. No evidence from FoldX, Rosetta, or premPS is available. Overall, the preponderance of predictions indicates that E419G is most likely pathogenic, which contrasts with the current ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.371949 | Uncertain | 0.961 | 0.261 | 0.000 | Uncertain | 1 | -10.589 | Likely Pathogenic | 0.956 | Likely Pathogenic | Likely Pathogenic | 1.41 | Ambiguous | 0.0 | 1.94 | Ambiguous | 1.68 | Ambiguous | 0.83 | Ambiguous | 0.469 | Likely Benign | -6.42 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.31 | Benign | 0.02 | Affected | 3.37 | 29 | 0.2992 | 0.5728 | 0 | -2 | 3.1 | -72.06 | 165.3 | 110.8 | 0.0 | 0.0 | -0.1 | 0.0 | X | Potentially Pathogenic | The carboxylate group of Glu419, located on an α helix (res. Met414-Glu436), forms a salt bridge with the side chain of either Arg716 or Lys418 from an opposing helix (res. Pro713-Arg726). The backbone amide group of Glu419 does not form H-bonds, resulting in a slight bend in the α helix. Thus, although glycine is known as an “α helix breaker,” the residue swap does not disrupt the continuity or integrity of the α helix. However, because Gly419 cannot form a salt bridge with the guanidinium group of the Arg716 side chain, the C2-GAP domain tertiary structure could be compromised during folding. | ||||||||||||||||
| c.125C>A | P42H 2D AIThe SynGAP1 missense variant P42H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote) which classifies the variant as Likely Benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized predicts Benign, and the SGM‑Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates Likely Benign. No Foldetta stability analysis is available for this variant. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.308712 | Structured | 0.431487 | Uncertain | 0.420 | 0.771 | 0.375 | -3.088 | Likely Benign | 0.167 | Likely Benign | Likely Benign | 0.140 | Likely Benign | -1.95 | Neutral | 0.992 | Probably Damaging | 0.977 | Probably Damaging | 4.16 | Benign | 0.00 | Affected | 0.1995 | 0.3705 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.125C>G | P42R 2D AIThe SynGAP1 missense variant P42R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Benign). In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT all predict a pathogenic impact. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized predicts benign, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Benign. No Foldetta stability result is available, so it does not influence the assessment. Overall, the majority of evidence points to a benign effect for P42R, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.308712 | Structured | 0.431487 | Uncertain | 0.420 | 0.771 | 0.375 | -2.498 | Likely Benign | 0.200 | Likely Benign | Likely Benign | 0.087 | Likely Benign | -1.79 | Neutral | 0.972 | Probably Damaging | 0.954 | Probably Damaging | 4.20 | Benign | 0.00 | Affected | 0.1619 | 0.3082 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||||||||||||
| c.1261G>A | A421T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A421T is not reported in ClinVar and is present in gnomAD (ID 6‑33438166‑G‑A). Functional prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, polyPhen‑2 (HumDiv and HumVar), SIFT, and FATHMM. Tools that predict a pathogenic effect are PROVEAN, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; the SGM‑Consensus score is “Likely Pathogenic.” Uncertain results from FoldX and premPS are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as benign. Overall, the majority of consensus predictions lean toward a benign impact, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.404927 | Uncertain | 0.965 | 0.257 | 0.000 | 6-33438166-G-A | 1 | 6.19e-7 | -9.217 | Likely Pathogenic | 0.975 | Likely Pathogenic | Likely Pathogenic | 0.75 | Ambiguous | 0.2 | 0.18 | Likely Benign | 0.47 | Likely Benign | 0.99 | Ambiguous | 0.179 | Likely Benign | -3.12 | Deleterious | 0.353 | Benign | 0.136 | Benign | 3.43 | Benign | 0.09 | Tolerated | 3.37 | 29 | 0.1346 | 0.4439 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||
| c.1262C>A | A421E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A421E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and FATHMM, whereas pathogenic calls are made by ESM1b, PROVEAN, AlphaMissense‑Default, AlphaMissense‑Optimized, Rosetta, premPS, and the SGM‑Consensus score (Likely Pathogenic). Stability‑based methods give mixed results: FoldX is uncertain, Foldetta is uncertain, and Rosetta alone predicts pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta remains uncertain. Overall, the majority of evidence, including the high‑accuracy tools, points to a pathogenic impact for A421E, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.404927 | Uncertain | 0.965 | 0.257 | 0.000 | -11.993 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.63 | Ambiguous | 0.2 | 2.07 | Destabilizing | 1.35 | Ambiguous | 1.30 | Destabilizing | 0.233 | Likely Benign | -4.24 | Deleterious | 0.368 | Benign | 0.144 | Benign | 3.44 | Benign | 0.14 | Tolerated | 0.1288 | 0.1830 | 0 | -1 | -5.3 | 58.04 | |||||||||||||||||||||||||||||
| c.1262C>T | A421V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A421V is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, FoldX, Foldetta, premPS, polyPhen‑2 HumVar, SIFT, and FATHMM, while pathogenic predictions are reported by PROVEAN, polyPhen‑2 HumDiv, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates a likely pathogenic outcome. High‑accuracy assessments further support this: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote) also predicts pathogenic, whereas Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta, predicts benign. Rosetta alone is uncertain and is treated as unavailable. Overall, the majority of high‑confidence tools favor a pathogenic effect, so A421V is most likely pathogenic, with no conflict with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.404927 | Uncertain | 0.965 | 0.257 | 0.000 | -8.167 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | -0.05 | Likely Benign | 0.1 | -0.82 | Ambiguous | -0.44 | Likely Benign | -0.06 | Likely Benign | 0.111 | Likely Benign | -3.15 | Deleterious | 0.538 | Possibly Damaging | 0.113 | Benign | 3.50 | Benign | 0.14 | Tolerated | 0.1055 | 0.3738 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1264G>A | E422K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E422K missense variant is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools show a split opinion: benign predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, and FATHMM, while pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support this view: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus also indicates Likely Pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts a benign effect. Overall, the balance of evidence leans toward pathogenicity, and this assessment does not contradict any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.426709 | Uncertain | 0.965 | 0.255 | 0.000 | -13.042 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.30 | Likely Benign | 0.0 | 0.10 | Likely Benign | 0.20 | Likely Benign | 0.32 | Likely Benign | 0.346 | Likely Benign | -3.52 | Deleterious | 0.998 | Probably Damaging | 0.975 | Probably Damaging | 3.39 | Benign | 0.04 | Affected | 0.1995 | 0.5129 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1265A>C | E422A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E422A missense variant is not reported in ClinVar and has no gnomAD entry. Prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, and FATHMM; pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized remains uncertain. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized is inconclusive; the SGM‑Consensus, which aggregates the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, indicates likely pathogenic; Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts benign. Overall, the balance of evidence (seven pathogenic versus six benign predictions) points to a likely pathogenic effect for E422A, and this conclusion is not contradicted by the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.426709 | Uncertain | 0.965 | 0.255 | 0.000 | -12.088 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 0.49 | Likely Benign | 0.0 | 0.25 | Likely Benign | 0.37 | Likely Benign | 0.26 | Likely Benign | 0.371 | Likely Benign | -5.43 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 3.31 | Benign | 0.01 | Affected | 0.2949 | 0.4459 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1265A>G | E422G 2D 3DClick to see structure in 3D Viewer AISynGAP1 E422G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split assessment: benign predictions come from REVEL, premPS, and FATHMM, while pathogenic predictions are reported by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Uncertain results are provided by FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy analyses indicate that the SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, predicts a likely pathogenic effect, whereas AlphaMissense‑Optimized remains inconclusive and Foldetta shows no definitive stability change. Overall, the majority of evidence points toward a pathogenic impact for E422G, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.426709 | Uncertain | 0.965 | 0.255 | 0.000 | -11.468 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 1.13 | Ambiguous | 0.0 | 1.25 | Ambiguous | 1.19 | Ambiguous | 0.39 | Likely Benign | 0.488 | Likely Benign | -6.38 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.31 | Benign | 0.01 | Affected | 0.2440 | 0.4184 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1267T>C | Y423H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 Y423H missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include REVEL, SIFT, ESM1b, and FATHMM. Tools that predict a pathogenic effect comprise FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and AlphaMissense‑Default. AlphaMissense‑Optimized is reported as uncertain. High‑accuracy assessments show that AlphaMissense‑Optimized is inconclusive, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is a 2‑vs‑2 tie and therefore inconclusive, while Foldetta predicts a pathogenic impact. Overall, the majority of evaluated tools (10 pathogenic vs. 4 benign) support a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.088832 | Structured | 0.421885 | Uncertain | 0.975 | 0.242 | 0.000 | -6.638 | Likely Benign | 0.873 | Likely Pathogenic | Ambiguous | 3.09 | Destabilizing | 0.1 | 2.44 | Destabilizing | 2.77 | Destabilizing | 1.66 | Destabilizing | 0.257 | Likely Benign | -3.88 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.39 | Benign | 0.61 | Tolerated | 0.1603 | 0.0352 | 0 | 2 | -1.9 | -26.03 | ||||||||||||||||||||||||||||||
| c.1268A>G | Y423C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y423C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM. The remaining tools—SGM‑Consensus, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—all predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain (treated as unavailable), SGM‑Consensus as Likely Pathogenic, and Foldetta as Pathogenic. With the majority of evidence pointing to deleterious effects, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.088832 | Structured | 0.421885 | Uncertain | 0.975 | 0.242 | 0.000 | -9.003 | Likely Pathogenic | 0.935 | Likely Pathogenic | Ambiguous | 4.01 | Destabilizing | 0.1 | 4.49 | Destabilizing | 4.25 | Destabilizing | 1.84 | Destabilizing | 0.286 | Likely Benign | -7.51 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.38 | Benign | 0.03 | Affected | 0.2381 | 0.1096 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||
| c.1271T>C | V424A 2D 3DClick to see structure in 3D Viewer AISynGAP1 V424A is not reported in ClinVar (ClinVar status: not reported) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on benign impact are REVEL and FATHMM, whereas the remaining tools—SGM‑Consensus, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—consistently predict pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as unavailable, SGM‑Consensus as likely pathogenic, and Foldetta as pathogenic. Based on the collective evidence, the variant is most likely pathogenic; this conclusion is not contradicted by the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.050641 | Structured | 0.411431 | Uncertain | 0.973 | 0.248 | 0.000 | -9.665 | Likely Pathogenic | 0.904 | Likely Pathogenic | Ambiguous | 2.31 | Destabilizing | 0.1 | 2.54 | Destabilizing | 2.43 | Destabilizing | 2.10 | Destabilizing | 0.245 | Likely Benign | -3.45 | Deleterious | 0.997 | Probably Damaging | 0.961 | Probably Damaging | 3.39 | Benign | 0.01 | Affected | 0.2105 | 0.1355 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1273A>C | T425P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T425P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Consensus from multiple in silico predictors indicates a pathogenic effect: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as deleterious, while only FATHMM predicts a benign outcome; premPS is uncertain. High‑accuracy tools reinforce this view: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports it as Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts a pathogenic effect. Taken together, the overwhelming majority of evidence supports a pathogenic classification, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.401218 | Uncertain | 0.964 | 0.280 | 0.000 | -12.318 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 2.81 | Destabilizing | 0.3 | 6.49 | Destabilizing | 4.65 | Destabilizing | 0.77 | Ambiguous | 0.512 | Likely Pathogenic | -5.16 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.37 | Benign | 0.03 | Affected | 0.1815 | 0.3469 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1274C>A | T425N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T425N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, FoldX, and FATHMM, whereas pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Predictions labeled uncertain (Foldetta, premPS, AlphaMissense‑Optimized, Rosetta) are treated as unavailable. High‑accuracy assessments further indicate a likely pathogenic outcome from the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and an uncertain result from AlphaMissense‑Optimized and Foldetta. Overall, the majority of definitive predictions support a pathogenic effect, and this conclusion does not contradict any ClinVar annotation because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.401218 | Uncertain | 0.964 | 0.280 | 0.000 | -10.709 | Likely Pathogenic | 0.925 | Likely Pathogenic | Ambiguous | 0.19 | Likely Benign | 0.1 | 0.81 | Ambiguous | 0.50 | Ambiguous | 0.82 | Ambiguous | 0.185 | Likely Benign | -3.87 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.44 | Benign | 0.04 | Affected | 0.1169 | 0.2843 | 0 | 0 | -2.8 | 13.00 | |||||||||||||||||||||||||||||
| c.1274C>T | T425I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T425I is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM; pathogenic predictions come from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further refine the picture: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic, whereas Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign stability. Overall, the balance of evidence slightly favors a pathogenic interpretation, with no conflict with ClinVar status because no ClinVar assertion exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.401218 | Uncertain | 0.964 | 0.280 | 0.000 | -10.443 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | -0.04 | Likely Benign | 0.2 | 0.07 | Likely Benign | 0.02 | Likely Benign | 0.30 | Likely Benign | 0.268 | Likely Benign | -5.30 | Deleterious | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 3.44 | Benign | 0.06 | Tolerated | 0.0827 | 0.4023 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.1277A>C | N426T 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense change N426T lies in the GAP domain. It is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, SIFT, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, and polyPhen‑2 HumVar. Predictions that are uncertain or inconclusive are FoldX, Foldetta, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also favors benign, while Foldetta remains uncertain and is not taken as evidence. Overall, the majority of tools, including the high‑accuracy methods, predict a benign effect. This consensus does not contradict any ClinVar annotation, as none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.042364 | Structured | 0.394941 | Uncertain | 0.959 | 0.287 | 0.000 | -7.389 | In-Between | 0.194 | Likely Benign | Likely Benign | 0.84 | Ambiguous | 0.0 | 0.38 | Likely Benign | 0.61 | Ambiguous | 0.12 | Likely Benign | 0.126 | Likely Benign | -3.14 | Deleterious | 0.983 | Probably Damaging | 0.926 | Probably Damaging | 3.47 | Benign | 0.10 | Tolerated | 0.1090 | 0.3002 | 0 | 0 | 2.8 | -13.00 | ||||||||||||||||||||||||||||||
| c.1279C>T | H427Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H427Y missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and polyPhen‑2 HumVar, as well as the SGM‑Consensus “Likely Benign” call. Tools that predict a pathogenic outcome are PROVEAN, polyPhen‑2 HumDiv, and SIFT. High‑accuracy assessments—AlphaMissense‑Optimized, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta (combining FoldX‑MD and Rosetta)—all uniformly indicate a benign effect. No prediction or folding‑stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.081712 | Structured | 0.394261 | Uncertain | 0.962 | 0.287 | 0.000 | -6.824 | Likely Benign | 0.328 | Likely Benign | Likely Benign | -0.08 | Likely Benign | 0.0 | -0.37 | Likely Benign | -0.23 | Likely Benign | 0.18 | Likely Benign | 0.180 | Likely Benign | -3.47 | Deleterious | 0.815 | Possibly Damaging | 0.073 | Benign | 3.44 | Benign | 0.01 | Affected | 0.0958 | 0.4029 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||
| c.1280A>C | H427P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H427P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools (FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus) predict a pathogenic impact. Two tools are uncertain: premPS and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as inconclusive, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) and Foldetta (combining FoldX‑MD and Rosetta outputs) both indicate a pathogenic effect. Overall, the preponderance of evidence points to a pathogenic classification for H427P, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.081712 | Structured | 0.394261 | Uncertain | 0.962 | 0.287 | 0.000 | -11.098 | Likely Pathogenic | 0.950 | Likely Pathogenic | Ambiguous | 4.74 | Destabilizing | 0.1 | 7.61 | Destabilizing | 6.18 | Destabilizing | 0.68 | Ambiguous | 0.324 | Likely Benign | -4.22 | Deleterious | 0.993 | Probably Damaging | 0.819 | Possibly Damaging | 3.51 | Benign | 0.01 | Affected | 0.2061 | 0.3442 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||
| c.1280A>G | H427R 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H427R is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438185‑A‑G). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and SIFT, while premPS and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also resolves to benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign stability change. No predictions or folding results are missing or inconclusive. Overall, the majority of evidence indicates the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.081712 | Structured | 0.394261 | Uncertain | 0.962 | 0.287 | 0.000 | 6-33438185-A-G | -3.259 | Likely Benign | 0.439 | Ambiguous | Likely Benign | -0.04 | Likely Benign | 0.1 | 0.48 | Likely Benign | 0.22 | Likely Benign | 0.72 | Ambiguous | 0.168 | Likely Benign | -2.61 | Deleterious | 0.174 | Benign | 0.018 | Benign | 3.51 | Benign | 0.03 | Affected | 3.38 | 25 | 0.2108 | 0.2121 | 0 | 2 | -1.3 | 19.05 | |||||||||||||||||||||||||||
| c.1283A>G | Y428C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y428C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect are REVEL and FATHMM. The remaining tools—FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus—consistently predict a pathogenic impact. High‑accuracy assessments show the SGM‑Consensus result as “Likely Pathogenic,” Foldetta predicts a destabilizing, pathogenic effect, while AlphaMissense‑Optimized is uncertain. No prediction or stability result is missing; the only inconclusive outcome is the AlphaMissense‑Optimized score. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation because no ClinVar status exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.118441 | Structured | 0.389652 | Uncertain | 0.965 | 0.292 | 0.000 | -11.312 | Likely Pathogenic | 0.852 | Likely Pathogenic | Ambiguous | 2.82 | Destabilizing | 0.0 | 2.99 | Destabilizing | 2.91 | Destabilizing | 2.02 | Destabilizing | 0.454 | Likely Benign | -8.59 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.39 | Benign | 0.01 | Affected | 0.3322 | 0.2220 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||
| c.1286G>C | R429P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R429P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and ESM1b; AlphaMissense‑Default and FoldX are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of tools and the high‑accuracy methods favor a pathogenic interpretation. Thus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar assertion exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.074921 | Structured | 0.390504 | Uncertain | 0.959 | 0.290 | 0.000 | -9.771 | Likely Pathogenic | 0.556 | Ambiguous | Likely Benign | 1.53 | Ambiguous | 0.2 | 5.13 | Destabilizing | 3.33 | Destabilizing | 1.01 | Destabilizing | 0.265 | Likely Benign | -2.89 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.43 | Benign | 0.25 | Tolerated | 0.1881 | 0.3790 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||
| c.1289T>A | M430K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M430K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN and ESM1b. The remaining tools—FoldX, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized classifies the variant as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) favors a pathogenic outcome; Foldetta remains uncertain. Overall, the majority of individual predictors lean toward a benign interpretation, whereas the SGM Consensus suggests pathogenicity. Given the lack of ClinVar evidence, there is no contradiction with existing clinical annotations. Based on the aggregate predictions, the variant is most likely benign, and this assessment does not conflict with the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.071867 | Structured | 0.385298 | Uncertain | 0.952 | 0.306 | 0.000 | -10.816 | Likely Pathogenic | 0.494 | Ambiguous | Likely Benign | 0.78 | Ambiguous | 0.1 | 0.44 | Likely Benign | 0.61 | Ambiguous | 0.86 | Ambiguous | 0.149 | Likely Benign | -2.66 | Deleterious | 0.134 | Benign | 0.033 | Benign | 3.47 | Benign | 0.32 | Tolerated | 0.1660 | 0.1271 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||
| c.1289T>G | M430R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M430R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, and ESM1b. The remaining tools—FoldX, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of consensus tools lean toward a benign classification, while a subset of high‑accuracy methods suggest pathogenicity, leaving the variant’s impact ambiguous. Based on the aggregate predictions, the variant is most likely benign, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.071867 | Structured | 0.385298 | Uncertain | 0.952 | 0.306 | 0.000 | -10.636 | Likely Pathogenic | 0.558 | Ambiguous | Likely Benign | 0.98 | Ambiguous | 0.1 | 0.47 | Likely Benign | 0.73 | Ambiguous | 0.83 | Ambiguous | 0.153 | Likely Benign | -2.87 | Deleterious | 0.620 | Possibly Damaging | 0.046 | Benign | 3.48 | Benign | 0.38 | Tolerated | 0.1775 | 0.1652 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||
| c.128G>C | G43A 2D AIThe SynGAP1 missense variant G43A is reported in gnomAD (variant ID 6-33423537‑G‑C) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus itself is likely benign; Foldetta results are not available. Overall, the preponderance of evidence points to a benign effect for G43A, and this conclusion does not contradict any ClinVar status, as none is currently assigned. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.308712 | Structured | 0.431462 | Uncertain | 0.396 | 0.762 | 0.375 | 6-33423537-G-C | 2 | 1.24e-6 | -3.925 | Likely Benign | 0.073 | Likely Benign | Likely Benign | 0.026 | Likely Benign | -0.52 | Neutral | 0.001 | Benign | 0.005 | Benign | 4.29 | Benign | 0.00 | Affected | 4.32 | 1 | 0.3677 | 0.3450 | 0 | 1 | 2.2 | 14.03 | ||||||||||||||||||||||||||||||||||
| c.1294T>A | C432S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C432S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, and FATHMM, whereas a majority of tools (SGM Consensus, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. Predictions that are inconclusive or unavailable are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show SGM Consensus as likely pathogenic, AlphaMissense‑Optimized as uncertain, and Foldetta as uncertain. Overall, the balance of evidence favors a pathogenic classification for C432S, and this assessment does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.111485 | Structured | 0.362533 | Uncertain | 0.960 | 0.285 | 0.000 | -8.229 | Likely Pathogenic | 0.913 | Likely Pathogenic | Ambiguous | 0.61 | Ambiguous | 0.1 | 0.96 | Ambiguous | 0.79 | Ambiguous | 1.52 | Destabilizing | 0.496 | Likely Benign | -9.55 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.53 | Benign | 0.12 | Tolerated | 0.4262 | 0.1415 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1295G>A | C432Y 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant C432Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely agree on a deleterious effect: pathogenic predictions come from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized, while benign predictions are reported by REVEL, Rosetta, FATHMM, and Foldetta. Tools with uncertain or missing results (FoldX, premPS) are not considered evidence. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predict pathogenicity, whereas Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a benign effect. Overall, the majority of predictions support a pathogenic impact, and this conclusion does not contradict any ClinVar annotation (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.111485 | Structured | 0.362533 | Uncertain | 0.960 | 0.285 | 0.000 | -13.720 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 0.62 | Ambiguous | 0.3 | 0.31 | Likely Benign | 0.47 | Likely Benign | 0.71 | Ambiguous | 0.441 | Likely Benign | -10.50 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | 3.45 | Benign | 0.02 | Affected | 0.0857 | 0.3438 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1295G>C | C432S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C432S is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, and FATHMM, whereas a majority of tools (SGM Consensus, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. Predictions that are inconclusive or unavailable are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show SGM Consensus as likely pathogenic, AlphaMissense‑Optimized as uncertain, and Foldetta as uncertain. Overall, the balance of evidence favors a pathogenic classification for C432S, and this assessment does not contradict any ClinVar status because no ClinVar entry exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.111485 | Structured | 0.362533 | Uncertain | 0.960 | 0.285 | 0.000 | -8.229 | Likely Pathogenic | 0.913 | Likely Pathogenic | Ambiguous | 0.61 | Ambiguous | 0.1 | 0.96 | Ambiguous | 0.79 | Ambiguous | 1.52 | Destabilizing | 0.417 | Likely Benign | -9.55 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.53 | Benign | 0.12 | Tolerated | 0.4262 | 0.1415 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1298C>A | A433E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A433E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, PROVEAN, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and ESM1b. Two tools (premPS and AlphaMissense‑Default) give uncertain results. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields a benign prediction; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports benign. No prediction is missing or inconclusive. Overall, the preponderance of evidence indicates that the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.352258 | Uncertain | 0.938 | 0.302 | 0.000 | -8.210 | Likely Pathogenic | 0.506 | Ambiguous | Likely Benign | -0.20 | Likely Benign | 0.0 | 0.15 | Likely Benign | -0.03 | Likely Benign | 0.58 | Ambiguous | 0.148 | Likely Benign | -1.65 | Neutral | 0.998 | Probably Damaging | 0.824 | Possibly Damaging | 3.42 | Benign | 0.18 | Tolerated | 0.0783 | 0.1695 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||
| c.1298C>T | A433V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A433V missense variant is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (ID 6‑33438203‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign and Foldetta as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive and therefore unavailable as evidence. Overall, the majority of predictions support a benign impact, and this is consistent with the lack of ClinVar pathogenic classification. Thus, the variant is most likely benign based on current computational evidence, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.098513 | Structured | 0.352258 | Uncertain | 0.938 | 0.302 | 0.000 | 6-33438203-C-T | 240 | 1.49e-4 | -8.200 | Likely Pathogenic | 0.129 | Likely Benign | Likely Benign | 0.48 | Likely Benign | 0.1 | -0.07 | Likely Benign | 0.21 | Likely Benign | 0.48 | Likely Benign | 0.096 | Likely Benign | -2.91 | Deleterious | 0.994 | Probably Damaging | 0.527 | Possibly Damaging | 3.43 | Benign | 0.04 | Affected | 3.37 | 29 | 0.0753 | 0.5104 | 0 | 0 | 2.4 | 28.05 | 214.5 | -45.8 | 0.0 | 0.0 | 0.0 | 0.2 | X | Potentially Benign | The methyl group of Ala433, located on the outer surface of an α helix (res. Met414-Glu436), does not interact with any nearby residues in the WT simulations. In the variant simulations, the iso-propyl side chain of Val433, which has a similarly hydrophobic profile as alanine, also does not form any lasting interactions at the helix surface. Accordingly, the residue swap does not negatively affect the protein structure based on the simulations. | ||||||||||||||||
| c.1301T>C | V434A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V434A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise Rosetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and ESM1b; FoldX and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as pathogenic. Overall, the majority of evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.158265 | Structured | 0.342846 | Uncertain | 0.954 | 0.306 | 0.000 | -8.531 | Likely Pathogenic | 0.441 | Ambiguous | Likely Benign | 1.72 | Ambiguous | 0.0 | 2.51 | Destabilizing | 2.12 | Destabilizing | 1.00 | Destabilizing | 0.238 | Likely Benign | -2.51 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.58 | Benign | 0.75 | Tolerated | 0.2338 | 0.2292 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||||||||
| c.1306G>A | E436K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E436K is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining evaluated algorithms (REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) uniformly predict a pathogenic impact; FoldX, Foldetta, and premPS are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for E436K, which does not conflict with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.239899 | Structured | 0.321046 | Uncertain | 0.934 | 0.289 | 0.000 | Uncertain | 1 | -13.869 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.56 | Ambiguous | 0.1 | 2.86 | Destabilizing | 1.71 | Ambiguous | 0.82 | Ambiguous | 0.829 | Likely Pathogenic | -3.77 | Deleterious | 0.994 | Probably Damaging | 0.951 | Probably Damaging | 4.71 | Benign | 0.02 | Affected | 3.37 | 29 | 0.2332 | 0.5995 | 0 | 1 | -0.4 | -0.94 | 186.8 | 39.8 | 0.0 | 0.0 | -0.2 | 0.0 | X | X | X | Potentially Pathogenic | The carboxylate group of Glu436, located on the α helix (res. Met414-Glu436), forms a salt bridge with the amino group of the Lys444 side chain on an opposing α helix (res. Val441-Ser457). The backbone carbonyl of Glu436 also H-bonds with the Lys444 side chain, which helps keep the ends of the two α helices tightly connected. In contrast, in the variant simulations, the salt bridge formation with Lys444 is not possible. Instead, the repelled Lys436 side chain rotates outward, causing a change in the α helix backbone H-bonding: the amide group of Lys444 H-bonds with the carbonyl of Ala433 instead of the carbonyl of Cys432. | ||||||||||||||
| c.1307A>C | E436A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E436A is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include Foldetta, premPS, and FATHMM, whereas the majority of algorithms—SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. Two high‑accuracy predictors give concordant results: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports a benign effect. No prediction is missing or inconclusive. Overall, the preponderance of evidence points to a pathogenic impact for E436A, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.239899 | Structured | 0.321046 | Uncertain | 0.934 | 0.289 | 0.000 | -12.678 | Likely Pathogenic | 0.971 | Likely Pathogenic | Likely Pathogenic | 1.03 | Ambiguous | 0.0 | -0.88 | Ambiguous | 0.08 | Likely Benign | 0.23 | Likely Benign | 0.825 | Likely Pathogenic | -5.68 | Deleterious | 0.998 | Probably Damaging | 0.991 | Probably Damaging | 4.65 | Benign | 0.05 | Affected | 0.3901 | 0.5116 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1307A>G | E436G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E436G missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FATHMM, while the majority of algorithms (SGM‑Consensus, REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact; FoldX, premPS, and Foldetta are inconclusive. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta’s stability prediction is uncertain. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.239899 | Structured | 0.321046 | Uncertain | 0.934 | 0.289 | 0.000 | -12.355 | Likely Pathogenic | 0.966 | Likely Pathogenic | Likely Pathogenic | 1.50 | Ambiguous | 0.1 | 2.02 | Destabilizing | 1.76 | Ambiguous | 0.58 | Ambiguous | 0.802 | Likely Pathogenic | -6.63 | Deleterious | 1.000 | Probably Damaging | 0.993 | Probably Damaging | 4.61 | Benign | 0.03 | Affected | 0.2910 | 0.4841 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1309C>A | P437T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P437T missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, FATHMM, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The remaining tools—FoldX, Foldetta, premPS, and AlphaMissense‑Default—return uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicting pathogenic, and Foldetta yielding an uncertain stability change. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict the lack of ClinVar annotation. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.175930 | Structured | 0.306196 | Uncertain | 0.921 | 0.298 | 0.000 | -13.011 | Likely Pathogenic | 0.484 | Ambiguous | Likely Benign | 1.35 | Ambiguous | 0.0 | -3.46 | Stabilizing | -1.06 | Ambiguous | 0.54 | Ambiguous | 0.305 | Likely Benign | -6.67 | Deleterious | 0.999 | Probably Damaging | 0.985 | Probably Damaging | 3.45 | Benign | 0.01 | Affected | 0.1659 | 0.5782 | 0 | -1 | 0.9 | 3.99 | ||||||||||||||||||||||||||||||
| c.1310C>A | P437H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P437H is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, and FATHMM, whereas those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Predictions that are inconclusive or unavailable are FoldX, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show that the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts a likely pathogenic outcome, while AlphaMissense‑Optimized and Foldetta remain uncertain. Overall, the balance of evidence favors a pathogenic classification for P437H, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.306196 | Uncertain | 0.921 | 0.298 | 0.000 | -13.496 | Likely Pathogenic | 0.826 | Likely Pathogenic | Ambiguous | 1.19 | Ambiguous | 0.0 | -3.28 | Stabilizing | -1.05 | Ambiguous | 0.31 | Likely Benign | 0.343 | Likely Benign | -7.75 | Deleterious | 1.000 | Probably Damaging | 0.995 | Probably Damaging | 3.44 | Benign | 0.01 | Affected | 0.1819 | 0.4328 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.1310C>G | P437R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant P437R is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, and FATHMM, whereas pathogenic predictions are made by SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. With the majority of tools indicating pathogenicity and the high‑accuracy consensus leaning toward pathogenic, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.306196 | Uncertain | 0.921 | 0.298 | 0.000 | -13.094 | Likely Pathogenic | 0.879 | Likely Pathogenic | Ambiguous | 0.34 | Likely Benign | 0.1 | -3.52 | Stabilizing | -1.59 | Ambiguous | 0.61 | Ambiguous | 0.461 | Likely Benign | -7.72 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.51 | Benign | 0.03 | Affected | 0.1448 | 0.2846 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1313C>A | A438D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A438D missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, polyPhen‑2 HumVar, and FATHMM; pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, SIFT, and AlphaMissense‑Default; the remaining tools (FoldX, Rosetta, Foldetta, premPS, ESM1b, AlphaMissense‑Optimized) are uncertain. High‑accuracy assessments indicate that AlphaMissense‑Optimized is inconclusive, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta is also inconclusive. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Thus, based on the available predictions, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.147574 | Structured | 0.290154 | Uncertain | 0.929 | 0.293 | 0.000 | -7.410 | In-Between | 0.808 | Likely Pathogenic | Ambiguous | 1.60 | Ambiguous | 0.1 | 0.70 | Ambiguous | 1.15 | Ambiguous | 0.92 | Ambiguous | 0.175 | Likely Benign | -3.07 | Deleterious | 0.859 | Possibly Damaging | 0.124 | Benign | 4.10 | Benign | 0.04 | Affected | 0.1490 | 0.1941 | 0 | -2 | -5.3 | 44.01 | ||||||||||||||||||||||||||||||
| c.1313C>T | A438V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A438V is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (gnomAD ID 6‑33438218‑C‑T). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a pathogenic outcome. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates “Likely Benign”; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is inconclusive (Uncertain). Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.147574 | Structured | 0.290154 | Uncertain | 0.929 | 0.293 | 0.000 | 6-33438218-C-T | 3 | 1.86e-6 | 1.405 | Likely Benign | 0.073 | Likely Benign | Likely Benign | -0.36 | Likely Benign | 0.0 | -0.70 | Ambiguous | -0.53 | Ambiguous | -1.21 | Stabilizing | 0.046 | Likely Benign | 1.03 | Neutral | 0.000 | Benign | 0.000 | Benign | 4.45 | Benign | 0.97 | Tolerated | 3.38 | 26 | 0.0904 | 0.5292 | 0 | 0 | 2.4 | 28.05 | 10.1016/j.ajhg.2020.11.011 | |||||||||||||||||||||||
| c.1319A>C | N440T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N440T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly favor a benign effect: AlphaMissense‑Default, AlphaMissense‑Optimized, ESM1b, FATHMM, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, REVEL, premPS, Rosetta, Foldetta, and the SGM‑Consensus (majority vote) all predict benign or likely benign. No tool predicts pathogenicity; the only inconclusive result is from FoldX, which is listed as uncertain. High‑accuracy methods corroborate the benign assessment: AlphaMissense‑Optimized is benign, the SGM‑Consensus is likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) is benign. Therefore, based on the available predictions, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.191378 | Structured | 0.267204 | Uncertain | 0.929 | 0.245 | 0.000 | -5.371 | Likely Benign | 0.143 | Likely Benign | Likely Benign | 0.58 | Ambiguous | 0.0 | 0.16 | Likely Benign | 0.37 | Likely Benign | 0.11 | Likely Benign | 0.079 | Likely Benign | -1.27 | Neutral | 0.007 | Benign | 0.005 | Benign | 3.48 | Benign | 0.14 | Tolerated | 0.0969 | 0.3341 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||
| c.1322T>C | V441A 2D 3DClick to see structure in 3D Viewer AISynGAP1 variant V441A is listed in ClinVar as uncertain and is present in gnomAD (ID 6‑33438227‑T‑C). Consensus from most in silico predictors favors a benign effect: REVEL, FoldX, Rosetta, Foldetta, polyPhen‑2 HumVar, SIFT, FATHMM, and AlphaMissense‑Optimized all report benign. Pathogenic predictions come from PROVEAN, polyPhen‑2 HumDiv, and ESM1b, while premPS and AlphaMissense‑Default remain uncertain. High‑accuracy assessments give a mixed picture: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports benign. Overall, the preponderance of evidence points to a benign impact, aligning with the ClinVar uncertain designation rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.161087 | Structured | 0.259875 | Uncertain | 0.918 | 0.249 | 0.000 | Conflicting | 2 | 6-33438227-T-C | 3 | 1.86e-6 | -9.439 | Likely Pathogenic | 0.359 | Ambiguous | Likely Benign | -0.14 | Likely Benign | 0.0 | 0.33 | Likely Benign | 0.10 | Likely Benign | 0.95 | Ambiguous | 0.053 | Likely Benign | -2.92 | Deleterious | 0.513 | Possibly Damaging | 0.214 | Benign | 3.44 | Benign | 0.93 | Tolerated | 3.37 | 29 | 0.2390 | 0.1800 | 0 | 0 | -2.4 | -28.05 | 195.0 | 44.6 | 0.0 | 0.1 | 0.5 | 0.0 | X | X | Uncertain | The iso-propyl side chain of Val441, located on the outer surface of an α helix (res. Asn440-Thr458), does not interact with other residues in the WT simulations. In the variant simulations, the methyl side chain of Ala441 is similarly hydrophobic and does not form any interactions on the outer helix surface. Although the residue swap does not negatively affect the protein structure based on the simulations, it is noteworthy that the residue faces the RasGTPase interface. Thus, the effect of the residue swap on the SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | |||||||||||||
| c.1324A>G | K442E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K442E is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split consensus: benign predictions come from REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments give mixed results: AlphaMissense‑Optimized remains uncertain; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts benign. No single evidence decisively outweighs the others. Therefore, the variant’s pathogenicity remains uncertain; the predictions do not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.170161 | Structured | 0.255766 | Uncertain | 0.912 | 0.225 | 0.000 | -12.813 | Likely Pathogenic | 0.939 | Likely Pathogenic | Ambiguous | -0.05 | Likely Benign | 0.1 | -0.21 | Likely Benign | -0.13 | Likely Benign | 0.24 | Likely Benign | 0.324 | Likely Benign | -3.34 | Deleterious | 0.997 | Probably Damaging | 0.966 | Probably Damaging | 3.47 | Benign | 0.16 | Tolerated | 0.2943 | 0.0789 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1325A>C | K442T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K442T missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM. Those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, while Foldetta (combining FoldX‑MD and Rosetta outputs) indicates a benign stability change; AlphaMissense‑Optimized remains inconclusive. Overall, the predictions are evenly split, with no single consensus. Thus, the variant is most likely benign based on the majority of evidence, and this assessment does not contradict any ClinVar record (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.170161 | Structured | 0.255766 | Uncertain | 0.912 | 0.225 | 0.000 | -11.273 | Likely Pathogenic | 0.865 | Likely Pathogenic | Ambiguous | 0.30 | Likely Benign | 0.2 | 0.21 | Likely Benign | 0.26 | Likely Benign | 0.22 | Likely Benign | 0.330 | Likely Benign | -5.01 | Deleterious | 0.999 | Probably Damaging | 0.999 | Probably Damaging | 3.43 | Benign | 0.07 | Tolerated | 0.1510 | 0.2859 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1330A>G | K444E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K444E missense variant is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and FATHMM; all other evaluated predictors—including FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently classify the variant as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also reports a pathogenic effect. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, and this assessment does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.203355 | Structured | 0.262172 | Uncertain | 0.955 | 0.213 | 0.000 | -15.571 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 2.99 | Destabilizing | 0.1 | 3.75 | Destabilizing | 3.37 | Destabilizing | 1.10 | Destabilizing | 0.437 | Likely Benign | -3.82 | Deleterious | 0.997 | Probably Damaging | 0.981 | Probably Damaging | 3.40 | Benign | 0.01 | Affected | 0.3486 | 0.0793 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1331A>C | K444T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K444T is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect are REVEL and FATHMM. Those that predict a pathogenic effect include FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools with uncertain or inconclusive results are Foldetta, Rosetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta as uncertain (treated as unavailable for pathogenicity inference). Overall, the majority of reliable predictions indicate a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.203355 | Structured | 0.262172 | Uncertain | 0.955 | 0.213 | 0.000 | -15.557 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 2.12 | Destabilizing | 0.1 | 1.17 | Ambiguous | 1.65 | Ambiguous | 0.96 | Ambiguous | 0.442 | Likely Benign | -5.73 | Deleterious | 0.999 | Probably Damaging | 1.000 | Probably Damaging | 3.45 | Benign | 0.01 | Affected | 0.1643 | 0.3416 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1331A>T | K444M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K444M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, and FATHMM, whereas pathogenic predictions are returned by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments further highlight the discrepancy: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, reports a benign effect. Overall, the majority of tools lean toward a pathogenic interpretation, and this is not contradicted by any ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.203355 | Structured | 0.262172 | Uncertain | 0.955 | 0.213 | 0.000 | -14.223 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 0.39 | Likely Benign | 0.1 | -0.47 | Likely Benign | -0.04 | Likely Benign | 0.33 | Likely Benign | 0.442 | Likely Benign | -5.73 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.41 | Benign | 0.00 | Affected | 0.0934 | 0.3890 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1333G>A | E445K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E445K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from FoldX, Rosetta, Foldetta, premPS, and FATHMM, whereas pathogenic calls arise from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. When high‑accuracy methods are considered separately, AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of evidence—including the high‑accuracy tools—supports a pathogenic effect. This conclusion does not conflict with ClinVar, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.270205 | Uncertain | 0.947 | 0.228 | 0.000 | -15.371 | Likely Pathogenic | 0.958 | Likely Pathogenic | Likely Pathogenic | 0.02 | Likely Benign | 0.0 | 0.02 | Likely Benign | 0.02 | Likely Benign | 0.46 | Likely Benign | 0.524 | Likely Pathogenic | -3.82 | Deleterious | 0.991 | Probably Damaging | 0.951 | Probably Damaging | 3.35 | Benign | 0.02 | Affected | 0.1784 | 0.5311 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1334A>C | E445A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E445A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, and FATHMM. Tools that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized and premPS are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) as Benign. Overall, the majority of predictions lean toward pathogenicity, and the high‑accuracy consensus supports a likely pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.270205 | Uncertain | 0.947 | 0.228 | 0.000 | -12.659 | Likely Pathogenic | 0.849 | Likely Pathogenic | Ambiguous | 0.09 | Likely Benign | 0.0 | -0.34 | Likely Benign | -0.13 | Likely Benign | 0.56 | Ambiguous | 0.476 | Likely Benign | -5.73 | Deleterious | 0.997 | Probably Damaging | 0.991 | Probably Damaging | 3.34 | Benign | 0.01 | Affected | 0.2846 | 0.4876 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1334A>G | E445G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E445G missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FATHMM, while the majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus) predict a pathogenic impact. Predictions that are inconclusive or uncertain are FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for E445G, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.270205 | Uncertain | 0.947 | 0.228 | 0.000 | -12.294 | Likely Pathogenic | 0.823 | Likely Pathogenic | Ambiguous | 0.80 | Ambiguous | 0.1 | 0.79 | Ambiguous | 0.80 | Ambiguous | 0.69 | Ambiguous | 0.523 | Likely Pathogenic | -6.68 | Deleterious | 1.000 | Probably Damaging | 0.993 | Probably Damaging | 3.39 | Benign | 0.01 | Affected | 0.2446 | 0.4202 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1337A>C | E446A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E446A is not reported in ClinVar and is absent from gnomAD. In silico predictors that classify the variant as benign include REVEL, Rosetta, FATHMM, and AlphaMissense‑Optimized. Predictors that classify it as pathogenic comprise SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, yielded an inconclusive result and is treated as unavailable. Overall, the majority of predictions lean toward pathogenicity, and this conclusion is not contradicted by the absence of a ClinVar entry. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.271506 | Structured | 0.276479 | Uncertain | 0.940 | 0.216 | 0.000 | -9.868 | Likely Pathogenic | 0.677 | Likely Pathogenic | Likely Benign | 1.66 | Ambiguous | 0.7 | 0.39 | Likely Benign | 1.03 | Ambiguous | 0.67 | Ambiguous | 0.443 | Likely Benign | -5.60 | Deleterious | 0.998 | Probably Damaging | 0.996 | Probably Damaging | 3.28 | Benign | 0.01 | Affected | 0.3195 | 0.5877 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1337A>G | E446G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E446G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only FATHMM. The majority of tools predict a pathogenic impact: SGM‑Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and Foldetta. Predictions that are uncertain or inconclusive are AlphaMissense‑Optimized, Rosetta, and premPS. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is Pathogenic. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.271506 | Structured | 0.276479 | Uncertain | 0.940 | 0.216 | 0.000 | -11.457 | Likely Pathogenic | 0.866 | Likely Pathogenic | Ambiguous | 2.62 | Destabilizing | 0.7 | 1.63 | Ambiguous | 2.13 | Destabilizing | 0.83 | Ambiguous | 0.510 | Likely Pathogenic | -6.42 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 3.24 | Benign | 0.00 | Affected | 0.2665 | 0.5202 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1340T>C | V447A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V447A missense variant is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise SGM‑Consensus, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and Foldetta both predict pathogenicity. No predictions are missing or inconclusive. Based on the overall distribution of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.250310 | Structured | 0.283801 | Uncertain | 0.970 | 0.243 | 0.000 | -9.852 | Likely Pathogenic | 0.692 | Likely Pathogenic | Likely Benign | 2.18 | Destabilizing | 0.0 | 2.72 | Destabilizing | 2.45 | Destabilizing | 1.56 | Destabilizing | 0.266 | Likely Benign | -2.84 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.37 | Benign | 0.15 | Tolerated | 0.2456 | 0.2124 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1343C>A | A448D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A448D is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include only FATHMM, whereas the remaining 13 tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact. High‑accuracy methods further support this: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. No prediction or folding result is missing or inconclusive. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.225814 | Structured | 0.292774 | Uncertain | 0.973 | 0.257 | 0.000 | -17.290 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 8.13 | Destabilizing | 0.2 | 4.35 | Destabilizing | 6.24 | Destabilizing | 1.40 | Destabilizing | 0.662 | Likely Pathogenic | -5.93 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 3.13 | Benign | 0.00 | Affected | 0.1541 | 0.1741 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1343C>T | A448V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A448V is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas a majority of tools predict a pathogenic impact: FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). Two tools (Rosetta and premPS) yield uncertain results. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Overall, the preponderance of evidence indicates that A448V is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.225814 | Structured | 0.292774 | Uncertain | 0.973 | 0.257 | 0.000 | -10.372 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 3.31 | Destabilizing | 0.2 | 1.80 | Ambiguous | 2.56 | Destabilizing | 0.66 | Ambiguous | 0.487 | Likely Benign | -3.95 | Deleterious | 0.998 | Probably Damaging | 0.955 | Probably Damaging | 3.26 | Benign | 0.02 | Affected | 0.0950 | 0.4955 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1345A>C | S449R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant S449R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, FATHMM, AlphaMissense‑Optimized, and polyPhen‑2 HumVar. Those that predict a pathogenic effect are SGM Consensus, PROVEAN, polyPhen‑2 HumDiv, AlphaMissense‑Default, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) classifying it as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) yielding an uncertain stability change. No folding‑stability prediction is definitive. Overall, the majority of tools predict a benign outcome, and this does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.254060 | Structured | 0.301437 | Uncertain | 0.958 | 0.251 | 0.000 | -8.486 | Likely Pathogenic | 0.677 | Likely Pathogenic | Likely Benign | -0.69 | Ambiguous | 0.2 | -1.33 | Ambiguous | -1.01 | Ambiguous | 0.50 | Likely Benign | 0.145 | Likely Benign | -3.36 | Deleterious | 0.950 | Possibly Damaging | 0.214 | Benign | 3.40 | Benign | 0.18 | Tolerated | 0.0762 | 0.3250 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1345A>G | S449G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S449G is listed in ClinVar with an “Uncertain” status and is present in the gnomAD database (variant ID 6‑33438250‑A‑G). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only polyPhen‑2 HumDiv predicts a pathogenic outcome, while Rosetta, Foldetta, and premPS are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as “Likely Benign,” and Foldetta as uncertain. Overall, the majority of evidence points to a benign effect, and this conclusion does not contradict the ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.254060 | Structured | 0.301437 | Uncertain | 0.958 | 0.251 | 0.000 | Uncertain | 1 | 6-33438250-A-G | 3 | 1.86e-6 | -5.936 | Likely Benign | 0.071 | Likely Benign | Likely Benign | 0.47 | Likely Benign | 0.0 | 0.55 | Ambiguous | 0.51 | Ambiguous | 0.85 | Ambiguous | 0.116 | Likely Benign | -2.32 | Neutral | 0.948 | Possibly Damaging | 0.124 | Benign | 3.35 | Benign | 0.13 | Tolerated | 3.37 | 32 | 0.2600 | 0.3718 | 0 | 1 | 0.4 | -30.03 | ||||||||||||||||||||||
| c.1345A>T | S449C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S449C is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors classify the substitution as benign: REVEL, FoldX, Rosetta, premPS (uncertain), PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts pathogenicity. High‑accuracy assessments concur: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also reports a benign effect. Based on the unanimous benign predictions and lack of ClinVar evidence, the variant is most likely benign, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.254060 | Structured | 0.301437 | Uncertain | 0.958 | 0.251 | 0.000 | -2.207 | Likely Benign | 0.050 | Likely Benign | Likely Benign | 0.48 | Likely Benign | 0.0 | -0.31 | Likely Benign | 0.09 | Likely Benign | -0.57 | Ambiguous | 0.067 | Likely Benign | 0.48 | Neutral | 0.000 | Benign | 0.000 | Benign | 3.32 | Benign | 0.21 | Tolerated | 0.0864 | 0.5012 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||
| c.1347T>A | S449R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S449R is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, SIFT, FATHMM, and polyPhen‑2 HumVar, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments provide a mixed picture: AlphaMissense‑Optimized predicts a benign effect, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome, and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, yields an inconclusive result. FoldX and Rosetta predictions are also uncertain and are treated as unavailable. Overall, the evidence is balanced, with an equal number of benign and pathogenic calls, and the high‑accuracy tools do not converge on a single conclusion. Consequently, the variant is most likely pathogenic based on the preponderance of pathogenic predictions, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.254060 | Structured | 0.301437 | Uncertain | 0.958 | 0.251 | 0.000 | -8.486 | Likely Pathogenic | 0.677 | Likely Pathogenic | Likely Benign | -0.69 | Ambiguous | 0.2 | -1.33 | Ambiguous | -1.01 | Ambiguous | 0.50 | Likely Benign | 0.168 | Likely Benign | -3.36 | Deleterious | 0.950 | Possibly Damaging | 0.214 | Benign | 3.40 | Benign | 0.18 | Tolerated | 0.0762 | 0.3250 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1347T>G | S449R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S449R is not reported in ClinVar (ClinVar status: None) and has no entry in gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include REVEL, premPS, SIFT, FATHMM, and polyPhen‑2 HumVar. Those that predict pathogenicity are SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicting a benign effect, while the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome; Foldetta’s stability prediction is uncertain and therefore treated as unavailable. Overall, the predictions are split, with an equal number of benign and pathogenic calls and conflicting high‑accuracy results. Consequently, the variant’s impact remains inconclusive, and there is no contradiction with the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.254060 | Structured | 0.301437 | Uncertain | 0.958 | 0.251 | 0.000 | -8.486 | Likely Pathogenic | 0.677 | Likely Pathogenic | Likely Benign | -0.69 | Ambiguous | 0.2 | -1.33 | Ambiguous | -1.01 | Ambiguous | 0.50 | Likely Benign | 0.167 | Likely Benign | -3.36 | Deleterious | 0.950 | Possibly Damaging | 0.214 | Benign | 3.40 | Benign | 0.18 | Tolerated | 0.0762 | 0.3250 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1349C>A | A450E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A450E is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that classify the variant as benign include SIFT and FATHMM, whereas the majority of tools predict it to be pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. No predictions are inconclusive. Overall, the evidence strongly favors a pathogenic impact for A450E, which does not contradict the current ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.321458 | Structured | 0.306281 | Uncertain | 0.963 | 0.234 | 0.000 | Uncertain | 1 | -16.578 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 3.86 | Destabilizing | 0.2 | 5.23 | Destabilizing | 4.55 | Destabilizing | 1.59 | Destabilizing | 0.653 | Likely Pathogenic | -4.67 | Deleterious | 0.999 | Probably Damaging | 0.992 | Probably Damaging | 3.38 | Benign | 0.07 | Tolerated | 3.37 | 32 | 0.0823 | 0.1695 | 0 | -1 | -5.3 | 58.04 | 240.1 | -82.6 | 0.0 | 0.0 | 0.7 | 0.0 | X | X | Potentially Pathogenic | The methyl group of Ala450, located in an α helix (res. Asn440-Thr458), packs against hydrophobic residues in the inter-helix space (e.g., Leu692). In the variant simulations, the carboxylate group of the Glu450 side chain rotates outward, away from the hydrophobic niche, where it does not form any lasting salt bridges or H-bonds. Although the residue swap does not negatively affect the protein structure based on the simulations, it is possible that the introduction of the negatively charged residue adversely affects the folding process or tertiary assembly. | |||||||||||||||
| c.1349C>T | A450V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A450V is not reported in ClinVar and is present in gnomAD (ID 6‑33438254‑C‑T). Functional prediction tools cluster into two groups: benign predictions from REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments give mixed results: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of tools lean toward a benign effect, and there is no ClinVar entry to contradict this assessment. Thus, the variant is most likely benign based on current predictions, with a single high‑accuracy tool suggesting pathogenicity but not overturning the overall benign consensus. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.321458 | Structured | 0.306281 | Uncertain | 0.963 | 0.234 | 0.000 | 6-33438254-C-T | 1 | 6.20e-7 | -11.489 | Likely Pathogenic | 0.578 | Likely Pathogenic | Likely Benign | 0.04 | Likely Benign | 0.2 | 0.27 | Likely Benign | 0.16 | Likely Benign | 0.46 | Likely Benign | 0.306 | Likely Benign | -3.69 | Deleterious | 0.998 | Probably Damaging | 0.955 | Probably Damaging | 3.55 | Benign | 0.04 | Affected | 3.37 | 32 | 0.0792 | 0.5638 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||
| c.134A>C | N45T 2D AIThe SynGAP1 missense variant N45T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.401658 | Structured | 0.431853 | Uncertain | 0.498 | 0.741 | 0.375 | -2.425 | Likely Benign | 0.367 | Ambiguous | Likely Benign | 0.075 | Likely Benign | -0.81 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.08 | Benign | 0.00 | Affected | 0.1642 | 0.8318 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||||||||||||
| c.1355T>C | V452A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V452A is not reported in ClinVar and has no entry in gnomAD. Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while SGM‑Consensus, REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as pathogenic. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, yields an uncertain result and is treated as unavailable evidence. High‑accuracy assessments reinforce the pathogenic prediction: AlphaMissense‑Optimized reports pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic, and Foldetta remains inconclusive. Overall, the consensus of the available tools points to a pathogenic effect for V452A, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.335645 | Structured | 0.315167 | Uncertain | 0.970 | 0.229 | 0.000 | -10.423 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 2.21 | Destabilizing | 0.0 | 1.51 | Ambiguous | 1.86 | Ambiguous | 2.18 | Destabilizing | 0.505 | Likely Pathogenic | -3.95 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.21 | Benign | 0.00 | Affected | 0.2642 | 0.2521 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1357C>T | H453Y 2D AIThe SynGAP1 missense variant H453Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign calls come from REVEL, premPS, FATHMM, and AlphaMissense‑Optimized; pathogenic calls come from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus. Predictions from FoldX, Rosetta, and Foldetta are uncertain. High‑accuracy methods provide a mixed picture: AlphaMissense‑Optimized predicts benign, SGM‑Consensus predicts likely pathogenic, and Foldetta is inconclusive. Overall, the majority of evidence points toward a pathogenic effect, aligning with the SGM‑Consensus but opposing the benign predictions from several tools. Because ClinVar contains no entry for this variant, there is no conflict with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.352862 | Structured | 0.316097 | Uncertain | 0.946 | 0.200 | 0.000 | -12.060 | Likely Pathogenic | 0.753 | Likely Pathogenic | Likely Benign | -0.52 | Ambiguous | 0.7 | -0.68 | Ambiguous | -0.60 | Ambiguous | 0.09 | Likely Benign | 0.320 | Likely Benign | -5.93 | Deleterious | 0.995 | Probably Damaging | 0.961 | Probably Damaging | 3.41 | Benign | 0.04 | Affected | 0.0815 | 0.3843 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||
| c.1358A>C | H453P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H453P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL and FATHMM. The majority of tools predict a pathogenic impact: FoldX, Rosetta, Foldetta, premPS (uncertain), PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. No predictions are missing or inconclusive. Based on the preponderance of evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.352862 | Structured | 0.316097 | Uncertain | 0.946 | 0.200 | 0.000 | -15.704 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 3.29 | Destabilizing | 0.4 | 7.04 | Destabilizing | 5.17 | Destabilizing | 0.84 | Ambiguous | 0.488 | Likely Benign | -9.88 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.40 | Benign | 0.04 | Affected | 0.2115 | 0.3938 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||
| c.1361T>C | I454T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I454T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only FATHMM, whereas all other evaluated algorithms—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. High‑accuracy methods further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No prediction or folding stability result is missing or inconclusive. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.254060 | Structured | 0.312811 | Uncertain | 0.965 | 0.182 | 0.000 | -9.045 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 3.20 | Destabilizing | 0.0 | 2.83 | Destabilizing | 3.02 | Destabilizing | 2.02 | Destabilizing | 0.502 | Likely Pathogenic | -4.74 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.26 | Benign | 0.00 | Affected | 0.0877 | 0.0708 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1369A>C | S457R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S457R is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split opinion: benign calls come from REVEL, Rosetta, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; FoldX, Foldetta, and premPS are inconclusive. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates pathogenic, whereas Foldetta’s stability analysis is uncertain. Overall, the majority of evidence points toward a pathogenic impact. This conclusion is not contradicted by ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.164327 | Structured | 0.297330 | Uncertain | 0.909 | 0.159 | 0.000 | -10.882 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | -1.04 | Ambiguous | 0.0 | -0.06 | Likely Benign | -0.55 | Ambiguous | 0.74 | Ambiguous | 0.468 | Likely Benign | -4.47 | Deleterious | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 3.30 | Benign | 0.05 | Affected | 0.0781 | 0.3426 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1369A>T | S457C 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant S457C is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split assessment: benign predictions come from REVEL, FoldX, Foldetta, and FATHMM, whereas pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy methods give mixed results: AlphaMissense‑Optimized is inconclusive and therefore not used as evidence; the SGM‑Consensus majority vote (AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a benign effect. Overall, the majority of individual predictors lean toward pathogenicity, while the high‑accuracy Foldetta result suggests benign stability. Given the predominance of pathogenic calls and the lack of ClinVar evidence, the variant is most likely pathogenic, with no contradiction to ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.164327 | Structured | 0.297330 | Uncertain | 0.909 | 0.159 | 0.000 | -8.152 | Likely Pathogenic | 0.815 | Likely Pathogenic | Ambiguous | -0.12 | Likely Benign | 0.0 | -0.79 | Ambiguous | -0.46 | Likely Benign | 0.56 | Ambiguous | 0.497 | Likely Benign | -4.81 | Deleterious | 0.999 | Probably Damaging | 0.987 | Probably Damaging | 3.30 | Benign | 0.01 | Affected | 0.0865 | 0.6284 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||
| c.136C>A | P46T 2D AIThe SynGAP1 missense variant P46T is reported in ClinVar as “Not submitted” and is not present in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, all of which classify the change as benign. In contrast, polyPhen‑2 (HumDiv and HumVar) and SIFT uniformly predict a pathogenic impact. AlphaMissense‑Default remains uncertain, and no Foldetta stability assessment is available. High‑accuracy methods give a consistent benign signal: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports “Likely Benign,” and Foldetta data are missing. Overall, the majority of reliable predictors and the consensus analysis indicate that P46T is most likely benign, and this conclusion does not contradict the ClinVar status, which contains no pathogenic assertion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.390993 | Structured | 0.433588 | Uncertain | 0.549 | 0.741 | 0.375 | -4.329 | Likely Benign | 0.383 | Ambiguous | Likely Benign | 0.092 | Likely Benign | -0.68 | Neutral | 0.909 | Possibly Damaging | 0.901 | Possibly Damaging | 4.13 | Benign | 0.00 | Affected | 0.2194 | 0.6400 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||||||||||||
| c.1371T>A | S457R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S457R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, and FATHMM, whereas the majority of tools predict a pathogenic impact: SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, SGM‑Consensus confirms a likely pathogenic status, and Foldetta (combining FoldX‑MD and Rosetta outputs) yields an uncertain result. No other high‑confidence stability predictions are available. Overall, the preponderance of evidence from multiple independent predictors indicates that S457R is most likely pathogenic, and this assessment does not contradict any existing ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.164327 | Structured | 0.297330 | Uncertain | 0.909 | 0.159 | 0.000 | -10.882 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | -1.04 | Ambiguous | 0.0 | -0.06 | Likely Benign | -0.55 | Ambiguous | 0.74 | Ambiguous | 0.343 | Likely Benign | -4.47 | Deleterious | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 3.30 | Benign | 0.05 | Affected | 0.0781 | 0.3426 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1371T>G | S457R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S457R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, and FATHMM, whereas the majority of tools predict a pathogenic impact: SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, SGM‑Consensus confirms a likely pathogenic status, and Foldetta (combining FoldX‑MD and Rosetta outputs) yields an uncertain result. No other high‑confidence stability predictions are available. Overall, the preponderance of evidence from multiple independent predictors indicates that S457R is most likely pathogenic, and this assessment does not contradict any existing ClinVar annotation because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.164327 | Structured | 0.297330 | Uncertain | 0.909 | 0.159 | 0.000 | -10.882 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | -1.04 | Ambiguous | 0.0 | -0.06 | Likely Benign | -0.55 | Ambiguous | 0.74 | Ambiguous | 0.343 | Likely Benign | -4.47 | Deleterious | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 3.30 | Benign | 0.05 | Affected | 0.0781 | 0.3426 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1372A>C | T458P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T458P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are SIFT and FATHMM, while the remaining tools (REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, ESM1b, and the SGM‑Consensus) all predict a pathogenic impact; premPS is uncertain. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts a pathogenic effect. No prediction or folding stability result is missing or inconclusive. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.185198 | Structured | 0.294848 | Uncertain | 0.915 | 0.144 | 0.000 | -13.547 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 3.05 | Destabilizing | 0.2 | 5.36 | Destabilizing | 4.21 | Destabilizing | 0.78 | Ambiguous | 0.557 | Likely Pathogenic | -5.33 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.35 | Benign | 0.06 | Tolerated | 0.2098 | 0.5395 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1373C>A | T458K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T458K is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, Rosetta, SIFT, and FATHMM. Tools that agree on a pathogenic effect include SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain results from FoldX and premPS are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized predicting pathogenicity, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicting pathogenicity, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicting benign stability. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.185198 | Structured | 0.294848 | Uncertain | 0.915 | 0.144 | 0.000 | -13.734 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | -0.59 | Ambiguous | 0.1 | -0.26 | Likely Benign | -0.43 | Likely Benign | 0.80 | Ambiguous | 0.373 | Likely Benign | -5.23 | Deleterious | 0.999 | Probably Damaging | 0.973 | Probably Damaging | 3.40 | Benign | 0.14 | Tolerated | 0.1153 | 0.3326 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||
| c.1373C>T | T458I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T458I missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM. Those that predict a pathogenic impact are SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of tools (seven versus six) and the two high‑accuracy pathogenic predictions suggest the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar classification is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.185198 | Structured | 0.294848 | Uncertain | 0.915 | 0.144 | 0.000 | -9.436 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | -0.40 | Likely Benign | 0.1 | 0.29 | Likely Benign | -0.06 | Likely Benign | 0.50 | Likely Benign | 0.337 | Likely Benign | -4.76 | Deleterious | 0.999 | Probably Damaging | 0.989 | Probably Damaging | 3.40 | Benign | 0.09 | Tolerated | 0.0724 | 0.6128 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.1378A>G | K460E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K460E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, SIFT, and FATHMM. In contrast, a majority of tools predict a pathogenic impact: AlphaMissense‑Default, AlphaMissense‑Optimized, ESM1b, PROVEAN, polyPhen‑2 (HumDiv and HumVar), Rosetta, and premPS all indicate pathogenicity, while the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also reports “Likely Pathogenic.” High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. No prediction or folding‑stability result is missing or inconclusive. Overall, the preponderance of evidence points to a pathogenic effect for K460E, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.155435 | Structured | 0.289547 | Uncertain | 0.938 | 0.150 | 0.125 | -13.304 | Likely Pathogenic | 0.982 | Likely Pathogenic | Likely Pathogenic | 1.68 | Ambiguous | 0.0 | 2.02 | Destabilizing | 1.85 | Ambiguous | 1.05 | Destabilizing | 0.398 | Likely Benign | -3.40 | Deleterious | 0.999 | Probably Damaging | 0.991 | Probably Damaging | 3.34 | Benign | 0.09 | Tolerated | 0.3966 | 0.1022 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1379A>C | K460T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K460T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two consensus groups: benign predictions come from REVEL, SIFT, and FATHMM, whereas pathogenic predictions are made by PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score (Likely Pathogenic). Stability‑based methods (FoldX, Rosetta, premPS, Foldetta) yield uncertain or inconclusive results. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus majority vote (AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic, while Foldetta’s combined FoldX‑MD/Rosetta analysis remains inconclusive. Overall, the majority of evidence points to a pathogenic impact for K460T, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.155435 | Structured | 0.289547 | Uncertain | 0.938 | 0.150 | 0.125 | -11.187 | Likely Pathogenic | 0.967 | Likely Pathogenic | Likely Pathogenic | 1.23 | Ambiguous | 0.1 | 1.16 | Ambiguous | 1.20 | Ambiguous | 0.77 | Ambiguous | 0.208 | Likely Benign | -5.02 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.35 | Benign | 0.07 | Tolerated | 0.1858 | 0.3848 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1379A>T | K460M 2D 3DClick to see structure in 3D Viewer AISynGAP1 K460M is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from REVEL, Rosetta, Foldetta, premPS, and FATHMM, while pathogenic calls arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. Two tools give uncertain results: FoldX and AlphaMissense‑Optimized. When high‑accuracy methods are considered separately, AlphaMissense‑Optimized remains inconclusive, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of predictions lean toward pathogenicity, and this assessment does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.155435 | Structured | 0.289547 | Uncertain | 0.938 | 0.150 | 0.125 | -10.351 | Likely Pathogenic | 0.943 | Likely Pathogenic | Ambiguous | 0.61 | Ambiguous | 0.0 | 0.18 | Likely Benign | 0.40 | Likely Benign | 0.20 | Likely Benign | 0.252 | Likely Benign | -4.92 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.29 | Benign | 0.02 | Affected | 0.1148 | 0.4522 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.137C>A | P46H 2D AIThe SynGAP1 missense variant P46H is evaluated by multiple in silico tools. Consensus from SGM‑Consensus indicates a likely benign effect, and the variant is not reported in ClinVar or gnomAD. Functional predictors that agree on a benign outcome include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized. Predictors that flag a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also reports likely benign, while Foldetta stability analysis is unavailable. Overall, the majority of evidence points to a benign impact for P46H, and this conclusion is not contradicted by any ClinVar annotation. The variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.390993 | Structured | 0.433588 | Uncertain | 0.549 | 0.741 | 0.375 | -4.022 | Likely Benign | 0.428 | Ambiguous | Likely Benign | 0.097 | Likely Benign | -0.43 | Neutral | 0.992 | Probably Damaging | 0.977 | Probably Damaging | 4.10 | Benign | 0.00 | Affected | 0.2360 | 0.5229 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||||||||||||
| c.1382C>A | A461D 2D AIThe SynGAP1 missense variant A461D is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL and FATHMM. The majority of tools predict a pathogenic impact: premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). FoldX and Foldetta are inconclusive, providing no definitive evidence. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta remains uncertain. Overall, the preponderance of evidence indicates that A461D is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.179055 | Structured | 0.292531 | Uncertain | 0.936 | 0.151 | 0.125 | -14.918 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.89 | Ambiguous | 1.1 | 2.18 | Destabilizing | 1.54 | Ambiguous | 1.09 | Destabilizing | 0.477 | Likely Benign | -5.47 | Deleterious | 0.997 | Probably Damaging | 0.792 | Possibly Damaging | 3.32 | Benign | 0.01 | Affected | 0.1533 | 0.2016 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1382C>T | A461V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A461V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, FATHMM, AlphaMissense‑Optimized, and polyPhen2_HumVar. Tools that predict a pathogenic effect are PROVEAN, polyPhen2_HumDiv, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Because the majority of standard predictors (nine benign vs. three pathogenic) favor a benign outcome, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.292531 | Uncertain | 0.936 | 0.151 | 0.125 | -9.968 | Likely Pathogenic | 0.436 | Ambiguous | Likely Benign | -0.08 | Likely Benign | 0.3 | 0.34 | Likely Benign | 0.13 | Likely Benign | 0.02 | Likely Benign | 0.141 | Likely Benign | -3.05 | Deleterious | 0.983 | Probably Damaging | 0.273 | Benign | 3.43 | Benign | 0.66 | Tolerated | 0.0819 | 0.5078 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||
| c.1384A>G | K462E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K462E is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, and FATHMM. Tools that agree on a pathogenic effect include SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Rosetta and Foldetta are uncertain and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts likely pathogenic, and Foldetta is uncertain. Overall, the majority of predictions (7 pathogenic vs. 5 benign) and the high‑accuracy tools support a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.297737 | Uncertain | 0.921 | 0.159 | 0.125 | -14.696 | Likely Pathogenic | 0.967 | Likely Pathogenic | Likely Pathogenic | 0.34 | Likely Benign | 0.0 | 1.56 | Ambiguous | 0.95 | Ambiguous | 0.33 | Likely Benign | 0.433 | Likely Benign | -3.88 | Deleterious | 0.998 | Probably Damaging | 0.991 | Probably Damaging | 3.49 | Benign | 0.15 | Tolerated | 0.4145 | 0.1022 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1385A>C | K462T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K462T missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, SIFT, and FATHMM, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Four tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Optimized) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as uncertain. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is not contradicted by ClinVar status, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.297737 | Uncertain | 0.921 | 0.159 | 0.125 | -11.586 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 0.55 | Ambiguous | 0.0 | 1.08 | Ambiguous | 0.82 | Ambiguous | 0.30 | Likely Benign | 0.414 | Likely Benign | -5.82 | Deleterious | 0.999 | Probably Damaging | 1.000 | Probably Damaging | 3.47 | Benign | 0.08 | Tolerated | 0.2095 | 0.3257 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1385A>T | K462M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K462M missense variant is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, FATHMM, premPS, and the protein‑folding stability method Foldetta; pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus score. Rosetta’s output is inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also pathogenic, while Foldetta indicates a benign effect on protein stability. No evidence from ClinVar contradicts these findings. Overall, the majority of predictive tools and the consensus score support a pathogenic classification, suggesting that K462M is most likely pathogenic rather than benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.297737 | Uncertain | 0.921 | 0.159 | 0.125 | -12.837 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | -0.23 | Likely Benign | 0.1 | 0.86 | Ambiguous | 0.32 | Likely Benign | 0.17 | Likely Benign | 0.475 | Likely Benign | -5.82 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.40 | Benign | 0.02 | Affected | 0.1282 | 0.3932 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1386G>C | K462N 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant K462N is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions come from REVEL, SIFT, and FATHMM, whereas pathogenic predictions are returned by PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts Pathogenic, and the SGM Consensus also indicates Likely Pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, yields an uncertain result, providing no definitive evidence. Overall, the majority of high‑confidence predictors lean toward pathogenicity, contradicting the absence of a ClinVar classification. Thus, the variant is most likely pathogenic based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.297737 | Uncertain | 0.921 | 0.159 | 0.125 | -12.823 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.69 | Ambiguous | 0.1 | 1.67 | Ambiguous | 1.18 | Ambiguous | 0.90 | Ambiguous | 0.304 | Likely Benign | -4.83 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.42 | Benign | 0.07 | Tolerated | 3.37 | 34 | 0.3967 | 0.1242 | 0 | 1 | 0.4 | -14.07 | |||||||||||||||||||||||||||
| c.1386G>T | K462N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K462N is reported in gnomAD (ID 6‑33438291‑G‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions from REVEL, SIFT, and FATHMM; pathogenic predictions from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Stability‑based methods (FoldX, Rosetta, premPS) and Foldetta are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM Consensus as likely pathogenic, while Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for K462N. This conclusion is not contradicted by ClinVar, which contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.297737 | Uncertain | 0.921 | 0.159 | 0.125 | 6-33438291-G-T | 1 | 6.20e-7 | -12.823 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.69 | Ambiguous | 0.1 | 1.67 | Ambiguous | 1.18 | Ambiguous | 0.90 | Ambiguous | 0.303 | Likely Benign | -4.83 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.42 | Benign | 0.07 | Tolerated | 3.37 | 34 | 0.3967 | 0.1242 | 0 | 1 | 0.4 | -14.07 | ||||||||||||||||||||||||
| c.1388A>C | D463A 2D 3DClick to see structure in 3D Viewer AISynGAP1 D463A is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, and FATHMM, while those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Tools with inconclusive results are AlphaMissense‑Optimized and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the predictions are split, with an equal number of benign and pathogenic calls, and the high‑accuracy methods provide conflicting outcomes. Therefore, the variant is most likely of uncertain significance, which does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.260850 | Structured | 0.305622 | Uncertain | 0.940 | 0.176 | 0.000 | Uncertain | 1 | -8.607 | Likely Pathogenic | 0.894 | Likely Pathogenic | Ambiguous | -0.04 | Likely Benign | 0.1 | 0.96 | Ambiguous | 0.46 | Likely Benign | 0.43 | Likely Benign | 0.425 | Likely Benign | -6.96 | Deleterious | 0.978 | Probably Damaging | 0.602 | Possibly Damaging | 3.33 | Benign | 0.29 | Tolerated | 0.3591 | 0.5169 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||
| c.1400A>C | D467A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D467A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS and SIFT, whereas the majority of tools predict a pathogenic impact: SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools with uncertain or inconclusive results—FoldX, Rosetta, and Foldetta—do not provide decisive evidence. High‑accuracy assessments reinforce the pathogenic prediction: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta remains uncertain. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict the current ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.268042 | Structured | 0.329932 | Uncertain | 0.940 | 0.246 | 0.000 | -12.499 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.83 | Ambiguous | 0.1 | 0.79 | Ambiguous | 0.81 | Ambiguous | 0.23 | Likely Benign | 0.790 | Likely Pathogenic | -7.71 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.19 | Pathogenic | 0.07 | Tolerated | 0.3134 | 0.5117 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||
| c.1403T>A | M468K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M468K is listed in ClinVar (ID 642691.0) as Pathogenic and is not reported in gnomAD. All available in silico predictors classify the variant as pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect. High‑accuracy assessments confirm this: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. Thus, the variant is most likely pathogenic, and this prediction aligns with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.284882 | Structured | 0.339253 | Uncertain | 0.932 | 0.257 | 0.000 | Likely Pathogenic | 1 | -16.982 | Likely Pathogenic | 0.978 | Likely Pathogenic | Likely Pathogenic | 3.21 | Destabilizing | 0.1 | 3.30 | Destabilizing | 3.26 | Destabilizing | 2.57 | Destabilizing | 0.828 | Likely Pathogenic | -4.61 | Deleterious | 0.878 | Possibly Damaging | 0.922 | Probably Damaging | -1.34 | Pathogenic | 0.04 | Affected | 3.37 | 31 | 0.1576 | 0.0456 | 0 | -1 | -5.8 | -3.02 | 188.7 | 69.3 | 0.0 | 0.0 | -0.1 | 0.2 | X | X | Potentially Pathogenic | The thioether group of Met468, located in the middle of an α helix (res. Ala461–Phe476), interacts with hydrophobic residues (e.g., Phe464, Leu465, Leu489) in an inter-helix space formed by two other α helices (res. Ala461–Phe476, res. Thr488–Gly502). In the variant simulations, the positively charged side chain of Lys468 rotates outward to escape the hydrophobic niche, forming an H-bond with the hydroxyl group of the Ser471 side chain and a salt bridge with the carboxylate group of the Glu472 side chain. This residue swap also disrupts the methionine-aromatic stacking with the phenyl ring of the Phe464 side chain. Although no large-scale structural changes are observed during the variant simulations, the importance of hydrophobic packing suggests that the effects could be more pronounced during protein folding. | |||||||||||||||
| c.1403T>G | M468R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M468R is not reported in ClinVar and is absent from gnomAD. All evaluated in‑silico predictors classify the substitution as pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized indicates pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. Consequently, the variant is most likely pathogenic based on the consensus of predictive tools, and this assessment does not contradict any ClinVar status (none available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.284882 | Structured | 0.339253 | Uncertain | 0.932 | 0.257 | 0.000 | -16.180 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 2.66 | Destabilizing | 0.2 | 2.23 | Destabilizing | 2.45 | Destabilizing | 2.43 | Destabilizing | 0.837 | Likely Pathogenic | -4.64 | Deleterious | 0.939 | Possibly Damaging | 0.943 | Probably Damaging | -1.34 | Pathogenic | 0.00 | Affected | 0.1717 | 0.0637 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||
| c.1406C>A | A469D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A469D is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that assess the variant’s effect fall into two groups: the single benign prediction from SIFT, and a consensus of pathogenic predictions from the remaining 15 tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus). High‑accuracy methods further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also indicates pathogenic. Taken together, the overwhelming majority of evidence points to a pathogenic effect, which is consistent with the ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.278302 | Structured | 0.343926 | Uncertain | 0.910 | 0.276 | 0.000 | Uncertain | 1 | -14.643 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 5.09 | Destabilizing | 0.2 | 4.16 | Destabilizing | 4.63 | Destabilizing | 1.68 | Destabilizing | 0.738 | Likely Pathogenic | -3.48 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | -1.34 | Pathogenic | 0.21 | Tolerated | 3.37 | 34 | 0.1372 | 0.1583 | 0 | -2 | -5.3 | 44.01 | 237.0 | -58.2 | -0.2 | 0.1 | 0.8 | 0.1 | X | X | Potentially Pathogenic | The methyl group of Ala469, located in an α helix (res. Ala461–Phe476), interacts with hydrophobic residues (e.g., Trp572, Leu588, Met470) in an inter-helix space formed by two other α helices (res. Glu582–Ser604, res. Arg563–Gly580). In the variant simulations, Asp469 introduces a negatively charged and bulky side chain into the hydrophobic niche. Consequently, the side chain of Asp469 rotates outward, allowing the carboxylate group to form a salt bridge with the guanidinium group of Arg575 on the protein surface. This interaction affects the continuity of the parent α helix (Ala461–Phe476). Due to the importance of hydrophobic packing, the structural effects could be more pronounced during actual protein folding. | |||||||||||||||
| c.1406C>T | A469V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A469V missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect comprise FoldX, Rosetta, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and Foldetta; premPS and AlphaMissense‑Default are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta predicts pathogenic. With seven tools supporting pathogenicity versus four supporting benignity, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.278302 | Structured | 0.343926 | Uncertain | 0.910 | 0.276 | 0.000 | -8.858 | Likely Pathogenic | 0.459 | Ambiguous | Likely Benign | 2.30 | Destabilizing | 0.4 | 2.66 | Destabilizing | 2.48 | Destabilizing | -0.77 | Ambiguous | 0.426 | Likely Benign | 0.26 | Neutral | 0.997 | Probably Damaging | 0.976 | Probably Damaging | -1.20 | Pathogenic | 0.60 | Tolerated | 0.0827 | 0.5421 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||
| c.1409T>A | M470K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant M470K is not reported in ClinVar (status: None) and is absent from gnomAD. Prediction tools cluster into two groups: the single benign call comes from SIFT, while all other evaluated algorithms—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—label the change as pathogenic or likely pathogenic. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also reports pathogenic. Thus, the variant is most likely pathogenic based on the consensus of available predictions, and this assessment does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.298791 | Structured | 0.351497 | Uncertain | 0.908 | 0.272 | 0.000 | -14.327 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 2.77 | Destabilizing | 0.1 | 2.67 | Destabilizing | 2.72 | Destabilizing | 1.55 | Destabilizing | 0.823 | Likely Pathogenic | -5.42 | Deleterious | 0.923 | Possibly Damaging | 0.922 | Probably Damaging | -1.06 | Pathogenic | 0.20 | Tolerated | 0.1157 | 0.0656 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1409T>G | M470R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M470R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only SIFT, whereas all other evaluated algorithms—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No prediction or folding stability result is missing or inconclusive. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.298791 | Structured | 0.351497 | Uncertain | 0.908 | 0.272 | 0.000 | -13.161 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 2.75 | Destabilizing | 0.1 | 2.75 | Destabilizing | 2.75 | Destabilizing | 1.57 | Destabilizing | 0.823 | Likely Pathogenic | -5.47 | Deleterious | 0.963 | Probably Damaging | 0.943 | Probably Damaging | -1.19 | Pathogenic | 0.17 | Tolerated | 0.1399 | 0.0757 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||
| c.140G>C | R47P 2D AIThe SynGAP1 missense variant R47P is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is inconclusive (two pathogenic, two benign) and therefore unavailable as evidence. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no reported result for this variant and is likewise unavailable. High‑accuracy tools specifically indicate a benign prediction from AlphaMissense‑Optimized, while the other high‑accuracy methods (SGM Consensus, Foldetta) provide no definitive assessment. Overall, the balance of evidence (five pathogenic vs. four benign predictions) suggests the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.318242 | Structured | 0.436559 | Uncertain | 0.520 | 0.719 | 0.125 | -10.316 | Likely Pathogenic | 0.746 | Likely Pathogenic | Likely Benign | 0.195 | Likely Benign | -1.63 | Neutral | 0.841 | Possibly Damaging | 0.809 | Possibly Damaging | 4.02 | Benign | 0.00 | Affected | 0.2050 | 0.4956 | 0 | -2 | 2.9 | -59.07 | ||||||||||||||||||||||||||||||||||||||||
| c.1414G>A | E472K 2D AIThe SynGAP1 missense variant E472K is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that assess pathogenicity unanimously classify the variant as deleterious: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect; the only inconclusive result is from premPS, which is listed as uncertain. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a destabilizing, pathogenic effect. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.359300 | Uncertain | 0.878 | 0.231 | 0.000 | -15.214 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 2.01 | Destabilizing | 1.2 | 3.23 | Destabilizing | 2.62 | Destabilizing | 0.78 | Ambiguous | 0.670 | Likely Pathogenic | -3.95 | Deleterious | 0.996 | Probably Damaging | 0.987 | Probably Damaging | 2.33 | Pathogenic | 0.03 | Affected | 0.3077 | 0.5651 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1415A>C | E472A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E472A is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a pathogenic effect include REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; all these methods uniformly classify the change as deleterious. Tools that are inconclusive or uncertain for this variant are FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts a pathogenic outcome, the SGM Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates pathogenicity, while Foldetta’s stability analysis remains uncertain. Taken together, the overwhelming majority of evidence points to a pathogenic effect for E472A, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.359300 | Uncertain | 0.878 | 0.231 | 0.000 | -15.356 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 1.81 | Ambiguous | 0.3 | 0.67 | Ambiguous | 1.24 | Ambiguous | 0.69 | Ambiguous | 0.732 | Likely Pathogenic | -5.90 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.32 | Pathogenic | 0.01 | Affected | 0.4639 | 0.6315 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1415A>G | E472G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E472G is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that assess the variant’s effect largely agree on a deleterious outcome: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify it as pathogenic or likely pathogenic. Only premPS predicts a benign effect. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized reports pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. No predictions are missing or inconclusive. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.359300 | Uncertain | 0.878 | 0.231 | 0.000 | -15.239 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 3.06 | Destabilizing | 0.2 | 2.86 | Destabilizing | 2.96 | Destabilizing | 0.24 | Likely Benign | 0.744 | Likely Pathogenic | -6.83 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.30 | Pathogenic | 0.00 | Affected | 0.3343 | 0.4950 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1418T>C | V473A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V473A missense variant is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM. Tools that agree on a pathogenic effect include SGM‑Consensus (Likely Pathogenic), premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized are all uncertain or unavailable, providing no decisive evidence. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta as uncertain. Overall, the majority of available predictions support a pathogenic impact. This conclusion is consistent with the lack of ClinVar annotation (no contradiction). Thus, the variant is most likely pathogenic based on current computational evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.191378 | Structured | 0.362529 | Uncertain | 0.884 | 0.239 | 0.000 | -10.867 | Likely Pathogenic | 0.925 | Likely Pathogenic | Ambiguous | 1.88 | Ambiguous | 0.0 | 1.76 | Ambiguous | 1.82 | Ambiguous | 2.17 | Destabilizing | 0.485 | Likely Benign | -3.95 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.18 | Benign | 0.00 | Affected | 0.2529 | 0.2521 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1421A>C | D474A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D474A missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from FoldX, Rosetta, Foldetta, premPS, and SIFT, while pathogenic predictions arise from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic. High‑accuracy assessments further show AlphaMissense‑Optimized as Pathogenic, SGM Consensus as Likely Pathogenic, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) as Benign. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.373433 | Uncertain | 0.864 | 0.255 | 0.000 | -11.082 | Likely Pathogenic | 0.979 | Likely Pathogenic | Likely Pathogenic | 0.08 | Likely Benign | 0.0 | 0.15 | Likely Benign | 0.12 | Likely Benign | 0.17 | Likely Benign | 0.757 | Likely Pathogenic | -6.73 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.22 | Pathogenic | 0.22 | Tolerated | 0.3265 | 0.4354 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||
| c.1424G>C | R475P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R475P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: benign‑predicting algorithms are not present, while pathogenic‑predicting tools—including REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—all classify the substitution as pathogenic. High‑accuracy assessments corroborate this view: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports a likely pathogenic outcome, and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts a pathogenic effect. Consequently, the variant is most likely pathogenic based on the available computational evidence, and this assessment does not contradict any ClinVar status, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.175930 | Structured | 0.382696 | Uncertain | 0.852 | 0.261 | 0.000 | -16.637 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 2.93 | Destabilizing | 0.4 | 5.40 | Destabilizing | 4.17 | Destabilizing | 1.02 | Destabilizing | 0.835 | Likely Pathogenic | -6.53 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.43 | Pathogenic | 0.00 | Affected | 0.1943 | 0.3295 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1430T>A | M477K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477K is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are premPS and FATHMM. Predictions that are uncertain or inconclusive are FoldX, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is unavailable due to mixed or uncertain inputs. Overall, the majority of evidence points to a benign impact. Thus, the variant is most likely benign, and this conclusion does not contradict the current ClinVar status, which contains no report of pathogenicity. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -7.519 | In-Between | 0.553 | Ambiguous | Likely Benign | 0.54 | Ambiguous | 0.1 | 0.37 | Likely Benign | 0.46 | Likely Benign | 1.12 | Destabilizing | 0.371 | Likely Benign | -1.32 | Neutral | 0.254 | Benign | 0.122 | Benign | -1.15 | Pathogenic | 0.14 | Tolerated | 0.1817 | 0.0847 | 0 | -1 | -5.8 | -3.02 | ||||||||||||||||||||||||||||||
| c.1430T>G | M477R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M477R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are premPS, polyPhen‑2 HumDiv, and FATHMM. Tools with uncertain or inconclusive results are Rosetta, Foldetta, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction, while Foldetta remains uncertain and is not taken as evidence. Overall, the majority of reliable predictors indicate a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.268042 | Structured | 0.408680 | Uncertain | 0.761 | 0.250 | 0.000 | -6.786 | Likely Benign | 0.552 | Ambiguous | Likely Benign | 0.48 | Likely Benign | 0.2 | 0.77 | Ambiguous | 0.63 | Ambiguous | 1.24 | Destabilizing | 0.442 | Likely Benign | -1.12 | Neutral | 0.720 | Possibly Damaging | 0.242 | Benign | -1.22 | Pathogenic | 0.11 | Tolerated | 0.1901 | 0.0828 | 0 | -1 | -6.4 | 24.99 | ||||||||||||||||||||||||||||||
| c.1432G>A | E478K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E478K is not reported in ClinVar (ClinVar ID: None) and has no entries in gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. Tools that predict a pathogenic effect are SGM‑Consensus (Likely Pathogenic), PROVEAN, SIFT, ESM1b, and AlphaMissense‑Default. The high‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic, and Foldetta as benign. Overall, the majority of evidence (eight benign versus five pathogenic predictions) points to a benign impact. This conclusion does not contradict ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | -12.654 | Likely Pathogenic | 0.899 | Likely Pathogenic | Ambiguous | 0.23 | Likely Benign | 0.0 | 0.12 | Likely Benign | 0.18 | Likely Benign | -0.02 | Likely Benign | 0.309 | Likely Benign | -3.45 | Deleterious | 0.320 | Benign | 0.117 | Benign | 3.49 | Benign | 0.05 | Affected | 0.2066 | 0.6192 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1433A>C | E478A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E478A missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No predictions are missing or inconclusive. Overall, the majority of high‑confidence tools lean toward a benign classification, and this does not contradict the lack of ClinVar annotation. Thus, the variant is most likely benign based on current computational predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | -8.499 | Likely Pathogenic | 0.612 | Likely Pathogenic | Likely Benign | 0.46 | Likely Benign | 0.0 | 0.45 | Likely Benign | 0.46 | Likely Benign | 0.02 | Likely Benign | 0.342 | Likely Benign | -4.74 | Deleterious | 0.585 | Possibly Damaging | 0.505 | Possibly Damaging | 3.42 | Benign | 0.05 | Affected | 0.3516 | 0.5847 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1433A>G | E478G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E478G missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN and polyPhen‑2 HumDiv. Uncertain predictions come from Rosetta, ESM1b, AlphaMissense‑Default, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive due to equal benign and pathogenic signals and two uncertain calls; Foldetta likewise yields an uncertain result. Overall, the majority of evaluated tools (seven benign vs. two pathogenic) support a benign classification. This consensus does not contradict any ClinVar status, as no ClinVar assertion exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.264545 | Structured | 0.414660 | Uncertain | 0.787 | 0.249 | 0.000 | -7.897 | In-Between | 0.418 | Ambiguous | Likely Benign | 0.40 | Likely Benign | 0.0 | 0.73 | Ambiguous | 0.57 | Ambiguous | 0.02 | Likely Benign | 0.306 | Likely Benign | -4.91 | Deleterious | 0.923 | Possibly Damaging | 0.427 | Benign | 3.41 | Benign | 0.15 | Tolerated | 0.2758 | 0.5572 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||
| c.1436G>C | R479P 2D 3DClick to see structure in 3D Viewer AIClinVar lists the SynGAP1 R479P variant as Uncertain, and it is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, and FATHMM. Those that predict a pathogenic effect are FoldX, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and ESM1b. Predictions that are inconclusive are AlphaMissense‑Optimized and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) as pathogenic. Overall, the majority of tools and the high‑accuracy methods support a pathogenic classification, which is in contrast to the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.216401 | Structured | 0.419256 | Uncertain | 0.820 | 0.249 | 0.000 | Uncertain | 1 | -11.795 | Likely Pathogenic | 0.938 | Likely Pathogenic | Ambiguous | 2.86 | Destabilizing | 0.2 | 3.88 | Destabilizing | 3.37 | Destabilizing | 0.81 | Ambiguous | 0.277 | Likely Benign | -3.52 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.41 | Benign | 0.18 | Tolerated | 0.1993 | 0.3747 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||
| c.1439A>C | E480A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E480A is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only SIFT, whereas a majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (derived from the four pathogenic‑predicted tools above) as likely pathogenic, and Foldetta as uncertain. Because most evidence points to a deleterious effect, the variant is most likely pathogenic, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.216401 | Structured | 0.426867 | Uncertain | 0.798 | 0.250 | 0.000 | -13.192 | Likely Pathogenic | 0.931 | Likely Pathogenic | Ambiguous | 0.91 | Ambiguous | 0.1 | 1.15 | Ambiguous | 1.03 | Ambiguous | 0.55 | Ambiguous | 0.694 | Likely Pathogenic | -5.04 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | -1.25 | Pathogenic | 0.09 | Tolerated | 0.3468 | 0.6635 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1439A>G | E480G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E480G missense variant is not reported in ClinVar and has no gnomAD entry. Consensus from multiple in‑silico predictors indicates a pathogenic effect: SGM‑Consensus, REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify it as pathogenic. Predictions that are uncertain—FoldX, premPS, and AlphaMissense‑Optimized—do not provide evidence for benignity. High‑accuracy assessments further support pathogenicity: the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic, and AlphaMissense‑Optimized remains uncertain. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because no ClinVar record exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.216401 | Structured | 0.426867 | Uncertain | 0.798 | 0.250 | 0.000 | -11.651 | Likely Pathogenic | 0.839 | Likely Pathogenic | Ambiguous | 1.83 | Ambiguous | 0.1 | 2.34 | Destabilizing | 2.09 | Destabilizing | 0.65 | Ambiguous | 0.778 | Likely Pathogenic | -5.44 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.32 | Pathogenic | 0.03 | Affected | 0.2806 | 0.6172 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1441C>T | H481Y 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant H481Y is listed in ClinVar as benign (ClinVar ID 1543764.0) and is present in the gnomAD database (gnomAD ID 6‑33438473‑C‑T). Prediction tools that classify the variant as benign include REVEL, Rosetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. FoldX and Foldetta report uncertain stability effects. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta as uncertain. Taking all available evidence together, the variant is most likely benign, which is consistent with its ClinVar benign annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.257454 | Structured | 0.430977 | Uncertain | 0.764 | 0.247 | 0.000 | Likely Benign | 1 | 6-33438473-C-T | 16 | 9.91e-6 | -10.910 | Likely Pathogenic | 0.565 | Likely Pathogenic | Likely Benign | -0.53 | Ambiguous | 0.1 | -0.46 | Likely Benign | -0.50 | Ambiguous | 0.20 | Likely Benign | 0.256 | Likely Benign | -3.32 | Deleterious | 0.988 | Probably Damaging | 0.979 | Probably Damaging | 3.40 | Benign | 0.59 | Tolerated | 3.37 | 33 | 0.0610 | 0.3558 | 0 | 2 | 1.9 | 26.03 | 256.5 | -44.4 | 0.0 | 0.0 | 0.2 | 0.2 | X | X | Uncertain | The imidazole ring of the His481 side chain is located in a short helical structure (res. Glu480-Leu482) within an α-α loop connecting the two α-helices (res. Ala461-Phe476 and Leu489-Glu519) at the GAP-Ras interface. In the WT simulations, His481 alternately stacks against Arg485, Arg587, and Glu480 without a definite role. In the variant simulations, Tyr481 also alternately stacks with nearby arginine residues, including Arg485, Arg587, and Arg479. The interaction between Tyr481 and Arg479 affects the α-α loop, causing it to fold into a distorted helical structure, an effect that might be more pronounced during protein folding. Finally, the potential effect of the residue swap on SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | ||||||||||||
| c.1442A>C | H481P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H481P is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, and FATHMM. Those that predict a pathogenic effect comprise SGM Consensus (Likely Pathogenic), PolyPhen‑2 HumDiv, PolyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and PROVEAN. Tools with inconclusive results are Foldetta (Uncertain) and premPS (Uncertain). High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, while the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely pathogenic outcome; Foldetta remains uncertain. Overall, the majority of predictions lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.257454 | Structured | 0.430977 | Uncertain | 0.764 | 0.247 | 0.000 | -10.205 | Likely Pathogenic | 0.630 | Likely Pathogenic | Likely Benign | -0.48 | Likely Benign | 0.3 | 3.69 | Destabilizing | 1.61 | Ambiguous | 0.67 | Ambiguous | 0.385 | Likely Benign | -5.84 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.43 | Benign | 0.26 | Tolerated | 0.1979 | 0.3553 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||
| c.1448T>C | I483T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I483T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are REVEL and FATHMM, whereas the remaining tools—FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts a pathogenic effect. Based on the preponderance of pathogenic predictions and the absence of any benign consensus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.206376 | Structured | 0.415850 | Uncertain | 0.798 | 0.254 | 0.000 | -10.692 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 2.32 | Destabilizing | 0.0 | 2.05 | Destabilizing | 2.19 | Destabilizing | 1.77 | Destabilizing | 0.474 | Likely Benign | -4.24 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.15 | Benign | 0.05 | Affected | 0.0888 | 0.0840 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1453C>A | R485S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R485S is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include only FoldX, which scores the variant as benign. In contrast, the majority of tools predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Tools that return uncertain results are Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as inconclusive. Overall, the preponderance of evidence points to a pathogenic effect for R485S, which is consistent with the ClinVar “Uncertain” classification rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.188120 | Structured | 0.377409 | Uncertain | 0.805 | 0.246 | 0.125 | Uncertain | 1 | -15.603 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 0.40 | Likely Benign | 0.1 | 1.07 | Ambiguous | 0.74 | Ambiguous | 0.82 | Ambiguous | 0.609 | Likely Pathogenic | -5.97 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.93 | Pathogenic | 0.00 | Affected | 0.2968 | 0.3266 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||
| c.1454G>A | R485H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 R485H missense variant is listed in ClinVar as Benign (ClinVar ID 3707943.0) and is present in the gnomAD database (gnomAD ID 6‑33438486‑G‑A). Functional prediction tools that agree on a benign effect are Rosetta and Foldetta, while the majority of tools (REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic impact. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM Consensus (derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Benign. Overall, the preponderance of evidence points to a pathogenic effect, which contradicts the ClinVar benign classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.188120 | Structured | 0.377409 | Uncertain | 0.805 | 0.246 | 0.125 | Likely Benign | 1 | 6-33438486-G-A | 13 | 8.05e-6 | -13.628 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 0.77 | Ambiguous | 0.1 | 0.12 | Likely Benign | 0.45 | Likely Benign | 1.13 | Destabilizing | 0.618 | Likely Pathogenic | -4.97 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.93 | Pathogenic | 0.00 | Affected | 3.37 | 35 | 0.2990 | 0.1602 | 0 | 2 | 1.3 | -19.05 | ||||||||||||||||||||||
| c.1454G>C | R485P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R485P is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that assess pathogenicity all agree that the variant is deleterious: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return pathogenic or likely pathogenic scores. No tool in the dataset predicts a benign effect. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, predicts a pathogenic impact. Based on the unanimous pathogenic predictions and the absence of any benign calls, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.188120 | Structured | 0.377409 | Uncertain | 0.805 | 0.246 | 0.125 | -16.356 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 5.26 | Destabilizing | 0.3 | 6.86 | Destabilizing | 6.06 | Destabilizing | 0.56 | Ambiguous | 0.692 | Likely Pathogenic | -6.96 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.94 | Pathogenic | 0.00 | Affected | 0.2059 | 0.3941 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1456G>A | E486K 2D 3DClick to see structure in 3D Viewer AISynGAP1 E486K is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that classify the variant as benign include REVEL, FoldX, Rosetta, Foldetta, premPS, SIFT, and FATHMM. Those that predict pathogenicity are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus as pathogenic, while Foldetta (combining FoldX‑MD and Rosetta outputs) indicates a benign effect. Because the predictions are split evenly and the high‑accuracy tools are contradictory, the variant’s impact remains uncertain; thus, the variant is most likely pathogenic based on the high‑accuracy predictions, a conclusion that contradicts its ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.196879 | Structured | 0.358545 | Uncertain | 0.833 | 0.245 | 0.125 | Uncertain | 2 | -14.545 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 0.06 | Likely Benign | 0.1 | 0.37 | Likely Benign | 0.22 | Likely Benign | 0.41 | Likely Benign | 0.435 | Likely Benign | -3.58 | Deleterious | 1.000 | Probably Damaging | 0.988 | Probably Damaging | 3.40 | Benign | 0.12 | Tolerated | 3.37 | 35 | 0.1940 | 0.6392 | 0 | 1 | -0.4 | -0.94 | 206.8 | 52.1 | -0.3 | 0.1 | 0.2 | 0.0 | X | X | Uncertain | Glu486 is located in an α-α loop connecting the two α-helices (res. Ala461-Phe476 and Leu489-Glu519) at the GAP-Ras interface. It is adjacent to the arginine finger (Arg485) and is expected to closely interact with Ras. The residue swap could affect complex formation with the GTPase and its activation. In the WT simulations, the carboxylate group of Glu486 forms salt bridges with Arg485 and Arg475 on the preceding α-helix (res. Ala461-Phe476). In the variant simulations, Lys486 does not form any specific interactions. Although the amino group of the Lys486 side chain cannot form these salt bridges, no negative effects on the protein structure are observed. Nevertheless, the potential role of Glu486 in SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations, and no definite conclusions can be drawn. | |||||||||||||||
| c.1457A>C | E486A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E486A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, Rosetta, premPS, SIFT, and FATHMM, whereas pathogenic predictions are made by SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods give a consistent pathogenic signal: AlphaMissense‑Optimized predicts pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta outputs, is inconclusive and therefore unavailable. Overall, the majority of evidence supports a pathogenic effect. The prediction aligns with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.196879 | Structured | 0.358545 | Uncertain | 0.833 | 0.245 | 0.125 | -11.902 | Likely Pathogenic | 0.980 | Likely Pathogenic | Likely Pathogenic | 0.63 | Ambiguous | 0.0 | 0.32 | Likely Benign | 0.48 | Likely Benign | -0.03 | Likely Benign | 0.398 | Likely Benign | -5.17 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 3.44 | Benign | 0.39 | Tolerated | 0.3561 | 0.5859 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1457A>G | E486G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E486G missense change is not listed in ClinVar and has no gnomAD entry. Functional prediction tools that agree on a benign effect include REVEL, premPS, SIFT, and FATHMM. Those that predict a damaging outcome are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, and ESM1b. Predictions from FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is consistent with the absence of a ClinVar annotation, so there is no contradiction with existing clinical data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.196879 | Structured | 0.358545 | Uncertain | 0.833 | 0.245 | 0.125 | -12.488 | Likely Pathogenic | 0.924 | Likely Pathogenic | Ambiguous | 1.09 | Ambiguous | 0.1 | 1.59 | Ambiguous | 1.34 | Ambiguous | -0.14 | Likely Benign | 0.328 | Likely Benign | -5.46 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.80 | Benign | 0.40 | Tolerated | 0.2918 | 0.5385 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.145T>A | C49S 2D AIThe SynGAP1 missense variant C49S is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs. 2 pathogenic votes). Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of standard predictors lean toward pathogenicity, but the high‑accuracy tools do not provide definitive support. Thus, the variant is most likely pathogenic based on the current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.575 | Likely Benign | 0.704 | Likely Pathogenic | Likely Benign | 0.260 | Likely Benign | -3.07 | Deleterious | 0.462 | Possibly Damaging | 0.478 | Possibly Damaging | 3.91 | Benign | 0.00 | Affected | 0.3855 | 0.1686 | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.1460A>C | N487T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant N487T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of tools predict a pathogenic impact: PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as “Likely Pathogenic.” High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, SGM‑Consensus (majority vote) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is inconclusive. No evidence from FoldX, Rosetta, or premPS is available to alter this assessment. Overall, the preponderance of computational evidence indicates that the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.209395 | Structured | 0.338511 | Uncertain | 0.890 | 0.243 | 0.125 | -12.618 | Likely Pathogenic | 0.981 | Likely Pathogenic | Likely Pathogenic | 1.92 | Ambiguous | 0.1 | 1.94 | Ambiguous | 1.93 | Ambiguous | 0.68 | Ambiguous | 0.481 | Likely Benign | -5.97 | Deleterious | 0.987 | Probably Damaging | 0.980 | Probably Damaging | 2.78 | Benign | 0.05 | Affected | 0.1200 | 0.3441 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||
| c.1462A>C | T488P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T488P is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (gnomAD ID: none). Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while the remaining twelve tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict pathogenicity; premPS is uncertain and therefore not counted. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenicity. Based on the overwhelming consensus of these predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.206376 | Structured | 0.332663 | Uncertain | 0.928 | 0.233 | 0.125 | -13.432 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 4.40 | Destabilizing | 0.7 | 5.68 | Destabilizing | 5.04 | Destabilizing | 0.68 | Ambiguous | 0.505 | Likely Pathogenic | -5.70 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.24 | Benign | 0.00 | Affected | 0.1518 | 0.3557 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1463C>A | T488K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T488K is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, while the remaining eleven tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) all classify the variant as pathogenic. The high‑accuracy methods—AlphaMissense‑Optimized, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta (combining FoldX‑MD and Rosetta outputs)—all predict pathogenicity. No prediction or folding‑stability result is missing or inconclusive. Based on the consensus of these tools, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.206376 | Structured | 0.332663 | Uncertain | 0.928 | 0.233 | 0.125 | -14.391 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 2.09 | Destabilizing | 0.2 | 2.33 | Destabilizing | 2.21 | Destabilizing | 0.90 | Ambiguous | 0.692 | Likely Pathogenic | -5.64 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 3.24 | Benign | 0.00 | Affected | 0.0871 | 0.2104 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||
| c.1468G>A | A490T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A490T is listed in gnomAD (variant ID 6‑33438500‑G‑A) but has no ClinVar entry. Prediction tools that agree on a pathogenic effect include REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Tools that are inconclusive or uncertain are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. No tool predicts a benign outcome. High‑accuracy assessments show that the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenicity, while AlphaMissense‑Optimized remains uncertain and Foldetta is also uncertain. Based on the preponderance of pathogenic predictions and the lack of any benign calls, the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.120615 | Structured | 0.322979 | Uncertain | 0.938 | 0.210 | 0.125 | 6-33438500-G-A | 1 | 6.20e-7 | -10.266 | Likely Pathogenic | 0.892 | Likely Pathogenic | Ambiguous | 0.80 | Ambiguous | 0.2 | 1.70 | Ambiguous | 1.25 | Ambiguous | 1.00 | Destabilizing | 0.821 | Likely Pathogenic | -3.87 | Deleterious | 0.998 | Probably Damaging | 0.993 | Probably Damaging | -1.34 | Pathogenic | 0.03 | Affected | 3.37 | 35 | 0.1065 | 0.4495 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||
| c.1469C>A | A490D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A490D is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that assess pathogenicity are unanimous: AlphaMissense‑Default, AlphaMissense‑Optimized, ESM1b, FATHMM, PROVEAN, SIFT, polyPhen‑2 (HumDiv and HumVar), REVEL, premPS, FoldX, Rosetta, and Foldetta all classify the variant as pathogenic. No tool predicts a benign effect, so the benign group is empty. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) also reports pathogenic. Based on the collective predictions, the variant is most likely pathogenic, and this assessment is consistent with the absence of ClinVar evidence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.120615 | Structured | 0.322979 | Uncertain | 0.938 | 0.210 | 0.125 | -16.643 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 3.33 | Destabilizing | 0.1 | 4.23 | Destabilizing | 3.78 | Destabilizing | 1.33 | Destabilizing | 0.946 | Likely Pathogenic | -5.70 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.48 | Pathogenic | 0.00 | Affected | 0.1532 | 0.1741 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1469C>T | A490V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A490V missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity uniformly favor a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all indicate pathogenicity. No tool predicts a benign outcome. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is inconclusive, the SGM‑Consensus remains likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also inconclusive. Overall, the consensus of the majority of algorithms points to a pathogenic effect, and this conclusion does not conflict with the absence of ClinVar annotation. Thus, the variant is most likely pathogenic, with no contradiction from ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.120615 | Structured | 0.322979 | Uncertain | 0.938 | 0.210 | 0.125 | -10.348 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 0.73 | Ambiguous | 0.3 | 0.62 | Ambiguous | 0.68 | Ambiguous | 0.69 | Ambiguous | 0.817 | Likely Pathogenic | -3.93 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | -1.26 | Pathogenic | 0.04 | Affected | 0.0983 | 0.4213 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.146G>A | C49Y 2D AIThe SynGAP1 missense variant C49Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the majority of algorithms (PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default) predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is classified as Likely Pathogenic, and Foldetta results are unavailable. Overall, the preponderance of evidence points to a pathogenic effect for C49Y. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with existing clinical database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -11.097 | Likely Pathogenic | 0.949 | Likely Pathogenic | Ambiguous | 0.312 | Likely Benign | -3.38 | Deleterious | 0.676 | Possibly Damaging | 0.761 | Possibly Damaging | 3.85 | Benign | 0.00 | Affected | 0.1016 | 0.3172 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||||||||||||
| c.146G>C | C49S 2D AIThe SynGAP1 missense variant C49S is not reported in ClinVar (ClinVar ID = None) and is absent from gnomAD (gnomAD ID = None). Prediction tools that agree on a benign effect include REVEL, ESM1b, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive (2 benign vs 2 pathogenic). Foldetta results are unavailable. Overall, the majority of conventional tools (five pathogenic vs four benign) lean toward a pathogenic interpretation, but the single high‑accuracy benign prediction and the inconclusive SGM Consensus leave the assessment uncertain. Thus, the variant is most likely pathogenic based on the prevailing predictions, and this does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | 0.209395 | Structured | 0.445316 | Uncertain | 0.541 | 0.704 | 0.000 | -6.575 | Likely Benign | 0.704 | Likely Pathogenic | Likely Benign | 0.224 | Likely Benign | -3.07 | Deleterious | 0.462 | Possibly Damaging | 0.478 | Possibly Damaging | 3.91 | Benign | 0.00 | Affected | 0.3855 | 0.1686 | 0 | -1 | -3.3 | -16.06 | ||||||||||||||||||||||||||||||||||||||||
| c.1471A>C | T491P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T491P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while premPS remains uncertain. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenic. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.064632 | Structured | 0.325158 | Uncertain | 0.929 | 0.188 | 0.125 | -13.603 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | 3.19 | Destabilizing | 0.6 | 4.32 | Destabilizing | 3.76 | Destabilizing | 0.96 | Ambiguous | 0.882 | Likely Pathogenic | -5.89 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.49 | Pathogenic | 0.00 | Affected | 0.2024 | 0.3759 | 0 | -1 | -0.9 | -3.99 | |||||||||||||||||||||||||||||
| c.1471A>G | T491A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T491A is not reported in ClinVar (ClinVar status: none) but is present in gnomAD (gnomAD ID: 6‑33438503‑A‑G). Prediction tools that agree on a benign effect include FoldX, Rosetta, and Foldetta. Tools that predict a pathogenic effect include SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of predictions (10 pathogenic vs. 3 benign) indicate a likely pathogenic impact, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.064632 | Structured | 0.325158 | Uncertain | 0.929 | 0.188 | 0.125 | 6-33438503-A-G | 1 | 6.20e-7 | -11.033 | Likely Pathogenic | 0.934 | Likely Pathogenic | Ambiguous | 0.10 | Likely Benign | 0.3 | -0.27 | Likely Benign | -0.09 | Likely Benign | 1.06 | Destabilizing | 0.851 | Likely Pathogenic | -4.82 | Deleterious | 0.998 | Probably Damaging | 0.989 | Probably Damaging | -1.47 | Pathogenic | 0.01 | Affected | 3.37 | 35 | 0.3903 | 0.2874 | 0 | 1 | 2.5 | -30.03 | ||||||||||||||||||||||||
| c.1472C>A | T491N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T491N is not reported in ClinVar and is absent from gnomAD. Prediction tools that reach consensus on pathogenicity include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, all of which classify the substitution as pathogenic. No tool predicts a benign effect. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports it as likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, yields an uncertain result. The remaining tools, Rosetta and Foldetta, provide inconclusive evidence. Overall, the collective evidence indicates that T491N is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.064632 | Structured | 0.325158 | Uncertain | 0.929 | 0.188 | 0.125 | -11.952 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 2.39 | Destabilizing | 0.4 | 1.44 | Ambiguous | 1.92 | Ambiguous | 1.43 | Destabilizing | 0.842 | Likely Pathogenic | -4.92 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.50 | Pathogenic | 0.02 | Affected | 0.0959 | 0.2902 | 0 | 0 | -2.8 | 13.00 | |||||||||||||||||||||||||||||
| c.1472C>T | T491I 2D AIThe SynGAP1 missense variant T491I is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only premPS; all other evaluated algorithms (SGM‑Consensus, REVEL, Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact, while FoldX is uncertain and therefore not counted in either group. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized reports a pathogenic change, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. Based on the collective predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.064632 | Structured | 0.325158 | Uncertain | 0.929 | 0.188 | 0.125 | -12.525 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 1.48 | Ambiguous | 1.8 | 3.73 | Destabilizing | 2.61 | Destabilizing | 0.49 | Likely Benign | 0.915 | Likely Pathogenic | -5.89 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.42 | Pathogenic | 0.00 | Affected | 0.0898 | 0.4942 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.1475A>C | K492T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K492T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FATHMM, whereas the majority—SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default and AlphaMissense‑Optimized—predict a pathogenic impact. Four methods (FoldX, Rosetta, Foldetta, premPS) return uncertain results. High‑accuracy predictors give a consistent pathogenic signal: AlphaMissense‑Optimized is pathogenic, SGM‑Consensus is likely pathogenic, and Foldetta remains uncertain. Overall, the evidence strongly favors a pathogenic classification, and this assessment does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.061840 | Structured | 0.327121 | Uncertain | 0.947 | 0.192 | 0.000 | -16.126 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 1.09 | Ambiguous | 0.1 | 0.88 | Ambiguous | 0.99 | Ambiguous | 0.98 | Ambiguous | 0.595 | Likely Pathogenic | -5.97 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.96 | Benign | 0.04 | Affected | 0.1691 | 0.2825 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1478C>A | A493D 2D AISynGAP1 missense variant A493D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as pathogenic. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenicity. Consequently, the variant is most likely pathogenic, and this assessment is consistent with the absence of a ClinVar entry (no contradiction). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.340081 | Uncertain | 0.966 | 0.182 | 0.000 | -16.834 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 4.45 | Destabilizing | 1.5 | 3.59 | Destabilizing | 4.02 | Destabilizing | 1.60 | Destabilizing | 0.925 | Likely Pathogenic | -5.25 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.41 | Pathogenic | 0.01 | Affected | 0.1437 | 0.1541 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1478C>T | A493V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A493V missense variant has no ClinVar entry and is reported in gnomAD (6‑33438510‑C‑T). Functional prediction tools largely agree on a deleterious effect: pathogenic calls come from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Benign predictions are limited to SIFT and Foldetta. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain, the SGM‑Consensus indicates likely pathogenic, and Foldetta predicts benign stability. No other tools provide conclusive evidence. Overall, the preponderance of pathogenic predictions, including the consensus and multiple independent algorithms, suggests the variant is most likely pathogenic. This conclusion does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.340081 | Uncertain | 0.966 | 0.182 | 0.000 | 6-33438510-C-T | 3 | 1.86e-6 | -12.511 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 0.56 | Ambiguous | 0.1 | -0.67 | Ambiguous | -0.06 | Likely Benign | 0.91 | Ambiguous | 0.735 | Likely Pathogenic | -3.84 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | -1.31 | Pathogenic | 0.10 | Tolerated | 3.37 | 35 | 0.0877 | 0.3860 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||
| c.1481T>C | I494T 2D AIThe SynGAP1 missense variant I494T is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include only AlphaMissense‑Optimized. All other evaluated tools—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—predict a pathogenic impact. High‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts likely pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No predictions are missing or inconclusive. Based on the overwhelming majority of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.155435 | Structured | 0.353330 | Uncertain | 0.941 | 0.157 | 0.000 | -10.033 | Likely Pathogenic | 0.754 | Likely Pathogenic | Likely Benign | 2.45 | Destabilizing | 0.7 | 2.12 | Destabilizing | 2.29 | Destabilizing | 1.73 | Destabilizing | 0.907 | Likely Pathogenic | -4.61 | Deleterious | 1.000 | Probably Damaging | 0.995 | Probably Damaging | -1.38 | Pathogenic | 0.01 | Affected | 0.1003 | 0.0640 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1484A>C | E495A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E495A missense variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that indicate a benign effect include FoldX, while the remaining evaluated algorithms (REVEL, SIFT, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, PROVEAN, AlphaMissense‑Default, and the SGM‑Consensus) all predict a pathogenic outcome. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Likely Pathogenic, and Foldetta as uncertain. No contradictory evidence is present from ClinVar or population databases. Overall, the preponderance of predictions supports a pathogenic classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.164327 | Structured | 0.364496 | Uncertain | 0.933 | 0.161 | 0.000 | -9.229 | Likely Pathogenic | 0.946 | Likely Pathogenic | Ambiguous | 0.42 | Likely Benign | 0.1 | 0.85 | Ambiguous | 0.64 | Ambiguous | 0.69 | Ambiguous | 0.834 | Likely Pathogenic | -5.84 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.38 | Pathogenic | 0.01 | Affected | 0.2711 | 0.4259 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1486G>A | E496K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E496K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FoldX, whereas the majority of tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) predict a pathogenic impact. Uncertain predictions come from Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as inconclusive. Overall, the evidence strongly favors a pathogenic classification for E496K, and this conclusion does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.383296 | Uncertain | 0.945 | 0.179 | 0.000 | -15.795 | Likely Pathogenic | 0.961 | Likely Pathogenic | Likely Pathogenic | 0.38 | Likely Benign | 0.1 | 1.77 | Ambiguous | 1.08 | Ambiguous | 0.76 | Ambiguous | 0.743 | Likely Pathogenic | -3.58 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | -1.40 | Pathogenic | 0.04 | Affected | 0.1810 | 0.3528 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1487A>C | E496A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E496A is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools show mixed results: benign calls come from Rosetta, Foldetta, premPS, SIFT, and AlphaMissense‑Optimized, while pathogenic calls are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). FoldX is inconclusive. High‑accuracy methods give a split verdict: AlphaMissense‑Optimized predicts benign, SGM‑Consensus predicts pathogenic, and Foldetta predicts benign. Overall, the majority of predictions lean toward pathogenicity, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.383296 | Uncertain | 0.945 | 0.179 | 0.000 | Uncertain | 1 | -11.159 | Likely Pathogenic | 0.598 | Likely Pathogenic | Likely Benign | 0.68 | Ambiguous | 0.0 | 0.23 | Likely Benign | 0.46 | Likely Benign | 0.47 | Likely Benign | 0.681 | Likely Pathogenic | -4.87 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.41 | Pathogenic | 0.09 | Tolerated | 0.2806 | 0.3548 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||
| c.1487A>G | E496G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E496G missense variant is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that assess the variant’s effect fall into two groups: no tool predicts a benign outcome, while eight tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) all predict a pathogenic effect. The SGM‑Consensus, which is a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates a likely pathogenic outcome. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain, the SGM‑Consensus remains likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Overall, the preponderance of evidence points to a pathogenic effect, contradicting the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.383296 | Uncertain | 0.945 | 0.179 | 0.000 | Uncertain | 1 | -13.529 | Likely Pathogenic | 0.850 | Likely Pathogenic | Ambiguous | 1.83 | Ambiguous | 0.1 | 1.76 | Ambiguous | 1.80 | Ambiguous | 0.92 | Ambiguous | 0.825 | Likely Pathogenic | -6.16 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.45 | Pathogenic | 0.02 | Affected | 3.37 | 35 | 0.2435 | 0.3473 | 0 | -2 | 3.1 | -72.06 | 173.9 | 103.1 | 0.0 | 0.0 | -0.7 | 0.0 | X | X | Potentially Pathogenic | Glu496 is located in the α-helix (res. Leu489-Glu519), and its carboxylate group forms salt bridges with the neighbouring residues Lys492 and Arg499 in the WT simulations. Glu496 also forms a hydrogen bond with Ser449 on an opposing helix (res. Val441-Ser457). In the variant simulations, Gly496 cannot form these salt bridges, which could weaken the secondary structure. Additionally, the loss of the hydrogen bond with Ser449 on the opposite helix can weaken the tertiary structure assembly. Moreover, glycine is an α-helix breaker, and it is seen to weaken the integrity of the helix as the hydrogen bonding between the backbone atoms of Gly496 and Ala493 breaks down. Also, due to its location at the GAP-Ras interface, the interaction of Glu496 with Arg499 and Lys492 might play a role in complex association and stability, which cannot be fully addressed using the SynGAP solvent-only simulations. | |||||||||||||||
| c.1489T>C | Y497H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y497H is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that assess pathogenicity all agree that the variant is deleterious: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify it as pathogenic. No tool predicts a benign effect. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, indicates a destabilizing, pathogenic effect. All available predictions are concordant and supportive. Based on these computational assessments, the variant is most likely pathogenic, and this conclusion does not contradict any existing ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.393608 | Uncertain | 0.950 | 0.181 | 0.000 | -10.970 | Likely Pathogenic | 0.981 | Likely Pathogenic | Likely Pathogenic | 2.46 | Destabilizing | 0.1 | 2.40 | Destabilizing | 2.43 | Destabilizing | 1.29 | Destabilizing | 0.819 | Likely Pathogenic | -4.94 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.65 | Pathogenic | 0.02 | Affected | 0.1983 | 0.0612 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.1490A>G | Y497C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y497C is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that assess pathogenicity all return a deleterious signal: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all predict pathogenic. No tool reports a benign outcome. High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain,” SGM Consensus as “Likely Pathogenic,” and Foldetta (combining FoldX‑MD and Rosetta outputs) as “Pathogenic.” Overall, the variant is most likely pathogenic based on the consensus of predictive algorithms, which contradicts the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.393608 | Uncertain | 0.950 | 0.181 | 0.000 | Uncertain | 1 | -11.872 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 3.88 | Destabilizing | 0.1 | 4.76 | Destabilizing | 4.32 | Destabilizing | 1.40 | Destabilizing | 0.806 | Likely Pathogenic | -8.82 | Deleterious | 1.000 | Probably Damaging | 0.995 | Probably Damaging | -1.65 | Pathogenic | 0.03 | Affected | 3.37 | 35 | 0.2905 | 0.1488 | 0 | -2 | 3.8 | -60.04 | 209.9 | 59.1 | -0.1 | 0.0 | -0.3 | 0.1 | X | X | Potentially Pathogenic | Tyr497 is located in the α-helix (res. Leu489-Glu519) within the inter-helix space of four α-helices (res. Leu489-Ile501, res. Val441-Ser457, res. Arg563-Glu578, res. Ala461-Val473). In the WT simulations, the phenol ring of Tyr497 hydrophobically packs with other residues in the inter-helix space (e.g., Leu465, Leu565, Val568). The hydroxyl group of Tyr497 also alternately forms hydrogen bonds with the carboxylate side chain of Gln456 and the backbone carbonyl of Glu564. Thus, Tyr497 plays a role in the folding and maintenance of the tertiary structure assembly between these four helices.In the variant simulations, the comparatively smaller residue, Cys497, cannot maintain any of the interactions seen with Tyr497 in the WT. Although no severe deleterious consequences are observed in the simulations, the structural effects could be more pronounced during actual protein folding. Indeed, the tertiary structure is seen to slightly break apart in the variant simulations. | |||||||||||||||
| c.1493T>A | M498K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M498K is not reported in ClinVar and is absent from gnomAD. Prediction tools largely agree on a deleterious effect: benign calls come only from polyPhen‑2 HumDiv and HumVar, whereas the remaining 15 tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus) all predict pathogenicity. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized scores the variant as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) labels it likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts a destabilizing, pathogenic effect. Taken together, the overwhelming majority of evidence indicates that M498K is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.399612 | Uncertain | 0.932 | 0.158 | 0.000 | -11.622 | Likely Pathogenic | 0.983 | Likely Pathogenic | Likely Pathogenic | 2.82 | Destabilizing | 0.1 | 2.86 | Destabilizing | 2.84 | Destabilizing | 1.79 | Destabilizing | 0.867 | Likely Pathogenic | -4.53 | Deleterious | 0.287 | Benign | 0.120 | Benign | -1.37 | Pathogenic | 0.00 | Affected | 0.1281 | 0.0656 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1493T>G | M498R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M498R is listed in ClinVar as Pathogenic (ClinVar ID 3907767.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include only polyPhen‑2 HumVar; all other evaluated algorithms (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. No predictions or stability results are missing or inconclusive. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, and this conclusion aligns with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.399612 | Uncertain | 0.932 | 0.158 | 0.000 | Likely Pathogenic | 1 | -8.812 | Likely Pathogenic | 0.988 | Likely Pathogenic | Likely Pathogenic | 3.85 | Destabilizing | 0.2 | 2.35 | Destabilizing | 3.10 | Destabilizing | 1.76 | Destabilizing | 0.869 | Likely Pathogenic | -4.53 | Deleterious | 0.464 | Possibly Damaging | 0.120 | Benign | -1.36 | Pathogenic | 0.00 | Affected | 0.1482 | 0.0757 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||
| c.149T>C | I50T 2D AIThe SynGAP1 missense variant I50T is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools largely agree that the change is benign: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM all classify it as benign. In contrast, SIFT and AlphaMissense‑Default predict a pathogenic effect. AlphaMissense‑Optimized returns an uncertain result, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available prediction for this variant. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, yields a “Likely Benign” verdict. High‑accuracy assessments therefore point to a benign outcome: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus is benign, and Foldetta data are missing. Based on the collective evidence, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.295083 | Structured | 0.449965 | Uncertain | 0.545 | 0.708 | 0.000 | -5.121 | Likely Benign | 0.949 | Likely Pathogenic | Ambiguous | 0.128 | Likely Benign | -1.61 | Neutral | 0.092 | Benign | 0.037 | Benign | 3.76 | Benign | 0.00 | Affected | 0.0950 | 0.0708 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||||||||||||
| c.14G>C | R5P 2D AIThe SynGAP1 missense variant R5P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the variant as benign. Only SIFT predicts a pathogenic outcome. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized is benign, and the SGM‑Consensus (majority vote) is likely benign; Foldetta results are unavailable. Overall, the preponderance of evidence indicates that R5P is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.595080 | Disordered | 0.547847 | Binding | 0.363 | 0.920 | 0.750 | -3.438 | Likely Benign | 0.189 | Likely Benign | Likely Benign | 0.286 | Likely Benign | -0.35 | Neutral | 0.233 | Benign | 0.013 | Benign | 4.10 | Benign | 0.00 | Affected | 0.2294 | 0.5530 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||||||||||||
| c.1502T>C | I501T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I501T is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and premPS, while Rosetta remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Overall, the majority of predictions lean toward a benign effect, and this does not contradict the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.079919 | Structured | 0.366596 | Uncertain | 0.886 | 0.153 | 0.000 | Uncertain | 1 | -5.996 | Likely Benign | 0.252 | Likely Benign | Likely Benign | 2.40 | Destabilizing | 0.1 | 1.81 | Ambiguous | 2.11 | Destabilizing | 1.57 | Destabilizing | 0.362 | Likely Benign | -3.48 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.44 | Benign | 0.16 | Tolerated | 3.37 | 35 | 0.0972 | 0.0640 | 0 | -1 | -5.2 | -12.05 | 214.5 | 26.9 | 0.0 | 0.0 | 0.5 | 0.0 | X | Potentially Pathogenic | Ile501 is located near a hinge in the middle of an α-helix (res. Leu489-Glu519). The sec-butyl side chain of Ile501 is hydrophobically packed with other residues in the inter-helix space (e.g., Leu500, Tyr497, Phe679) in the WT simulations. In the variant simulations, the hydroxyl group of Thr501 forms a hydrogen bond with the backbone atoms of Tyr497 on the same α-helix, which may weaken the α-helix integrity. Additionally, the polar hydroxyl group of Thr501 is not suitable for the hydrophobic inter-helix space, and thus, the residue swap could affect protein folding. However, Ile501 is followed by Gly502, which facilitates a hinge in the middle of the α-helix, making further weakening caused by Thr501 unlikely to be harmful to the α-helix integrity. | ||||||||||||||||
| c.1508A>C | Q503P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Q503P is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only AlphaMissense‑Optimized. Those that predict a pathogenic effect comprise SGM‑Consensus, REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and FATHMM; the remaining tools (FoldX, premPS, AlphaMissense‑Default) are inconclusive. High‑accuracy methods give a consistent pathogenic signal: AlphaMissense‑Optimized predicts benign, but the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and Foldetta both predict pathogenic. Because the majority of evidence points to a deleterious effect, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.040537 | Structured | 0.322935 | Uncertain | 0.848 | 0.168 | 0.000 | -10.915 | Likely Pathogenic | 0.414 | Ambiguous | Likely Benign | 1.06 | Ambiguous | 0.7 | 5.94 | Destabilizing | 3.50 | Destabilizing | 0.75 | Ambiguous | 0.860 | Likely Pathogenic | -5.20 | Deleterious | 0.999 | Probably Damaging | 0.991 | Probably Damaging | -1.53 | Pathogenic | 0.03 | Affected | 0.2076 | 0.3610 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||||
| c.1510A>G | K504E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K504E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise SGM‑Consensus (Likely Pathogenic), premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. FoldX, Rosetta, and Foldetta give uncertain results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic; Foldetta remains uncertain. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.028107 | Structured | 0.304984 | Uncertain | 0.850 | 0.189 | 0.000 | -9.890 | Likely Pathogenic | 0.587 | Likely Pathogenic | Likely Benign | 0.63 | Ambiguous | 0.4 | 0.78 | Ambiguous | 0.71 | Ambiguous | 1.06 | Destabilizing | 0.386 | Likely Benign | -3.40 | Deleterious | 0.924 | Possibly Damaging | 0.674 | Possibly Damaging | -1.25 | Pathogenic | 0.21 | Tolerated | 0.2583 | 0.0650 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1511A>C | K504T 2D 3DClick to see structure in 3D Viewer AISynGAP1 K504T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Other tools (AlphaMissense‑Default, Foldetta, premPS, Rosetta) were inconclusive and are not considered evidence. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of predictions support a pathogenic classification, and this is not contradicted by the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.028107 | Structured | 0.304984 | Uncertain | 0.850 | 0.189 | 0.000 | -9.572 | Likely Pathogenic | 0.494 | Ambiguous | Likely Benign | 0.12 | Likely Benign | 0.3 | -0.85 | Ambiguous | -0.37 | Likely Benign | 0.84 | Ambiguous | 0.498 | Likely Benign | -5.36 | Deleterious | 0.961 | Probably Damaging | 0.990 | Probably Damaging | -1.44 | Pathogenic | 0.10 | Tolerated | 0.1482 | 0.2619 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1514A>G | Y505C 2D AIThe SynGAP1 missense variant Y505C is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only FATHMM, whereas all other evaluated algorithms—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy methods further support this: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates a likely pathogenic outcome; and Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, also classifies the variant as pathogenic. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.047319 | Structured | 0.292227 | Uncertain | 0.909 | 0.188 | 0.000 | -11.784 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | 3.41 | Destabilizing | 0.5 | 4.37 | Destabilizing | 3.89 | Destabilizing | 2.32 | Destabilizing | 0.578 | Likely Pathogenic | -8.95 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.59 | Benign | 0.00 | Affected | 0.3203 | 0.1814 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||
| c.1516C>T | L506F 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L506F is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include only REVEL, whereas the majority of tools (FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) predict a pathogenic impact; premPS and AlphaMissense‑Optimized are inconclusive. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is uncertain, the SGM Consensus (derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, which is consistent with its ClinVar “Uncertain” classification and does not contradict the available data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.034884 | Structured | 0.279180 | Uncertain | 0.924 | 0.196 | 0.000 | Uncertain | 1 | -11.262 | Likely Pathogenic | 0.883 | Likely Pathogenic | Ambiguous | 4.92 | Destabilizing | 0.8 | 5.76 | Destabilizing | 5.34 | Destabilizing | 0.91 | Ambiguous | 0.464 | Likely Benign | -3.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 1.62 | Pathogenic | 0.01 | Affected | 3.37 | 35 | 0.0566 | 0.1471 | 0 | 2 | -1.0 | 34.02 | |||||||||||||||||||||||||
| c.1519A>G | K507E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K507E missense change is not listed in ClinVar and has no gnomAD allele, indicating it is not a common polymorphism. Functional prediction tools cluster into two groups: benign predictions come from Rosetta, PROVEAN, SIFT, and AlphaMissense‑Optimized, while pathogenic predictions arise from REVEL, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Four tools give uncertain results: FoldX, Foldetta, premPS, and AlphaMissense‑Default. High‑accuracy assessments provide a mixed picture: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) leans pathogenic, and Foldetta remains uncertain. Overall, the majority of standard predictors favor a pathogenic effect, and the high‑accuracy consensus also tilts toward pathogenicity, though not decisively. Therefore, the variant is most likely pathogenic, and this assessment does not contradict ClinVar status because the variant is not yet reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.019401 | Structured | 0.262601 | Uncertain | 0.885 | 0.222 | 0.000 | -11.507 | Likely Pathogenic | 0.448 | Ambiguous | Likely Benign | 0.70 | Ambiguous | 0.0 | 0.31 | Likely Benign | 0.51 | Ambiguous | 0.67 | Ambiguous | 0.536 | Likely Pathogenic | -0.84 | Neutral | 0.999 | Probably Damaging | 0.995 | Probably Damaging | -1.45 | Pathogenic | 0.21 | Tolerated | 0.2537 | 0.0488 | 0 | 1 | 0.4 | 0.94 | ||||||||||||||||||||||||||||||
| c.1520A>C | K507T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K507T missense variant has no ClinVar record and is not reported in gnomAD. Prediction tools that agree on a benign effect include SIFT, Rosetta, and AlphaMissense‑Optimized. Those that predict a pathogenic impact are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, SGM‑Consensus (a majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifying it as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) yielding an uncertain result. Overall, the majority of available predictions lean toward pathogenicity, and this conclusion does not contradict the absence of a ClinVar classification. **The variant is most likely pathogenic based on the current predictive evidence.** Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.019401 | Structured | 0.262601 | Uncertain | 0.885 | 0.222 | 0.000 | -10.244 | Likely Pathogenic | 0.468 | Ambiguous | Likely Benign | 0.82 | Ambiguous | 0.1 | 0.31 | Likely Benign | 0.57 | Ambiguous | 0.78 | Ambiguous | 0.724 | Likely Pathogenic | -2.86 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.55 | Pathogenic | 0.12 | Tolerated | 0.1501 | 0.2451 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1520A>T | K507M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K507M missense variant is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include FoldX, Foldetta, premPS, PROVEAN, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are SGM‑Consensus, REVEL, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Rosetta remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, more tools (8) predict pathogenicity than benign (5), and the high‑accuracy consensus leans toward pathogenicity. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.019401 | Structured | 0.262601 | Uncertain | 0.885 | 0.222 | 0.000 | -9.548 | Likely Pathogenic | 0.576 | Likely Pathogenic | Likely Benign | 0.12 | Likely Benign | 0.1 | -0.74 | Ambiguous | -0.31 | Likely Benign | -0.22 | Likely Benign | 0.850 | Likely Pathogenic | -2.39 | Neutral | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.63 | Pathogenic | 0.03 | Affected | 0.0783 | 0.2251 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1523A>C | D508A 2D 3DClick to see structure in 3D Viewer AISynGAP1 D508A is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include REVEL, FoldX, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict pathogenicity are SGM Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. The high‑accuracy methods give a split result: AlphaMissense‑Optimized predicts benign, SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) is uncertain. No prediction or stability result is missing; all available outputs are considered. Overall, the predictions are evenly divided between benign and pathogenic, with no clear consensus. Therefore, the variant is inconclusive; it is not contradictory to the ClinVar status, which has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.019401 | Structured | 0.255890 | Uncertain | 0.890 | 0.228 | 0.000 | -11.434 | Likely Pathogenic | 0.704 | Likely Pathogenic | Likely Benign | -0.18 | Likely Benign | 0.2 | 1.84 | Ambiguous | 0.83 | Ambiguous | 0.09 | Likely Benign | 0.339 | Likely Benign | -7.37 | Deleterious | 0.988 | Probably Damaging | 0.762 | Possibly Damaging | 3.37 | Benign | 0.06 | Tolerated | 0.3758 | 0.4241 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||
| c.1526C>A | A509D 2D AIThe SynGAP1 missense variant A509D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a “Likely Pathogenic” verdict; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports a pathogenic effect. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.250110 | Uncertain | 0.923 | 0.256 | 0.000 | -17.026 | Likely Pathogenic | 0.973 | Likely Pathogenic | Likely Pathogenic | 4.02 | Destabilizing | 1.6 | 3.09 | Destabilizing | 3.56 | Destabilizing | 1.36 | Destabilizing | 0.915 | Likely Pathogenic | -4.94 | Deleterious | 0.963 | Probably Damaging | 0.844 | Possibly Damaging | -1.40 | Pathogenic | 0.00 | Affected | 0.1610 | 0.1360 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1526C>T | A509V 2D 3DClick to see structure in 3D Viewer AISynGAP1 A509V is not reported in ClinVar and is absent from gnomAD. High‑accuracy predictors give mixed results: AlphaMissense‑Optimized classifies the variant as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—labels it pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta outputs, is inconclusive because FoldX is uncertain and Rosetta is benign. Among the remaining tools, benign predictions come from REVEL, Rosetta, premPS, polyPhen2_HumDiv, polyPhen2_HumVar, and SIFT, whereas pathogenic predictions come from SGM‑Consensus, PROVEAN, ESM1b, and FATHMM. AlphaMissense‑Default and FoldX remain uncertain. Overall, the majority of evidence points toward a benign effect, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.250110 | Uncertain | 0.923 | 0.256 | 0.000 | -11.987 | Likely Pathogenic | 0.382 | Ambiguous | Likely Benign | 0.52 | Ambiguous | 0.5 | 0.35 | Likely Benign | 0.44 | Likely Benign | -0.25 | Likely Benign | 0.474 | Likely Benign | -3.11 | Deleterious | 0.064 | Benign | 0.048 | Benign | -1.15 | Pathogenic | 0.18 | Tolerated | 0.1333 | 0.5706 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1529T>C | I510T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I510T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only AlphaMissense‑Optimized. All other evaluated algorithms—SGM‑Consensus, REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and Foldetta—classify the variant as pathogenic, while Rosetta remains uncertain. High‑accuracy assessments further support a pathogenic interpretation: AlphaMissense‑Optimized predicts benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts likely pathogenic, and Foldetta predicts pathogenic. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.250630 | Uncertain | 0.945 | 0.273 | 0.000 | -9.993 | Likely Pathogenic | 0.701 | Likely Pathogenic | Likely Benign | 3.08 | Destabilizing | 0.2 | 1.99 | Ambiguous | 2.54 | Destabilizing | 1.95 | Destabilizing | 0.914 | Likely Pathogenic | -3.63 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.43 | Pathogenic | 0.00 | Affected | 0.0960 | 0.0440 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.152T>C | I51T 2D AIThe SynGAP1 missense variant I51T is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default, while AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show that the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Benign, AlphaMissense‑Optimized remains Uncertain, and no Foldetta stability result is available. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.291804 | Structured | 0.454181 | Uncertain | 0.606 | 0.710 | 0.000 | -5.861 | Likely Benign | 0.881 | Likely Pathogenic | Ambiguous | 0.135 | Likely Benign | -1.07 | Neutral | 0.084 | Benign | 0.050 | Benign | 4.16 | Benign | 0.00 | Affected | 0.1180 | 0.1265 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||||||||||||
| c.1532G>A | G511E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G511E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools (FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. No prediction or stability result is missing or inconclusive. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.048328 | Structured | 0.244404 | Uncertain | 0.924 | 0.287 | 0.000 | -12.263 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 2.17 | Destabilizing | 0.2 | 2.62 | Destabilizing | 2.40 | Destabilizing | 1.15 | Destabilizing | 0.479 | Likely Benign | -7.69 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.24 | Benign | 0.02 | Affected | 0.1882 | 0.4240 | 0 | -2 | -3.1 | 72.06 | |||||||||||||||||||||||||||||
| c.1534G>A | E512K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E512K missense variant is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, FATHMM, premPS, and Foldetta, while those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized; Rosetta is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta as benign. Overall, seven tools predict pathogenicity versus six predicting benignity, with one uncertain result. Thus, the variant is most likely pathogenic based on the current computational evidence, and this assessment does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.247079 | Uncertain | 0.923 | 0.273 | 0.000 | -13.927 | Likely Pathogenic | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.17 | Likely Benign | 0.1 | 0.63 | Ambiguous | 0.40 | Likely Benign | -0.03 | Likely Benign | 0.344 | Likely Benign | -3.85 | Deleterious | 0.962 | Probably Damaging | 0.658 | Possibly Damaging | 3.32 | Benign | 0.06 | Tolerated | 0.2844 | 0.4776 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1535A>C | E512A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E512A missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, and FATHMM. Those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Uncertain results come from Foldetta, AlphaMissense‑Optimized, and Rosetta. High‑accuracy assessments show SGM‑Consensus as likely pathogenic, AlphaMissense‑Optimized as uncertain, and Foldetta as uncertain. Overall, the majority of evidence points toward a pathogenic impact, and this conclusion does not contradict any ClinVar annotation because none exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.247079 | Uncertain | 0.923 | 0.273 | 0.000 | -10.979 | Likely Pathogenic | 0.861 | Likely Pathogenic | Ambiguous | 0.45 | Likely Benign | 0.1 | 0.97 | Ambiguous | 0.71 | Ambiguous | 0.04 | Likely Benign | 0.309 | Likely Benign | -5.71 | Deleterious | 0.987 | Probably Damaging | 0.937 | Probably Damaging | 3.31 | Benign | 0.12 | Tolerated | 0.4293 | 0.5162 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1535A>G | E512G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E512G is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, premPS, and FATHMM, whereas a majority of tools (SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default) predict a pathogenic impact. The remaining tools—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—return uncertain or inconclusive results and are treated as unavailable evidence. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for E512G, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.247079 | Uncertain | 0.923 | 0.273 | 0.000 | -11.996 | Likely Pathogenic | 0.817 | Likely Pathogenic | Ambiguous | 0.86 | Ambiguous | 0.1 | 1.84 | Ambiguous | 1.35 | Ambiguous | 0.17 | Likely Benign | 0.381 | Likely Benign | -6.46 | Deleterious | 0.997 | Probably Damaging | 0.915 | Probably Damaging | 3.30 | Benign | 0.02 | Affected | 0.3262 | 0.4185 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1540A>T | I514F 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I514F is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Prediction tools that classify the variant as benign include only FATHMM. All other evaluated algorithms—REVEL, FoldX, Rosetta, Foldetta, premPS (uncertain), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—report it as pathogenic or likely pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, classifies the variant as pathogenic. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, which is consistent with its ClinVar uncertain status rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.049374 | Structured | 0.221408 | Uncertain | 0.948 | 0.266 | 0.000 | Uncertain | 1 | -13.383 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 2.35 | Destabilizing | 0.3 | 3.74 | Destabilizing | 3.05 | Destabilizing | 0.93 | Ambiguous | 0.601 | Likely Pathogenic | -3.98 | Deleterious | 0.997 | Probably Damaging | 0.993 | Probably Damaging | 2.89 | Benign | 0.00 | Affected | 3.37 | 35 | 0.0574 | 0.1629 | 0 | 1 | -1.7 | 34.02 | |||||||||||||||||||||||||
| c.1541T>C | I514T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I514T has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect are limited to FATHMM, whereas the majority of algorithms—SGM‑Consensus, REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. Rosetta reports an uncertain outcome and is not included in the consensus groups. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenic. Taken together, the evidence overwhelmingly points to a pathogenic effect, and this conclusion is not contradicted by the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.049374 | Structured | 0.221408 | Uncertain | 0.948 | 0.266 | 0.000 | -8.820 | Likely Pathogenic | 0.963 | Likely Pathogenic | Likely Pathogenic | 2.92 | Destabilizing | 0.1 | 1.88 | Ambiguous | 2.40 | Destabilizing | 1.94 | Destabilizing | 0.617 | Likely Pathogenic | -4.77 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.82 | Benign | 0.00 | Affected | 0.0962 | 0.0480 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1543C>A | R515S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 R515S missense variant is not reported in ClinVar (status: None) and has no entry in gnomAD. Prediction tools that agree on benign impact include only SIFT, while the remaining tools—REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus—consistently predict pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (derived from the four pathogenic‑predicted tools) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Other stability‑based predictions (FoldX, Rosetta, premPS) are also uncertain. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.055536 | Structured | 0.191256 | Uncertain | 0.924 | 0.275 | 0.000 | -10.615 | Likely Pathogenic | 0.928 | Likely Pathogenic | Ambiguous | 1.54 | Ambiguous | 0.3 | 1.11 | Ambiguous | 1.33 | Ambiguous | 0.92 | Ambiguous | 0.586 | Likely Pathogenic | -3.03 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.28 | Pathogenic | 0.17 | Tolerated | 0.2707 | 0.1849 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||
| c.1544G>C | R515P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant R515P has no ClinVar entry and is absent from gnomAD. In silico prediction tools uniformly indicate a deleterious effect: benign‑predicting algorithms are not reported, while pathogenic‑predicting tools—including REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—all classify the substitution as pathogenic. High‑accuracy assessments corroborate this trend: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports a likely pathogenic outcome; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts a pathogenic effect. Consequently, the variant is most likely pathogenic based on the collective computational evidence, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.055536 | Structured | 0.191256 | Uncertain | 0.924 | 0.275 | 0.000 | -14.291 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 3.69 | Destabilizing | 0.1 | 8.87 | Destabilizing | 6.28 | Destabilizing | 1.02 | Destabilizing | 0.823 | Likely Pathogenic | -4.24 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.36 | Pathogenic | 0.03 | Affected | 0.2223 | 0.2725 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1547C>A | A516D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A516D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only SIFT, which scores the substitution as tolerated. In contrast, the majority of algorithms predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Uncertain or inconclusive results come from FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus is likely pathogenic, while Foldetta remains uncertain. Overall, the preponderance of evidence indicates that A516D is most likely pathogenic, and this conclusion does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.167423 | Uncertain | 0.938 | 0.284 | 0.000 | -14.621 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.65 | Ambiguous | 0.2 | 1.04 | Ambiguous | 0.85 | Ambiguous | 0.62 | Ambiguous | 0.725 | Likely Pathogenic | -5.17 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | -1.23 | Pathogenic | 0.15 | Tolerated | 0.1890 | 0.1679 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1547C>T | A516V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A516V missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess the variant’s effect fall into two groups: benign predictions come from FoldX, Rosetta, and SIFT, while pathogenic predictions are made by SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; premPS is uncertain. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely pathogenic, whereas Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign. Overall, the majority of tools, including the high‑accuracy predictors, lean toward a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.067594 | Structured | 0.167423 | Uncertain | 0.938 | 0.284 | 0.000 | -11.545 | Likely Pathogenic | 0.962 | Likely Pathogenic | Likely Pathogenic | 0.42 | Likely Benign | 0.1 | 0.16 | Likely Benign | 0.29 | Likely Benign | 0.62 | Ambiguous | 0.639 | Likely Pathogenic | -3.61 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | -1.32 | Pathogenic | 0.09 | Tolerated | 0.1336 | 0.5263 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1552T>C | Y518H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y518H is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, SIFT, and FATHMM, whereas a majority of tools predict a pathogenic impact: FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), AlphaMissense‑Default, and ESM1b. Predictions that are inconclusive are Foldetta, AlphaMissense‑Optimized, and Rosetta. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as “Likely Pathogenic,” and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for Y518H, which does not contradict the current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.139895 | Structured | 0.126970 | Uncertain | 0.897 | 0.321 | 0.000 | Uncertain | 1 | -9.797 | Likely Pathogenic | 0.943 | Likely Pathogenic | Ambiguous | 2.39 | Destabilizing | 0.4 | 0.82 | Ambiguous | 1.61 | Ambiguous | 1.31 | Destabilizing | 0.496 | Likely Benign | -4.74 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.40 | Benign | 0.08 | Tolerated | 0.1927 | 0.0212 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||
| c.1553A>G | Y518C 2D AIThe SynGAP1 missense variant Y518C is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into three groups: benign predictions from REVEL, SIFT, and FATHMM; pathogenic predictions from SGM‑Consensus, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default; and uncertain results from Rosetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus indicates likely pathogenic, and Foldetta predicts pathogenic folding instability. Overall, the majority of evidence points to a pathogenic impact, and this conclusion does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.139895 | Structured | 0.126970 | Uncertain | 0.897 | 0.321 | 0.000 | -9.813 | Likely Pathogenic | 0.794 | Likely Pathogenic | Ambiguous | 2.99 | Destabilizing | 0.9 | 1.70 | Ambiguous | 2.35 | Destabilizing | 0.69 | Ambiguous | 0.415 | Likely Benign | -8.35 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.45 | Benign | 0.16 | Tolerated | 0.2905 | 0.1128 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||
| c.1556A>C | E519A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E519A missense variant is listed in ClinVar as Pathogenic (ClinVar ID 1029087.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, SIFT, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, AlphaMissense‑Default, ESM1b, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). Uncertain predictions from Rosetta and AlphaMissense‑Optimized are treated as unavailable. High‑accuracy results are: AlphaMissense‑Optimized – unavailable; SGM‑Consensus – Pathogenic; Foldetta – Benign. Overall, the predictions are balanced, but the high‑accuracy Foldetta result leans toward benign while the consensus leans toward pathogenic, leaving the assessment inconclusive. Based on the available predictions, the variant is most likely benign, contradicting the ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.078022 | Structured | 0.104514 | Uncertain | 0.899 | 0.328 | 0.000 | Likely Pathogenic | 1 | -8.557 | Likely Pathogenic | 0.904 | Likely Pathogenic | Ambiguous | -0.05 | Likely Benign | 0.0 | 0.55 | Ambiguous | 0.25 | Likely Benign | 0.00 | Likely Benign | 0.384 | Likely Benign | -5.23 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 3.33 | Benign | 0.10 | Tolerated | 3.37 | 35 | 0.3544 | 0.3545 | 0 | -1 | 5.3 | -58.04 | 162.4 | 83.5 | -0.1 | 0.1 | -0.2 | 0.0 | X | Potentially Benign | Glu519 is located at the beginning of an α-α loop between the two α-helices (res. Gly502-Tyr518 and Ala533-Val560). In the WT simulations, the carboxylate side chain of Glu519 does not make any specific interactions. Accordingly, the Ala residue swap does not show any negative structural effects in the variant simulations. However, it should be noted that Glu519 faces the missing part of the N-terminal in the model, and thus its potential role in maintaining the tertiary structure might be de-emphasized in the current model. | ||||||||||||||||
| c.1556A>G | E519G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E519G missense variant is not reported in ClinVar (ClinVar status: None) and has no entry in gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, FATHMM, and premPS. Tools that predict a pathogenic effect comprise SGM‑Consensus (Likely Pathogenic), PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. Uncertain predictions come from Foldetta, AlphaMissense‑Optimized, and Rosetta. High‑accuracy assessments: AlphaMissense‑Optimized is uncertain; SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic; Foldetta is uncertain. Overall, the majority of available predictions (7 pathogenic vs. 4 benign) lean toward a pathogenic impact. This conclusion is consistent with the lack of ClinVar annotation, so there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.078022 | Structured | 0.104514 | Uncertain | 0.899 | 0.328 | 0.000 | -10.835 | Likely Pathogenic | 0.884 | Likely Pathogenic | Ambiguous | 0.22 | Likely Benign | 0.1 | 0.92 | Ambiguous | 0.57 | Ambiguous | 0.22 | Likely Benign | 0.458 | Likely Benign | -6.12 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 3.21 | Benign | 0.01 | Affected | 0.2729 | 0.3471 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1559C>G | S520C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S520C is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign predictions come from FoldX, Foldetta, and ESM1b, while pathogenic predictions arise from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, FATHMM, and AlphaMissense‑Default; Rosetta, premPS, and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of evidence points toward a pathogenic effect. Thus, the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status (which is currently unreported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.094817 | Structured | 0.084894 | Uncertain | 0.887 | 0.337 | 0.000 | -6.947 | Likely Benign | 0.917 | Likely Pathogenic | Ambiguous | 0.05 | Likely Benign | 0.2 | 0.76 | Ambiguous | 0.41 | Likely Benign | 0.53 | Ambiguous | 0.720 | Likely Pathogenic | -4.57 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | -1.36 | Pathogenic | 0.03 | Affected | 0.1029 | 0.4948 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||
| c.1561G>A | E521K 2D 3DClick to see structure in 3D Viewer AISynGAP1 E521K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments give mixed results: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is benign. Overall, the majority of conventional tools lean toward a benign interpretation, while the high‑accuracy methods are split. Thus, the variant is most likely benign, and this assessment does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.086953 | Structured | 0.062387 | Uncertain | 0.865 | 0.349 | 0.000 | -9.596 | Likely Pathogenic | 0.911 | Likely Pathogenic | Ambiguous | -0.48 | Likely Benign | 0.2 | 0.14 | Likely Benign | -0.17 | Likely Benign | -0.10 | Likely Benign | 0.379 | Likely Benign | -3.05 | Deleterious | 0.994 | Probably Damaging | 0.994 | Probably Damaging | 3.57 | Benign | 0.45 | Tolerated | 0.2895 | 0.6513 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1562A>C | E521A 2D 3DClick to see structure in 3D Viewer AISynGAP1 E521A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign calls (REVEL, FoldX, Rosetta, premPS, SIFT, FATHMM) and pathogenic calls (SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default). AlphaMissense‑Optimized is uncertain. High‑accuracy assessments give conflicting results: AlphaMissense‑Optimized is uncertain; the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts likely pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts benign. No prediction or stability result is missing. Overall, the evidence is evenly split, with six benign and six pathogenic calls, and the two high‑accuracy tools disagree. Therefore, the variant’s impact remains uncertain; it is not contradicted by ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.086953 | Structured | 0.062387 | Uncertain | 0.865 | 0.349 | 0.000 | -8.997 | Likely Pathogenic | 0.892 | Likely Pathogenic | Ambiguous | 0.18 | Likely Benign | 0.1 | 0.40 | Likely Benign | 0.29 | Likely Benign | 0.11 | Likely Benign | 0.395 | Likely Benign | -4.12 | Deleterious | 0.998 | Probably Damaging | 0.999 | Probably Damaging | 3.28 | Benign | 0.13 | Tolerated | 0.4041 | 0.5918 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1562A>G | E521G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E521G is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, premPS, SIFT, ESM1b, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The high‑accuracy methods give the following results: AlphaMissense‑Optimized predicts benign; Foldetta (combining FoldX‑MD and Rosetta outputs) predicts benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive because it is a 2‑vs‑2 split and is treated as unavailable. Overall, the majority of evidence points to a benign impact. The variant is most likely benign based on current predictions, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.086953 | Structured | 0.062387 | Uncertain | 0.865 | 0.349 | 0.000 | -6.636 | Likely Benign | 0.767 | Likely Pathogenic | Likely Benign | 0.18 | Likely Benign | 0.1 | 0.58 | Ambiguous | 0.38 | Likely Benign | 0.14 | Likely Benign | 0.289 | Likely Benign | -3.64 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.31 | Benign | 0.07 | Tolerated | 0.2932 | 0.5455 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||
| c.1564G>A | E522K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E522K has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a split: benign calls come from Rosetta, Foldetta, premPS, and SIFT, while pathogenic calls are made by REVEL, PROVEAN, both polyPhen‑2 versions, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments further indicate AlphaMissense‑Optimized predicts Pathogenic, SGM‑Consensus also predicts Likely Pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts Benign. No prediction is available from FoldX (Uncertain). Overall, the majority of high‑confidence tools lean toward pathogenicity, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.046216 | Uncertain | 0.823 | 0.376 | 0.000 | -12.637 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | -0.69 | Ambiguous | 0.1 | 0.14 | Likely Benign | -0.28 | Likely Benign | -0.13 | Likely Benign | 0.631 | Likely Pathogenic | -3.81 | Deleterious | 0.994 | Probably Damaging | 0.994 | Probably Damaging | -1.09 | Pathogenic | 0.23 | Tolerated | 0.2019 | 0.4291 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1565A>C | E522A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E522A is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, and premPS, whereas the majority of tools predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus. High‑accuracy assessments further support this: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicts pathogenicity; and Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the preponderance of evidence (10 pathogenic vs. 4 benign predictions) indicates that E522A is most likely pathogenic, and this conclusion does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.046216 | Uncertain | 0.823 | 0.376 | 0.000 | -10.509 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.18 | Likely Benign | 0.1 | 0.01 | Likely Benign | 0.10 | Likely Benign | 0.40 | Likely Benign | 0.739 | Likely Pathogenic | -5.67 | Deleterious | 0.998 | Probably Damaging | 0.999 | Probably Damaging | -1.34 | Pathogenic | 0.02 | Affected | 0.3137 | 0.4128 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1565A>G | E522G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E522G missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Consensus from multiple in‑silico predictors indicates a pathogenic effect: SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as deleterious. No tool reports a benign outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts a damaging effect, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is labeled Likely Pathogenic, while Foldetta’s stability analysis is inconclusive. FoldX, Rosetta, and premPS are uncertain and are treated as unavailable evidence. Overall, the computational evidence overwhelmingly points to a pathogenic impact, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.092881 | Structured | 0.046216 | Uncertain | 0.823 | 0.376 | 0.000 | -10.053 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 0.72 | Ambiguous | 0.0 | 1.62 | Ambiguous | 1.17 | Ambiguous | 0.58 | Ambiguous | 0.823 | Likely Pathogenic | -6.59 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.34 | Pathogenic | 0.01 | Affected | 0.2672 | 0.3654 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1568A>C | N523T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N523T is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. Tools that predict pathogenicity are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The premPS score is uncertain and does not contribute to the consensus. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or folding result is missing or inconclusive. Overall, the majority of tools (10/14) predict pathogenicity, whereas four predict benign. Therefore, the variant is most likely pathogenic based on current predictions, and this assessment does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.069024 | Structured | 0.033426 | Uncertain | 0.883 | 0.383 | 0.125 | -10.056 | Likely Pathogenic | 0.704 | Likely Pathogenic | Likely Benign | 0.47 | Likely Benign | 0.2 | -0.22 | Likely Benign | 0.13 | Likely Benign | 0.65 | Ambiguous | 0.646 | Likely Pathogenic | -5.33 | Deleterious | 0.898 | Possibly Damaging | 0.592 | Possibly Damaging | -1.40 | Pathogenic | 0.02 | Affected | 0.0920 | 0.3780 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||
| c.1571G>A | C524Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C524Y is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that assess the variant’s effect largely agree on a deleterious outcome: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify it as pathogenic, while premPS is the sole tool predicting a benign effect. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenicity. With the overwhelming majority of evidence pointing to a damaging effect and no ClinVar annotation to contradict this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.134866 | Structured | 0.024729 | Uncertain | 0.916 | 0.385 | 0.125 | -11.032 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 3.20 | Destabilizing | 1.4 | 6.24 | Destabilizing | 4.72 | Destabilizing | 0.18 | Likely Benign | 0.863 | Likely Pathogenic | -10.94 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | -1.36 | Pathogenic | 0.00 | Affected | 0.1496 | 0.3911 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1574A>C | E525A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E525A is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include Foldetta, premPS, and FATHMM, whereas the majority of other in silico predictors (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a pathogenic effect. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled Likely Pathogenic, whereas Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts a benign impact. Overall, the preponderance of evidence from both general and high‑accuracy tools points to a pathogenic classification for E525A, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.206376 | Structured | 0.023618 | Uncertain | 0.937 | 0.382 | 0.125 | -12.627 | Likely Pathogenic | 0.977 | Likely Pathogenic | Likely Pathogenic | 1.23 | Ambiguous | 0.6 | -0.62 | Ambiguous | 0.31 | Likely Benign | 0.09 | Likely Benign | 0.680 | Likely Pathogenic | -5.97 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | 2.68 | Benign | 0.00 | Affected | 0.3554 | 0.3960 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1574A>G | E525G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E525G is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include only FATHMM. All other evaluated algorithms—REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—predict it to be pathogenic. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized returns a pathogenic score, the SGM‑Consensus also indicates pathogenicity, while Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, yields an uncertain result. Taken together, the overwhelming majority of evidence points to a pathogenic impact for E525G. This conclusion is consistent with the absence of ClinVar annotation and gnomAD data, and there is no contradiction with ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.206376 | Structured | 0.023618 | Uncertain | 0.937 | 0.382 | 0.125 | -13.181 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 1.71 | Ambiguous | 0.6 | 0.90 | Ambiguous | 1.31 | Ambiguous | 0.78 | Ambiguous | 0.691 | Likely Pathogenic | -6.96 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 2.68 | Benign | 0.00 | Affected | 0.2947 | 0.3886 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1577T>C | V526A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V526A is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity all agree that the variant is deleterious: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify it as pathogenic or likely pathogenic. No tool predicts a benign effect. High‑accuracy assessments further support this: AlphaMissense‑Optimized is uncertain, SGM‑Consensus is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a pathogenic impact. Based on the overwhelming consensus of pathogenic predictions and the lack of benign evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.139895 | Structured | 0.023118 | Uncertain | 0.943 | 0.403 | 0.000 | -12.055 | Likely Pathogenic | 0.852 | Likely Pathogenic | Ambiguous | 2.30 | Destabilizing | 0.1 | 2.49 | Destabilizing | 2.40 | Destabilizing | 2.05 | Destabilizing | 0.908 | Likely Pathogenic | -3.93 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.44 | Pathogenic | 0.00 | Affected | 0.2224 | 0.1922 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1580A>C | D527A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D527A missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only premPS, while the remaining evaluated methods (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict pathogenicity. FoldX, Rosetta, and Foldetta are inconclusive and are treated as unavailable. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta remains uncertain. Overall, the variant is most likely pathogenic based on the consensus of predictive tools, and this assessment does not contradict any ClinVar status, as none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.139895 | Structured | 0.021908 | Uncertain | 0.913 | 0.408 | 0.000 | -15.473 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | 0.81 | Ambiguous | 0.9 | 1.75 | Ambiguous | 1.28 | Ambiguous | -0.24 | Likely Benign | 0.929 | Likely Pathogenic | -7.79 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -2.39 | Pathogenic | 0.00 | Affected | 0.2850 | 0.3799 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||
| c.1582C>A | P528T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P528T is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a pathogenic effect include SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. No tools predict a benign outcome. Predictions that are uncertain or inconclusive are Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as Uncertain, SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta as Uncertain. Based on the preponderance of pathogenic predictions and the lack of benign evidence, the variant is most likely pathogenic. This conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.225814 | Structured | 0.020396 | Uncertain | 0.909 | 0.403 | 0.000 | -13.782 | Likely Pathogenic | 0.798 | Likely Pathogenic | Ambiguous | 2.05 | Destabilizing | 0.3 | 1.01 | Ambiguous | 1.53 | Ambiguous | 0.66 | Ambiguous | 0.673 | Likely Pathogenic | -7.69 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | 2.49 | Pathogenic | 0.00 | Affected | 0.1367 | 0.4360 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||
| c.1583C>A | P528H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P528H is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely indicate a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all classify the change as pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also reports it as likely pathogenic. No tool predicts a benign outcome; the remaining predictions (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized) are uncertain or inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta as uncertain. Taken together, the preponderance of evidence supports a pathogenic interpretation, and this is consistent with the absence of a ClinVar entry; there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.225814 | Structured | 0.020396 | Uncertain | 0.909 | 0.403 | 0.000 | -15.365 | Likely Pathogenic | 0.940 | Likely Pathogenic | Ambiguous | 1.93 | Ambiguous | 0.1 | 0.74 | Ambiguous | 1.34 | Ambiguous | 0.61 | Ambiguous | 0.565 | Likely Pathogenic | -8.61 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.47 | Pathogenic | 0.00 | Affected | 0.1434 | 0.3523 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.1583C>G | P528R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 P528R missense variant is not reported in ClinVar and is absent from gnomAD. No in silico predictor classifies it as benign; the following tools predict pathogenicity: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Predictions that are uncertain or inconclusive (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized) are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as likely pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.225814 | Structured | 0.020396 | Uncertain | 0.909 | 0.403 | 0.000 | -16.367 | Likely Pathogenic | 0.952 | Likely Pathogenic | Ambiguous | 0.76 | Ambiguous | 0.4 | 0.67 | Ambiguous | 0.72 | Ambiguous | 0.73 | Ambiguous | 0.615 | Likely Pathogenic | -8.58 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.49 | Pathogenic | 0.01 | Affected | 0.1261 | 0.2543 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1585A>T | I529F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I529F is not reported in ClinVar and is present in gnomAD (variant ID 6‑33438828‑A‑T). Consensus from most in‑silico predictors classifies the change as benign: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic effect. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Benign. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus is Likely Benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also reports a benign effect. Overall, the preponderance of evidence indicates that I529F is most likely benign, and this conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.318242 | Structured | 0.019545 | Uncertain | 0.901 | 0.403 | 0.000 | 6-33438828-A-T | 4 | 2.48e-6 | -5.669 | Likely Benign | 0.235 | Likely Benign | Likely Benign | -0.21 | Likely Benign | 0.0 | 0.06 | Likely Benign | -0.08 | Likely Benign | 0.06 | Likely Benign | 0.320 | Likely Benign | -0.97 | Neutral | 0.266 | Benign | 0.054 | Benign | -1.30 | Pathogenic | 0.23 | Tolerated | 3.37 | 35 | 0.0522 | 0.2713 | 0 | 1 | -1.7 | 34.02 | ||||||||||||||||||||||||
| c.1586T>C | I529T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I529T is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus “Likely Benign” call. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also yields a benign prediction; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, predicts benign. Overall, the majority of evidence points to a benign effect, and this is consistent with the ClinVar “Uncertain” classification—there is no contradiction between the predictions and the current ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.318242 | Structured | 0.019545 | Uncertain | 0.901 | 0.403 | 0.000 | Uncertain | 1 | -0.539 | Likely Benign | 0.336 | Likely Benign | Likely Benign | 0.22 | Likely Benign | 0.2 | 0.16 | Likely Benign | 0.19 | Likely Benign | 0.17 | Likely Benign | 0.343 | Likely Benign | 0.24 | Neutral | 0.872 | Possibly Damaging | 0.820 | Possibly Damaging | -1.23 | Pathogenic | 0.55 | Tolerated | 3.37 | 35 | 0.0897 | 0.0989 | 0 | -1 | -5.2 | -12.05 | 207.2 | 29.8 | 0.2 | 0.0 | 0.2 | 0.1 | X | Potentially Benign | Ile529 is located on an α-α loop between the two α-helices (res. Gly502-Tyr518 and Ala533-Val560). In the WT simulations, the sec-butyl side chain of Ile529 faces the membrane interface and shows no specific interactions. In the variant simulations, the hydroxyl group of Thr529 forms a hydrogen bond with the carboxylate side chain of Asp527, but no negative structural changes are observed. However, due to its location near the SynGAP-membrane interface, the effect of the residue swap cannot be fully addressed using the SynGAP solvent-only simulations. | ||||||||||||||||
| c.1588A>G | K530E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K530E is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include premPS and polyPhen‑2 HumVar, whereas a majority of tools predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (Likely Pathogenic). High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain, the SGM‑Consensus remains Likely Pathogenic, and Foldetta is uncertain. Overall, the preponderance of evidence points to a pathogenic effect for K530E. This conclusion is consistent with the lack of a ClinVar entry, so there is no contradiction with existing clinical annotations. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.308712 | Structured | 0.018455 | Uncertain | 0.891 | 0.409 | 0.000 | -14.450 | Likely Pathogenic | 0.951 | Likely Pathogenic | Ambiguous | 0.79 | Ambiguous | 0.2 | 1.27 | Ambiguous | 1.03 | Ambiguous | 0.43 | Likely Benign | 0.581 | Likely Pathogenic | -3.45 | Deleterious | 0.703 | Possibly Damaging | 0.276 | Benign | -1.57 | Pathogenic | 0.00 | Affected | 0.2505 | 0.0810 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1589A>C | K530T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K530T missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include only premPS, whereas the remaining nine tools—SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (a majority vote of the pathogenic‑predominant tools) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points to a deleterious impact. Consequently, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently contains no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.308712 | Structured | 0.018455 | Uncertain | 0.891 | 0.409 | 0.000 | -11.506 | Likely Pathogenic | 0.852 | Likely Pathogenic | Ambiguous | 1.06 | Ambiguous | 0.3 | 1.06 | Ambiguous | 1.06 | Ambiguous | 0.27 | Likely Benign | 0.610 | Likely Pathogenic | -5.17 | Deleterious | 0.921 | Possibly Damaging | 0.950 | Probably Damaging | -1.63 | Pathogenic | 0.00 | Affected | 0.1351 | 0.2780 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1589A>T | K530M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K530M missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only premPS. Tools that agree on a pathogenic effect are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. Predictions that are uncertain or inconclusive are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments: AlphaMissense‑Optimized is uncertain; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic; Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. Based on the predominance of pathogenic predictions and the SGM Consensus result, the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.308712 | Structured | 0.018455 | Uncertain | 0.891 | 0.409 | 0.000 | -12.235 | Likely Pathogenic | 0.953 | Likely Pathogenic | Ambiguous | 0.51 | Ambiguous | 0.0 | 1.26 | Ambiguous | 0.89 | Ambiguous | 0.24 | Likely Benign | 0.671 | Likely Pathogenic | -5.17 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | -1.69 | Pathogenic | 0.00 | Affected | 0.0745 | 0.3123 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.158G>A | G53E 2D AIThe SynGAP1 missense variant G53E is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as “Likely Benign.” High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain” and the SGM‑Consensus as “Likely Benign”; Foldetta results are not available. Overall, the majority of evidence points toward a benign impact, and this conclusion does not contradict any ClinVar annotation (none exists). Thus, the variant is most likely benign based on current predictive data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.268042 | Structured | 0.460894 | Uncertain | 0.386 | 0.666 | 0.000 | -2.061 | Likely Benign | 0.853 | Likely Pathogenic | Ambiguous | 0.198 | Likely Benign | -0.25 | Neutral | 0.994 | Probably Damaging | 0.986 | Probably Damaging | 4.29 | Benign | 0.00 | Affected | 0.1413 | 0.4378 | 0 | -2 | -3.1 | 72.06 | |||||||||||||||||||||||||||||||||||||||
| c.1591T>A | C531S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C531S is not reported in ClinVar and has no entries in gnomAD. Prediction tools that assess pathogenicity largely agree on a deleterious effect: SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default all predict pathogenic. In contrast, only three tools predict a benign outcome: FoldX, Foldetta, and AlphaMissense‑Optimized. High‑accuracy methods provide a mixed picture: AlphaMissense‑Optimized reports a benign effect, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic, and Foldetta also predicts benign stability. Overall, the preponderance of evidence points to a pathogenic impact for C531S. This conclusion is not contradicted by ClinVar, which contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.281712 | Structured | 0.017941 | Uncertain | 0.878 | 0.401 | 0.000 | -8.213 | Likely Pathogenic | 0.575 | Likely Pathogenic | Likely Benign | -0.02 | Likely Benign | 0.0 | 0.77 | Ambiguous | 0.38 | Likely Benign | 1.23 | Destabilizing | 0.519 | Likely Pathogenic | -8.00 | Deleterious | 0.958 | Probably Damaging | 0.533 | Possibly Damaging | -1.18 | Pathogenic | 0.01 | Affected | 0.3716 | 0.1782 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1592G>A | C531Y 2D AIThe SynGAP1 missense variant C531Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only Rosetta, whereas the remaining tools—REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM Consensus—consistently predict a pathogenic impact. Uncertain or inconclusive results come from AlphaMissense‑Optimized, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for C531Y. This prediction does not contradict ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.281712 | Structured | 0.017941 | Uncertain | 0.878 | 0.401 | 0.000 | -11.667 | Likely Pathogenic | 0.914 | Likely Pathogenic | Ambiguous | 3.09 | Destabilizing | 4.6 | 0.15 | Likely Benign | 1.62 | Ambiguous | 0.65 | Ambiguous | 0.551 | Likely Pathogenic | -8.95 | Deleterious | 0.976 | Probably Damaging | 0.480 | Possibly Damaging | -1.24 | Pathogenic | 0.00 | Affected | 0.0843 | 0.3266 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1592G>C | C531S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C531S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from FoldX, Foldetta, and AlphaMissense‑Optimized, whereas pathogenic predictions are made by SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default; Rosetta remains uncertain. High‑accuracy assessments further support a pathogenic bias: AlphaMissense‑Optimized indicates benign, but the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—classifies the variant as pathogenic, and Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, predicts benign. Overall, the preponderance of evidence points to a pathogenic effect for C531S, and this conclusion does not conflict with the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.281712 | Structured | 0.017941 | Uncertain | 0.878 | 0.401 | 0.000 | -8.213 | Likely Pathogenic | 0.575 | Likely Pathogenic | Likely Benign | -0.02 | Likely Benign | 0.0 | 0.77 | Ambiguous | 0.38 | Likely Benign | 1.23 | Destabilizing | 0.508 | Likely Pathogenic | -8.00 | Deleterious | 0.958 | Probably Damaging | 0.533 | Possibly Damaging | -1.18 | Pathogenic | 0.01 | Affected | 0.3716 | 0.1782 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1594A>C | T532P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T532P is listed in ClinVar as Benign (ClinVar ID 1598909.0) and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote). Only FATHMM predicts a pathogenic outcome. High‑accuracy assessments—AlphaMissense‑Optimized, the SGM Consensus, and Foldetta (combining FoldX‑MD and Rosetta outputs)—all indicate a benign impact. No prediction or folding‑stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion is consistent with its ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.275179 | Structured | 0.021478 | Uncertain | 0.889 | 0.385 | 0.000 | Benign | 1 | -2.143 | Likely Benign | 0.061 | Likely Benign | Likely Benign | -0.30 | Likely Benign | 0.2 | 0.06 | Likely Benign | -0.12 | Likely Benign | 0.08 | Likely Benign | 0.201 | Likely Benign | -0.90 | Neutral | 0.005 | Benign | 0.008 | Benign | -1.28 | Pathogenic | 0.18 | Tolerated | 3.37 | 35 | 0.1850 | 0.3811 | 0 | -1 | -0.9 | -3.99 | 174.2 | 35.1 | 0.4 | 0.0 | 0.1 | 0.0 | X | Potentially Benign | Thr532 is located on an α-α loop between the two α-helices (res. Gly502-Tyr518 and Ala533-Val560) facing the membrane. In the WT simulations, the hydroxyl group of Thr532 occasionally forms hydrogen bonds with the backbone atoms of other loop residues without any specific interaction. In the variant simulations, the Pro532 residue swap does not cause structural changes. Although hydrophilic residues seem more favorable in the loop, the pyrrolidine side chain of proline is well suited for unstructured protein regions such as loops. However, due to its location at the SynGAP-membrane interface, the effect of the residue swap cannot be fully addressed using the SynGAP solvent-only simulations. | ||||||||||||||||
| c.1595C>A | T532K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 T532K missense variant is not reported in ClinVar and has no entries in gnomAD. Prediction tools that classify the change as benign include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Those that predict pathogenicity are ESM1b, FATHMM, AlphaMissense‑Default, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM Consensus as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. With ten benign versus four pathogenic predictions and two of the three high‑accuracy tools supporting a benign outcome, the variant is most likely benign. This assessment does not contradict the ClinVar status, which currently has no classification for T532K. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.021478 | Uncertain | 0.889 | 0.385 | 0.000 | -8.460 | Likely Pathogenic | 0.644 | Likely Pathogenic | Likely Benign | -0.44 | Likely Benign | 0.2 | 0.24 | Likely Benign | -0.10 | Likely Benign | 0.34 | Likely Benign | 0.376 | Likely Benign | -1.97 | Neutral | 0.259 | Benign | 0.033 | Benign | -1.21 | Pathogenic | 0.29 | Tolerated | 0.0943 | 0.1992 | 0 | -1 | -3.2 | 27.07 | |||||||||||||||||||||||||||||
| c.1595C>T | T532I 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant T532I is not reported in ClinVar and has no entries in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Foldetta, premPS, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. The high‑accuracy methods give a mixed picture: AlphaMissense‑Optimized classifies the variant as benign, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) reports a benign effect. Overall, the balance of evidence leans toward pathogenicity, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.021478 | Uncertain | 0.889 | 0.385 | 0.000 | -9.031 | Likely Pathogenic | 0.591 | Likely Pathogenic | Likely Benign | -0.11 | Likely Benign | 0.2 | 0.87 | Ambiguous | 0.38 | Likely Benign | 0.22 | Likely Benign | 0.428 | Likely Benign | -3.31 | Deleterious | 0.633 | Possibly Damaging | 0.202 | Benign | -1.31 | Pathogenic | 0.03 | Affected | 0.0713 | 0.4983 | 0 | -1 | 5.2 | 12.05 | |||||||||||||||||||||||||||||
| c.1597G>A | A533T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A533T is catalogued in gnomAD (6-33438840‑G‑A) but has no ClinVar entry. In silico predictors overwhelmingly favor a benign effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all report benign or tolerated outcomes, whereas only FATHMM predicts pathogenicity. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, the SGM Consensus is Likely Benign, and Foldetta (combining FoldX‑MD and Rosetta) predicts a benign stability change. Overall, the evidence points to a benign impact, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.179055 | Structured | 0.026324 | Uncertain | 0.843 | 0.393 | 0.000 | 6-33438840-G-A | 2 | 1.24e-6 | -5.396 | Likely Benign | 0.084 | Likely Benign | Likely Benign | 0.30 | Likely Benign | 0.1 | 0.31 | Likely Benign | 0.31 | Likely Benign | 0.19 | Likely Benign | 0.147 | Likely Benign | -0.48 | Neutral | 0.002 | Benign | 0.001 | Benign | -1.26 | Pathogenic | 0.11 | Tolerated | 3.37 | 35 | 0.1413 | 0.6541 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||
| c.1598C>A | A533E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A533E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are ESM1b and FATHMM, while AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact. This conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.026324 | Uncertain | 0.843 | 0.393 | 0.000 | -10.810 | Likely Pathogenic | 0.553 | Ambiguous | Likely Benign | -0.13 | Likely Benign | 0.0 | -0.26 | Likely Benign | -0.20 | Likely Benign | 0.43 | Likely Benign | 0.314 | Likely Benign | -1.89 | Neutral | 0.259 | Benign | 0.107 | Benign | -1.18 | Pathogenic | 0.18 | Tolerated | 0.1147 | 0.1498 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||
| c.1598C>T | A533V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A533V is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized, Foldetta, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are SIFT and FATHMM. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as likely benign, and Foldetta as benign, all supporting a non‑pathogenic interpretation. No prediction or folding‑stability result is missing or inconclusive. Based on the overall consensus of the available predictions, the variant is most likely benign, and this conclusion does not contradict any ClinVar status because no ClinVar evidence exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.179055 | Structured | 0.026324 | Uncertain | 0.843 | 0.393 | 0.000 | -6.948 | Likely Benign | 0.148 | Likely Benign | Likely Benign | 0.23 | Likely Benign | 0.0 | 0.51 | Ambiguous | 0.37 | Likely Benign | 0.25 | Likely Benign | 0.170 | Likely Benign | -2.41 | Neutral | 0.149 | Benign | 0.024 | Benign | -1.28 | Pathogenic | 0.05 | Affected | 0.1243 | 0.6205 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1601C>G | S534C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S534C is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that indicate a benign effect include REVEL, FoldX, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. High‑accuracy assessments show AlphaMissense‑Optimized as benign; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta is uncertain, so both are treated as unavailable. No other high‑accuracy predictions are available. Overall, the evidence is evenly split between benign and pathogenic predictions, leaving the variant’s clinical significance inconclusive. There is no ClinVar status to contradict this balanced prediction. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.167087 | Structured | 0.032173 | Uncertain | 0.860 | 0.362 | 0.000 | -8.077 | Likely Pathogenic | 0.247 | Likely Benign | Likely Benign | 0.18 | Likely Benign | 0.1 | 0.90 | Ambiguous | 0.54 | Ambiguous | 0.56 | Ambiguous | 0.308 | Likely Benign | -4.05 | Deleterious | 0.998 | Probably Damaging | 0.997 | Probably Damaging | 3.25 | Benign | 0.00 | Affected | 0.0984 | 0.4861 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.1603A>T | S535C 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant S535C is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include Rosetta, Foldetta, premPS, polyPhen‑2 HumVar, AlphaMissense‑Default, and AlphaMissense‑Optimized. Those that predict pathogenicity are REVEL, PROVEAN, polyPhen‑2 HumDiv, SIFT, and FATHMM. Two tools, FoldX and ESM1b, return uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, Foldetta as benign, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic. Overall, the majority of predictions lean toward a benign effect, and this conclusion is not contradicted by ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.236433 | Structured | 0.041365 | Uncertain | 0.918 | 0.343 | 0.000 | -7.526 | In-Between | 0.165 | Likely Benign | Likely Benign | 0.57 | Ambiguous | 0.1 | 0.25 | Likely Benign | 0.41 | Likely Benign | 0.47 | Likely Benign | 0.500 | Likely Pathogenic | -2.96 | Deleterious | 0.933 | Possibly Damaging | 0.419 | Benign | -1.33 | Pathogenic | 0.02 | Affected | 0.1139 | 0.5633 | 0 | -1 | 3.3 | 16.06 | ||||||||||||||||||||||||||||||
| c.1609G>A | A537T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A537T is catalogued in gnomAD (6‑33438852‑G‑A) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, classifies the variant as Likely Benign. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized predicts Benign; the SGM‑Consensus (majority vote) is Likely Benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts Benign. No prediction or folding stability result is missing or inconclusive. Based on the collective evidence, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.116183 | Structured | 0.037313 | Uncertain | 0.928 | 0.362 | 0.000 | 6-33438852-G-A | 2 | 1.24e-6 | -4.704 | Likely Benign | 0.097 | Likely Benign | Likely Benign | 0.35 | Likely Benign | 0.0 | 0.29 | Likely Benign | 0.32 | Likely Benign | 0.37 | Likely Benign | 0.210 | Likely Benign | -1.41 | Neutral | 0.953 | Possibly Damaging | 0.602 | Possibly Damaging | -1.27 | Pathogenic | 0.44 | Tolerated | 3.37 | 35 | 0.1520 | 0.5440 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||
| c.1610C>A | A537E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A537E is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM; AlphaMissense‑Default is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as benign, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction or stability result is missing or inconclusive. Based on the overall consensus of the available tools, the variant is most likely benign, and this conclusion does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.116183 | Structured | 0.037313 | Uncertain | 0.928 | 0.362 | 0.000 | -5.120 | Likely Benign | 0.450 | Ambiguous | Likely Benign | -0.14 | Likely Benign | 0.1 | 0.00 | Likely Benign | -0.07 | Likely Benign | 0.23 | Likely Benign | 0.378 | Likely Benign | -1.78 | Neutral | 0.987 | Probably Damaging | 0.838 | Possibly Damaging | -1.04 | Pathogenic | 1.00 | Tolerated | 0.1347 | 0.1566 | 0 | -1 | -5.3 | 58.04 | ||||||||||||||||||||||||||||||
| c.1610C>T | A537V 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A537V is listed in ClinVar as Benign (ClinVar ID 766762.0) and is present in gnomAD (ID 6‑33438853‑C‑T). Functional prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments further support a benign outcome: AlphaMissense‑Optimized is benign; the SGM Consensus, derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is benign; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, is benign. FoldX alone is uncertain and therefore not considered evidence. Overall, the consensus of available predictions indicates that the variant is most likely benign, in agreement with its ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | GAP | 0.116183 | Structured | 0.037313 | Uncertain | 0.928 | 0.362 | 0.000 | Likely Benign | 1 | 6-33438853-C-T | 7 | 4.34e-6 | -6.888 | Likely Benign | 0.120 | Likely Benign | Likely Benign | 0.54 | Ambiguous | 0.0 | -0.05 | Likely Benign | 0.25 | Likely Benign | 0.41 | Likely Benign | 0.382 | Likely Benign | -1.97 | Neutral | 0.977 | Probably Damaging | 0.469 | Possibly Damaging | -1.26 | Pathogenic | 0.24 | Tolerated | 3.37 | 35 | 0.1328 | 0.4748 | 0 | 0 | 2.4 | 28.05 | 220.3 | -45.1 | 0.0 | 0.0 | -0.7 | 0.1 | X | Potentially Benign | Ala537 is located on the outer surface of an α-helix (res. Ala533-Val560). The methyl group of Ala537 is on the surface and does not form any interactions. In the variant simulations, the iso-propyl side chain of Val537 is also on the surface, similar to Ala537 in the WT, causing no negative structural effects. | |||||||||||||
| c.1612G>A | E538K 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant E538K has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show mixed results: benign predictions come from REVEL, FoldX, Rosetta, Foldetta, premPS, polyPhen‑2 HumVar, SIFT, and FATHMM, whereas pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and AlphaMissense‑Default. The high‑accuracy assessment indicates that AlphaMissense‑Optimized is uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) supports a pathogenic outcome, and Foldetta (combining FoldX‑MD and Rosetta) predicts benign stability. Overall, the majority of tools lean toward a benign effect, but the consensus of high‑accuracy predictors and several individual pathogenic scores suggest uncertainty. The variant is most likely benign based on the bulk of evidence, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.122885 | Structured | 0.033501 | Uncertain | 0.938 | 0.359 | 0.000 | -11.345 | Likely Pathogenic | 0.922 | Likely Pathogenic | Ambiguous | -0.03 | Likely Benign | 0.0 | -0.16 | Likely Benign | -0.10 | Likely Benign | -0.22 | Likely Benign | 0.215 | Likely Benign | -2.97 | Deleterious | 0.848 | Possibly Damaging | 0.294 | Benign | 3.46 | Benign | 0.16 | Tolerated | 0.2257 | 0.3818 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1613A>C | E538A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E538A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, and FATHMM. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (majority vote). High‑accuracy methods give mixed results: AlphaMissense‑Optimized predicts benign, SGM‑Consensus predicts likely pathogenic, and Foldetta predicts benign. No prediction or folding stability result is missing or inconclusive. Overall, the majority of tools (7 pathogenic vs 6 benign) lean toward a pathogenic interpretation, and this does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.122885 | Structured | 0.033501 | Uncertain | 0.938 | 0.359 | 0.000 | -9.888 | Likely Pathogenic | 0.770 | Likely Pathogenic | Likely Benign | 0.33 | Likely Benign | 0.0 | -0.06 | Likely Benign | 0.14 | Likely Benign | 0.16 | Likely Benign | 0.269 | Likely Benign | -4.66 | Deleterious | 0.944 | Possibly Damaging | 0.761 | Possibly Damaging | 3.36 | Benign | 0.05 | Affected | 0.3995 | 0.4187 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1613A>G | E538G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E538G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, FATHMM, and AlphaMissense‑Optimized; pathogenic predictions arise from PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is labeled Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM Consensus as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. No folding‑stability result is definitive. Overall, the balance of evidence, including the majority‑vote SGM Consensus and the higher number of pathogenic predictions, points to a pathogenic effect for E538G. This conclusion does not contradict ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.122885 | Structured | 0.033501 | Uncertain | 0.938 | 0.359 | 0.000 | -9.258 | Likely Pathogenic | 0.701 | Likely Pathogenic | Likely Benign | 0.91 | Ambiguous | 0.0 | 1.07 | Ambiguous | 0.99 | Ambiguous | 0.31 | Likely Benign | 0.285 | Likely Benign | -5.10 | Deleterious | 0.993 | Probably Damaging | 0.700 | Possibly Damaging | 3.33 | Benign | 0.03 | Affected | 0.3173 | 0.3713 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1615C>T | H539Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H539Y has no ClinVar entry and is not reported in gnomAD. Prediction tools that indicate a benign effect include only premPS, whereas the remaining evaluated algorithms (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, and AlphaMissense‑Default) all predict a pathogenic impact. Predictions marked as uncertain are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show that AlphaMissense‑Optimized is inconclusive, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) supports pathogenicity, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also inconclusive. Overall, the preponderance of evidence points to a pathogenic effect for H539Y. This conclusion is not contradicted by ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.031398 | Uncertain | 0.948 | 0.360 | 0.000 | -13.177 | Likely Pathogenic | 0.923 | Likely Pathogenic | Ambiguous | -1.22 | Ambiguous | 0.0 | -1.12 | Ambiguous | -1.17 | Ambiguous | 0.34 | Likely Benign | 0.906 | Likely Pathogenic | -5.60 | Deleterious | 0.998 | Probably Damaging | 0.990 | Probably Damaging | -1.26 | Pathogenic | 0.01 | Affected | 0.0776 | 0.2552 | 0 | 2 | 1.9 | 26.03 | |||||||||||||||||||||||||||||
| c.1616A>C | H539P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H539P missense variant is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that indicate a benign effect include only FoldX, whereas all other evaluated algorithms—REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the change as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenic. Taken together, the overwhelming majority of evidence points to a pathogenic effect, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.031398 | Uncertain | 0.948 | 0.360 | 0.000 | -15.182 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.22 | Likely Benign | 0.3 | 9.08 | Destabilizing | 4.65 | Destabilizing | 1.03 | Destabilizing | 0.913 | Likely Pathogenic | -9.18 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.29 | Pathogenic | 0.01 | Affected | 0.1900 | 0.2794 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||
| c.1619A>C | Q540P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q540P is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: SGM‑Consensus, REVEL, FoldX, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the substitution as pathogenic, while premPS is uncertain. High‑accuracy assessments corroborate this: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also reports a pathogenic effect. No tool predicts a benign outcome. Consequently, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently has no entry for this change. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.085092 | Structured | 0.029522 | Uncertain | 0.958 | 0.371 | 0.000 | -16.096 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 3.95 | Destabilizing | 0.3 | 10.06 | Destabilizing | 7.01 | Destabilizing | 0.73 | Ambiguous | 0.922 | Likely Pathogenic | -5.96 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.32 | Pathogenic | 0.01 | Affected | 0.1882 | 0.3654 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||||
| c.161A>C | N54T 2D AIThe SynGAP1 missense variant N54T is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while those that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus from SGM (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of computational evidence points to a benign impact, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.196879 | Structured | 0.464669 | Uncertain | 0.504 | 0.659 | 0.000 | -5.808 | Likely Benign | 0.386 | Ambiguous | Likely Benign | 0.070 | Likely Benign | -0.47 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.20 | Benign | 0.00 | Affected | 0.1268 | 0.7498 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||||||||||||
| c.1622C>A | A541D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A541D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include SIFT and FoldX, whereas a larger group—SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default—consistently predict pathogenicity. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.050641 | Structured | 0.029947 | Uncertain | 0.955 | 0.365 | 0.000 | -11.510 | Likely Pathogenic | 0.864 | Likely Pathogenic | Ambiguous | 0.36 | Likely Benign | 0.0 | 0.65 | Ambiguous | 0.51 | Ambiguous | 0.65 | Ambiguous | 0.571 | Likely Pathogenic | -3.18 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.15 | Pathogenic | 0.14 | Tolerated | 0.1581 | 0.1902 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1622C>T | A541V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A541V is not reported in ClinVar and is absent from gnomAD. Benign predictions come from REVEL, Foldetta, premPS, SIFT, Rosetta, and AlphaMissense‑Optimized, whereas pathogenic predictions come from SGM‑Consensus, PROVEAN, polyPhen2_HumDiv, polyPhen2_HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy tools give mixed results: AlphaMissense‑Optimized predicts benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts pathogenic, and Foldetta predicts benign. FoldX is uncertain and therefore not considered. Overall, the majority of tools predict pathogenicity, and this assessment is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.050641 | Structured | 0.029947 | Uncertain | 0.955 | 0.365 | 0.000 | -9.777 | Likely Pathogenic | 0.762 | Likely Pathogenic | Likely Benign | 0.63 | Ambiguous | 0.1 | 0.06 | Likely Benign | 0.35 | Likely Benign | 0.43 | Likely Benign | 0.497 | Likely Benign | -3.09 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | -1.33 | Pathogenic | 0.06 | Tolerated | 0.0985 | 0.3648 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1625A>C | N542T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N542T is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are SIFT and Rosetta. Tools that predict a pathogenic effect include SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain or inconclusive results come from FoldX, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact. This conclusion is not contradicted by ClinVar status, which has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.053060 | Structured | 0.026143 | Uncertain | 0.953 | 0.331 | 0.000 | -9.918 | Likely Pathogenic | 0.974 | Likely Pathogenic | Likely Pathogenic | 1.13 | Ambiguous | 0.1 | 0.20 | Likely Benign | 0.67 | Ambiguous | 0.75 | Ambiguous | 0.784 | Likely Pathogenic | -5.37 | Deleterious | 0.997 | Probably Damaging | 0.990 | Probably Damaging | -1.41 | Pathogenic | 0.12 | Tolerated | 0.1120 | 0.5872 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||
| c.1631G>C | R544P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant R544P is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Across the available in‑silico predictors, none indicate a benign effect; all 13 tools (REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic outcome. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) is pathogenic. Consequently, the variant is most likely pathogenic based on current predictions, which contradicts the ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.038858 | Structured | 0.016004 | Uncertain | 0.967 | 0.333 | 0.000 | Uncertain | 2 | -16.905 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 4.70 | Destabilizing | 0.1 | 4.19 | Destabilizing | 4.45 | Destabilizing | 1.14 | Destabilizing | 0.762 | Likely Pathogenic | -4.88 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.48 | Pathogenic | 0.05 | Affected | 3.37 | 35 | 0.2024 | 0.3038 | 0 | -2 | 2.9 | -59.07 | 192.0 | 123.8 | 0.1 | 0.0 | -0.3 | 0.0 | X | X | Potentially Pathogenic | Arg544 is located in the middle of an α-helix (res. Ala533-Val560). In the WT simulations, the guanidinium side chain of Arg544 forms a salt bridge with the carboxylate groups of Glu548 on the same α-helix, and with Glu651 and Glu656 on an opposing α-helix (res. Glu666-Asp644). In the variant simulations, the pyrrolidine side chain of Pro544 cannot form any of the salt bridges that Arg544 does in the WT, potentially weakening the tertiary structure assembly. Additionally, Pro544 lacks the amide group, and thus, unlike Arg544 in the WT, is unable to form a hydrogen bond with the carbonyl of Gln540. This disruption breaks the continuity of the secondary structure element, causing the α-helix to bend slightly in the variant simulations. These negative structural effects could be more pronounced during protein folding and are likely to be undermined in the MD simulations. | |||||||||||||||
| c.1634T>A | M545K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M545K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from SIFT, FoldX, and Rosetta, while pathogenic predictions are made by REVEL, premPS, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and PROVEAN. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic. High‑accuracy assessments further show AlphaMissense‑Optimized as Pathogenic, SGM Consensus as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as Benign. Overall, the majority of evidence points toward a pathogenic effect, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.012875 | Uncertain | 0.955 | 0.311 | 0.000 | -11.723 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 0.02 | Likely Benign | 0.1 | 0.46 | Likely Benign | 0.24 | Likely Benign | 1.07 | Destabilizing | 0.809 | Likely Pathogenic | -4.80 | Deleterious | 0.972 | Probably Damaging | 0.960 | Probably Damaging | -1.17 | Pathogenic | 0.43 | Tolerated | 0.1144 | 0.0488 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1634T>G | M545R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M545R is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Prediction tools that classify the variant as benign include SIFT, FoldX, and Foldetta, whereas the majority of tools predict it to be pathogenic: SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Rosetta’s assessment is uncertain. High‑accuracy methods give a mixed picture: AlphaMissense‑Optimized predicts pathogenicity; the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also indicates likely pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts benign. Overall, the preponderance of evidence from both general and high‑accuracy predictors points to a pathogenic effect. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.025762 | Structured | 0.012875 | Uncertain | 0.955 | 0.311 | 0.000 | -9.223 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | -0.27 | Likely Benign | 0.1 | 0.95 | Ambiguous | 0.34 | Likely Benign | 1.07 | Destabilizing | 0.773 | Likely Pathogenic | -4.76 | Deleterious | 0.987 | Probably Damaging | 0.971 | Probably Damaging | -1.23 | Pathogenic | 0.36 | Tolerated | 0.1345 | 0.0837 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||
| c.1637G>A | C546Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C546Y is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that classify the variant as benign include FoldX, premPS, and SIFT, whereas the remaining tools—SGM‑Consensus, REVEL, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict it to be pathogenic. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized reports a pathogenic effect; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also indicates pathogenicity; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, predicts a destabilizing, pathogenic impact. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.027463 | Structured | 0.009041 | Uncertain | 0.960 | 0.288 | 0.000 | -10.771 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | -0.08 | Likely Benign | 1.8 | 4.75 | Destabilizing | 2.34 | Destabilizing | 0.09 | Likely Benign | 0.794 | Likely Pathogenic | -8.74 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.07 | Pathogenic | 0.29 | Tolerated | 0.1433 | 0.3015 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1639T>A | C547S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C547S is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. No tools predict a benign outcome. Predictions that are uncertain or inconclusive are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.045352 | Structured | 0.007912 | Uncertain | 0.971 | 0.275 | 0.000 | -11.888 | Likely Pathogenic | 0.946 | Likely Pathogenic | Ambiguous | 0.85 | Ambiguous | 0.0 | 1.73 | Ambiguous | 1.29 | Ambiguous | 1.76 | Destabilizing | 0.884 | Likely Pathogenic | -9.64 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.27 | Pathogenic | 0.04 | Affected | 0.4542 | 0.1792 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1640G>A | C547Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C547Y (ClinVar ID 1404191.0) is listed as Pathogenic in ClinVar and is not reported in gnomAD. Prediction tools that agree on a benign effect include only SIFT, whereas the remaining tools—REVEL, FoldX, Rosetta, Foldetta, premPS (uncertain), PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is pathogenic. Taken together, the overwhelming majority of computational evidence indicates a pathogenic effect, which is in agreement with the ClinVar classification. Thus, the variant is most likely pathogenic and does not contradict the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.045352 | Structured | 0.007912 | Uncertain | 0.971 | 0.275 | 0.000 | Pathogenic | 1 | -15.871 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 8.53 | Destabilizing | 1.8 | 6.20 | Destabilizing | 7.37 | Destabilizing | 0.62 | Ambiguous | 0.874 | Likely Pathogenic | -10.57 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.33 | Pathogenic | 0.06 | Tolerated | 3.37 | 35 | 0.1393 | 0.2742 | 0 | -2 | -3.8 | 60.04 | 280.1 | -54.8 | 0.0 | 0.0 | 0.0 | 0.0 | X | X | X | Potentially Pathogenic | Cys547 is located in an α-helix (res. Ala533-Val560). The thiol side chain of Cys547 is situated in a hydrophobic inter-helix space, where it packs hydrophobically with other residues such as Ile626, Leu551, and Phe652. Additionally, the thiol side chain of Cys weakly hydrogen bonds with the carbonyl group of Leu543 in the same α-helix. In the variant simulations, the bulkier phenol ring of Tyr547, with its polar hydroxyl group, is less suited for the hydrophobic space. Consequently, it moves outside and forms a hydrogen bond with the carbonyl group of Phe652 in the neighboring α-helix (res. Glu666-Asp644). This causes the two helices to slightly separate, negatively affecting the secondary structure integrity of the latter helix. These negative structural effects could be more pronounced during protein folding and are likely to be undermined in the MD simulations. | ||||||||||||||
| c.1640G>C | C547S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C547S is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default. No tools predict a benign outcome. Predictions that are uncertain or inconclusive are FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect. This conclusion is consistent with the lack of ClinVar annotation and does not contradict any existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.045352 | Structured | 0.007912 | Uncertain | 0.971 | 0.275 | 0.000 | -11.888 | Likely Pathogenic | 0.946 | Likely Pathogenic | Ambiguous | 0.85 | Ambiguous | 0.0 | 1.73 | Ambiguous | 1.29 | Ambiguous | 1.76 | Destabilizing | 0.825 | Likely Pathogenic | -9.64 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.27 | Pathogenic | 0.04 | Affected | 0.4542 | 0.1792 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1642G>A | E548K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E548K missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, premPS, SIFT, and FATHMM, while those that predict a pathogenic impact are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as pathogenic, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also predicting likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) indicating a benign effect on protein stability. Overall, the balance of evidence leans toward a pathogenic interpretation, with no conflict with ClinVar status because the variant has not yet been reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.054297 | Structured | 0.008632 | Uncertain | 0.965 | 0.288 | 0.000 | -13.734 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | -0.34 | Likely Benign | 0.0 | -0.19 | Likely Benign | -0.27 | Likely Benign | 0.13 | Likely Benign | 0.348 | Likely Benign | -3.85 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.33 | Benign | 0.09 | Tolerated | 0.1990 | 0.4491 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1643A>C | E548A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E548A is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify it as benign include REVEL, FoldX, FATHMM, premPS, and Foldetta, whereas pathogenic predictions come from SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy methods give a pathogenic call from AlphaMissense‑Optimized, a likely pathogenic verdict from the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN), and a benign result from Foldetta. Overall, the majority of evidence points to a pathogenic effect, so the variant is most likely pathogenic, and this assessment does not contradict the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.054297 | Structured | 0.008632 | Uncertain | 0.965 | 0.288 | 0.000 | -13.543 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.18 | Likely Benign | 0.1 | 0.56 | Ambiguous | 0.37 | Likely Benign | 0.48 | Likely Benign | 0.499 | Likely Benign | -5.83 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.27 | Benign | 0.05 | Affected | 0.3035 | 0.4328 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1643A>G | E548G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E548G missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include SIFT and FATHMM, while the majority of other in silico predictors (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a pathogenic impact; SGM‑Consensus also classifies the variant as likely pathogenic. Uncertain results are reported by FoldX, Rosetta, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized predicting pathogenicity, SGM‑Consensus confirming a likely pathogenic status, and Foldetta yielding an inconclusive stability change. Overall, the consensus of the available predictions points to a pathogenic effect for E548G, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.054297 | Structured | 0.008632 | Uncertain | 0.965 | 0.288 | 0.000 | -12.010 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 0.97 | Ambiguous | 0.1 | 1.66 | Ambiguous | 1.32 | Ambiguous | 0.59 | Ambiguous | 0.521 | Likely Pathogenic | -6.73 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.32 | Benign | 0.06 | Tolerated | 0.2638 | 0.3654 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1649C>A | A550D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A550D is not reported in ClinVar and is absent from gnomAD. Functional prediction tools largely converge on a deleterious effect. Benign predictions: none. Pathogenic predictions: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized. Uncertain predictions: Rosetta and Foldetta. High‑accuracy assessments further support a damaging outcome: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains uncertain. Overall, the overwhelming majority of evidence indicates a pathogenic effect. Based on the aggregate predictions, the variant is most likely pathogenic, and this is not contradicted by ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.018106 | Structured | 0.007241 | Uncertain | 0.954 | 0.265 | 0.000 | -18.844 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 2.51 | Destabilizing | 0.1 | 1.46 | Ambiguous | 1.99 | Ambiguous | 1.01 | Destabilizing | 0.879 | Likely Pathogenic | -5.43 | Deleterious | 0.999 | Probably Damaging | 0.971 | Probably Damaging | -1.32 | Pathogenic | 0.01 | Affected | 0.1333 | 0.1783 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1649C>T | A550V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A550V variant has no ClinVar entry and is catalogued in gnomAD (ID 6‑33438892‑C‑T). Prediction tools that agree on a benign effect include Foldetta, premPS, SIFT, and AlphaMissense‑Optimized. Those that predict a pathogenic outcome are REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Uncertain results come from FoldX and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM‑Consensus as pathogenic, and Foldetta as benign. Overall, the majority of tools (seven versus four) favor a pathogenic interpretation, and this does not contradict any ClinVar classification because none is available. Thus, based on the current predictions, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.018106 | Structured | 0.007241 | Uncertain | 0.954 | 0.265 | 0.000 | 6-33438892-C-T | 1 | 6.20e-7 | -10.461 | Likely Pathogenic | 0.441 | Ambiguous | Likely Benign | 0.77 | Ambiguous | 0.2 | -0.05 | Likely Benign | 0.36 | Likely Benign | 0.48 | Likely Benign | 0.540 | Likely Pathogenic | -2.93 | Deleterious | 0.984 | Probably Damaging | 0.494 | Possibly Damaging | -1.05 | Pathogenic | 0.39 | Tolerated | 3.37 | 33 | 0.0874 | 0.4370 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||
| c.1654T>A | C552S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C552S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, and SIFT, whereas a majority of tools predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). The high‑accuracy assessments are mixed: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM‑Consensus also indicates a likely pathogenic outcome, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, reports a benign effect. premPS is inconclusive. Overall, the preponderance of evidence from multiple independent predictors points to a pathogenic effect for C552S. This conclusion is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.013265 | Structured | 0.005714 | Uncertain | 0.955 | 0.256 | 0.000 | -9.309 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | -0.27 | Likely Benign | 0.0 | -0.34 | Likely Benign | -0.31 | Likely Benign | 0.97 | Ambiguous | 0.748 | Likely Pathogenic | -8.21 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.15 | Pathogenic | 0.69 | Tolerated | 0.3469 | 0.1415 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1655G>A | C552Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C552Y is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions from premPS and SIFT, and pathogenic predictions from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain results are reported by FoldX, Rosetta, and Foldetta. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic, while Foldetta’s stability analysis is inconclusive. Overall, the majority of evidence points to a pathogenic impact for C552Y. This conclusion is consistent with the absence of ClinVar annotation and does not contradict any existing database status. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.013265 | Structured | 0.005714 | Uncertain | 0.955 | 0.256 | 0.000 | -13.195 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | -0.96 | Ambiguous | 0.1 | -0.57 | Ambiguous | -0.77 | Ambiguous | 0.41 | Likely Benign | 0.750 | Likely Pathogenic | -9.37 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.23 | Pathogenic | 0.07 | Tolerated | 0.0988 | 0.2563 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1655G>C | C552S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C552S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include FoldX, Rosetta, Foldetta, and SIFT, whereas a majority of tools predict a pathogenic impact: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). The high‑accuracy assessments are mixed: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM‑Consensus also indicates a likely pathogenic outcome, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability predictions, reports a benign effect. premPS is inconclusive. Overall, the preponderance of evidence from multiple independent predictors points to a pathogenic effect for C552S. This conclusion is not contradicted by ClinVar status, as the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.013265 | Structured | 0.005714 | Uncertain | 0.955 | 0.256 | 0.000 | -9.309 | Likely Pathogenic | 0.985 | Likely Pathogenic | Likely Pathogenic | -0.27 | Likely Benign | 0.0 | -0.34 | Likely Benign | -0.31 | Likely Benign | 0.97 | Ambiguous | 0.684 | Likely Pathogenic | -8.21 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.15 | Pathogenic | 0.69 | Tolerated | 0.3469 | 0.1415 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1657A>G | K553E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K553E missense variant is not reported in ClinVar (status: None) and has no entries in gnomAD. Prediction tools that agree on a benign effect include only SIFT, whereas the remaining tools (REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict pathogenicity. FoldX and Rosetta give uncertain results, and Foldetta also reports an uncertain stability change. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a deleterious effect. Thus, the variant is most likely pathogenic based on current predictions, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.012270 | Structured | 0.006539 | Uncertain | 0.949 | 0.246 | 0.000 | -17.415 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 1.08 | Ambiguous | 0.3 | 1.15 | Ambiguous | 1.12 | Ambiguous | 1.04 | Destabilizing | 0.828 | Likely Pathogenic | -3.85 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.35 | Pathogenic | 0.12 | Tolerated | 0.2890 | 0.0650 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1658A>C | K553T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant K553T is listed in ClinVar with an uncertain significance (ClinVar ID 2007142.0) and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include Rosetta and SIFT, whereas the majority of tools predict a pathogenic impact: REVEL, PROVEAN, both polyPhen‑2 HumDiv and HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (Likely Pathogenic). Uncertain results are reported by FoldX, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as Likely Pathogenic, and Foldetta as inconclusive. Overall, the consensus of the available predictions indicates that K553T is most likely pathogenic, which does not contradict the current ClinVar status of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.012270 | Structured | 0.006539 | Uncertain | 0.949 | 0.246 | 0.000 | Uncertain | 1 | -15.328 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 1.06 | Ambiguous | 0.2 | 0.48 | Likely Benign | 0.77 | Ambiguous | 0.79 | Ambiguous | 0.761 | Likely Pathogenic | -5.77 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.34 | Pathogenic | 0.14 | Tolerated | 3.37 | 35 | 0.1733 | 0.2619 | 0 | -1 | 3.2 | -27.07 | 218.2 | -10.7 | 0.0 | 0.0 | -0.2 | 0.5 | X | Potentially Pathogenic | Lys533 is located on an α-helix (res. Ala533-Val560). In the WT simulations, Lys533 packs against Phe513, and its amino side chain occasionally forms an ionic interaction with the carboxylate group of Glu512 from an opposing α-helix (res. Gln503-Tyr518). In the variant simulations, Thr533 is unable to reproduce these interactions, potentially weakening the integrity of the tertiary structure. Additionally, Thr533 forms a hydrogen bond with the backbone carbonyl group of Leu549 in the same helix, which could potentially weaken the secondary structure. Regardless, the residue swap does not cause significant structural effects based on the simulations. | ||||||||||||||||
| c.1658A>T | K553M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K553M is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include FoldX, Rosetta, Foldetta, and premPS, whereas the majority of tools predict it to be pathogenic: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Based on the overall distribution of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.012270 | Structured | 0.006539 | Uncertain | 0.949 | 0.246 | 0.000 | -16.086 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | -0.08 | Likely Benign | 0.0 | -0.06 | Likely Benign | -0.07 | Likely Benign | 0.32 | Likely Benign | 0.854 | Likely Pathogenic | -5.76 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.39 | Pathogenic | 0.01 | Affected | 0.0921 | 0.2756 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1659G>C | K553N 2D 3DClick to see structure in 3D Viewer AISynGAP1 K553N is not reported in ClinVar and is present in gnomAD (6-33438902-G-C). Functional prediction tools largely agree on a deleterious effect: SIFT is the sole benign predictor, whereas REVEL, SGM‑Consensus, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity. FoldX, Rosetta, and Foldetta return uncertain results. High‑accuracy assessments reinforce the pathogenic signal: AlphaMissense‑Optimized is pathogenic, SGM‑Consensus is likely pathogenic, and Foldetta remains uncertain. Overall, the consensus of the majority of tools indicates that K553N is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar entry (no contradiction). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.012270 | Structured | 0.006539 | Uncertain | 0.949 | 0.246 | 0.000 | 6-33438902-G-C | 1 | 6.20e-7 | -13.664 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.63 | Ambiguous | 0.0 | 0.62 | Ambiguous | 0.63 | Ambiguous | 1.11 | Destabilizing | 0.566 | Likely Pathogenic | -4.77 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.24 | Pathogenic | 0.11 | Tolerated | 3.37 | 35 | 0.2758 | 0.0926 | 0 | 1 | 0.4 | -14.07 | ||||||||||||||||||||||||
| c.1659G>T | K553N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K553N missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only SIFT, whereas the majority of algorithms (SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a pathogenic impact; FoldX, Rosetta, and Foldetta are inconclusive. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, SGM‑Consensus as likely pathogenic, and Foldetta as uncertain. Taken together, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.012270 | Structured | 0.006539 | Uncertain | 0.949 | 0.246 | 0.000 | -13.664 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 0.63 | Ambiguous | 0.0 | 0.62 | Ambiguous | 0.63 | Ambiguous | 1.11 | Destabilizing | 0.566 | Likely Pathogenic | -4.77 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.24 | Pathogenic | 0.11 | Tolerated | 3.37 | 35 | 0.2758 | 0.0926 | 0 | 1 | 0.4 | -14.07 | |||||||||||||||||||||||||||
| c.1661T>C | V554A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V554A is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are REVEL and FATHMM. All other evaluated predictors—SGM‑Consensus, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, and AlphaMissense‑Default—consistently predict a pathogenic impact. High‑accuracy assessments further support this: AlphaMissense‑Optimized is uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts Pathogenic. No prediction or stability result is missing or inconclusive. Based on the overwhelming majority of pathogenic predictions, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because none is available. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.020522 | Structured | 0.007349 | Uncertain | 0.955 | 0.226 | 0.000 | -9.730 | Likely Pathogenic | 0.870 | Likely Pathogenic | Ambiguous | 2.07 | Destabilizing | 0.1 | 2.34 | Destabilizing | 2.21 | Destabilizing | 2.00 | Destabilizing | 0.419 | Likely Benign | -3.97 | Deleterious | 0.998 | Probably Damaging | 0.981 | Probably Damaging | 3.22 | Benign | 0.02 | Affected | 0.2131 | 0.1633 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1664T>C | V555A 2D 3DClick to see structure in 3D Viewer AISynGAP1 V555A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that assess sequence conservation and structural impact uniformly indicate a deleterious effect: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM Consensus (derived from the majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) all classify the variant as pathogenic. No tool predicts a benign outcome. Predictions that are inconclusive—FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized—are listed as uncertain and do not alter the overall assessment. High‑accuracy methods give a pathogenic consensus from SGM and an uncertain result from AlphaMissense‑Optimized and Foldetta. Consequently, the variant is most likely pathogenic, and this conclusion is consistent with the absence of a ClinVar entry. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.013265 | Structured | 0.008218 | Uncertain | 0.943 | 0.225 | 0.000 | -8.781 | Likely Pathogenic | 0.857 | Likely Pathogenic | Ambiguous | 0.91 | Ambiguous | 0.0 | 0.90 | Ambiguous | 0.91 | Ambiguous | 1.04 | Destabilizing | 0.633 | Likely Pathogenic | -3.48 | Deleterious | 0.979 | Probably Damaging | 0.956 | Probably Damaging | -1.34 | Pathogenic | 0.01 | Affected | 0.2602 | 0.1761 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1667A>C | N556T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant N556T is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include SIFT, premPS, AlphaMissense‑Optimized, and Rosetta. Tools that predict a pathogenic effect are REVEL, SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized classifying the variant as benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) labeling it likely pathogenic, and Foldetta reporting an uncertain stability change. FoldX‑MD and Foldetta results are inconclusive and are treated as unavailable. Overall, the majority of evidence points to a pathogenic impact. This conclusion does not contradict ClinVar status, as no ClinVar classification exists for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.015078 | Structured | 0.008655 | Uncertain | 0.925 | 0.225 | 0.000 | -8.636 | Likely Pathogenic | 0.642 | Likely Pathogenic | Likely Benign | 1.03 | Ambiguous | 0.1 | 0.08 | Likely Benign | 0.56 | Ambiguous | 0.06 | Likely Benign | 0.695 | Likely Pathogenic | -4.73 | Deleterious | 0.996 | Probably Damaging | 0.990 | Probably Damaging | -1.36 | Pathogenic | 0.09 | Tolerated | 0.1189 | 0.3171 | 0 | 0 | 2.8 | -13.00 | |||||||||||||||||||||||||||||
| c.1670C>G | S557C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S557C is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include premPS and AlphaMissense‑Optimized, whereas the majority of other in silico predictors (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) classify the change as pathogenic. FoldX, Rosetta, and Foldetta provide uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized predicting benign, while the SGM‑Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—predicts pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains uncertain. Overall, the preponderance of evidence from multiple pathogenic predictors and the SGM‑Consensus suggests the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because the variant is not yet reported there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.028107 | Structured | 0.010261 | Uncertain | 0.924 | 0.215 | 0.000 | -9.845 | Likely Pathogenic | 0.577 | Likely Pathogenic | Likely Benign | 1.43 | Ambiguous | 0.1 | 1.74 | Ambiguous | 1.59 | Ambiguous | 0.49 | Likely Benign | 0.923 | Likely Pathogenic | -4.52 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | -1.77 | Pathogenic | 0.00 | Affected | 0.1287 | 0.5678 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||
| c.1672C>T | H558Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 H558Y missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, AlphaMissense‑Default, and AlphaMissense‑Optimized. Only FATHMM predicts a pathogenic outcome, while FoldX, premPS, and ESM1b are uncertain. High‑accuracy assessments further support a benign classification: AlphaMissense‑Optimized is benign; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) yields a benign result; and Foldetta also predicts benign. No prediction or folding stability result is missing or inconclusive. Overall, the variant is most likely benign, and this conclusion does not contradict any ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.033407 | Structured | 0.011039 | Uncertain | 0.897 | 0.200 | 0.000 | -7.208 | In-Between | 0.187 | Likely Benign | Likely Benign | -0.58 | Ambiguous | 0.0 | -0.36 | Likely Benign | -0.47 | Likely Benign | -0.56 | Ambiguous | 0.359 | Likely Benign | 0.55 | Neutral | 0.019 | Benign | 0.013 | Benign | -1.24 | Pathogenic | 1.00 | Tolerated | 0.0938 | 0.2363 | 0 | 2 | 1.9 | 26.03 | ||||||||||||||||||||||||||||||
| c.1673A>C | H558P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant H558P is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are SIFT and AlphaMissense‑Optimized; those that agree on a pathogenic effect include SGM‑Consensus (Likely Pathogenic), REVEL, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. Uncertain predictions come from FoldX and premPS. High‑accuracy assessments show AlphaMissense‑Optimized predicts benign, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) predicts Likely Pathogenic, and Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) predicts Pathogenic. Overall, the majority of tools, including the high‑accuracy ones, indicate a pathogenic effect. Thus, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.033407 | Structured | 0.011039 | Uncertain | 0.897 | 0.200 | 0.000 | -13.706 | Likely Pathogenic | 0.675 | Likely Pathogenic | Likely Benign | 0.63 | Ambiguous | 0.2 | 6.93 | Destabilizing | 3.78 | Destabilizing | 0.94 | Ambiguous | 0.782 | Likely Pathogenic | -6.28 | Deleterious | 0.999 | Probably Damaging | 0.989 | Probably Damaging | -1.25 | Pathogenic | 0.09 | Tolerated | 0.2061 | 0.2405 | 0 | -2 | 1.6 | -40.02 | |||||||||||||||||||||||||||||
| c.1673A>G | H558R 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant H558R is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from AlphaMissense‑Optimized, Rosetta, SIFT, and polyPhen‑2 HumVar, while pathogenic predictions arise from REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, ESM1b, and FATHMM. Four tools give inconclusive results: AlphaMissense‑Default, SGM‑Consensus, FoldX, and Foldetta. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign effect, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, remains uncertain. Overall, the majority of evidence points toward a pathogenic impact, which does not conflict with the ClinVar designation of uncertain significance. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.033407 | Structured | 0.011039 | Uncertain | 0.897 | 0.200 | 0.000 | Uncertain | 1 | -14.445 | Likely Pathogenic | 0.554 | Ambiguous | Likely Benign | -1.14 | Ambiguous | 0.1 | -0.23 | Likely Benign | -0.69 | Ambiguous | 1.03 | Destabilizing | 0.587 | Likely Pathogenic | -4.94 | Deleterious | 0.677 | Possibly Damaging | 0.239 | Benign | -1.24 | Pathogenic | 0.14 | Tolerated | 3.37 | 35 | 0.1500 | 0.1512 | 0 | 2 | -1.3 | 19.05 | |||||||||||||||||||||||||
| c.1675T>A | C559S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C559S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, and FATHMM. Tools that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. The remaining tools, premPS and AlphaMissense‑Optimized, are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta) as Benign. Based on the overall distribution of predictions, the variant is most likely pathogenic; this conclusion does not contradict the ClinVar status, which has no classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.016021 | Structured | 0.010460 | Uncertain | 0.842 | 0.204 | 0.000 | -10.756 | Likely Pathogenic | 0.785 | Likely Pathogenic | Ambiguous | -0.03 | Likely Benign | 0.0 | -0.31 | Likely Benign | -0.17 | Likely Benign | 0.65 | Ambiguous | 0.450 | Likely Benign | -7.26 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.58 | Benign | 0.64 | Tolerated | 0.3940 | 0.1319 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1676G>A | C559Y 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant C559Y is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include FoldX, Rosetta, Foldetta, premPS, SIFT, and FATHMM. Those that predict pathogenicity are REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Because the evidence is split evenly between benign and pathogenic predictions and the high‑accuracy tools disagree, the variant is best classified as of uncertain significance. This assessment does not contradict ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.016021 | Structured | 0.010460 | Uncertain | 0.842 | 0.204 | 0.000 | -11.767 | Likely Pathogenic | 0.868 | Likely Pathogenic | Ambiguous | -0.37 | Likely Benign | 0.0 | -0.19 | Likely Benign | -0.28 | Likely Benign | 0.36 | Likely Benign | 0.561 | Likely Pathogenic | -8.39 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 3.45 | Benign | 0.10 | Tolerated | 0.2094 | 0.2135 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1676G>C | C559S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C559S has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, SIFT, and FATHMM. Those that predict a pathogenic impact are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Two tools give inconclusive results: premPS (Uncertain) and AlphaMissense‑Optimized (Uncertain). High‑accuracy assessments show SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, Foldetta (combining FoldX‑MD and Rosetta) as Benign, and AlphaMissense‑Optimized as Uncertain. Overall, the majority of predictions lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which is currently unreported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.016021 | Structured | 0.010460 | Uncertain | 0.842 | 0.204 | 0.000 | -10.756 | Likely Pathogenic | 0.785 | Likely Pathogenic | Ambiguous | -0.03 | Likely Benign | 0.0 | -0.31 | Likely Benign | -0.17 | Likely Benign | 0.65 | Ambiguous | 0.469 | Likely Benign | -7.26 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.58 | Benign | 0.64 | Tolerated | 0.3940 | 0.1319 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1679T>C | V560A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant V560A is not reported in ClinVar (ClinVar status: none) and has no entry in gnomAD (gnomAD ID: none). Prediction tools that agree on a benign effect include REVEL, SIFT, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect comprise SGM‑Consensus, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default. FoldX, Rosetta, and Foldetta provide uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic; Foldetta remains uncertain. Overall, the majority of predictions (eight pathogenic vs. three benign) indicate that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.021381 | Structured | 0.013872 | Uncertain | 0.853 | 0.204 | 0.000 | -8.260 | Likely Pathogenic | 0.701 | Likely Pathogenic | Likely Benign | 0.54 | Ambiguous | 0.1 | 1.33 | Ambiguous | 0.94 | Ambiguous | 1.19 | Destabilizing | 0.447 | Likely Benign | -3.15 | Deleterious | 0.911 | Possibly Damaging | 0.657 | Possibly Damaging | -1.20 | Pathogenic | 0.31 | Tolerated | 0.2549 | 0.2212 | 0 | 0 | -2.4 | -28.05 | |||||||||||||||||||||||||||||
| c.1681T>A | F561I 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant F561I is not reported in ClinVar (ClinVar ID None) but is present in gnomAD (variant ID 6‑33440733‑T‑A). Prediction tools that agree on a benign effect include only SIFT. All other evaluated predictors—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No prediction or folding stability result is missing or inconclusive. Based on the overwhelming consensus of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.024826 | Structured | 0.018013 | Uncertain | 0.903 | 0.196 | 0.000 | 6-33440733-T-A | 1 | 6.32e-7 | -11.708 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 3.82 | Destabilizing | 0.1 | 2.13 | Destabilizing | 2.98 | Destabilizing | 1.46 | Destabilizing | 0.710 | Likely Pathogenic | -5.97 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | -1.06 | Pathogenic | 0.09 | Tolerated | 3.37 | 35 | 0.1825 | 0.1925 | 0 | 1 | 1.7 | -34.02 | ||||||||||||||||||||||||
| c.1684C>A | P562T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P562T is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign outcome; the only uncertain predictions come from Foldetta and premPS. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, while Foldetta remains uncertain. Taken together, the overwhelming majority of computational evidence indicates that P562T is likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.022306 | Structured | 0.023606 | Uncertain | 0.893 | 0.200 | 0.000 | -14.747 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 2.62 | Destabilizing | 0.1 | 0.51 | Ambiguous | 1.57 | Ambiguous | 0.81 | Ambiguous | 0.697 | Likely Pathogenic | -7.96 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 0.58 | Pathogenic | 0.00 | Affected | 0.1892 | 0.3454 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||
| c.1685C>G | P562R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P562R has no ClinVar entry and is not reported in gnomAD. Across the available in‑silico predictors, none classify the change as benign; the majority (REVEL, FoldX, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, SGM‑Consensus) predict it to be pathogenic. Two tools (Rosetta and premPS) return uncertain results. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized scores it as pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates pathogenicity, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts a destabilizing, pathogenic impact. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.022306 | Structured | 0.023606 | Uncertain | 0.893 | 0.200 | 0.000 | -17.309 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 5.88 | Destabilizing | 1.2 | 0.96 | Ambiguous | 3.42 | Destabilizing | 0.82 | Ambiguous | 0.820 | Likely Pathogenic | -8.96 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 0.58 | Pathogenic | 0.02 | Affected | 0.1640 | 0.2489 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.1689G>C | R563S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 R563S missense change is not reported in ClinVar and has no entries in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Five tools (FoldX, Rosetta, Foldetta, premPS, AlphaMissense‑Optimized) return uncertain or inconclusive results. High‑accuracy assessments are likewise ambiguous: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is a 2‑to‑2 tie and therefore unavailable; Foldetta also reports an uncertain stability change. Consequently, the evidence does not strongly support either benign or pathogenic status. The variant’s classification is therefore inconclusive and does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.031987 | Uncertain | 0.876 | 0.209 | 0.000 | -6.503 | Likely Benign | 0.949 | Likely Pathogenic | Ambiguous | 1.16 | Ambiguous | 0.1 | 1.46 | Ambiguous | 1.31 | Ambiguous | 0.69 | Ambiguous | 0.251 | Likely Benign | -4.49 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.48 | Benign | 0.23 | Tolerated | 0.3031 | 0.2534 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||
| c.1689G>T | R563S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant R563S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, SIFT, ESM1b, and FATHMM, while pathogenic predictions arise from PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. The remaining tools—FoldX, Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized—yield uncertain or inconclusive results and are treated as unavailable. High‑accuracy assessments are likewise inconclusive: AlphaMissense‑Optimized is uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is a 2‑to‑2 tie, and Foldetta is uncertain. Consequently, the evidence does not strongly favor either outcome. Based on the current predictions, the variant is most likely benign, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.039760 | Structured | 0.031987 | Uncertain | 0.876 | 0.209 | 0.000 | -6.503 | Likely Benign | 0.949 | Likely Pathogenic | Ambiguous | 1.16 | Ambiguous | 0.1 | 1.46 | Ambiguous | 1.31 | Ambiguous | 0.69 | Ambiguous | 0.251 | Likely Benign | -4.49 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.48 | Benign | 0.23 | Tolerated | 0.3031 | 0.2534 | 0 | -1 | 3.7 | -69.11 | ||||||||||||||||||||||||||||||
| c.1691A>C | E564A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E564A is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include premPS and SIFT, whereas the majority of tools predict a pathogenic impact: SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Uncertain results are reported by FoldX, Rosetta, and Foldetta. High‑accuracy assessments reinforce the pathogenic prediction: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta remains uncertain. Overall, the preponderance of evidence from multiple independent predictors indicates that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.023534 | Structured | 0.038418 | Uncertain | 0.891 | 0.208 | 0.000 | -14.506 | Likely Pathogenic | 0.957 | Likely Pathogenic | Likely Pathogenic | 0.64 | Ambiguous | 0.1 | 1.49 | Ambiguous | 1.07 | Ambiguous | 0.24 | Likely Benign | 0.765 | Likely Pathogenic | -5.91 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | -1.33 | Pathogenic | 0.12 | Tolerated | 0.3201 | 0.4918 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1696A>G | K566E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K566E missense variant is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Across the evaluated tools, every predictor that returned a definitive call classified the variant as pathogenic: SGM‑Consensus, REVEL, FoldX, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool reported a benign outcome; the only inconclusive result was from Rosetta, which was labeled “Uncertain.” High‑accuracy assessments—AlphaMissense‑Optimized, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta—each returned a pathogenic prediction. Consequently, the variant is most likely pathogenic based on the consensus of predictive algorithms, and this assessment does not contradict any existing ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.027463 | Structured | 0.047887 | Uncertain | 0.924 | 0.219 | 0.000 | -10.759 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 3.76 | Destabilizing | 0.2 | 1.13 | Ambiguous | 2.45 | Destabilizing | 1.58 | Destabilizing | 0.832 | Likely Pathogenic | -3.58 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.38 | Pathogenic | 0.04 | Affected | 0.3275 | 0.0850 | 0 | 1 | 0.4 | 0.94 | |||||||||||||||||||||||||||||
| c.1697A>C | K566T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 K566T missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only Rosetta, which scores the variant as benign. All other evaluated predictors—REVEL, SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized—classify the variant as pathogenic. FoldX, Foldetta, and premPS are inconclusive, giving uncertain results. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic; Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for K566T, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.027463 | Structured | 0.047887 | Uncertain | 0.924 | 0.219 | 0.000 | -12.866 | Likely Pathogenic | 0.986 | Likely Pathogenic | Likely Pathogenic | 1.70 | Ambiguous | 0.2 | -0.21 | Likely Benign | 0.75 | Ambiguous | 0.99 | Ambiguous | 0.788 | Likely Pathogenic | -5.37 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.43 | Pathogenic | 0.04 | Affected | 0.1804 | 0.3847 | 0 | -1 | 3.2 | -27.07 | |||||||||||||||||||||||||||||
| c.1697A>T | K566M 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K566M is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign are Foldetta and premPS, whereas the remaining tools (REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict it to be pathogenic; FoldX and Rosetta give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Based on the overall distribution of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because the variant is not yet catalogued there. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.027463 | Structured | 0.047887 | Uncertain | 0.924 | 0.219 | 0.000 | -13.208 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.80 | Ambiguous | 0.2 | -0.51 | Ambiguous | 0.15 | Likely Benign | 0.37 | Likely Benign | 0.804 | Likely Pathogenic | -5.51 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.47 | Pathogenic | 0.01 | Affected | 0.0936 | 0.3826 | 0 | -1 | 5.8 | 3.02 | |||||||||||||||||||||||||||||
| c.1698G>C | K566N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K566N is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that assess pathogenicity uniformly indicate a deleterious effect: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity, while Rosetta is uncertain. No tool predicts a benign outcome. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Consequently, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.027463 | Structured | 0.047887 | Uncertain | 0.924 | 0.219 | 0.000 | -11.255 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 2.83 | Destabilizing | 0.3 | 1.33 | Ambiguous | 2.08 | Destabilizing | 1.32 | Destabilizing | 0.683 | Likely Pathogenic | -4.04 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.43 | Pathogenic | 0.03 | Affected | 3.37 | 35 | 0.3042 | 0.1567 | 0 | 1 | 0.4 | -14.07 | |||||||||||||||||||||||||||
| c.1698G>T | K566N 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant K566N is listed in gnomAD (variant ID 6‑33440750‑G‑T) but has no ClinVar entry. All available in‑silico predictors that provide a definitive call classify the substitution as pathogenic: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The only tool with an inconclusive result is Rosetta (Uncertain), which is treated as unavailable evidence. High‑accuracy methods reinforce this assessment: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. Consequently, the variant is most likely pathogenic based on current predictions, and this conclusion does not contradict any ClinVar status because none is reported. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.027463 | Structured | 0.047887 | Uncertain | 0.924 | 0.219 | 0.000 | 6-33440750-G-T | -11.255 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 2.83 | Destabilizing | 0.3 | 1.33 | Ambiguous | 2.08 | Destabilizing | 1.32 | Destabilizing | 0.683 | Likely Pathogenic | -4.04 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.43 | Pathogenic | 0.03 | Affected | 3.37 | 35 | 0.3042 | 0.1567 | 0 | 1 | 0.4 | -14.07 | ||||||||||||||||||||||||||
| c.1699G>A | E567K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E567K is not reported in ClinVar (no entry) and is absent from gnomAD. Prediction tools cluster into two groups: benign calls come from REVEL, FoldX, SIFT, and FATHMM, while pathogenic calls arise from Rosetta, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus (likely pathogenic). High‑accuracy methods reinforce the pathogenic assessment: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates likely pathogenic, whereas Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, remains uncertain. PremPS is likewise inconclusive. Overall, the majority of evidence points to a pathogenic effect, and this conclusion is not contradicted by ClinVar status or gnomAD presence. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.021816 | Structured | 0.051008 | Uncertain | 0.916 | 0.234 | 0.000 | -15.568 | Likely Pathogenic | 0.984 | Likely Pathogenic | Likely Pathogenic | -0.18 | Likely Benign | 0.0 | 2.61 | Destabilizing | 1.22 | Ambiguous | 0.65 | Ambiguous | 0.380 | Likely Benign | -3.75 | Deleterious | 0.999 | Probably Damaging | 0.994 | Probably Damaging | 3.45 | Benign | 0.16 | Tolerated | 0.2115 | 0.5552 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.16G>A | A6T 2D AIThe SynGAP1 missense variant A6T is reported in gnomAD (variant ID 6-33420280‑G‑A) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates a likely benign outcome. High‑accuracy assessments confirm this: AlphaMissense‑Optimized is benign, and the SGM‑Consensus also reports likely benign; Foldetta results are not available. Overall, the consensus of the majority of prediction tools and the high‑accuracy methods points to a benign impact. This conclusion is consistent with the absence of a ClinVar pathogenic classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.566480 | Disordered | 0.549054 | Binding | 0.377 | 0.920 | 0.875 | 6-33420280-G-A | -3.711 | Likely Benign | 0.130 | Likely Benign | Likely Benign | 0.089 | Likely Benign | -0.13 | Neutral | 0.027 | Benign | 0.004 | Benign | 4.11 | Benign | 0.00 | Affected | 4.32 | 1 | 0.1521 | 0.6836 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||||||||||||||
| c.1700A>C | E567A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E567A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, SIFT, and FATHMM, while pathogenic predictions arise from SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, and AlphaMissense‑Default. Four tools (Foldetta, premPS, Rosetta, AlphaMissense‑Optimized) yield uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, SGM‑Consensus as Likely Pathogenic (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN), and Foldetta as uncertain. Overall, the majority of evidence points toward a pathogenic effect. This conclusion is not contradicted by ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.021816 | Structured | 0.051008 | Uncertain | 0.916 | 0.234 | 0.000 | -12.854 | Likely Pathogenic | 0.902 | Likely Pathogenic | Ambiguous | 0.43 | Likely Benign | 0.1 | 1.79 | Ambiguous | 1.11 | Ambiguous | 0.63 | Ambiguous | 0.418 | Likely Benign | -5.74 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.43 | Benign | 0.17 | Tolerated | 0.3170 | 0.5189 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1700A>G | E567G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E567G missense variant is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on benign impact include REVEL, SIFT, and FATHMM, whereas those that agree on pathogenic impact include AlphaMissense‑Default, ESM1b, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, Rosetta, and Foldetta. Predictions that are inconclusive are FoldX, premPS, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as pathogenic. Taken together, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.021816 | Structured | 0.051008 | Uncertain | 0.916 | 0.234 | 0.000 | -13.402 | Likely Pathogenic | 0.912 | Likely Pathogenic | Ambiguous | 1.47 | Ambiguous | 0.0 | 3.48 | Destabilizing | 2.48 | Destabilizing | 0.75 | Ambiguous | 0.456 | Likely Benign | -6.67 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.40 | Benign | 0.07 | Tolerated | 0.2586 | 0.4715 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1703T>C | V568A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 V568A missense variant is not reported in ClinVar (ClinVar status: none) but is present in the gnomAD database (gnomAD ID: 6‑33440755‑T‑C). Prediction tools that agree on a benign effect include only SIFT, whereas the majority of tools predict a pathogenic impact: REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (Likely Pathogenic). Uncertain or inconclusive results come from FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as pathogenic, and Foldetta as uncertain. Based on the preponderance of pathogenic predictions and the consensus from high‑accuracy tools, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status (which has no entry). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.024826 | Structured | 0.053503 | Uncertain | 0.937 | 0.257 | 0.000 | 6-33440755-T-C | 2 | 1.25e-6 | -10.929 | Likely Pathogenic | 0.946 | Likely Pathogenic | Ambiguous | 1.90 | Ambiguous | 0.1 | 1.77 | Ambiguous | 1.84 | Ambiguous | 2.16 | Destabilizing | 0.834 | Likely Pathogenic | -3.82 | Deleterious | 0.999 | Probably Damaging | 0.990 | Probably Damaging | -1.38 | Pathogenic | 0.06 | Tolerated | 3.37 | 35 | 0.2413 | 0.1961 | 0 | 0 | -2.4 | -28.05 | ||||||||||||||||||||||||
| c.1709C>A | A570D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A570D is not reported in ClinVar (ClinVar status: not listed) and is absent from gnomAD (gnomAD ID: none). Prediction tools that assess pathogenicity unanimously classify the variant as deleterious: SGM‑Consensus (Likely Pathogenic), REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. No tool predicts a benign effect, so the benign group is empty. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts Pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also indicates Likely Pathogenic; and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts Pathogenic. Based on the collective evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.046336 | Structured | 0.054494 | Uncertain | 0.932 | 0.263 | 0.000 | -14.117 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 2.47 | Destabilizing | 1.2 | 2.33 | Destabilizing | 2.40 | Destabilizing | 1.36 | Destabilizing | 0.805 | Likely Pathogenic | -5.31 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.28 | Pathogenic | 0.03 | Affected | 0.2006 | 0.2206 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1709C>T | A570V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A570V missense variant is catalogued in gnomAD (ID 6‑33440761‑C‑T) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from premPS and SIFT, while pathogenic predictions are made by REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. Four tools report uncertainty: FoldX, Rosetta, Foldetta, and AlphaMissense‑Optimized. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as Likely Pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as uncertain. Overall, the majority of evidence points to a pathogenic effect. This conclusion is not contradicted by ClinVar status, which currently contains no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.046336 | Structured | 0.054494 | Uncertain | 0.932 | 0.263 | 0.000 | 6-33440761-C-T | 1 | 6.22e-7 | -13.083 | Likely Pathogenic | 0.882 | Likely Pathogenic | Ambiguous | 0.88 | Ambiguous | 0.3 | 1.63 | Ambiguous | 1.26 | Ambiguous | 0.46 | Likely Benign | 0.669 | Likely Pathogenic | -3.75 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | -1.30 | Pathogenic | 0.06 | Tolerated | 3.37 | 35 | 0.1050 | 0.3173 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||
| c.1718G>C | R573P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R573P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity all converge on a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all indicate a pathogenic or likely pathogenic outcome. No tool in the dataset predicts a benign effect. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized classifies the variant as pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports it as likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts a pathogenic impact. Based on the uniform predictions, the variant is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.134866 | Structured | 0.032433 | Uncertain | 0.934 | 0.235 | 0.000 | -16.684 | Likely Pathogenic | 1.000 | Likely Pathogenic | Likely Pathogenic | 5.76 | Destabilizing | 1.1 | 8.58 | Destabilizing | 7.17 | Destabilizing | 1.21 | Destabilizing | 0.831 | Likely Pathogenic | -5.78 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.47 | Pathogenic | 0.01 | Affected | 0.1961 | 0.3239 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1723C>A | R575S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R575S is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify it as benign include only SIFT, whereas the majority—REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict pathogenicity. FoldX, Rosetta, Foldetta, and premPS give uncertain results, which are treated as unavailable evidence. High‑accuracy methods reinforce the pathogenic signal: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta remains uncertain. Overall, the preponderance of evidence points to a pathogenic effect for R575S, and this assessment does not contradict the absence of a ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.061840 | Structured | 0.021362 | Uncertain | 0.916 | 0.259 | 0.000 | -11.124 | Likely Pathogenic | 0.957 | Likely Pathogenic | Likely Pathogenic | 1.66 | Ambiguous | 0.1 | 0.55 | Ambiguous | 1.11 | Ambiguous | 0.76 | Ambiguous | 0.582 | Likely Pathogenic | -2.71 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.21 | Pathogenic | 0.33 | Tolerated | 0.2669 | 0.2394 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||
| c.1724G>C | R575P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R575P is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include only SIFT, whereas all other evaluated algorithms—REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—classify the variant as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also predicts pathogenicity. Based on the overwhelming consensus of pathogenic predictions and the absence of any benign consensus, the variant is most likely pathogenic. This conclusion is not contradicted by ClinVar status, as no ClinVar entry exists for R575P. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.061840 | Structured | 0.021362 | Uncertain | 0.916 | 0.259 | 0.000 | -16.008 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 3.50 | Destabilizing | 0.1 | 4.97 | Destabilizing | 4.24 | Destabilizing | 1.13 | Destabilizing | 0.774 | Likely Pathogenic | -3.69 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.33 | Pathogenic | 0.10 | Tolerated | 0.2080 | 0.2670 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1726T>A | C576S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C576S is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign predictions from REVEL and FATHMM, while the majority of other in silico methods (premPS, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, ESM1b, AlphaMissense‑Default) predict pathogenicity. FoldX and Rosetta give uncertain stability changes, and Foldetta likewise reports no definitive effect. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized predicts pathogenic, and the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic. Foldetta remains inconclusive. Overall, the preponderance of evidence from multiple pathogenic‑oriented tools and the high‑accuracy predictions indicates that C576S is most likely pathogenic, which is consistent with the absence of a ClinVar entry and gnomAD data. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.017684 | Uncertain | 0.913 | 0.245 | 0.000 | -10.474 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.77 | Ambiguous | 0.1 | 1.57 | Ambiguous | 1.17 | Ambiguous | 1.61 | Destabilizing | 0.414 | Likely Benign | -8.91 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.40 | Benign | 0.02 | Affected | 0.4968 | 0.1464 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1727G>A | C576Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C576Y is not reported in ClinVar and has no gnomAD allele. Prediction tools largely agree on a deleterious effect: FATHMM is the sole benign predictor, whereas SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default and AlphaMissense‑Optimized all classify the change as pathogenic. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports a likely pathogenic outcome; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also indicates pathogenicity. No tool yields an inconclusive result. Based on the consensus of available predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.017684 | Uncertain | 0.913 | 0.245 | 0.000 | -13.891 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 8.77 | Destabilizing | 0.5 | 3.90 | Destabilizing | 6.34 | Destabilizing | 0.63 | Ambiguous | 0.612 | Likely Pathogenic | -9.98 | Deleterious | 0.999 | Probably Damaging | 0.996 | Probably Damaging | 3.38 | Benign | 0.00 | Affected | 0.1398 | 0.3009 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1727G>C | C576S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C576S is not reported in ClinVar and has no entries in gnomAD. Prediction tools that indicate a benign effect are limited to FATHMM, whereas the remaining 11 tools (SGM‑Consensus, REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) all predict a pathogenic or likely pathogenic outcome. High‑accuracy methods reinforce this trend: AlphaMissense‑Optimized reports pathogenic; the SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, also returns likely pathogenic; Foldetta, a protein‑folding stability predictor combining FoldX‑MD and Rosetta, is inconclusive. Folding‑stability tools FoldX and Rosetta individually yield uncertain results and are treated as unavailable. Taken together, the majority of evidence points to a pathogenic effect. This conclusion is consistent with the absence of ClinVar annotation and gnomAD data, so there is no contradiction with existing database status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.017684 | Uncertain | 0.913 | 0.245 | 0.000 | -10.474 | Likely Pathogenic | 0.991 | Likely Pathogenic | Likely Pathogenic | 0.77 | Ambiguous | 0.1 | 1.57 | Ambiguous | 1.17 | Ambiguous | 1.61 | Destabilizing | 0.523 | Likely Pathogenic | -8.91 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.40 | Benign | 0.02 | Affected | 0.4968 | 0.1464 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1730C>A | A577E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A577E missense variant is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, PROVEAN, and SIFT, whereas those that agree on a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is classified as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, while Foldetta (combining FoldX‑MD and Rosetta outputs) is also uncertain; Rosetta and premPS are inconclusive. Overall, the majority of evaluated tools (five pathogenic vs. four benign) and the SGM‑Consensus support a pathogenic classification. Thus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.113710 | Structured | 0.019074 | Uncertain | 0.913 | 0.239 | 0.000 | -9.607 | Likely Pathogenic | 0.794 | Likely Pathogenic | Ambiguous | 0.29 | Likely Benign | 0.0 | 0.72 | Ambiguous | 0.51 | Ambiguous | 0.62 | Ambiguous | 0.419 | Likely Benign | -1.91 | Neutral | 0.999 | Probably Damaging | 0.996 | Probably Damaging | -1.12 | Pathogenic | 0.76 | Tolerated | 0.1487 | 0.1799 | 0 | -1 | -5.3 | 58.04 | |||||||||||||||||||||||||||||
| c.1730C>T | A577V 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A577V is catalogued in gnomAD (ID 6‑33440782‑C‑T) but has no ClinVar entry. Functional prediction tools that agree on a benign effect include REVEL, Rosetta, premPS, PROVEAN, SIFT, ESM1b, and AlphaMissense‑Optimized. Tools that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and FATHMM. High‑accuracy assessments further support a benign interpretation: AlphaMissense‑Optimized scores benign; the SGM consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) yields benign; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also reports benign. No prediction or stability result is missing or inconclusive in these key analyses. Consequently, the variant is most likely benign, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.113710 | Structured | 0.019074 | Uncertain | 0.913 | 0.239 | 0.000 | 6-33440782-C-T | 2 | 1.24e-6 | -4.265 | Likely Benign | 0.432 | Ambiguous | Likely Benign | 0.69 | Ambiguous | 0.1 | 0.00 | Likely Benign | 0.35 | Likely Benign | 0.16 | Likely Benign | 0.417 | Likely Benign | -2.11 | Neutral | 0.997 | Probably Damaging | 0.976 | Probably Damaging | -1.32 | Pathogenic | 0.26 | Tolerated | 3.37 | 34 | 0.1278 | 0.4790 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||
| c.1732G>A | E578K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E578K is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, and SIFT, whereas polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, and AlphaMissense‑Default predict a pathogenic outcome. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show that the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) classifies the variant as Likely Pathogenic, AlphaMissense‑Optimized remains Uncertain, and Foldetta predicts a benign effect. Overall, the majority of tools (seven benign vs. five pathogenic) suggest a benign impact, and this assessment does not contradict the absence of ClinVar evidence. Thus, the variant is most likely benign based on the current predictions, with no conflicting ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -13.391 | Likely Pathogenic | 0.870 | Likely Pathogenic | Ambiguous | 0.07 | Likely Benign | 0.1 | -0.19 | Likely Benign | -0.06 | Likely Benign | -0.27 | Likely Benign | 0.450 | Likely Benign | -1.65 | Neutral | 0.996 | Probably Damaging | 0.987 | Probably Damaging | -1.30 | Pathogenic | 0.49 | Tolerated | 0.1954 | 0.5480 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1733A>C | E578A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E578A missense change is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from FoldX, Rosetta, Foldetta, premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized; pathogenic predictions come from REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. ESM1b is uncertain. High‑accuracy assessment shows AlphaMissense‑Optimized predicts benign, while the SGM consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) favors pathogenic, and Foldetta also predicts benign. Overall, the majority of conventional tools lean benign, but the high‑accuracy consensus and AlphaMissense‑Optimized suggest a pathogenic effect. Thus, the variant is most likely pathogenic according to the most reliable predictors, and this assessment does not contradict the absence of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -7.369 | In-Between | 0.604 | Likely Pathogenic | Likely Benign | 0.34 | Likely Benign | 0.1 | -0.48 | Likely Benign | -0.07 | Likely Benign | -0.02 | Likely Benign | 0.525 | Likely Pathogenic | -2.30 | Neutral | 0.999 | Probably Damaging | 0.998 | Probably Damaging | -1.40 | Pathogenic | 0.44 | Tolerated | 0.3326 | 0.5118 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||
| c.1733A>G | E578G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E578G missense change is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include premPS, PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, polyPhen‑2 (HumDiv and HumVar), FATHMM, and AlphaMissense‑Default. Four tools give uncertain or inconclusive results (FoldX, Rosetta, Foldetta, ESM1b). High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus—derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN—predicts pathogenic. Foldetta, a protein‑folding stability method, yields an uncertain result. Overall, the majority of high‑confidence predictions lean toward pathogenicity, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.059222 | Structured | 0.020971 | Uncertain | 0.902 | 0.240 | 0.000 | -7.680 | In-Between | 0.601 | Likely Pathogenic | Likely Benign | 0.98 | Ambiguous | 0.1 | 0.63 | Ambiguous | 0.81 | Ambiguous | 0.08 | Likely Benign | 0.589 | Likely Pathogenic | -2.40 | Neutral | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.49 | Pathogenic | 0.19 | Tolerated | 0.2608 | 0.4843 | 0 | -2 | 3.1 | -72.06 | ||||||||||||||||||||||||||||||
| c.1736G>C | R579P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R579P is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts a benign outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. No predictions or stability results are missing or inconclusive. Based on the unanimous pathogenic predictions and the absence of any ClinVar or gnomAD evidence to the contrary, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.053060 | Structured | 0.022872 | Uncertain | 0.877 | 0.244 | 0.000 | -14.826 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 3.00 | Destabilizing | 0.2 | 6.36 | Destabilizing | 4.68 | Destabilizing | 0.93 | Ambiguous | 0.821 | Likely Pathogenic | -6.26 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.41 | Pathogenic | 0.01 | Affected | 0.2207 | 0.3081 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.173T>A | M58K 2D AIThe SynGAP1 missense variant M58K is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a likely benign classification. Foldetta, a protein‑folding stability method combining FoldX‑MD and Rosetta outputs, has no available result for this variant. Overall, the majority of evidence points to a benign impact. There is no ClinVar entry to contradict this assessment, so the variant is most likely benign based on current predictions. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.188120 | Structured | 0.484415 | Uncertain | 0.515 | 0.665 | 0.000 | -6.059 | Likely Benign | 0.939 | Likely Pathogenic | Ambiguous | 0.199 | Likely Benign | -1.52 | Neutral | 0.018 | Benign | 0.184 | Benign | 4.08 | Benign | 0.00 | Affected | 0.1615 | 0.1163 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||||||||||||
| c.173T>G | M58R 2D AIThe SynGAP1 missense variant M58R is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and FATHMM. Tools that predict a pathogenic effect are SIFT and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain, and no Foldetta stability assessment is available. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Benign. High‑accuracy assessments therefore indicate a benign likelihood: AlphaMissense‑Optimized is inconclusive, SGM‑Consensus is benign, and Foldetta data are missing. Based on the collective predictions, the variant is most likely benign, and this conclusion does not contradict the ClinVar status, which has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.188120 | Structured | 0.484415 | Uncertain | 0.515 | 0.665 | 0.000 | -5.035 | Likely Benign | 0.940 | Likely Pathogenic | Ambiguous | 0.237 | Likely Benign | -1.78 | Neutral | 0.042 | Benign | 0.184 | Benign | 4.07 | Benign | 0.00 | Affected | 0.1745 | 0.1113 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||||||||||||
| c.1744G>A | E582K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E582K missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Functional prediction tools cluster into two groups: benign predictions come from REVEL, FoldX, Rosetta, premPS, PROVEAN, SIFT, and FATHMM, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. AlphaMissense‑Optimized is uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is inconclusive, and Foldetta predicts a benign effect on protein stability. Taken together, the majority of evidence points to a benign impact, and this conclusion does not contradict any ClinVar annotation (none is available). Thus, the variant is most likely benign. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -11.826 | Likely Pathogenic | 0.814 | Likely Pathogenic | Ambiguous | 0.20 | Likely Benign | 0.1 | 0.07 | Likely Benign | 0.14 | Likely Benign | 0.02 | Likely Benign | 0.168 | Likely Benign | -1.83 | Neutral | 0.994 | Probably Damaging | 0.994 | Probably Damaging | 3.31 | Benign | 0.33 | Tolerated | 0.2038 | 0.3762 | 0 | 1 | -0.4 | -0.94 | ||||||||||||||||||||||||||||||
| c.1745A>C | E582A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E582A missense variant is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, Rosetta, Foldetta, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Default. Two tools (FoldX and ESM1b) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as benign. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the lack of ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -7.432 | In-Between | 0.661 | Likely Pathogenic | Likely Benign | 0.78 | Ambiguous | 0.2 | 0.15 | Likely Benign | 0.47 | Likely Benign | 0.27 | Likely Benign | 0.263 | Likely Benign | -2.99 | Deleterious | 0.998 | Probably Damaging | 0.999 | Probably Damaging | 3.19 | Benign | 0.26 | Tolerated | 0.3236 | 0.4000 | 0 | -1 | 5.3 | -58.04 | ||||||||||||||||||||||||||||||
| c.1745A>G | E582G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E582G is not reported in ClinVar (ClinVar ID: None) and has no entry in gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include REVEL, premPS, SIFT, FATHMM, and AlphaMissense‑Optimized. Those that predict a pathogenic effect are SGM‑Consensus, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. FoldX, Rosetta, and Foldetta give uncertain or inconclusive results. High‑accuracy methods give mixed signals: AlphaMissense‑Optimized predicts benign, the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic, and Foldetta’s stability assessment is uncertain. Overall, six tools predict pathogenicity while five predict benign, and the high‑accuracy consensus is split. Thus, the variant is most likely pathogenic based on the preponderance of evidence, and this assessment does not contradict ClinVar status, which currently has no classification for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.102787 | Structured | 0.033838 | Uncertain | 0.845 | 0.235 | 0.000 | -9.621 | Likely Pathogenic | 0.630 | Likely Pathogenic | Likely Benign | 1.35 | Ambiguous | 0.2 | 1.24 | Ambiguous | 1.30 | Ambiguous | 0.37 | Likely Benign | 0.224 | Likely Benign | -3.95 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 3.13 | Benign | 0.13 | Tolerated | 0.2835 | 0.3325 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1748A>C | D583A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 D583A missense variant is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include Rosetta, Foldetta, premPS, SIFT, and ESM1b, while those that agree on a pathogenic effect are SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; FoldX is uncertain and thus not counted. High‑accuracy methods give AlphaMissense‑Optimized as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. Overall, the majority of predictions (10 pathogenic vs. 5 benign) and the two high‑accuracy pathogenic calls outweigh the single high‑accuracy benign call, indicating that the variant is most likely pathogenic. This assessment does not contradict ClinVar status, as no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.116183 | Structured | 0.038478 | Uncertain | 0.805 | 0.247 | 0.000 | -4.545 | Likely Benign | 0.967 | Likely Pathogenic | Likely Pathogenic | 0.98 | Ambiguous | 0.2 | -0.16 | Likely Benign | 0.41 | Likely Benign | 0.13 | Likely Benign | 0.787 | Likely Pathogenic | -7.64 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.39 | Pathogenic | 0.14 | Tolerated | 0.3025 | 0.3705 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||
| c.1751T>C | I584T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I584T is not reported in ClinVar and is absent from gnomAD. Consensus from most in silico predictors indicates a pathogenic effect: REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify it as likely pathogenic. Only AlphaMissense‑Optimized predicts a benign outcome, while Rosetta and Foldetta are uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign, SGM‑Consensus as likely pathogenic, and Foldetta as uncertain. With the overwhelming majority of tools supporting pathogenicity and no ClinVar entry to contradict this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.059222 | Structured | 0.046673 | Uncertain | 0.846 | 0.244 | 0.000 | -10.413 | Likely Pathogenic | 0.765 | Likely Pathogenic | Likely Benign | 2.05 | Destabilizing | 0.1 | 1.70 | Ambiguous | 1.88 | Ambiguous | 1.66 | Destabilizing | 0.748 | Likely Pathogenic | -4.63 | Deleterious | 0.999 | Probably Damaging | 0.993 | Probably Damaging | -1.11 | Pathogenic | 0.02 | Affected | 0.0911 | 0.0608 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1753G>A | A585T 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A585T is reported in gnomAD (ID 6‑33440805‑G‑A) but has no ClinVar entry. Functional prediction tools cluster into two groups: benign predictions come from REVEL, premPS, PROVEAN, and SIFT, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, and AlphaMissense‑Default. Four tools (FoldX, Rosetta, Foldetta, AlphaMissense‑Optimized) give uncertain results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the balance of evidence favors a pathogenic effect for A585T. This conclusion is not contradicted by ClinVar, which contains no classification for the variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.055884 | Uncertain | 0.880 | 0.244 | 0.000 | 6-33440805-G-A | 13 | 8.05e-6 | -10.063 | Likely Pathogenic | 0.876 | Likely Pathogenic | Ambiguous | 1.66 | Ambiguous | 0.2 | 1.97 | Ambiguous | 1.82 | Ambiguous | 0.23 | Likely Benign | 0.465 | Likely Benign | -1.73 | Neutral | 1.000 | Probably Damaging | 0.994 | Probably Damaging | -1.30 | Pathogenic | 0.26 | Tolerated | 3.37 | 35 | 0.1132 | 0.4212 | 0 | 1 | -2.5 | 30.03 | ||||||||||||||||||||||||
| c.1754C>A | A585E 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant A585E is not reported in ClinVar (status: None) and is absent from gnomAD. Prediction tools cluster into two groups: the single benign call comes from SIFT, while all other evaluated algorithms—including REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus—label the change as pathogenic or likely pathogenic. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) indicates likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, also reports pathogenic. Thus, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.055884 | Uncertain | 0.880 | 0.244 | 0.000 | -14.715 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 5.38 | Destabilizing | 0.7 | 3.70 | Destabilizing | 4.54 | Destabilizing | 1.42 | Destabilizing | 0.539 | Likely Pathogenic | -2.59 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.30 | Pathogenic | 0.28 | Tolerated | 0.1160 | 0.1212 | 0 | -1 | -5.3 | 58.04 | |||||||||||||||||||||||||||||
| c.1754C>T | A585V 2D AIThe SynGAP1 missense variant A585V is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and SIFT, whereas a majority of tools predict a pathogenic impact: SGM‑Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. Tools with uncertain or inconclusive results (FoldX, Rosetta, Foldetta, premPS) are treated as unavailable. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic, while Foldetta remains uncertain. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.055884 | Uncertain | 0.880 | 0.244 | 0.000 | -10.843 | Likely Pathogenic | 0.970 | Likely Pathogenic | Likely Pathogenic | 0.95 | Ambiguous | 1.4 | 1.12 | Ambiguous | 1.04 | Ambiguous | 0.51 | Ambiguous | 0.420 | Likely Benign | -3.35 | Deleterious | 0.999 | Probably Damaging | 0.988 | Probably Damaging | -1.27 | Pathogenic | 0.10 | Tolerated | 0.1004 | 0.3496 | 0 | 0 | 2.4 | 28.05 | |||||||||||||||||||||||||||||
| c.1757A>C | D586A 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant D586A is not reported in ClinVar and is absent from gnomAD. Functional prediction tools that agree on a benign effect include FoldX, SIFT, and premPS, whereas a majority of predictors (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) indicate a pathogenic impact. High‑accuracy assessments further support a deleterious outcome: AlphaMissense‑Optimized classifies the variant as pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) labels it Likely Pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, remains inconclusive. No evidence from the uncertain tools (Rosetta, Foldetta) alters this consensus. Consequently, the variant is most likely pathogenic, and this prediction does not contradict any ClinVar annotation, as none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.060549 | Structured | 0.066018 | Uncertain | 0.866 | 0.241 | 0.000 | -9.955 | Likely Pathogenic | 0.960 | Likely Pathogenic | Likely Pathogenic | 0.41 | Likely Benign | 0.2 | 0.88 | Ambiguous | 0.65 | Ambiguous | 0.25 | Likely Benign | 0.693 | Likely Pathogenic | -4.58 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | -1.22 | Pathogenic | 0.31 | Tolerated | 0.3397 | 0.4673 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||
| c.1760G>T | R587M 2D  3DClick to see structure in 3D Viewer AISynGAP1 missense variant R587M is not reported in ClinVar and is absent from gnomAD. Prediction tools cluster into two groups: benign calls from FoldX, Rosetta, and Foldetta; pathogenic calls from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, and AlphaMissense‑Default; and two uncertain calls from premPS and AlphaMissense‑Optimized. High‑accuracy assessments further clarify the picture: AlphaMissense‑Optimized remains uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, predicts pathogenicity; and Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, predicts a benign effect. Overall, the majority of evidence points to a pathogenic impact for R587M, and this conclusion is not contradicted by any ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.054297 | Structured | 0.077330 | Uncertain | 0.862 | 0.216 | 0.000 | -15.106 | Likely Pathogenic | 0.931 | Likely Pathogenic | Ambiguous | 0.23 | Likely Benign | 0.0 | -0.10 | Likely Benign | 0.07 | Likely Benign | 0.84 | Ambiguous | 0.787 | Likely Pathogenic | -5.24 | Deleterious | 1.000 | Probably Damaging | 0.979 | Probably Damaging | -1.30 | Pathogenic | 0.02 | Affected | 0.1734 | 0.3910 | 0 | -1 | 6.4 | -24.99 | |||||||||||||||||||||||||||||
| c.1762C>T | L588F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L588F is reported in gnomAD (variant ID 6‑33440814‑C‑T) but has no ClinVar entry. Across the available in‑silico predictors, every tool examined classifies the substitution as pathogenic: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all return pathogenic or likely pathogenic scores. No tool predicts a benign effect. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is pathogenic. Because every available prediction converges on a deleterious effect and there is no ClinVar annotation to contradict this, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.038042 | Structured | 0.082229 | Uncertain | 0.887 | 0.214 | 0.000 | 6-33440814-C-T | -14.050 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 2.90 | Destabilizing | 0.2 | 2.55 | Destabilizing | 2.73 | Destabilizing | 1.12 | Destabilizing | 0.736 | Likely Pathogenic | -3.98 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | -1.38 | Pathogenic | 0.04 | Affected | 3.38 | 34 | 0.0816 | 0.2007 | 0 | 2 | -1.0 | 34.02 | ||||||||||||||||||||||||||
| c.1766T>C | I589T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I589T is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that assess pathogenicity all agree that the variant is deleterious: SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a pathogenic effect. No tool predicts a benign outcome. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenicity. All available predictions are consistent and conclusive. Based on the unanimous computational evidence, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar annotation (none is present). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.018415 | Structured | 0.084536 | Uncertain | 0.927 | 0.214 | 0.000 | -12.128 | Likely Pathogenic | 0.995 | Likely Pathogenic | Likely Pathogenic | 2.60 | Destabilizing | 0.0 | 2.54 | Destabilizing | 2.57 | Destabilizing | 2.07 | Destabilizing | 0.935 | Likely Pathogenic | -4.98 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.97 | Pathogenic | 0.02 | Affected | 0.1153 | 0.1231 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1768A>C | S590R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S590R is not reported in ClinVar (status: None) and has no entries in gnomAD. Prediction tools that agree on a benign effect are limited to FATHMM, whereas the majority of tools predict a pathogenic impact: SGM‑Consensus (likely pathogenic), REVEL, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No predictions are inconclusive or missing. Based on the collective evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.022667 | Structured | 0.088943 | Uncertain | 0.918 | 0.199 | 0.000 | -18.228 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 1.69 | Ambiguous | 0.6 | 3.21 | Destabilizing | 2.45 | Destabilizing | 1.28 | Destabilizing | 0.640 | Likely Pathogenic | -4.88 | Deleterious | 1.000 | Probably Damaging | 0.979 | Probably Damaging | 3.11 | Benign | 0.01 | Affected | 0.0983 | 0.3388 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1768A>T | S590C 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant S590C has no ClinVar entry and is not reported in gnomAD. Functional prediction tools show a split assessment: benign predictions come from FoldX, Rosetta, Foldetta, SIFT, FATHMM, and AlphaMissense‑Optimized, whereas pathogenic predictions arise from REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, AlphaMissense‑Default, and the SGM‑Consensus. The high‑accuracy subset indicates that AlphaMissense‑Optimized predicts a benign effect, while the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) and Foldetta both support a pathogenic or neutral outcome, respectively. Overall, the majority of tools lean toward a pathogenic interpretation, and this aligns with the SGM‑Consensus designation. Because there is no ClinVar classification, the predictions do not contradict existing clinical data. Thus, based on the available computational evidence, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.022667 | Structured | 0.088943 | Uncertain | 0.918 | 0.199 | 0.000 | -10.823 | Likely Pathogenic | 0.698 | Likely Pathogenic | Likely Benign | -0.09 | Likely Benign | 0.1 | 0.35 | Likely Benign | 0.13 | Likely Benign | 0.56 | Ambiguous | 0.631 | Likely Pathogenic | -4.48 | Deleterious | 1.000 | Probably Damaging | 0.968 | Probably Damaging | 3.08 | Benign | 0.07 | Tolerated | 0.1094 | 0.5332 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||
| c.1770C>A | S590R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S590R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools (AlphaMissense‑Default, AlphaMissense‑Optimized, SGM‑Consensus, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, Rosetta, and Foldetta) all predict a pathogenic impact; FoldX is uncertain and therefore not counted. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.022667 | Structured | 0.088943 | Uncertain | 0.918 | 0.199 | 0.000 | -18.228 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 1.69 | Ambiguous | 0.6 | 3.21 | Destabilizing | 2.45 | Destabilizing | 1.28 | Destabilizing | 0.391 | Likely Benign | -4.88 | Deleterious | 1.000 | Probably Damaging | 0.979 | Probably Damaging | 3.11 | Benign | 0.01 | Affected | 0.0983 | 0.3388 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1770C>G | S590R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S590R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining tools (AlphaMissense‑Default, AlphaMissense‑Optimized, SGM‑Consensus, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, Rosetta, and Foldetta) all predict a pathogenic impact; FoldX is uncertain and therefore not counted. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Based on the preponderance of pathogenic predictions and the lack of benign consensus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.022667 | Structured | 0.088943 | Uncertain | 0.918 | 0.199 | 0.000 | -18.228 | Likely Pathogenic | 0.999 | Likely Pathogenic | Likely Pathogenic | 1.69 | Ambiguous | 0.6 | 3.21 | Destabilizing | 2.45 | Destabilizing | 1.28 | Destabilizing | 0.389 | Likely Benign | -4.88 | Deleterious | 1.000 | Probably Damaging | 0.979 | Probably Damaging | 3.11 | Benign | 0.01 | Affected | 0.0983 | 0.3388 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1772C>A | A591D 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A591D is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL and FATHMM, whereas the remaining evaluated algorithms (AlphaMissense‑Default, ESM1b, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, premPS, FoldX‑MD, Rosetta, Foldetta, and the SGM Consensus) uniformly predict a pathogenic or likely pathogenic outcome; FoldX‑MD is uncertain but does not counter the overall trend. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. Based on the preponderance of pathogenic predictions and the absence of benign evidence, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar status (none reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.018787 | Structured | 0.093848 | Uncertain | 0.882 | 0.185 | 0.000 | -14.747 | Likely Pathogenic | 0.990 | Likely Pathogenic | Likely Pathogenic | 1.55 | Ambiguous | 0.2 | 2.77 | Destabilizing | 2.16 | Destabilizing | 1.40 | Destabilizing | 0.414 | Likely Benign | -5.02 | Deleterious | 0.919 | Possibly Damaging | 0.495 | Possibly Damaging | 3.42 | Benign | 0.00 | Affected | 0.1550 | 0.1741 | 0 | -2 | -5.3 | 44.01 | |||||||||||||||||||||||||||||
| c.1772C>T | A591V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 A591V missense variant is not reported in ClinVar and is present in gnomAD (ID 6‑33440824‑C‑T). Functional prediction tools show discordant results: benign calls come from REVEL, polyPhen‑2 HumVar, and FATHMM, while pathogenic calls are made by PROVEAN, polyPhen‑2 HumDiv, SIFT, ESM1b, and AlphaMissense‑Default. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels the variant as Likely Pathogenic. High‑accuracy assessments are mixed: AlphaMissense‑Optimized is uncertain, Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain, and the SGM Consensus remains Likely Pathogenic. Overall, the majority of evidence points toward a pathogenic effect, and this assessment is not contradicted by ClinVar, which contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.018787 | Structured | 0.093848 | Uncertain | 0.882 | 0.185 | 0.000 | 6-33440824-C-T | 2 | 1.24e-6 | -12.282 | Likely Pathogenic | 0.926 | Likely Pathogenic | Ambiguous | 1.35 | Ambiguous | 0.4 | 0.98 | Ambiguous | 1.17 | Ambiguous | 0.86 | Ambiguous | 0.321 | Likely Benign | -3.79 | Deleterious | 0.970 | Probably Damaging | 0.373 | Benign | 3.35 | Benign | 0.02 | Affected | 3.37 | 35 | 0.1128 | 0.4228 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||
| c.1786C>A | R596S 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant R596S is not reported in ClinVar and is absent from gnomAD. In silico assessment shows a consensus of pathogenicity: all evaluated tools (REVEL, FoldX, premPS, PROVEAN, polyPhen‑2 HumDiv/HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized) predict a deleterious effect, while Rosetta remains uncertain. Grouping by agreement, no tool predicts benign; the pathogenic group includes 13 predictions, with Rosetta excluded as inconclusive. High‑accuracy methods further support a damaging outcome: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. Consequently, the variant is most likely pathogenic based on the available predictions, and this assessment does not contradict ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.017797 | Structured | 0.135423 | Uncertain | 0.918 | 0.134 | 0.000 | -13.921 | Likely Pathogenic | 0.998 | Likely Pathogenic | Likely Pathogenic | 3.51 | Destabilizing | 0.2 | 1.53 | Ambiguous | 2.52 | Destabilizing | 1.17 | Destabilizing | 0.626 | Likely Pathogenic | -5.97 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.42 | Pathogenic | 0.00 | Affected | 0.3129 | 0.2680 | 0 | -1 | 3.7 | -69.11 | |||||||||||||||||||||||||||||
| c.1787G>C | R596P 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant R596P is not reported in ClinVar and is absent from gnomAD. Prediction tools that assess pathogenicity all converge on a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict pathogenicity, while no tool predicts a benign outcome. High‑accuracy methods reinforce this consensus: AlphaMissense‑Optimized indicates pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) reports “Likely Pathogenic,” and Foldetta (combining FoldX‑MD and Rosetta outputs) also predicts pathogenic. No contradictory evidence is present. Based on the uniform predictions, the variant is most likely pathogenic, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.017797 | Structured | 0.135423 | Uncertain | 0.918 | 0.134 | 0.000 | -13.786 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 6.80 | Destabilizing | 0.1 | 4.78 | Destabilizing | 5.79 | Destabilizing | 1.15 | Destabilizing | 0.780 | Likely Pathogenic | -6.97 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 2.42 | Pathogenic | 0.00 | Affected | 0.2335 | 0.3356 | 0 | -2 | 2.9 | -59.07 | |||||||||||||||||||||||||||||
| c.1795T>A | C599S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C599S is not reported in ClinVar (ClinVar ID None) and is absent from gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include only SIFT, which scores the variant as benign. All other evaluated algorithms—REVEL, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and the SGM Consensus—predict a pathogenic or likely pathogenic outcome. High‑accuracy assessments show the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) as Likely Pathogenic, AlphaMissense‑Optimized as Uncertain, and Foldetta (combining FoldX‑MD and Rosetta) as Uncertain. No prediction or folding stability result is missing; all available data are reported. Based on the preponderance of pathogenic predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.009865 | Structured | 0.151725 | Uncertain | 0.960 | 0.151 | 0.000 | -13.336 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.85 | Ambiguous | 0.0 | 1.57 | Ambiguous | 1.21 | Ambiguous | 1.27 | Destabilizing | 0.919 | Likely Pathogenic | -9.95 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.45 | Pathogenic | 0.06 | Tolerated | 0.3414 | 0.1415 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1796G>A | C599Y 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C599Y is not reported in ClinVar (ClinVar status: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that indicate a benign effect include only premPS, whereas all other evaluated algorithms—SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—consistently predict a pathogenic impact. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized reports pathogenicity; the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is labeled Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. No prediction or folding stability result is missing or inconclusive. Based on the overwhelming agreement among the majority of tools, the variant is most likely pathogenic, and this assessment does not contradict the ClinVar status, which currently contains no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.009865 | Structured | 0.151725 | Uncertain | 0.960 | 0.151 | 0.000 | -13.563 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 4.00 | Destabilizing | 1.4 | 2.80 | Destabilizing | 3.40 | Destabilizing | -0.29 | Likely Benign | 0.905 | Likely Pathogenic | -10.95 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.49 | Pathogenic | 0.00 | Affected | 0.1092 | 0.2555 | 0 | -2 | -3.8 | 60.04 | |||||||||||||||||||||||||||||
| c.1796G>C | C599S 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant C599S is not reported in ClinVar and has no entries in gnomAD. Prediction tools cluster into two groups: the sole benign prediction comes from SIFT, whereas the remaining nine tools—REVEL, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus—indicate pathogenicity. High‑accuracy assessments give a mixed picture: AlphaMissense‑Optimized is uncertain, SGM‑Consensus predicts likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is uncertain. FoldX and Rosetta individually report uncertain stability changes. Overall, the majority of evidence points to a deleterious effect, so the variant is most likely pathogenic, with no ClinVar annotation to contradict this assessment. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.009865 | Structured | 0.151725 | Uncertain | 0.960 | 0.151 | 0.000 | -13.336 | Likely Pathogenic | 0.954 | Likely Pathogenic | Ambiguous | 0.85 | Ambiguous | 0.0 | 1.57 | Ambiguous | 1.21 | Ambiguous | 1.27 | Destabilizing | 0.866 | Likely Pathogenic | -9.95 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.45 | Pathogenic | 0.06 | Tolerated | 0.3414 | 0.1415 | 0 | -1 | -3.3 | -16.06 | |||||||||||||||||||||||||||||
| c.1798C>A | P600T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P600T is not reported in ClinVar and is absent from gnomAD. Prediction tools that classify the variant as benign include Foldetta, Rosetta, and premPS. Those that predict pathogenicity are REVEL, FoldX, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN). High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM Consensus as likely pathogenic, while Foldetta indicates a benign effect on protein folding stability. Overall, the majority of computational evidence supports a pathogenic effect for P600T, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.009728 | Structured | 0.162960 | Uncertain | 0.947 | 0.147 | 0.000 | -11.945 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 2.61 | Destabilizing | 0.1 | -2.90 | Stabilizing | -0.15 | Likely Benign | 0.24 | Likely Benign | 0.737 | Likely Pathogenic | -7.97 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 1.34 | Pathogenic | 0.01 | Affected | 0.1871 | 0.4954 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||
| c.1799C>A | P600Q 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P600Q is not reported in ClinVar and has no entry in gnomAD. Functional prediction tools largely agree on a deleterious effect: benign predictions are limited to Rosetta, whereas the remaining tools—REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—all classify the variant as pathogenic. Uncertain results come from FoldX, Foldetta, and premPS. High‑accuracy assessments show AlphaMissense‑Optimized as pathogenic, the SGM‑Consensus as likely pathogenic, and Foldetta as inconclusive. Overall, the preponderance of evidence indicates that P600Q is most likely pathogenic, and this conclusion is not contradicted by any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.009728 | Structured | 0.162960 | Uncertain | 0.947 | 0.147 | 0.000 | -14.187 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 1.76 | Ambiguous | 0.0 | -3.09 | Stabilizing | -0.67 | Ambiguous | 0.53 | Ambiguous | 0.739 | Likely Pathogenic | -7.97 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 1.34 | Pathogenic | 0.00 | Affected | 0.1740 | 0.3970 | 0 | -1 | -1.9 | 31.01 | |||||||||||||||||||||||||||||
| c.1799C>G | P600R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P600R is not reported in ClinVar and is absent from gnomAD. Prediction tools that indicate a benign effect include Rosetta and premPS, whereas the majority of tools—REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized—predict a pathogenic outcome; FoldX and Foldetta are inconclusive. High‑accuracy methods give a consistent pathogenic signal: AlphaMissense‑Optimized is pathogenic, the SGM Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta remains uncertain. Overall, the computational evidence strongly favors a pathogenic classification, and this does not contradict any existing ClinVar annotation. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.009728 | Structured | 0.162960 | Uncertain | 0.947 | 0.147 | 0.000 | -15.304 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 1.48 | Ambiguous | 0.1 | -2.48 | Stabilizing | -0.50 | Ambiguous | 0.37 | Likely Benign | 0.756 | Likely Pathogenic | -8.96 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 1.34 | Pathogenic | 0.00 | Affected | 0.1604 | 0.2734 | 0 | -2 | -2.9 | 59.07 | |||||||||||||||||||||||||||||
| c.179A>C | D60A 2D AIThe SynGAP1 D60A missense variant has no ClinVar record and is not reported in gnomAD. Prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, and FATHMM, while those that predict a pathogenic outcome are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, and AlphaMissense‑Default. The SGM‑Consensus, which aggregates AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports a “Likely Benign” classification. High‑accuracy assessments show AlphaMissense‑Optimized as “Uncertain” and the SGM‑Consensus as “Likely Benign”; Foldetta results are unavailable. Overall, the majority of evaluated tools lean toward a benign interpretation, with no evidence of pathogenicity from the high‑confidence methods. Consequently, the variant is most likely benign, and this assessment does not contradict any ClinVar status, as none exists for this allele. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.284882 | Structured | 0.480942 | Uncertain | 0.521 | 0.676 | 0.000 | -4.500 | Likely Benign | 0.918 | Likely Pathogenic | Ambiguous | 0.167 | Likely Benign | -2.17 | Neutral | 0.909 | Possibly Damaging | 0.857 | Possibly Damaging | 3.96 | Benign | 0.00 | Affected | 0.3851 | 0.7405 | 0 | -2 | 5.3 | -44.01 | |||||||||||||||||||||||||||||||||||||||
| c.17C>T | A6V 2D AIThe SynGAP1 A6V missense variant is reported in gnomAD (ID 6‑33420281‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a benign effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as benign. Only SIFT predicts it to be pathogenic, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) labels it “Likely Benign.” High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized is benign, and the SGM‑Consensus is also benign; Foldetta results are not available. Overall, the preponderance of evidence indicates the variant is most likely benign, and this conclusion does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.566480 | Disordered | 0.549054 | Binding | 0.377 | 0.920 | 0.875 | 6-33420281-C-T | -3.781 | Likely Benign | 0.191 | Likely Benign | Likely Benign | 0.123 | Likely Benign | 0.32 | Neutral | 0.117 | Benign | 0.007 | Benign | 4.09 | Benign | 0.00 | Affected | 4.32 | 1 | 0.0987 | 0.5799 | 0 | 0 | 2.4 | 28.05 | ||||||||||||||||||||||||||||||||||||
| c.1802C>A | A601E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A601E is listed in ClinVar with an “Uncertain” status and is not reported in gnomAD. Prediction tools that assess pathogenicity largely agree: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized all predict a pathogenic effect, while only FATHMM predicts a benign outcome. High‑accuracy assessments reinforce this consensus: AlphaMissense‑Optimized indicates pathogenicity; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Pathogenic”; and Foldetta, which integrates FoldX‑MD and Rosetta stability calculations, also predicts pathogenicity. No predictions or stability results are missing or inconclusive. Based on the overwhelming agreement among these tools, the variant is most likely pathogenic, which does not contradict its current ClinVar “Uncertain” classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.008895 | Structured | 0.174517 | Uncertain | 0.955 | 0.156 | 0.000 | Conflicting | 2 | -16.752 | Likely Pathogenic | 0.992 | Likely Pathogenic | Likely Pathogenic | 6.68 | Destabilizing | 0.8 | 5.76 | Destabilizing | 6.22 | Destabilizing | 1.24 | Destabilizing | 0.588 | Likely Pathogenic | -4.98 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 2.54 | Benign | 0.00 | Affected | 3.37 | 35 | 0.1346 | 0.1444 | 0 | -1 | -5.3 | 58.04 | 240.0 | -82.3 | 0.0 | 0.0 | 0.7 | 0.1 | X | X | X | Potentially Pathogenic | The methyl side chain of Ala601, located on an α helix (res. Glu582-Met603), packs hydrophobically against other hydrophobic residues in the inter-helix space (e.g., Phe597, Leu598, Leu506, Phe608).In the variant simulations, the carboxylate group of Glu601 faces the inter-helix space and is forced to shift slightly away from the hydrophobic niche. Additionally, in two of the simulations, Glu601 forms a salt bridge with Arg499, causing the otherwise stable salt bridge between Arg499 and Glu496 at the outer surface of an α helix (res. Leu489-Glu519) to break due to the residue swap.These effects suggest that the protein folding process could be seriously affected. Moreover, due to its location at the GAP-Ras interface, it could also impact the complex formation with the GTPase. | ||||||||||||||
| c.1802C>G | A601G 2D  3DClick to see structure in 3D Viewer AIThe SynGAP1 A601G missense variant is listed in gnomAD (ID 6‑33440854‑C‑G) but has no ClinVar entry. Prediction tools that agree on a benign effect are FATHMM and AlphaMissense‑Optimized; those that agree on a pathogenic effect are REVEL, FoldX, Rosetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, and ESM1b. The high‑accuracy consensus methods give a pathogenic verdict: the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is also pathogenic. AlphaMissense‑Default and premPS are uncertain, and no evidence is available from other folding‑stability tools. Overall, the preponderance of evidence points to a pathogenic impact for A601G, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.008895 | Structured | 0.174517 | Uncertain | 0.955 | 0.156 | 0.000 | 6-33440854-C-G | 1 | 6.19e-7 | -11.772 | Likely Pathogenic | 0.543 | Ambiguous | Likely Benign | 2.03 | Destabilizing | 0.0 | 2.31 | Destabilizing | 2.17 | Destabilizing | 0.94 | Ambiguous | 0.511 | Likely Pathogenic | -3.98 | Deleterious | 1.000 | Probably Damaging | 0.997 | Probably Damaging | 2.55 | Benign | 0.01 | Affected | 3.37 | 35 | 0.2337 | 0.4370 | 0 | 1 | -2.2 | -14.03 | |||||||||||||||||||||||||
| c.1802C>T | A601V 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant A601V is listed in ClinVar (ID 968190.0) with an uncertain clinical significance and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from Rosetta and FATHMM, while pathogenic predictions are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, AlphaMissense‑Default, and the SGM‑Consensus score. Four tools (FoldX, Foldetta, premPS, AlphaMissense‑Optimized) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic effect, which is consistent with the ClinVar designation of uncertain significance rather than contradicting it. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.008895 | Structured | 0.174517 | Uncertain | 0.955 | 0.156 | 0.000 | Uncertain | 1 | -10.447 | Likely Pathogenic | 0.853 | Likely Pathogenic | Ambiguous | 1.64 | Ambiguous | 0.1 | 0.35 | Likely Benign | 1.00 | Ambiguous | 0.81 | Ambiguous | 0.535 | Likely Pathogenic | -3.98 | Deleterious | 1.000 | Probably Damaging | 0.989 | Probably Damaging | 2.74 | Benign | 0.03 | Affected | 3.37 | 35 | 0.1338 | 0.5263 | 0 | 0 | 2.4 | 28.05 | 228.5 | -45.5 | 0.0 | 0.0 | 0.4 | 0.5 | X | Potentially Benign | The methyl side chain of Ala601, located on an α helix (res. Glu582-Met603), packs hydrophobically against other hydrophobic residues in the inter-helix space (e.g., Phe597, Leu598, Leu506, Phe608).In the variant simulations, Val601, which has similar size and physicochemical properties to alanine, resides in the inter-helix hydrophobic space in a similar manner to Ala601 in the WT, causing no apparent negative effect on the protein structure. However, the effect of the residue swap on the SynGAP-Ras complex formation or GTPase activation cannot be fully addressed using the SynGAP solvent-only simulations. | ||||||||||||||||
| c.1805T>C | I602T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant I602T is not reported in ClinVar and is absent from gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all classify it as pathogenic or likely pathogenic. No tool reports a benign outcome. High‑accuracy assessments corroborate this trend: AlphaMissense‑Optimized is uncertain, SGM‑Consensus is likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenicity. Consequently, the variant is most likely pathogenic according to the available computational evidence, and this assessment does not contradict any ClinVar annotation (none exists). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.010221 | Structured | 0.186541 | Uncertain | 0.963 | 0.171 | 0.000 | -12.238 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 2.39 | Destabilizing | 0.1 | 2.14 | Destabilizing | 2.27 | Destabilizing | 1.94 | Destabilizing | 0.931 | Likely Pathogenic | -4.82 | Deleterious | 1.000 | Probably Damaging | 0.996 | Probably Damaging | -2.00 | Pathogenic | 0.00 | Affected | 0.0890 | 0.0989 | 0 | -1 | -5.2 | -12.05 | |||||||||||||||||||||||||||||
| c.1808T>A | M603K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M603K is not reported in ClinVar and has no entries in gnomAD. Prediction tools largely agree on a deleterious effect: pathogenic calls come from REVEL, SGM Consensus, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized, while only FoldX reports a benign outcome. High‑accuracy assessments reinforce this trend: AlphaMissense‑Optimized predicts pathogenic, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) indicates likely pathogenic, and Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is inconclusive. No other tools provide a clear benign signal. Consequently, the variant is most likely pathogenic, and this assessment does not conflict with the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.011342 | Structured | 0.197847 | Uncertain | 0.942 | 0.176 | 0.000 | -15.561 | Likely Pathogenic | 0.994 | Likely Pathogenic | Likely Pathogenic | 0.17 | Likely Benign | 0.0 | 1.19 | Ambiguous | 0.68 | Ambiguous | 0.67 | Ambiguous | 0.933 | Likely Pathogenic | -5.64 | Deleterious | 0.923 | Possibly Damaging | 0.922 | Probably Damaging | -1.35 | Pathogenic | 0.00 | Affected | 0.1268 | 0.0488 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1808T>G | M603R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M603R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include only FoldX, which scores the variant as benign. All other evaluated predictors—REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN)—classify the variant as pathogenic. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, the SGM‑Consensus also indicates likely pathogenic, while Foldetta (combining FoldX‑MD and Rosetta outputs) yields an uncertain result. No other tools provide conflicting evidence. Based on the preponderance of pathogenic predictions and the absence of benign consensus, the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.011342 | Structured | 0.197847 | Uncertain | 0.942 | 0.176 | 0.000 | -14.968 | Likely Pathogenic | 0.993 | Likely Pathogenic | Likely Pathogenic | 0.08 | Likely Benign | 0.0 | 1.13 | Ambiguous | 0.61 | Ambiguous | 0.65 | Ambiguous | 0.935 | Likely Pathogenic | -5.64 | Deleterious | 0.963 | Probably Damaging | 0.943 | Probably Damaging | -1.35 | Pathogenic | 0.00 | Affected | 0.1516 | 0.0837 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||
| c.1813C>A | P605T 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P605T is not reported in ClinVar (ClinVar status: none) and is absent from gnomAD (gnomAD ID: none). Prediction tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts a benign outcome. High‑accuracy methods specifically show pathogenicity: AlphaMissense‑Optimized predicts pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) also predicts pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) predicts pathogenic. No predictions or stability results are missing or inconclusive. Based on the unanimous pathogenic predictions and the absence of any ClinVar or gnomAD evidence to the contrary, the variant is most likely pathogenic and does not contradict ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.023087 | Structured | 0.192737 | Uncertain | 0.929 | 0.231 | 0.000 | -11.533 | Likely Pathogenic | 0.989 | Likely Pathogenic | Likely Pathogenic | 2.87 | Destabilizing | 0.4 | 2.14 | Destabilizing | 2.51 | Destabilizing | 0.72 | Ambiguous | 0.801 | Likely Pathogenic | -7.96 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | 0.69 | Pathogenic | 0.00 | Affected | 0.1626 | 0.5228 | 0 | -1 | 0.9 | 3.99 | |||||||||||||||||||||||||||||
| c.1814C>A | P605H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P605H is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized; no tool predicts a benign outcome. High‑accuracy assessments further support pathogenicity: AlphaMissense‑Optimized is pathogenic, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) is pathogenic. All available predictions are consistent and indicate a deleterious effect. Consequently, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status, as no ClinVar entry exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.023087 | Structured | 0.192737 | Uncertain | 0.929 | 0.231 | 0.000 | -13.846 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 8.80 | Destabilizing | 2.7 | 6.69 | Destabilizing | 7.75 | Destabilizing | 0.87 | Ambiguous | 0.828 | Likely Pathogenic | -8.95 | Deleterious | 1.000 | Probably Damaging | 0.999 | Probably Damaging | 0.68 | Pathogenic | 0.00 | Affected | 0.1778 | 0.4142 | 0 | -2 | -1.6 | 40.02 | |||||||||||||||||||||||||||||
| c.1814C>G | P605R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant P605R is listed in ClinVar with an uncertain significance and is not reported in gnomAD. Functional prediction tools uniformly indicate a deleterious effect: REVEL, FoldX, Rosetta, Foldetta, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized all classify the change as pathogenic, while premPS remains inconclusive. Grouping by consensus, the benign category is empty and the pathogenic category contains all available predictions. High‑accuracy assessments reinforce this view: AlphaMissense‑Optimized predicts pathogenic; the SGM Consensus (derived from AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) also yields pathogenic; and Foldetta, integrating FoldX‑MD and Rosetta outputs, reports a destabilizing, pathogenic effect. Consequently, the variant is most likely pathogenic, which does not contradict its current ClinVar uncertain status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.023087 | Structured | 0.192737 | Uncertain | 0.929 | 0.231 | 0.000 | Uncertain | 1 | -13.745 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 8.71 | Destabilizing | 2.5 | 6.46 | Destabilizing | 7.59 | Destabilizing | 0.92 | Ambiguous | 0.845 | Likely Pathogenic | -8.95 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 0.69 | Pathogenic | 0.00 | Affected | 3.37 | 35 | 0.1458 | 0.3131 | 0 | -2 | -2.9 | 59.07 | 281.7 | -118.1 | -0.2 | 0.0 | 0.5 | 0.1 | X | X | X | X | Potentially Pathogenic | Pro605 is located in a short turn between an α helix (res. Glu582-Met603) and a short α helical section (res. Ser606-Phe608). The pyrrolidine side chain of Pro605 packs hydrophobically with nearby hydrophobic residues (e.g., Ile514, Leu623, Leu610) in the inter-helix space. Additionally, proline lacks a free backbone amide group, which breaks the α helix and facilitates the turn in the WT structure.In the variant simulations, the guanidinium side chain of Arg605 is bulkier than proline, and its positively charged guanidinium group faces mostly hydrophobic residues (e.g., Ile514, Leu623, Leu610). As a result, it needs to rotate away from the hydrophobic niche. The residue swap could have a more profound effect on the actual folding process, for example, by preventing the bending at the α helix end.Moreover, due to its location at the GAP-Ras interface, the residue swap could affect the GAP-Ras association. | |||||||||||||
| c.1816A>C | S606R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S606R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, and FATHMM, whereas a separate group predicts pathogenicity: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Tools with uncertain outcomes are Foldetta, premPS, and Rosetta. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Pathogenic, and Foldetta remains inconclusive. Overall, the majority of high‑confidence predictions and the consensus vote indicate a pathogenic effect. Because ClinVar contains no entry for this variant, there is no contradiction between the computational evidence and clinical annotation. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.191720 | Uncertain | 0.875 | 0.247 | 0.000 | -12.900 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.15 | Likely Benign | 0.4 | 1.80 | Ambiguous | 0.98 | Ambiguous | 0.82 | Ambiguous | 0.252 | Likely Benign | -4.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 3.38 | Benign | 0.08 | Tolerated | 0.0819 | 0.2750 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1816A>T | S606C 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S606C is not reported in ClinVar (ClinVar ID None) and has no entry in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect include REVEL, FoldX, Rosetta, Foldetta, premPS, FATHMM, and AlphaMissense‑Optimized. Tools that predict a pathogenic effect are PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, and AlphaMissense‑Default. The SGM‑Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, reports the variant as Likely Pathogenic. High‑accuracy assessments show AlphaMissense‑Optimized as Benign, Foldetta (combining FoldX‑MD and Rosetta outputs) as Benign, and the SGM‑Consensus as Likely Pathogenic. No prediction or folding stability result is missing or inconclusive. Based on the overall distribution of predictions, the variant is most likely benign, although the SGM‑Consensus suggests pathogenicity; this does not contradict any ClinVar status because the variant is not yet classified in ClinVar. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.191720 | Uncertain | 0.875 | 0.247 | 0.000 | -11.122 | Likely Pathogenic | 0.774 | Likely Pathogenic | Likely Benign | -0.34 | Likely Benign | 0.0 | -0.31 | Likely Benign | -0.33 | Likely Benign | 0.49 | Likely Benign | 0.348 | Likely Benign | -4.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 3.31 | Benign | 0.00 | Affected | 0.0986 | 0.4580 | 0 | -1 | 3.3 | 16.06 | |||||||||||||||||||||||||||||
| c.1818T>A | S606R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S606R is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect include REVEL, FoldX, SIFT, and FATHMM, whereas a separate group predicts pathogenicity: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, and AlphaMissense‑Default. Tools with uncertain outcomes are Foldetta, premPS, and Rosetta. High‑accuracy assessments further support a deleterious impact: AlphaMissense‑Optimized predicts pathogenicity, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) is Likely Pathogenic, and Foldetta remains inconclusive. Overall, the majority of high‑confidence predictions and the consensus vote indicate a pathogenic effect. Because ClinVar contains no entry for this variant, there is no contradiction between the computational evidence and clinical annotation. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.191720 | Uncertain | 0.875 | 0.247 | 0.000 | -12.900 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.15 | Likely Benign | 0.4 | 1.80 | Ambiguous | 0.98 | Ambiguous | 0.82 | Ambiguous | 0.246 | Likely Benign | -4.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 3.38 | Benign | 0.08 | Tolerated | 0.0819 | 0.2750 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1818T>G | S606R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant S606R is not reported in ClinVar or gnomAD. Prediction tools that indicate a benign effect include REVEL, FoldX, SIFT, and FATHMM, whereas a majority predict pathogenicity: PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is labeled Likely Pathogenic, while Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. High‑accuracy methods confirm the pathogenic trend: AlphaMissense‑Optimized is pathogenic, SGM Consensus is Likely Pathogenic, and Foldetta remains uncertain. Overall, the evidence points to the variant being most likely pathogenic, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.041405 | Structured | 0.191720 | Uncertain | 0.875 | 0.247 | 0.000 | -12.900 | Likely Pathogenic | 0.997 | Likely Pathogenic | Likely Pathogenic | 0.15 | Likely Benign | 0.4 | 1.80 | Ambiguous | 0.98 | Ambiguous | 0.82 | Ambiguous | 0.246 | Likely Benign | -4.98 | Deleterious | 0.999 | Probably Damaging | 0.997 | Probably Damaging | 3.38 | Benign | 0.08 | Tolerated | 0.0819 | 0.2750 | 0 | -1 | -3.7 | 69.11 | |||||||||||||||||||||||||||||
| c.1819C>T | L607F 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant L607F is catalogued in gnomAD (6‑33440871‑C‑T) but has no ClinVar entry. Functional prediction tools largely agree on a deleterious effect: REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) all report pathogenic or likely pathogenic. Only FoldX predicts a benign outcome, while Rosetta, Foldetta, premPS, and AlphaMissense‑Optimized are uncertain. High‑accuracy assessments show the SGM‑Consensus as likely pathogenic, AlphaMissense‑Optimized as uncertain, and Foldetta as uncertain. Overall, the preponderance of evidence points to a pathogenic effect for L607F, and this conclusion is not contradicted by ClinVar status (none). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.048328 | Structured | 0.194229 | Uncertain | 0.869 | 0.250 | 0.000 | 6-33440871-C-T | 1 | 6.19e-7 | -13.654 | Likely Pathogenic | 0.948 | Likely Pathogenic | Ambiguous | 0.23 | Likely Benign | 0.1 | 1.20 | Ambiguous | 0.72 | Ambiguous | 0.61 | Ambiguous | 0.758 | Likely Pathogenic | -3.98 | Deleterious | 0.998 | Probably Damaging | 0.997 | Probably Damaging | -1.54 | Pathogenic | 0.01 | Affected | 3.37 | 35 | 0.0872 | 0.2816 | 0 | 2 | -1.0 | 34.02 | ||||||||||||||||||||||||
| c.181G>A | E61K 2D AIThe SynGAP1 E61K missense variant has no ClinVar entry and is not reported in gnomAD. Functional prediction tools cluster into two consensus groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus, SGM‑Consensus, classifies the variant as Likely Benign, and AlphaMissense‑Optimized also predicts Benign. Foldetta, a protein‑folding stability method that integrates FoldX‑MD and Rosetta outputs, has no available result for this variant. Based on the aggregate evidence, the variant is most likely benign, and this assessment does not contradict any ClinVar status (none is reported). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | -4.953 | Likely Benign | 0.425 | Ambiguous | Likely Benign | 0.120 | Likely Benign | -0.22 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.34 | Benign | 0.00 | Affected | 0.2736 | 0.5691 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||||||||||||
| c.1826G>A | G609E 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant G609E has no ClinVar entry and is not reported in gnomAD. Prediction tools that agree on a benign effect include PROVEAN, SIFT, and AlphaMissense‑Optimized. Those that agree on a pathogenic effect are REVEL, FoldX, polyPhen‑2 (HumDiv and HumVar), ESM1b, and FATHMM. The remaining tools (Rosetta, Foldetta, premPS, AlphaMissense‑Default) give uncertain or inconclusive results. High‑accuracy assessments show AlphaMissense‑Optimized as benign, the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as pathogenic, and Foldetta as uncertain. Overall, the majority of evidence points to a pathogenic impact, and this conclusion is not contradicted by any ClinVar annotation. Thus, the variant is most likely pathogenic. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | GAP | 0.179055 | Structured | 0.203786 | Uncertain | 0.851 | 0.252 | 0.000 | -10.470 | Likely Pathogenic | 0.421 | Ambiguous | Likely Benign | 2.95 | Destabilizing | 0.4 | -1.65 | Ambiguous | 0.65 | Ambiguous | 0.61 | Ambiguous | 0.531 | Likely Pathogenic | -2.28 | Neutral | 0.916 | Possibly Damaging | 0.588 | Possibly Damaging | -1.41 | Pathogenic | 0.17 | Tolerated | 0.1709 | 0.4491 | 0 | -2 | -3.1 | 72.06 | ||||||||||||||||||||||||||||||
| c.182A>C | E61A 2D AIThe SynGAP1 missense variant E61A is listed in ClinVar (ID 3767543.0) with an *Uncertain* clinical significance and is not reported in gnomAD. Functional prediction tools that agree on a benign effect include REVEL, PROVEAN, ESM1b, FATHMM, AlphaMissense‑Optimized, and the SGM‑Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). Tools that predict a pathogenic effect are polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and SIFT; AlphaMissense‑Default remains uncertain. High‑accuracy assessments show AlphaMissense‑Optimized as benign and the SGM‑Consensus as likely benign, while Foldetta (a protein‑folding stability method combining FoldX‑MD and Rosetta outputs) has no available result for this variant. Overall, the majority of evidence points to a benign impact, and this conclusion does not contradict the current ClinVar status of uncertainty. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | Uncertain | 1 | -5.235 | Likely Benign | 0.453 | Ambiguous | Likely Benign | 0.074 | Likely Benign | -1.52 | Neutral | 0.458 | Possibly Damaging | 0.678 | Possibly Damaging | 4.12 | Benign | 0.00 | Affected | 0.4499 | 0.5878 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||||||||||
| c.182A>G | E61G 2D AIThe SynGAP1 missense variant E61G is not reported in ClinVar and is absent from gnomAD. Functional prediction tools cluster into two groups: benign predictions come from REVEL, PROVEAN, ESM1b, FATHMM, and AlphaMissense‑Optimized, while pathogenic predictions arise from polyPhen‑2 (HumDiv and HumVar) and SIFT. AlphaMissense‑Default remains uncertain. The high‑accuracy consensus methods reinforce the benign assessment: AlphaMissense‑Optimized predicts benign, the SGM Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) reports “Likely Benign,” and the protein‑folding stability tool Foldetta is not available for this variant. Overall, the majority of evidence points to a benign effect. There is no ClinVar entry to contradict this conclusion. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Benign | 0.352862 | Structured | 0.477329 | Uncertain | 0.518 | 0.699 | 0.125 | -5.574 | Likely Benign | 0.469 | Ambiguous | Likely Benign | 0.072 | Likely Benign | -1.68 | Neutral | 0.659 | Possibly Damaging | 0.775 | Possibly Damaging | 4.08 | Benign | 0.00 | Affected | 0.3593 | 0.4965 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||||||||||||
| c.1832T>A | M611K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 M611K variant has no ClinVar entry and is not reported in gnomAD. Prediction tools that classify it as benign include polyPhen‑2 HumDiv, polyPhen‑2 HumVar, and AlphaMissense‑Optimized. The majority of other in silico predictors (SGM‑Consensus, REVEL, Rosetta, Foldetta, premPS, PROVEAN, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) indicate a pathogenic effect; FoldX is uncertain and therefore not considered evidence. High‑accuracy assessments show AlphaMissense‑Optimized as benign, while the SGM Consensus (derived from the unanimous pathogenic vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) and Foldetta (combining FoldX‑MD and Rosetta outputs) both predict pathogenicity. Based on the preponderance of pathogenic predictions and the high‑accuracy consensus, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.236433 | Structured | 0.210791 | Uncertain | 0.870 | 0.253 | 0.000 | -13.021 | Likely Pathogenic | 0.630 | Likely Pathogenic | Likely Benign | 1.86 | Ambiguous | 0.6 | 2.65 | Destabilizing | 2.26 | Destabilizing | 1.65 | Destabilizing | 0.665 | Likely Pathogenic | -4.10 | Deleterious | 0.250 | Benign | 0.120 | Benign | -1.26 | Pathogenic | 0.03 | Affected | 0.1193 | 0.0688 | 0 | -1 | -5.8 | -3.02 | |||||||||||||||||||||||||||||
| c.1832T>G | M611R 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant M611R is not reported in ClinVar (ClinVar ID None) and has no entries in gnomAD (gnomAD ID None). Prediction tools that agree on a benign effect are AlphaMissense‑Optimized, polyPhen2_HumVar, and SIFT. Tools that agree on a pathogenic effect include SGM‑Consensus, REVEL, Rosetta, premPS, PROVEAN, polyPhen2_HumDiv, ESM1b, FATHMM, and AlphaMissense‑Default. High‑accuracy assessments show AlphaMissense‑Optimized predicts a benign outcome, whereas the SGM‑Consensus (a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN) predicts pathogenic. Foldetta, which integrates FoldX‑MD and Rosetta stability outputs, is reported as uncertain. No prediction or folding result is missing; all available data are considered. Based on the overall distribution of predictions, the variant is most likely pathogenic, and this assessment does not contradict any ClinVar status because no ClinVar classification exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.236433 | Structured | 0.210791 | Uncertain | 0.870 | 0.253 | 0.000 | -11.050 | Likely Pathogenic | 0.642 | Likely Pathogenic | Likely Benign | 1.80 | Ambiguous | 0.8 | 2.00 | Destabilizing | 1.90 | Ambiguous | 1.58 | Destabilizing | 0.644 | Likely Pathogenic | -4.10 | Deleterious | 0.779 | Possibly Damaging | 0.159 | Benign | -1.21 | Pathogenic | 0.21 | Tolerated | 0.1399 | 0.0837 | 0 | -1 | -6.4 | 24.99 | |||||||||||||||||||||||||||||
| c.1835A>C | Q612P 2D 3DClick to see structure in 3D Viewer AISynGAP1 missense variant Q612P is listed in ClinVar (ID 3660462.0) with an uncertain significance annotation and is not reported in gnomAD. Functional prediction tools cluster into two groups: benign predictions come from FoldX, SIFT, and AlphaMissense‑Optimized; pathogenic predictions arise from REVEL, PolyPhen‑2 (HumDiv and HumVar), ESM1b, FATHMM, AlphaMissense‑Default, PROVEAN, and the SGM Consensus score (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN). The high‑accuracy AlphaMissense‑Optimized predicts benign, whereas the SGM Consensus predicts likely pathogenic; Foldetta, a folding‑stability method combining FoldX‑MD and Rosetta outputs, returns an uncertain result and is therefore not factored into the consensus. Overall, the majority of evidence supports a pathogenic effect, which contrasts with the ClinVar uncertain classification. Thus, based on current predictions, the variant is most likely pathogenic, contradicting the ClinVar status. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.203988 | Uncertain | 0.822 | 0.263 | 0.000 | Uncertain | 1 | -9.684 | Likely Pathogenic | 0.673 | Likely Pathogenic | Likely Benign | -0.19 | Likely Benign | 0.3 | 3.06 | Destabilizing | 1.44 | Ambiguous | 0.56 | Ambiguous | 0.671 | Likely Pathogenic | -5.84 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | -1.31 | Pathogenic | 0.19 | Tolerated | 0.2252 | 0.4050 | 0 | -1 | 1.9 | -31.01 | |||||||||||||||||||||||||||
| c.1837G>A | E613K 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E613K is not reported in ClinVar and is absent from gnomAD. Functional prediction tools show a split: benign calls come from FoldX, Foldetta, and premPS, whereas pathogenic calls are made by REVEL, PROVEAN, polyPhen‑2 (HumDiv and HumVar), SIFT, ESM1b, FATHMM, AlphaMissense‑Default, and AlphaMissense‑Optimized. The SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is also pathogenic. High‑accuracy assessments give AlphaMissense‑Optimized as pathogenic, SGM Consensus as pathogenic, and Foldetta (combining FoldX‑MD and Rosetta outputs) as benign. No prediction is inconclusive. Overall, the majority of evidence points to a pathogenic effect, and this conclusion does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.193489 | Uncertain | 0.816 | 0.254 | 0.000 | -11.892 | Likely Pathogenic | 0.987 | Likely Pathogenic | Likely Pathogenic | 0.34 | Likely Benign | 0.6 | -0.54 | Ambiguous | -0.10 | Likely Benign | -0.08 | Likely Benign | 0.567 | Likely Pathogenic | -3.72 | Deleterious | 0.996 | Probably Damaging | 0.987 | Probably Damaging | -1.15 | Pathogenic | 0.04 | Affected | 0.2979 | 0.5942 | 0 | 1 | -0.4 | -0.94 | |||||||||||||||||||||||||||||
| c.1838A>C | E613A 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 E613A missense variant is not reported in ClinVar (ClinVar ID: None) and is absent from gnomAD (gnomAD ID: None). Prediction tools that agree on a benign effect include Rosetta, premPS, and Foldetta, whereas the majority of tools predict a pathogenic impact: REVEL, SIFT, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, ESM1b, FATHMM, AlphaMissense‑Default, and the SGM‑Consensus score. High‑accuracy assessments show AlphaMissense‑Optimized as uncertain, the SGM‑Consensus (majority vote of AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) as likely pathogenic, and Foldetta (combining FoldX‑MD and Rosetta stability outputs) as benign. Overall, the preponderance of evidence (10 pathogenic vs. 3 benign predictions) indicates that the variant is most likely pathogenic, and this conclusion does not contradict the ClinVar status, which currently has no entry for this variant. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.193489 | Uncertain | 0.816 | 0.254 | 0.000 | -10.841 | Likely Pathogenic | 0.906 | Likely Pathogenic | Ambiguous | 0.90 | Ambiguous | 0.5 | -0.17 | Likely Benign | 0.37 | Likely Benign | 0.32 | Likely Benign | 0.688 | Likely Pathogenic | -5.57 | Deleterious | 0.999 | Probably Damaging | 0.998 | Probably Damaging | -1.26 | Pathogenic | 0.02 | Affected | 0.4696 | 0.5929 | 0 | -1 | 5.3 | -58.04 | |||||||||||||||||||||||||||||
| c.1838A>G | E613G 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant E613G is not reported in ClinVar and has no entry in gnomAD. Prediction tools that agree on a benign effect include only premPS, whereas the remaining tools (SGM‑Consensus, REVEL, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, FATHMM, AlphaMissense‑Default) uniformly predict a pathogenic impact. The high‑accuracy methods give the following results: AlphaMissense‑Optimized is uncertain; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, is pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, is uncertain. No prediction or stability assessment is missing or inconclusive beyond the uncertain labels. Overall, the preponderance of evidence points to a pathogenic effect for E613G, and this assessment does not contradict the absence of a ClinVar classification. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.275179 | Structured | 0.193489 | Uncertain | 0.816 | 0.254 | 0.000 | -12.417 | Likely Pathogenic | 0.911 | Likely Pathogenic | Ambiguous | 1.49 | Ambiguous | 0.3 | 1.34 | Ambiguous | 1.42 | Ambiguous | 0.08 | Likely Benign | 0.641 | Likely Pathogenic | -6.56 | Deleterious | 1.000 | Probably Damaging | 0.998 | Probably Damaging | -1.26 | Pathogenic | 0.01 | Affected | 0.3422 | 0.5266 | 0 | -2 | 3.1 | -72.06 | |||||||||||||||||||||||||||||
| c.1840T>C | Y614H 2D 3DClick to see structure in 3D Viewer AIThe SynGAP1 missense variant Y614H is not reported in ClinVar (no ClinVar ID) and is absent from gnomAD (no gnomAD ID). Prediction tools that agree on a benign effect include REVEL, SIFT, and FATHMM, whereas a majority of tools predict a pathogenic impact: AlphaMissense‑Default, AlphaMissense‑Optimized, ESM1b, PROVEAN, polyPhen‑2 (HumDiv and HumVar), and Rosetta. Tools with uncertain or inconclusive results (FoldX, Foldetta, premPS) are not considered evidence for either side. High‑accuracy assessments further support a deleterious effect: AlphaMissense‑Optimized is pathogenic; the SGM Consensus (majority vote from AlphaMissense‑Default, ESM1b, FATHMM, PROVEAN) is Likely Pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, remains uncertain. Overall, the preponderance of evidence indicates that the variant is most likely pathogenic, and this conclusion does not contradict any ClinVar annotation (none is available). Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.314870 | Structured | 0.182134 | Uncertain | 0.852 | 0.254 | 0.000 | -9.678 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 1.17 | Ambiguous | 0.5 | 2.38 | Destabilizing | 1.78 | Ambiguous | 0.91 | Ambiguous | 0.397 | Likely Benign | -4.95 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.58 | Benign | 0.27 | Tolerated | 0.1948 | 0.0373 | 0 | 2 | -1.9 | -26.03 | |||||||||||||||||||||||||||||
| c.1841A>G | Y614C 2D AIThe SynGAP1 missense variant Y614C is not reported in ClinVar and is absent from gnomAD. Prediction tools that agree on a benign effect are limited to FATHMM, whereas the remaining 13 tools (SGM‑Consensus, REVEL, FoldX, Rosetta, Foldetta, premPS, PROVEAN, polyPhen‑2 HumDiv, polyPhen‑2 HumVar, SIFT, ESM1b, AlphaMissense‑Default, AlphaMissense‑Optimized) uniformly predict a pathogenic impact. High‑accuracy methods reinforce this view: AlphaMissense‑Optimized scores the variant as pathogenic; the SGM Consensus, derived from a majority vote of AlphaMissense‑Default, ESM1b, FATHMM, and PROVEAN, labels it likely pathogenic; and Foldetta, which integrates FoldX‑MD and Rosetta outputs, also predicts pathogenicity. Consequently, the variant is most likely pathogenic based on the collective predictions, and this assessment does not contradict any ClinVar annotation because none exists. Disclaimer: This summary was generated using AI and should be interpreted alongside expert review. | Likely Pathogenic | GAP | 0.314870 | Structured | 0.182134 | Uncertain | 0.852 | 0.254 | 0.000 | -11.716 | Likely Pathogenic | 0.996 | Likely Pathogenic | Likely Pathogenic | 1.56 | Ambiguous | 0.7 | 3.21 | Destabilizing | 2.39 | Destabilizing | 1.76 | Destabilizing | 0.539 | Likely Pathogenic | -8.83 | Deleterious | 1.000 | Probably Damaging | 1.000 | Probably Damaging | 3.41 | Benign | 0.02 | Affected | 0.3081 | 0.1679 | 0 | -2 | 3.8 | -60.04 | |||||||||||||||||||||||||||||
Found 8751 rows. Show 800 rows per page. Page 4/11 « Previous | Next »